


 下载:
下载:
 下载:
下载:
-
菊花Chrysanthemum × morifolium是多年生宿根草本花卉,其鲜切花年产量居所有花卉品种的前列。菊花具短日、喜冷凉的特性,光周期不敏感和耐热特性的夏菊是菊花育种的重要方向。夏菊由于生长开花季节处于炎热的夏季,营养生长和生殖生长都受到一定程度的影响。不同夏菊品种的耐热特性差异较大,部分品种夏季高温条件下难以正常开放,开展相应的田间和生理生化指标评价极有必要。水杨酸(salicylic acid,SA)对植物的生长发育有重要的调控作用,被认为是一种新的植物生长调节物质,它不仅能够调控植物的生长,也能够提高植物的抗逆性,抵抗不良的环境条件。杨华庚等[1]运用水杨酸对蝴蝶兰Phalaenopsis幼苗进行处理,结果显示外源水杨酸处理可以提高蝴蝶兰幼苗超氧化物歧化酶(SOD)和过氧化物酶(POD)等抗氧化酶的活性来提高其耐热性。李天来等[2]通过水杨酸处理番茄Lycopersicon esculentum幼苗表明,适宜浓度的水杨酸能够降低叶片丙二醛(MDA)的质量摩尔浓度,提高植物幼苗叶片中脯氨酸(Pro)质量分数,提高植物幼苗的抗高温能力。曹淑红等[3]运用水杨酸对高温下的百日草Zinnia elegans幼苗进行处理,结果表明:水杨酸能够明显的降低百日草幼苗的热害指数,降低叶片丙二醛含量,增加脯氨酸的质量分数,提高百日草的耐热性。目前,菊花耐热方面的相关研究较少[4-5]。本研究以夏菊品种‘粉荷’Chrysanthemum × morifolium ‘Fenhe’为材料,通过叶面喷施不同浓度的水杨酸,研究水杨酸对菊花幼苗耐热性的影响,为夏菊幼苗耐热性研究和菊花的耐热育种提供理论依据。
-
本研究于2014年3月至9月在江苏农林职业技术学院实验室进行。供试材料为夏菊品种‘粉荷’。3月20日选取生长整齐一致的植株脚芽,修剪整齐后在扦插床[蛭石:珍珠岩=2:1(体积比)]上生根。20 d后将生根苗移入8 cm的营养钵,在15~28 ℃温室中进行正常栽培管理。5月1日将幼苗放入光照培养箱,内温度设置18~28 ℃进行5 d预培养,将浓度设为0(空白),15,30,45,60 mmol·L-1的水杨酸溶液均匀喷施于菊花幼苗叶面,至溶液欲滴为度,然后转入光照培养箱进行40 ℃/30 ℃高温胁迫处理,采用倒数取点移入培养箱,分别进行0(对照),24,48,72 h高温处理,处理结束后统一取样。自上往下取第3~4片成熟的叶片,用蒸馏水洗净、混匀,测定SOD,POD,过氧化氢酶(CAT)活性和叶绿素、可溶性蛋白、可溶性糖、丙二醛、脯氨酸,测定方法参照李合生[6]的方法。3次重复,测幼苗10株·次-1。采用Excel 2010进行数据处理,结合SPSS 16.0软件进行方差分析。
-
由图 1可知:在高温胁迫下菊花幼苗‘粉荷’叶绿素的质量分数呈现先升高再降低的趋势,并且在处理24 h时达到最高值,其中30 mmol·L-1水杨酸处理的幼苗叶绿素质量分数最高,比对照增加了26.0%,达到了24.05 mg·g-1,此后叶绿素的质量分数便逐渐降低。在处理72 h后,叶绿素质量分数达到了最低值,各处理分别比对照降低了18.0%,13.1%,1.7%,10.5%和10.6%,水杨酸处理幼苗的叶绿素质量分数始终高于对照,并以30 mmol·L-1水杨酸处理的植株降低最少,仍保持相对较高水平。

图 1 高温胁迫下喷施不同浓度水杨酸对叶片叶绿素质量分数的影响
Figure 1. Chlorophyll content in response to different contents of SA under high temperature stress
-
图 2显示:随着处理时间的延长,菊花幼苗叶片丙二醛的质量摩尔浓度逐渐增加,在处理72 h以后都达到最大值,对照叶片的丙二醛质量摩尔浓度增加了158.2%。水杨酸处理叶片的丙二醛质量摩尔浓度也均增加,30 mmol·L-1处理的叶片丙二醛质量摩尔浓度增加了86.8%,为4.76 μmol·g-1,其余3个处理的叶片丙二醛质量摩尔浓度均增加了110.0%以上,但水杨酸处理的幼苗叶片丙二醛质量摩尔浓度均低于对照。
