






-
浙江山核桃Carya cathayensis属胡桃科Juglandaceae山核桃属Carya,在浙江临安、淳安已形成规模化栽培体系,是当地农业经济的重要组成部分[1]。山核桃在自然生长条件下存在严重的生理落果现象。数据显示:山核桃生理落果率普遍超过60%[2]。这一现象直接导致山核桃减产减收,因此阐明其落果机制并研发调控技术已成为提升山核桃产业效益的关键。
植物器官脱落过程受控于离层(abscission zone)的形成与激活机制。当果实启动脱落程序时,其与果柄连接处的细胞通过纤维素酶、果胶酶等水解酶类,使细胞壁中1~2层细胞发生胞间分离,形成结构脆弱的离层组织[3]。QIU等[4]在樱桃Prunus avium落果研究中提出,离层形成诱因可归为3类:果实的成熟衰老、器官发育资源竞争、逆境胁迫。离层形成的调控网络具有多层级特征,既涉及生长素浓度变化与乙烯合成通路的激素级联反应,也依赖于多聚半乳糖醛酸酶、β-半乳糖苷酶等细胞壁降解酶系的时序性表达[5−6]。植物器官脱落是一种细胞衰老死亡过程,与体内活性氧(ROS)水平紧密相关,因此有必要探究活性氧水平与山核桃生理落果的关系。
活性氧作为植物氧化代谢的重要副产物,在体内过量积累时,会引发膜脂过氧化、蛋白质变性及DNA损伤等病理效应[7]。离层形成作为程序性细胞死亡的特殊表现形式,其发生过程与活性氧代谢存在相关性[8−9]。活性氧调控技术已广泛应用于苹果Malus pumila、蓝莓Vaccinium spp.等果实的保鲜处理,通过外源添加活性氧清除剂可有效抑制细胞内过氧化氢积累,阻止氧化应激和细胞死亡,抑制膜脂过氧化进程[10]。本研究选取碘化钾和没食子酸丙酯作为模式抗氧化剂,两者均具有环境兼容性好、作用机制明确、施用成本低等优势[11],适用于在大田条件下山核桃落果调控研究。
当前山核桃研究体系聚焦于栽培模式优化、病虫害综合防治、高产品种选育及功能成分开发利用等应用领域[12]。魏海林等[13]研究表明:引起薄壳山核桃C. illinoinensis落花落果的原因主要有授粉不良、营养不足、栽植过密、低温冻害和病虫害。杨蕊等[14]发现:外源褪黑素处理可通过上调抗氧化酶活性,显著抑制青皮核桃Juglans hopeiensis储藏期间的膜脂过氧化反应进程。然而,关于活性氧代谢在山核桃离层形成中的具体作用尚未明确。本研究以浙江山核桃为对象,通过连续动态观测,明确落果高峰期的时空分布特征,结合组织切片技术解析离层发育的细胞学模式与活性氧积累位点;通过正交试验设计优化碘化钾与没食子酸丙酯的施用浓度与频次,最终构建基于活性氧清除的落果防控技术体系。研究成果将对山核桃生理落果控制具有重要实践指导意义。
-
本研究以山核桃为材料,试验在山核桃核心栽培区杭州市临安区示范园及淳安县临岐镇规模化种植基地开展。分别于2023和2024年生长季(4—7月)选择株高3~5 m,树龄8~10 a,冠层完整且无病虫害侵染的健康植株作为研究对象,同时确保每年的试验气候环境相似。通过物候学观测确定自花授粉完成期(雌花柱头褐变率达90%以上)后,对进入幼果发育阶段(子房直径>3 mm)的果实进行定位标记,系统观测其落果动态规律。试验采用的活性氧清除剂为碘化钾(西陇科学,CAS
7681 -11-0)和没食子酸丙酯(Sigma-Aldrich,CAS 121-79-9)。 -
选定幼果膨大初期作为处理窗口期。采用分层随机抽样法选取着果量10~30个的二次分枝作为处理小区,每个处理重复3次,以清水为对照组。处理分别为碘化钾(3.0、5.0、10.0 mmol·L−1)和没食子酸丙酯(0.1、0.3、0.5 mmol·L−1),配制时添加体积分数为0.1%的Triton X-100作为表面活性剂以增强叶面附着性,将试剂喷施于山核桃果实与枝条连接处。处理时段为日均温25~28 ℃、相对湿度60%~75%的晴日8:00—11:00,每次处理间隔7 d,共处理3次。
-
建立基于时间序列的果实追踪数据库。处理前进行基线果实计数(S0),随后在第1次(S1)、第2次(S2)、第3次(S3)处理后第7天分别进行全株果实普查。统计参数包括阶段落果量(ΔSn = Sn−1−Sn,n=1,2,3)和累计落果率[L = (S0−S3)/S0×100%]。2023年初始统计时间为5月13日,2024年初始统计时间为5月18日。
-
采集正常果与自然脱落果的果柄离区组织(长度为5 mm),立即浸入预冷的质量分数为2.5%戊二醛(0.1 mol·L−1 PBS缓冲液,pH 7.2)进行渗透固定(2 h),再用体积分数为75%乙醇浸泡洗脱2次。
活性氧原位检测:过氧化氢定位采用3,3'-二氨基联苯胺(DAB,1 g·L−1)染色液避光浸渍16 h,利用内源性过氧化物酶催化过氧化氢与3,3'-二氨基联苯胺生成棕色聚合物。${\mathrm{O}}_{2}^{-} $检测采用质量分数为0.1%硝基蓝四氮唑 [NBT,含10.0 mmol·L−1 三氮化钠(NaN3)]染色12 h,通过超氧阴离子介导的硝基蓝四氮唑还原反应形成蓝色沉淀。染色完成后,用体积分数为75%乙醇将果实组织进行脱色处理,期间更换3~4次脱色溶液,直至脱色完成。脱色完成后的组织样品用作解剖材料。
-
使用Leica VT1200S振荡切片机,将脱色完成的组织样品用质量分数为5%琼脂糖包埋,用强力胶固定在载物台上,调制参数收集50 μm的切片,于试管内加入去离子水保存切片,用于后续显微镜观察。使用Leica DM6B全电动正置显微镜(HC PL FLUOTAR 10×/0.30,20×/0.40)进行明场成像,配合LAS X软件(v3.7)进行图像采集。