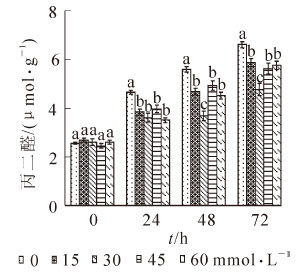
图 2 高温胁迫下喷施不同浓度水杨酸对叶片丙二醛质量摩尔浓度的影响
Figure 2. MDA content in response to different contents of SA under high temperature stress
由图 3A表明:高温胁迫下细胞脯氨酸质量分数均有不同程度的增加,其中水杨酸处理的菊花幼苗叶片脯氨酸质量分数都显著高于对照,在处理48 h时45 和60 mmol·L-1处理的叶片分别达到了最大值,为3.27 mg·g-1和3.25 mg·g-1,分别比对照增加了90.6%和86.4%;处理72 h时,30 mmol·L-1水杨酸处理幼苗的叶片脯氨酸质量分数达到最大值为3.55 mg·g-1,比对照增加了94.3%。
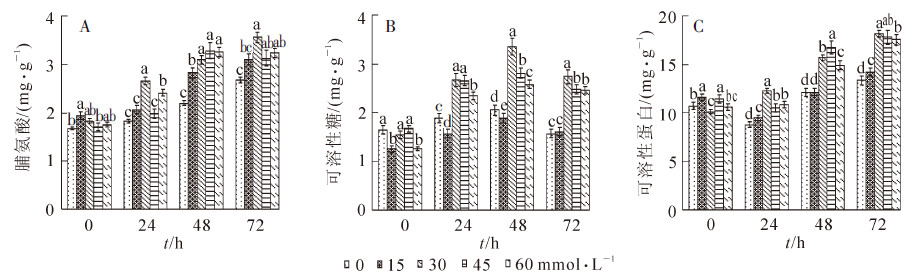
图 3 喷施不同浓度水杨酸在高温胁迫下菊花叶片脯氨酸、可溶性糖和可溶性蛋白质量分数的变化
Figure 3. roline, soluble sugar and soluble content in leaf in response to different contents of SA under high temperature stress
可溶性糖也是渗透调节的一类重要物质,在高温处理后均出现先升高后降低的趋势,其中30~60 mmol·L-1的水杨酸处理的菊花幼苗叶片的可溶性糖质量分数均显著高于其他2个处理(图 3B),且30 mmol·L-1的水杨酸处理的菊花叶片可溶性糖质量分数最高,为3.35 mg·g-1,比对照增加了112.1%;45和60 mmol·L-1水杨酸处理的叶片比对照也分别增加了67.7%和104.2%。而0(空白)和15 mmol·L-1水杨酸处理的菊花幼苗叶片可溶性糖质量分数的增加较少,分别仅有24.9%和50.6%。
可溶性蛋白质量分数随着处理时间的加长先降低再升高,其中在72 h时均达到最大值,15~60 mmol·L-1水杨酸处理的菊花幼苗叶片比对照分别增加了22.5%,80.0%,56.7%和65.9%(图 3C),0 mmol·L-1(空白)仅比对照增加了24.8%。
-
由图 4A可知:在高温胁迫的过程中,叶片的SOD活性变化趋势基本相同。在高温胁迫24 h后SOD的活性升高到最大值,60 mmol·L-1水杨酸处理的幼苗SOD活性最高为274×16.67 nkat·g-1,比对照增加53.7%,30 mmol·L-1水杨酸处理的幼苗活性比对照增加了37.4%;胁迫48 h后SOD的活性开始降低,水杨酸处理的对幼苗SOD活性的增加不明显,但30 mmol·L-1的水杨酸处理幼苗SOD活性比对照的高;72 h后水杨酸处理的幼苗SOD活性均比对照活性高,说明水杨酸处理能够提高菊花幼苗SOD活性,并且以30 mmol·L-1的水杨酸效果最好。
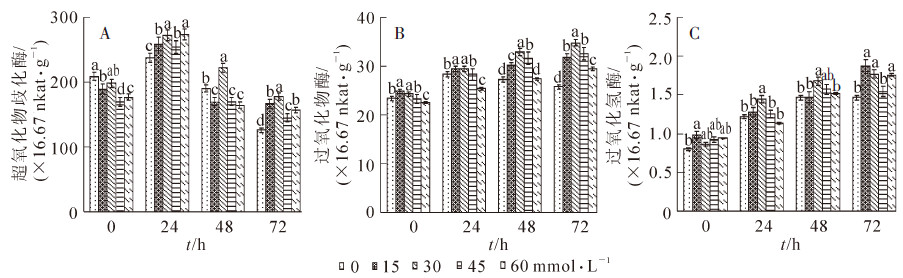
图 4 喷施不同浓度水杨酸对高温胁迫下菊花叶片超氧化物歧化酶(SOD)、过氧化氢酶(POD)、过氧化氢酶(CAT)的活性变化
Figure 4. Activities of SOD, POD, CAT in leaf in response to different contents of SA under high temperature stress
POD活性的变化趋势与SOD活性变化不同,随着处理时间的延长POD的活性逐渐升高,并且在处理72 h后达最高值,并且水杨酸处理对于菊花幼苗POD活性的增加效果显著,且30 mmol·L-1的水杨酸处理对于菊花幼苗POD活性的增加最显著,达到了1.77×16.67 nkat·g-1·min-1(图 4B),比对照增加了43.0%。CAT活性的变化与POD有相似的趋势,也是随着处理时间的延长活性逐渐增加,并且在72 h达到了最大值。各处理幼苗叶片CAT活性均比对照增加了80%以上(图 4C),其中30 mmol·L-1水杨酸处理的幼苗叶片比对照增高了103.8%,增加幅度最大。
-
植物在受到逆境胁迫时会导致体内一系列的生理变化,如叶绿素降解,膜透性增大,活性氧增多,诱导使体内的渗透调节系统和抗氧化系统应答。水杨酸作为一种新型的植物生长调节物质,能够有效减轻高温、干旱等逆境对植物的影响[7]。水杨酸主要通过提高植物体内可溶性物质质量分数、增加抗氧化系统的活性来提高植物适应逆境的能力。近年来对于水杨酸的研究成了新的热点。高温胁迫会引起植物膜脂过氧化,甚至破坏叶绿体结构,降低叶绿素质量分数,但随着胁迫时间的延长,叶绿素质量分数则趋于稳定[8]。本研究中叶绿素的质量分数也出现先升高后降低,逐渐趋于稳定。虽然水杨酸处理并没有改变叶绿素降低的趋势,但水杨酸处理的幼苗叶片的叶绿素质量分数始终高于对照,说明水杨酸处理能够减轻叶片的高温伤害,提高叶片的抗高温能力。这与杨岚等[8]在铁皮石斛Dendrobium officinale中的研究相一致。
植物受高温胁迫时,细胞内活性氧产生加速,清除活性氧的能力下降,活性氧的增加使得细胞膜发生脂质过氧化作用同时产生丙二醛,植物体内丙二醛质量摩尔浓度的高低是植物活性氧浓度和脂质过氧化程度的重要指标。植物在受到逆境伤害后也能够通过自身的游离脯氨酸、可溶性糖等渗透调节物质来增加细胞的渗透势从而减轻细胞所受的伤害,它也可以通过诱导特异蛋白的表达来减轻细胞的伤害程度。可溶性渗透调节物质在植物的抗逆过程中起着重要作用,它们可以保持原生质体与环境的渗透平衡,防止细胞质脱水[1, 8]。本试验中水杨酸处理减少了夏菊叶片丙二醛的质量摩尔浓度,增加了叶片可溶性糖、可溶性蛋白和脯氨酸的质量分数,并且以30 mmol·L-1的水杨酸效果最好。这说明水杨酸处理降低了高温下细胞膜脂质过氧化作用,通过增加渗透调节物质从而减轻了细胞膜的伤害,与孙军利[9]、曹淑红等[3]的研究结果一致。
超氧化物岐化酶(SOD),过氧化物酶(POD)以及过氧化氢酶(CAT)是细胞清除活性氧系统的重要酶[10-11],在植物抵抗不良环境中起到重要的作用。叶凡等[12]研究表明,耐热性强的白菜Brassica pekinensis幼苗在受到高温胁迫时,SOD,CAT和抗坏血酸过氧化物酶(APX)的活性都较不耐热的品种稳定。水德聚等[7]在研究外源水杨酸对高温下白菜耐热性时表明,外源水杨酸能够提高SOD和POD等酶的活性,来提高白菜幼苗的耐热性。本试验中通过测量SOD,CAT和POD等3种酶的活性变化表明:在高温胁迫下水杨酸处理的夏菊幼苗的3种酶活性均高于对照,其中高温胁迫对于SOD活性影响最大,在水杨酸处理后呈现先升高后降低的趋势。POD和CAT的活性在高温处理后则一直呈现上升的趋势,说明高温胁迫下夏菊叶片中POD和CAT更好地起到了清除胁迫产生的活性氧和过氧化物的作用,防止细胞膜发生膜脂过氧化,降低高温对叶片的伤害。在整个过程中,水杨酸处理后叶片CAT和POD活性与对照相比大部分能提高,说明水杨酸处理起到了提高保护酶的活性作用,这与前人的研究一致[13-14],并且30 mmol·L-1的水杨酸处理效果最好。
Physiological and biochemical indexes of exogenous salicylic acid on chrysanthemum seedlings with high temperature stress
-
摘要: 为研究高温胁迫下水杨酸(SA)对菊花Chrysanthemum × morifolium幼苗热伤害的缓解作用,以扦插40 d的夏菊幼苗为材料,向叶片喷施不同浓度[0(空白),15,30,45,60 mmol·L-1]的水杨酸溶液,研究在不同高温时间[0(对照),24,48,72 h]处理下叶片叶绿素、丙二醛、脯氨酸、可溶性糖和可溶性蛋白的含量,以及超氧化物歧化酶(SOD),过氧化物酶(POD),过氧化氢酶(CAT)的活性。结果表明:喷施水杨酸能够减轻菊花叶片叶绿素质量分数的降低速度,用30 mmol·L-1水杨酸处理72 h后叶片叶绿素质量分数仅比对照(未进行高温处理)降低了1.7%;高温处理后,使用不同浓度的水杨酸处理能够减轻菊花叶片丙二醛质量摩尔浓度的增加,增加脯氨酸、可溶性蛋白、可溶性糖的质量分数,以30 mmol·L-1的水杨酸处理72 h的效果最好。