-
计算不同处理后的山核桃落果率,通过单因素方差分析不同组别的显著性差异(P<0.05),采用Excel 2016整理数据,SPSS 27.0对数据进行ANOVA方差检验和Duncan’s多重比较,并用GraphPad Prism 10.1.2软件绘制图表,用Power Point处理图片。
-
对浙江淳安、临安地区发育约30 d的山核桃果实进行显微解剖学观察发现:自然脱落进程中果柄与枝条交界处形成典型的离层结构(图1 A和B)。离层细胞是植物在器官脱落过程中形成的一种特殊细胞层,主要存在于木本双子叶植物及裸子植物的叶柄或叶基部,形成一群小型细胞。这些细胞失水皱缩,细胞壁逐渐胶化,对植物器官的支持力量变弱。切取果实与枝条相连的组织在显微镜下观察,发现离层细胞比周围细胞更小,排列更为紧密(图1 C和D),说明山核桃果实与枝条形成离层过程中产生了细胞和组织的形态变化。
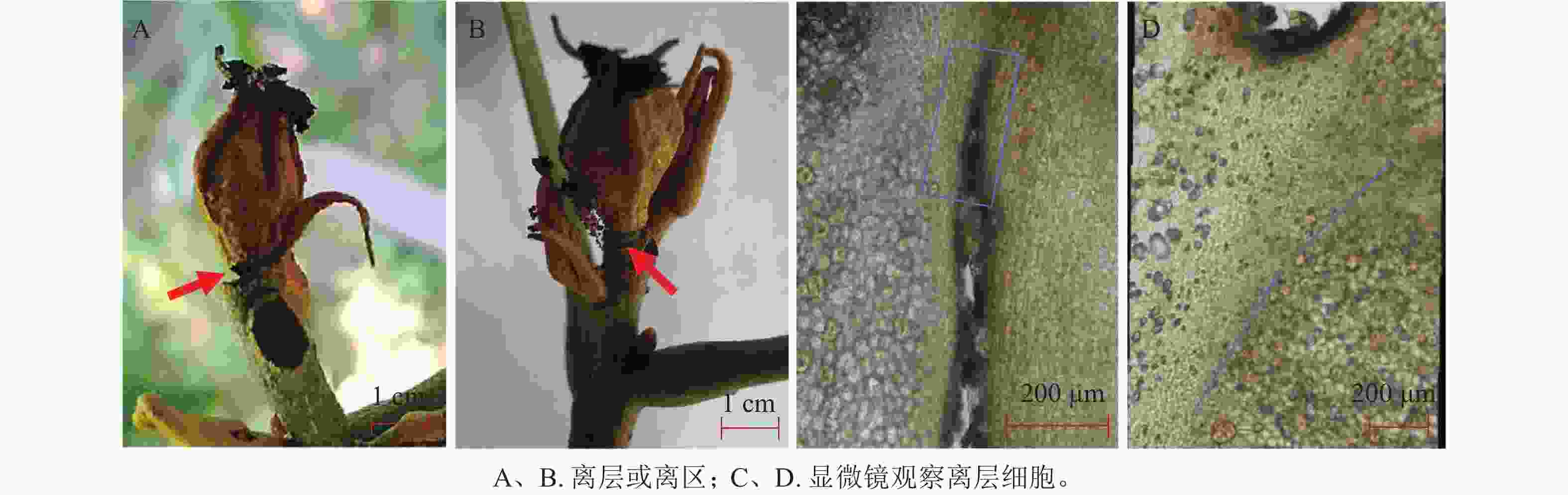
图 1 山核桃果实脱落过程中产生离层
Figure 1. Formation of the abscission zone during the C. cathayensis fruit abscission process
-
通过系统解剖学追踪,发现山核桃离层主要分布于2类微解剖位点:果柄-枝条连接处和果实-果柄过渡区(图2)。在山核桃果实的脱落过程中,果柄与枝条、果实与枝条的连结处为新生的组织,质地柔软,牢固性差,表明落果的形成可能与组织结构、细胞紧密程度相关,影响离层的形成。

图 2 山核桃果实脱落位置
Figure 2. Different abscission sites of C. cathayensis fruits
-
表1为2023—2024年的时序监测数据。由表1可知:2023和2024年对照组平均落果率高达89.55%±2.43%和80.09%±4.05%。在喷施不同浓度的活性氧清除剂后,在2023年中,3.0 mmol·L−1碘化钾、0.3 mmol·L−1 没食子酸丙酯、0.5 mmol·L−1没食子酸丙酯处理组的落果率相较于对照显著降低(P<0.05),分别为57.24%±8.60%、66.66%±3.89%、51.85%±3.15%;2024年,3.0 mmol·L−1碘化钾、0.1 mmol·L−1没食子酸丙酯、0.3 mmol·L−1没食子酸丙酯、0.5 mmol·L−1没食子酸丙酯处理组的落果率相较于对照显著降低(P<0.05),分别为 61.22%±2.75%、54.47%±1.34%、58.62%±2.68%、45.77%±2.17%。跨年度数据表明:3.0 mmol·L−1碘化钾与0.5 mmol·L−1没食子酸丙酯具有稳定的落果抑制效果,表明这2种试剂更适宜用作山核桃落果的活性氧清除剂,可进一步用于探究山核桃果实体内活性氧水平的变化差异。
表 1 山核桃在不同试剂处理后的落果率
Table 1. Fruit drop rate of C. cathayensis after treatment with different reagents
试剂 处理浓度/
(mmol·L−1)落果率/% 2023年 2024年 对照 89.55±2.43 a 80.09±4.05 ab 碘化钾 3.0 57.24±8.60 c 61.22±2.75 c 5.0 83.62±7.38 a 76.42±0.80 b 10.0 76.72±4.51 ab 88.75±5.73 a 没食子
酸丙酯0.1 78.03±2.46 ab 54.47±1.34 cd 0.3 66.66±3.89 bc 58.62±2.68 c 0.5 51.85±3.15 c 45.77±2.17 d 说明:同列不同字母表示不同处理与对照间差异显著 (P<0.05)。 -
山核桃果实在脱落过程中会产生离层。对正常生长未脱落和已脱落的山核桃果实切片分别进行3,3'-二氨基联苯胺和硝基蓝四氮唑组织化学定位,显微镜观察其着色差异(图3A~B)。结果显示:正常生长的果实内活性氧染色无显著差异,已脱落的果实在离层位置显色加深,说明果实脱落后离层位置的活性氧水平更高。
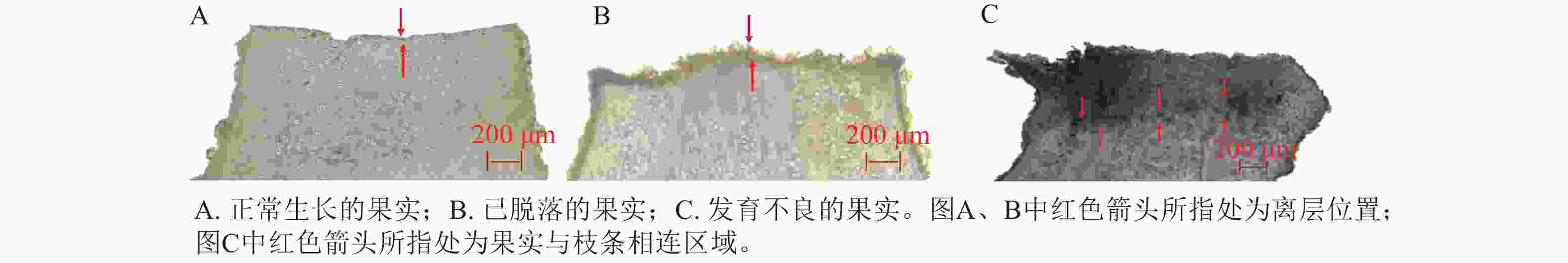
图 3 山核桃果实3,3'-二氨基联苯胺染色(A、B)和硝基蓝四氮唑染色(C)
Figure 3. DAB staining (A, B) and NBT staining (C) of C. cathayensis fruits
为了研究山核桃果实未完全脱落时体内活性氧的情况,采集瘦弱暗黄生长差的果实切片进行硝基蓝四氮唑试剂染色,观察发现:果实与枝条相连区域有明显的着色差异(图3C),表明活性氧水平的增加可能是形成离层的化学信号之一。果实体内活性氧水平增加,细胞形态发生变化乃至死亡,果实与枝条之间形成离层直至完全脱落。
-
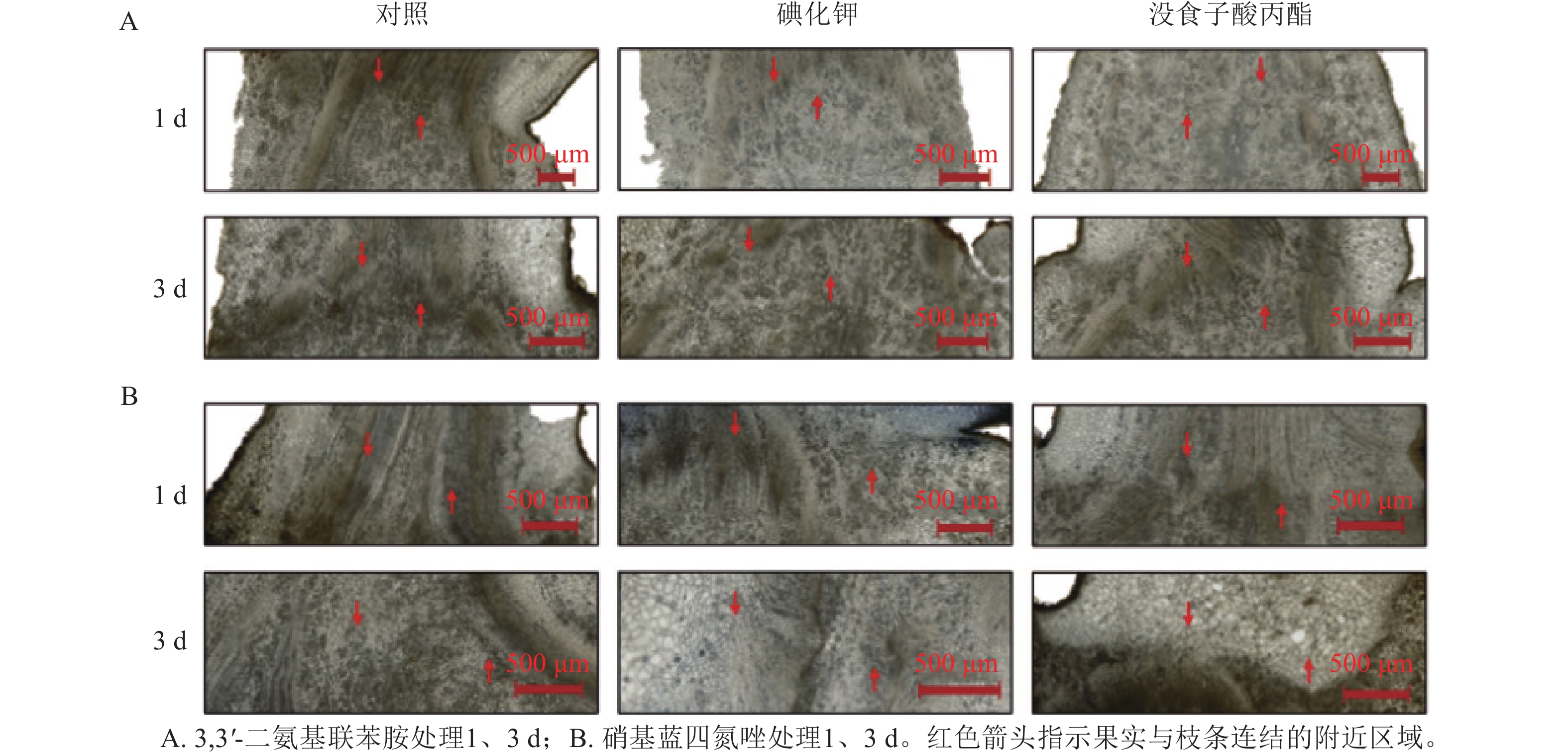
用3.0 mmol·L−1碘化钾和0.5 mmol·L−1没食子酸丙酯对正常生长的山核桃果实分别处理1、3 d,采集后同一时间进行组织固定和3,3'-二氨基联苯胺、硝基蓝四氮唑染色,观察切片显色差异(图4)。对比果实与枝条连结的组织切片染色情况。结果表明:正常生长的山核桃果实经过碘化钾、没食子酸丙酯的处理后,比对照处理显色更浅,体内活性氧水平更低,通过3,3'-二氨基联苯胺、硝基蓝四氮唑染色,碘化钾、没食子酸丙酯对活性氧的清除效果得到进一步验证。