菊花叶片丙二醛质量摩尔浓度仅比对照增加了86.8%,为4.76 μmol·g-1,脯氨酸、可溶性糖、可溶性蛋白的质量分数则分别增加了94.3%,112.1%和80.0%,均高于其他处理;高温处理后,SOD活性呈现先升高再降低的趋势,POD和CAT活性逐渐升高,而且30 mmol·L-1水杨酸对于SOD和POD的影响最为明显,15 mmol·L-1水杨酸对于CAT的影响最显著。结果表明:短期高温胁迫引起了菊花叶片的叶绿素、可溶性渗透调节物质和抗氧化系统都受到了不容程度的伤害,水杨酸能起到一定的保护作用,减轻高温对叶片的影响。图4参14Abstract: To study the relief of salicylic acid (SA) on thermal damage in chrysanthemum seedlings with high temperature stresses of 0, 24, 48, and 72 h, cuttings from 40-day-old summer chrysanthemum seedlings were used as materials. Leaves were sprayed with different concentrations of SA (0, 15, 30, 45, and 60 mmol·L-1). Then the content of chlorophyll, malondialdehyde (MDA), proline, free soluble sugar, and free soluble protein, were studied along with activities of superoxide dismutase (SOD), peroxidase (POD), and hydrogen peroxide (CAT) enzyme. Results showed that compared with the controls, the 30 mmol·L-1 salicylic acid treatment on the leaf for 72 h reduced leaf chlorophyll content 1.7%, have significant difference (P < 0.05). After high temperature treatment, best results were with the 30 mmol·L-1 salicylic acid treatment where compared to the control the MDA content increased 86.8%, have significant difference(P < 0.05). The content of proline (94.3%), free soluble sugar (112.1%), and free soluble protein (80%) significantly increased than other treatments (P < 0.05). SOD activity first increased, then decreased, whereas POD and catalase (CAT) activity increased. The 30 mmol·L-1 SA concentration had the greatest effect on SOD and POD, and 15 mmol·L-1 SA had the greatest effect on CAT. Thus, a short duration of high temperature stress damaged chrysanthemum leaves, but SA could play a protective role mitigating the damage. [Ch, 4 fig. 14 ref.]
-
在城市化快速发展的背景下,城镇建设用地的扩张导致生态空间衰减、系统结构失衡、生态功能下降等问题凸显[1],生态环境面临多重压力和干扰,引起的景观生态风险值得关注。绿色空间是城镇地域范围内对于改善区域生态环境、维持生态系统物质能量循环具有重要作用的生态空间,是由耕地、林地、草地、水域等不同土地单元镶嵌而成的复合生态系统[2−4]。当前,国内外学者对绿色空间的研究主要集中在绿色空间结构与功能[5]、景观格局动态演化[6−7]及生态环境效益[8−9]等方面。景观生态风险评价用于评估自然或人为因素干扰对生态系统及其组分产生不利影响的可能性及损失[10],基于景观格局指数构建景观生态风险评价模型能够定量揭示生态环境健康程度及风险压力的时空分布特征[11]。现有研究主要集中于景观生态风险的静态分析,对时空动态分析视角下景观生态风险演变特征的分析相对薄弱,且研究尺度集中在城市[12−13]、城市群[14−15]、流域[16−17]等典型地区,对具有特殊地域特征的苏南水网地区的研究相对较少。
苏南水网地区位于经济发达、人口密集的长江三角洲,河流、湖荡众多,水系纵横交错,形成了独特的地域生态空间特征。随着城镇建设用地的迅速扩张,苏南水网地区绿色空间日趋破碎化,生态系统稳定性下降。本研究以苏南水网地区江苏省昆山市为研究对象,利用2000、2010、2020年土地利用数据,定量测度其绿色空间景观格局变化引起的景观生态风险,并探究景观生态风险时空演变特征,依据风险等级转移变化特征划定绿色空间管控分区,提出分区调控策略,为优化水网地区空间景观布局,保护地区生态安全,合理开发绿色空间资源提供理论依据,也为地区景观生态风险管理提供决策支持。
1. 研究区概况与数据来源
1.1 研究区概况
昆山市位于长江三角洲地区江苏省苏州市东部,31°06′~31°32′N,120°48′~121°09′E,全市下辖周庄镇、锦溪镇、淀山湖镇等10个镇,总面积为931 km2。根据《昆山市统计年鉴》,2000—2020年昆山市户籍总人数增加47.3万人,城镇化率由57.31%提升至78.95%,国内生产总值(GDP)增长4 075.96亿元,经济建设水平居于全国经济百强县首位。昆山市境内地势平坦,属北亚热带季风性湿润气候,四季分明,雨量充沛。境内河港纵横交错,湖荡星罗棋布,水域面积占16.4%,包含白莲湖、傀儡湖、明镜荡等湖荡,水网地区风貌特征明显。
1.2 数据来源及处理
采用2000、2010、2020年3期 Landsat TM/OLI 遥感影像,数据集来源于地理空间数据云平台(
http://www.gscloud.cn/ ),空间分辨率为30 m×30 m。利用ENVI 5.3软件对各期遥感影像数据进行校准、图像拼接裁剪等处理。参考中国科学院土地利用/土地覆盖分类系统及GB/T 21010—2017《土地利用现状分类》相关标准,结合苏南水网地区地域特点,将研究区划分为耕地、林地、草地、水域、建设用地和未利用地等6类土地利用类型,其中耕地、林地、草地和水域为绿色空间,建设用地和未利用地为非绿色空间。对解译后的土地利用类型数据进行精度验证,Kappa系数均>0.85,符合解译精度要求。2. 研究方法