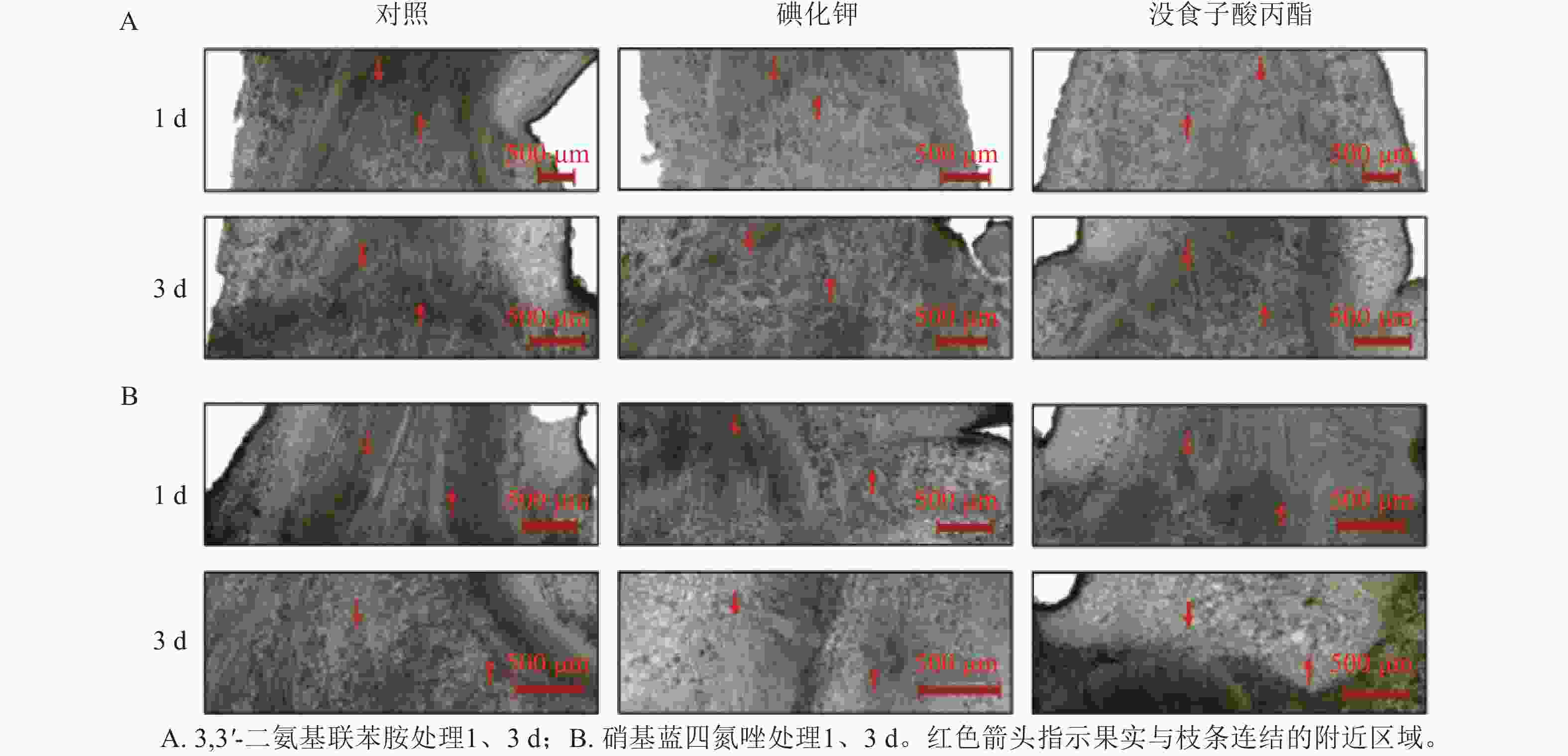
图 4 碘化钾和没食子酸丙酯处理后山核桃果实3,3'-二氨基联苯胺和硝基蓝四氮唑染色结果
Figure 4. DAB and NBT staining of potassium iodide and propyl gallate treated C. cathayensis fruits
-
山核桃的生理落果现象呈现复杂的时空动态特征,其发生机制受气候条件、营养分配及解剖结构等多因素协同调控。①受精不良。受精是果实发育的关键环节。如果花粉质量差、授粉时机不当或花粉与雌蕊的亲和性差,都会导致受精失败,从而引起落果。例如,薄壳山核桃柱头张角大于90°时为雌花最佳授粉期[2],坐果率最高。②营养竞争。在果实生长发育过程中,尤其是果实膨大期,需要大量的营养物质。如果营养供应不足或分配不均,会导致一些果实因缺乏营养而脱落。研究表明:6月10日至7月10日是山核桃果实快速增长期,也是落果高峰期,这一时期果实生长需要大量营养,竞争激烈[15]。③环境因素。气候条件对山核桃的落果也有显著影响。干旱、低温、高温等极端天气条件会影响花粉活力和花的发育,进而导致落果。此外,连续的阴雨天气会使果实含水率增高,影响果实的正常发育[16]。
本研究发现:山核桃果实脱落主要发生在2个解剖学界面,分别为果实-枝条离层区和果柄-枝条离层区。组织学观察显示:这2处离层组织的细胞排列方式存在明显差异,提示不同脱落位点的生物力学特性可能与其细胞学特征密切相关。曹艺洁等[17]对香梨Pyrus sinkiangensis 果柄形态的研究发现:不同类型果柄的形态结构、细胞结构物质和营养物质均存在显著差异,果柄处的纤维素、半纤维素、木质素含量以及厚壁组织的细胞壁厚度发生变化,导致香梨的抗机械损伤能力不同。ŽNIDARIČ等[18]在番茄Solanum lycopersicum模型中的研究证实,离层区远端与近端细胞的核内复制差异会通过影响膜运输系统调控器官脱落进程,这与李松刚等[19]在对龙眼Dimocarpus longan的研究中揭示的果柄离层分离力动态变化规律具有相似性。值得注意的是,4—6月梅雨期与山核桃果实快速发育阶段高度重叠,持续低温和过量降水导致幼果细胞膨压下降,离层区纤维素酶活性升高,最终使该时段落果率增加。进入7月后,日均温升至26~30 ℃,促进离层区愈伤组织形成,使落果率下降[20]。基于上述规律,建议在5—6月关键窗口期实施靶向干预,现有保果技术主要依赖外源生长调节剂和营养调控。未来研究可聚焦于:①离层特异性启动子驱动RNAi技术抑制脱落相关基因表达;②纳米载体递送系统实现离层区精准施药[21]。
-
活性氧的积累与植物细胞的氧化损伤密切相关。在山核桃的生理落果过程中,活性氧的过度积累可能导致细胞膜的脂质过氧化,进而破坏细胞膜的完整性,导致细胞内容物渗漏,激活植物防御系统,最终引发果实脱落[22]。植物体内的抗氧化系统能够清除过量的活性氧,维持细胞内的氧化还原平衡。此外,植物通过生理和环境信号传递的方式也可以调节活性氧动态平衡。在山核桃中,抗氧化酶的活性变化与落果现象密切相关。例如,当山核桃受到干旱胁迫时,外源应用褪黑素可以调节抗氧化酶的活性,减少活性氧的积累,从而减轻氧化损伤并提高果实的抗逆性[23]。植物器官脱落过程伴随显著的氧化还原状态重塑。在植物正常的生长发育过程中,活性氧水平较低,当植物受到干旱、盐、病虫害等逆境胁迫时,体内活性氧显著增加。植物器官脱落受到活性氧和抗氧化酶的影响。已有研究表明:活性氧的水平能够影响拟南芥Arabidopsis thaliana细胞壁的降解和细胞分离过程,进而控制花器官脱落区细胞的程序性死亡[24]。转录组分析也发现拟南芥果荚的脱落与过氧化物酶基因的表达相关[25],因此活性氧可能是植物器官脱落的重要信号。
碘化钾和没食子酸丙酯是2种还原性化合物,具有活性氧清除能力,在化工领域研究上常被用来检测活性氧的水平,但在果实发育及落果控制上的应用潜力还少有研究。本研究利用3.0 mmol·L−1 碘化钾和0.5 mmol·L−1 没食子酸丙酯处理山核桃果实,发现处理组离层区活性氧积累显著降低。显微观察显示:处理组离层细胞保持完整层状结构,而对照组出现典型离层空腔化特征。通过活性氧清除剂碘化钾和没食子酸丙酯的作用,山核桃果实体内的活性氧水平降低,且不同清除剂可能通过差异化的分子途径发挥作用。
-
活性氧与果实脱落之间存在着密切的关系。对拟南芥果实进行3,3'-二氨基联苯胺和硝基蓝四氮唑染色可以测定体内活性氧水平[26]。3,3'-二氨基联苯胺和硝基蓝四氮唑染色表明:已脱落山核桃果实中离层位置的活性氧水平高于正常生长的果实。从山核桃树的落果率统计可知:活性氧清除剂可以调控山核桃树的落果率,表明活性氧水平的高低影响果实的脱落,因此降低山核桃果实体内活性氧水平可以减少落果发生。在山核桃果实上喷施不同浓度的碘化钾和没食子酸丙酯,定期统计落果数量,可以筛选合适的活性氧清除剂浓度。2023和2024年统计数据表明:5—6月山核桃树的平均落果率超过80%。有研究表明:麻核桃Juglans hopeiensis的落果率为76.73%~94.77%[2],薄壳山核桃等也有较高的落果率[27]。经过3.0 mmol·L−1 碘化钾和0.5 mmol·L−1 没食子酸丙酯处理后的山核桃落果率均显著降低,经过3.0 mmol·L−1 碘化钾处理后,2023和2024年落果率分别为57.24%±8.60%和61.22%±2.75%;经过0.5 mmol·L−1 没食子酸丙酯处理后,2023和2024年落果率分别为51.85%±3.15%、45.77%±2.17%。活性氧清除剂的使用显著减少了山核桃落果的发生。