2.1 风险小区的划分
为了便于景观生态风险指数的空间化表达,本研究基于ArcGIS的渔网分析功能划分景观生态风险小区。依据研究区面积大小及数据精度,采用等间距采样的方法将研究区划分为1.5 km×1.5 km正方形格网,共划分景观生态风险样本小区489个(图1),利用 Fragstats 4.2软件计算各个样本小区内的生态风险指数,作为每个风险小区中心点的景观生态风险值。
2.2 景观生态风险评价模型
景观格局指数是反映景观结构组成和空间配置特征的定量指标[18]。基于景观格局指数的生态风险评价方法能够有效评估生态系统受到外部干扰的强弱和内部抵抗力的大小[19]。根据相关研究成果[20−21],依据景观格局与生态风险之间的关联,选取景观干扰度指数($ {E}_{i} $)、景观脆弱度指数($ {V}_{i} $)和景观损失度指数($ {R}_{i} $)来构建景观生态风险评价模型。
各景观格局指数计算方法及生态学含义详见表1。
表 1 景观格局指数及计算方法Table 1 Landscape pattern index and their calculation methods指数名称 计算方法 生态学含义 土地利用生态风险指数
(IERk)${I_{{\text{ER}}k}} = \displaystyle \sum \limits_{i = 1}^N \dfrac{{{A_{ki}}}}{{{A_k}}} \times {R_i} $ Aki为第k个风险小区内土地利用类型i的面积;Ak为第k个风险小区的面积;Ri为第i类景观的景观损失度指数 景观损失度指数(Ri) Ri=Ei×Vi Ei为景观干扰度指数,Vi为景观脆弱度指数 景观干扰度指数($ {E}_{i} $) $ {E}_{i}={aC}_{i}+{bN}_{i}+{cD}_{i} $ 表示不同类型景观生态系统所受外界干扰的程度,主要与人类的开发活动有关。其中:$ a、b、c $分别为$ {C}_{i} $、$ {N}_{i}{\mathrm{、}D}_{i} $的权重,且$ a+b+c= $1,参考前人研究[11, 22],将$ a、b、c $分别赋值为0.5、0.3和0.2 景观破碎度指数($ {C}_{i} $) $ {C}_{i}=\dfrac{{n}_{i}}{{A}_{i}} $ 表示景观被分割的破碎化程度,值越大表明景观破碎程度越高 景观分离度指数($ {N}_{i} $) $ {N}_{i}=\dfrac{A}{2{A}_{i}}\sqrt{\dfrac{{n}_{i}}{A}} $ 表示某一景观类型中不同斑块间的分离程度,值越大表明景观空间分布越离散,景观结构稳定性越低。