当前保果技术多聚焦于栽培管理(如环剥、施肥),对分子调控机制的研究相对滞后。本研究为提高山核桃产量提供了新思路:①开发基于活性氧传感器的精准施药系统;②构建“清除剂-微肥-生长调节剂”复合配方;③筛选耐脱落种质资源。这些技术对香榧Torreya grandis ‘Merrillii’ (胚败育率>60%)、柑橘Citrus reticulata (6月落果率约50%)等木本经济作物同样具有借鉴价值。未来需重点解析:活性氧信号与激素网络(乙烯和脱落酸)的互作机制;离层区细胞壁重构的氧化还原调控途径;施用清除剂对果实品质的长期影响。
-
本研究系统揭示了山核桃的生理落果特征,发现在5—6月山核桃的落果率高达72.22%~95.00%,主要发生在幼果形成初期,授粉不良和营养竞争是主要原因。此外,山核桃果实的结缔位置和支撑形态也可能影响落果的发生。研究还发现:山核桃果实发生落果时离层处的活性氧水平更高,而喷施3.0 mmol·L−1 碘化钾和0.5 mmol·L−1 没食子酸丙酯等活性氧清除剂可有效降低落果率。
Study on the physiological abscission patterns and control techniques in Carya cathayensis
-
摘要:
目的 山核桃 Carya cathayensis落果严重影响了产量和经济效益。揭示浙江山核桃果实脱落特征,探讨2种活性氧清除剂(碘化钾和没食子酸丙酯)在缓解山核桃落果中的调控作用。 方法 以浙江淳安、临安地区山核桃为研究对象,在2023和2024年分别喷施不同浓度的碘化钾(3.0、5.0和10.0 mmol·L−1)和没食子酸丙酯(0.1、0.3和0.5 mmol·L−1)溶液,以喷施水作为对照组,计算落果率。通过3,3'-二氨基联苯胺(DAB)和硝基蓝四氮唑(NBT)染色法检测果实组织活性氧水平,分析果实脱落过程中活性氧的动态变化。 结果 山核桃在4—6月出现大量生理落果,2023和2024年的自然落果率分别达89.55%±2.43%和80.09%±4.05%。果实脱落有2种特征:果实直接与枝条分离或果实连同果柄与枝条分离。2023和2024年喷施活性氧清除剂处理后,3.0 mmol·L−1碘化钾处理组的落果率分别为57.24%±8.60%和61.22%±2.75%;0.5 mmol·L−1 没食子酸丙酯处理组的落果率分别为51.85%±3.15%和45.77%±2.17%,均显著低于对照(P<0.05)。组织化学分析显示:离区活性氧水平在脱落前明显升高,而碘化钾和没食子酸丙酯处理有效降低了果实组织中的活性氧积累。 结论 山核桃落果集中发生于4—6月。活性氧在离层形成过程中起关键作用,3.0 mmol·L−1 碘化钾和0.5 mmol·L−1 没食子酸丙酯通过清除活性氧显著降低了落果率。图4表1参27 Abstract:Objective Carya cathayensis is important economic tree species in China, suffers from severe fruit abscission, which significantly affects yield and economic benefits. This study aims to reveal the fruit abscission characteristics of C. cathayensis and explore the regulatory effects of 2 reactive oxygen species (ROS) scavengers (potassium iodide and propyl gallate) on fruit abscission. Method C. cathayensis from Chun’an and Lin’an in Zhejiang were used as research subjects. Solutions of potassium iodide (KI) and propyl gallate (PG) at different concentrations were sprayed in 2023 and 2024, water spray served as the control, and the fruit abscission rate was calculated, KI concentrations were 3.0, 5.0, and 10.0 mmol·L−1; PG concentrations were 0.1, 0.3, and 0.5 mmol·L−1, all confidence intervals for mean differences were calculated at the 95% confidence level. The ROS content in fruit tissues was detected by 3,3'-diaminobenzidine (DAB) and nitroblue tetrazolium (NBT) staining methods to analyze the dynamic changes of ROS during fruit abscission. Result A large amount of physiological fruit abscission occurred in C. cathayensis from April to June, with natural abscission rates reaching 89.55%±2.43% in 2023 and 80.09%±4.05% in 2024. There were 2 types of fruit abscission: the fruit directly separated from the branch or the fruit separated from the branch together with the pedicel. After treatment with ROS scavengers, the fruit abscission rates were 57.24%±8.60% (2023) and 61.22%±2.75% (2024) for the 3 mmol·L−1 KI treatment group, and 51.85%±3.15% (2023) and 45.77%±2.17% (2024) for the 0.5 mmol·L−1 PG treatment group. Histological analysis showed that the ROS levels in the abscission zone significantly increased before abscission, while KI and PG treatments effectively reduced ROS accumulation in fruit tissues. Conclusion Fruit abscission in C. cathayensis occurring predominantly from April to June. ROS play a key role in the formation of the abscission layer, and 3 mmol·L−1 KI and 0.5 mmol·L−1 PG significantly reduced the fruit abscission rate by scavenging ROS. [Ch, 4 fig. 1 tab. 27 ref.] -
Key words:
- Carya cathayensis /
- fruit abscission /
- abscission zone /
- reactive oxygen species (ROS) /
- ROS scavengers
-
图 1 山核桃果实脱落过程中产生离层
Figure 1 Formation of the abscission zone during the C. cathayensis fruit abscission process
图 3 山核桃果实3,3'-二氨基联苯胺染色(A、B)和硝基蓝四氮唑染色(C)
Figure 3 DAB staining (A, B) and NBT staining (C) of C. cathayensis fruits
图 4 碘化钾和没食子酸丙酯处理后山核桃果实3,3'-二氨基联苯胺和硝基蓝四氮唑染色结果
Figure 4 DAB and NBT staining of potassium iodide and propyl gallate treated C. cathayensis fruits
表 1 山核桃在不同试剂处理后的落果率
Table 1. Fruit drop rate of C. cathayensis after treatment with different reagents
试剂 处理浓度/
(mmol·L−1)落果率/% 2023年 2024年 对照 89.55±2.43 a 80.09±4.05 ab 碘化钾 3.0 57.24±8.60 c 61.22±2.75 c 5.0 83.62±7.38 a 76.42±0.80 b 10.0 76.72±4.51 ab 88.75±5.73 a 没食子
酸丙酯0.1 78.03±2.46 ab 54.47±1.34 cd 0.3 66.66±3.89 bc 58.62±2.68 c 0.5 51.85±3.15 c 45.77±2.17 d 说明:同列不同字母表示不同处理与对照间差异显著 (P<0.05)。  下载: 导出CSV
下载: 导出CSV
-
[1] 苑轲, 黄坚钦, 王克涛, 等. 山核桃寡核苷酸探针开发及其应用[J]. 林业科学, 2023, 59(5): 88−99. YUAN Ke, HUANG Jianqin, WANG Ketao, et al. Development and application of oligonucleotide probes for the genus Carya[J]. Scientia Silvae Sinicae, 2023, 59(5): 88−99. DOI: 10.11707/j.1001-7488.LYKX20220397. [2] 朱轶群, 王红霞, 刘凯, 等. 麻核桃落果规律及影响因素[J]. 北方园艺, 2020(9): 46−54. ZHU Yiqun, WANG Hongxia, LIU Kai, et al. Regularity and influence factors of fruits drop in Juglans hopeiensis Hu[J]. Northern Horticulture, 2020(9): 46−54. DOI: 10.11937/bfyy.20193429. [3] WEN Yue, SHAO B, HAO Zhichao, et al. Preliminary study on programmed cell death during Calyx abscission of Korla fragrant pear[J]. Horticulturae, 2024, 10(6): 637. DOI: 10.3390/horticulturae10060637. [4] QIU Zhilang, WEN Zhuang, YANG Kun, et al. Comparative proteomics profiling illuminates the fruitlet abscission mechanism of sweet cherry as induced by embryo abortion[J]. International Journal of Molecular Sciences, 2020, 21(4): 1200. DOI: 10.3390/ijms21041200. [5] 杨加虎, 丁志伟, 李莎, 等. 果树器官脱落研究现状及其展望[J]. 中国蚕业, 2023, 44(3): 49−57. YANG Jiahu, DING Zhiwei, LI Sha, et al. Research status and prospect of organ abscission of fruit trees[J]. China Sericulture, 2023, 44(3): 49−57. DOI: 10.16839/j.cnki.zgcy.2023.03.010. [6] LI J, SU Shihao. Abscission in plants: from mechanism to applications[J]. Advanced Biotechnology, 2024, 2(3): 27. DOI: 10.1007/s44307-024-00033-9. [7] TAVANTI T R, MELO A A R, MOREIRA L D K, et al. Micronutrient fertilization enhances ROS scavenging system for alleviation of abiotic stresses in plants[J]. Plant Physiology and Biochemistry, 2021, 160: 386−396. DOI: 10.1016/j.plaphy.2021.01.040. [8] 郝紫微, 吴潇, 戴雨沁, 等. 赤霉素延缓‘丰水’梨落叶的生理机制[J]. 南京农业大学学报, 2021, 44(1): 61−67. HAO Ziwei, WU Xiao, DAI Yuqin, et al. The physiological mechanism of gibberellin effects on delaying leaf abscission in ‘Hosui’ pear[J]. Journal of Nanjing Agricultural University, 2021, 44(1): 61−67. DOI: 10.7685/jnau.202004030. [9] 陈静. SmMYB113调控茄子花脱落的分子机制研究[D]. 泰安: 山东农业大学, 2022. CHEN Jing. Study of the Molecular Mechanism of SmMYB113 Regulate Flower Abscission in Eggplant[D]. Tai’an: Shandong Agricultural University, 2022. DOI: 10.27277/d.cnki.gsdnu.2022.001177. [10] 史文灏, 张立新, 高品一, 等. 植物抗氧化活性成分研究进展[J]. 广州化工, 2024, 52(10): 7−10. SHI Wenhao, ZHANG Lixin, GAO Pinyi, et al. Research progress anti-oxidative gradients in natural plants[J]. GuangZhou Chemical Industry, 2024, 52(10): 7−10. DOI: 10.3969/j.issn.1001-9677.2024.10.004. [11] YANG Shengji, WANG Yongsheng, ZHANG Lidan, et al. High-dose synthetic phenolic antioxidant propyl gallate impairs mouse oocyte meiotic maturation through inducing mitochondrial dysfunction and DNA damage[J]. Environmental Toxicology, 2023, 38(8): 1800−1810. DOI: 10.1002/tox.23807. [12] 胡闰茹, 丁继垲, 王皓悦, 等. 临安山核桃研究综述[J]. 南方农业, 2024, 18(2): 22−25. HU Runru, DING Jikai, WANG Haoyue, et al. A review of research on Lin’an walnut[J]. South China Agriculture, 2024, 18(2): 22−25. DOI: 10.19415/j.cnki.1673-890x.2024.02.008. [13] 魏海林, 欧阳群文, 蒋瑶. 薄壳山核桃成花坐果的主要机理、影响因素及其促进措施[J]. 湖南农业科学, 2021(8): 106−111. WEI Hailin, OUYANG Qunwen, JIANG Yao. Review on the main mechanism and factors affecting the flowering and fruit-setting of Carya illinoensis and its technical measures of high yield cultivation[J]. Hunan Agricultural Sciences, 2021(8): 106−111. DOI: 10.16498/j.cnki.hnnykx.2021.008.027. [14] 杨蕊, 张皓波, 龚加顺, 等. 外源褪黑素对青皮核桃品质及活性氧代谢的影响[J]. 食品科技, 2024, 49(10): 23−30. YANG Rui, ZHANG Haobo, GONG Jiashun, et al. Effects of exogenous melatonin on quality and active oxygen metabolism of green walnut[J]. Food Science and Technology, 2024, 49(10): 23−30. [15] 王世明. 营养物质不足是麻核桃生理落果的主要原因[J]. 中国果业信息, 2020, 37(6): 49−50. WANG Shiming. Lack of nutrients is the main reason for physiological fruit drop of Juglans regia[J]. China Fruit News, 2020, 37(6): 49−50. [16] 刘旭鹏, 王敏, 韩守安, 等. 植物器官脱落调控因素及分子机理研究进展[J]. 植物学报, 2025, 60(3): 472−482. LIU Xupeng, WANG Min, HAN Shouan, et al. Research progress on factors and molecular mechanisms regulating plant organ abscission[J]. Chinese Bulletin of Botany, 2025, 60(3): 472−482. DOI: 10.11983/CBB24133. [17] 曹艺洁, 艾沙江·买买提, 仙米斯娅·塔依甫, 等. 库尔勒香梨不同类型果柄差异比较[J]. 新疆农业科学, 2023, 60(6): 1442−1450. CAO Yijie, Aishajiang Maimaiti, Xianmisiya Tayifu, et al. Comparison of fruit stalk difference between different types of Korla fragrant pear[J]. Xinjiang Agricultural Sciences, 2023, 60(6): 1442−1450. DOI: 10.6048/j.issn.1001-4330.2023.06.017. [18] ŽNIDARIČ M T, ZAGORŠČAK M, RAMŠAK Ž, et al. Chloroplast vesiculation and induced Chloroplast vesiculation and Senescence-associated gene 12 expression during Toma to flower pedicel abscission[J]. Plant Direct, 2025, 9(1): e70035. DOI: 10.1002/pld3.70035. [19] 李松刚, 张蕾, 洪继旺, 等. 环剥与药剂处理对龙眼果实落果及果柄离层响应的影响[J]. 广东农业科学, 2023, 50(4): 60−65. LI Songgang, ZHANG Lei, HONG Jiwang, et al. Effects of girdling and chemical treatments on the response of Longan fruit and layer[J]. Guangdong Agricultural Sciences, 2023, 50(4): 60−65. DOI: 10.16768/j.issn.1004-874X.2023.04.007. [20] VAHDATI K, SADEGHI-MAJD R, SESTRAS A F, et al. Clonal propagation of walnuts (Juglans spp. ): a review on evolution from traditional techniques to application of biotechnology[J]. Plants, 2022, 11(22): 3040. DOI: 10.3390/plants11223040. [21] 李杰, 罗江宏, 杨萍. 病毒诱导基因沉默在蔬菜作物上应用的研究进展[J]. 中国农业科学, 2021, 54(10): 2154−2166. LI Jie, LUO Jianghong, YANG Ping. Research advances of applying virus-induced gene silencing in vegetables[J]. Scientia Agricultura Sinica, 2021, 54(10): 2154−2166. DOI: 10.3864/j.issn.0578-1752.2021.10.011. [22] 徐一荻, 刘春花, 谢富, 等. 供磷水平对核桃实生苗生长生理特性及酶活性的影响[J]. 西南林业大学学报(自然科学), 2021, 41(5): 27−35. XU Yidi, LIU Chunhua, XIE Fu, et al. Effects of phosphorus supply on the growth, physiological characteristics and enzyme activity of Juglans regia seedlings[J]. Journal of Southwest Forestry University (Natural Sciences), 2021, 41(5): 27−35. DOI: 10.11929/j.swfu.202004052. [23] HUANG Qian, YAN Haijing, YOU Mingyuan, et al. Enhancing drought tolerance and fruit characteristics in Toma to through exogenous melatonin application[J]. Horticulturae, 2023, 9(10): 1083. DOI: 10.3390/horticulturae9101083. [24] LEE Jinsu, CHEN Huize, LEE G, et al. MSD2-mediated ROS metabolism fine-tunes the timing of floral organ abscission in Arabidopsis[J]. New Phytologist, 2022, 235(6): 2466−2480. DOI: 10.1111/nph.18303. [25] ELJEBBAWI A, SAVELLI B, LIBOUREL C, et al. Class Ⅲ peroxidases in response to multiple abiotic stresses in Arabidopsis thaliana Pyrenean populations[J]. International Journal of Molecular Sciences, 2022, 23(7): 3960. DOI: 10.3390/ijms23073960. [26] WANG Jiehua, MOEEN-UD-DIN M, YIN Rong, et al. ROS homeostasis involved in dose-dependent responses of Arabidopsis seedlings to copper toxicity[J]. Genes, 2022, 14(1): 11. DOI: 10.3390/genes14010011. [27] 黄锈贤, 黄琼, 谢伟东, 等. 传粉及授粉方式对薄壳山核桃‘Shaoxing’结实的影响[J]. 中国南方果树, 2022, 51(1): 124−129. HUANG Xiuxian, HUANG Qiong, XIE Weidong, et al. Effects of pollination and pollination methods on the fruiting of Carya illinoensis ‘Shaoxing’[J]. South China Fruits, 2022, 51(1): 124−129. DOI: 10.13938/j.issn.1007-1431.20210183. -

-
链接本文:
https://zlxb.zafu.edu.cn/article/doi/10.11833/j.issn.2095-0756.20250284
 点击查看大图
点击查看大图

计量
- 文章访问数: 441
- HTML全文浏览量: 77
- PDF下载量: 92
- 被引次数: 0