$ {n}_{i} $为景观类型$ i $的斑块个数;$ {A}_{i} $为景观类型$ i $的面积;$ A $为景观总面积 景观优势度指数($ {D}_{i} $) $ {D}_{i}=\dfrac{\left(\dfrac{{n}_{i}}{N}+\dfrac{{q}_{i}}{Q}\right)}{4}+\dfrac{{A}_{i}}{2A} $ 表示斑块在景观中的地位,值越大代表斑块对景观格局演变影响越大。$ {q}_{i} $为景观类型$ i $斑块出现的样方数;$ Q $为样方总数;$ N $为斑块总数 景观脆弱度指数($ {V}_{i} $) $ {V}_{i}={I}_{{\mathrm{LS}}}\times \left(1-{I}_{{\mathrm{LA}}}\right) $ 表示不同景观类型抵抗外界干扰的敏感程度。其中:ILS为景观敏感度指数,可通过景观干扰度指数和景观易损度指数相乘而得,景观易损度指数根据前人研究成果[23−24],结合研究区实际情况赋以权重:未利用地为6,水域为5,耕地为4,草地为3,林地为2,建设用地为1;ILA为景观适应度指数,由斑块丰富密度指数、香农多样性指数、香农均匀度指数相乘而得。3种指数均由Fragstats软件计算而得 3. 结果与分析
3.1 昆山市绿色空间演变特征分析
3.1.1 绿色空间面积组成对比分析
通过ArcGIS软件对遥感影像图进行分类处理,得到昆山市2000、2010和2020年3个时期土地利用类型图(图2),并统计得到各土地利用类型面积与占比(表2)。从分析结果来看:2000—2020年昆山市各绿色空间类型面积发生了较大变化,其中耕地面积下降明显,减少20 203.11 hm2,占比下降21.70%;水域面积在2000—2010年小幅增加,占比上升2.24%,2010—2020年水域面积明显下降,减少了
5905.17 hm2,占比下降6.34%;林地面积共减少72.90 hm2,而草地面积则增加了143.64 hm2,两者在绿色空间中占比很小。总体而言,研究期间昆山市绿色空间总面积明显减少,反映了建设用地扩张不断侵占市域内的绿色空间,以耕地面积的缩减最为突出。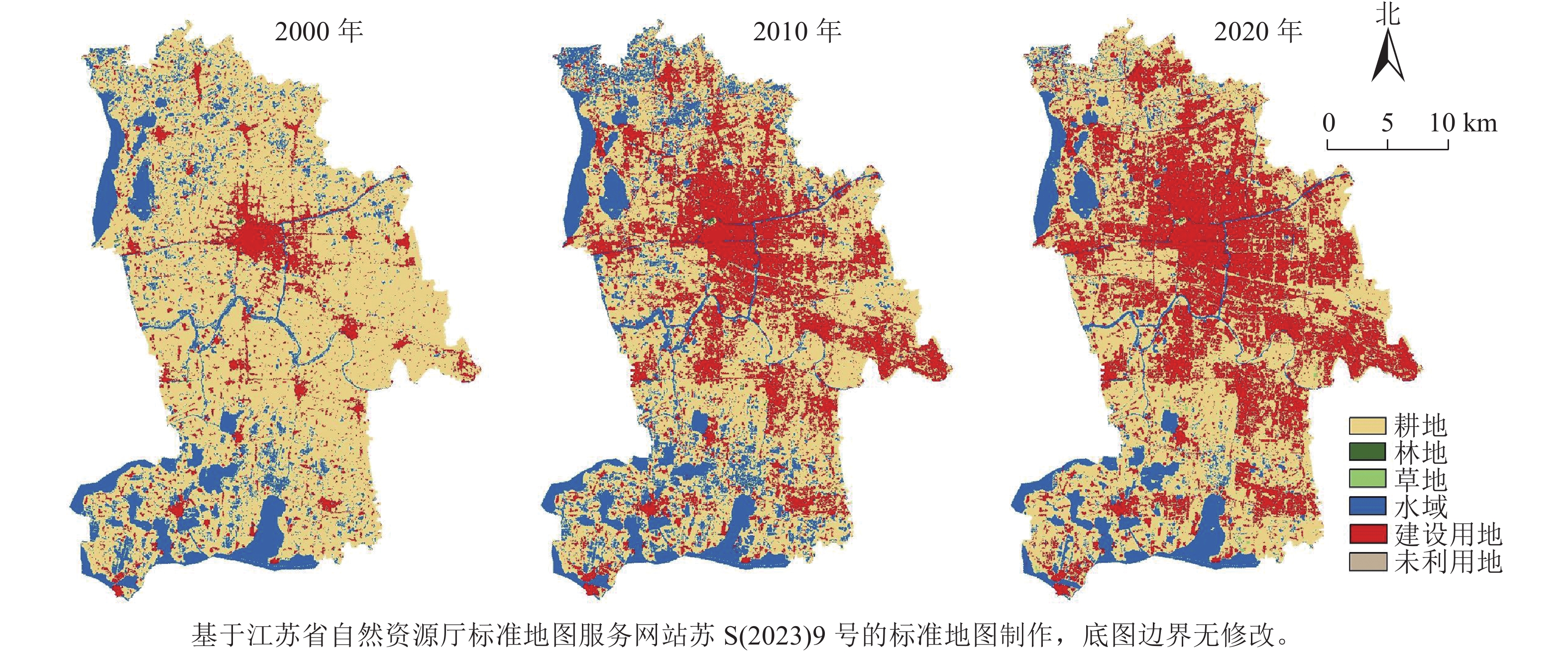 图 2 2000—2020年昆山市土地利用类型示意图Figure 2 Land use type map of Kunshan City from 2000 to 2020表 2 2000—2020年昆山市各用地类型面积变化Table 2 Changes in the area of various land types in Kunshan City from 2000 to 2020
图 2 2000—2020年昆山市土地利用类型示意图Figure 2 Land use type map of Kunshan City from 2000 to 2020表 2 2000—2020年昆山市各用地类型面积变化Table 2 Changes in the area of various land types in Kunshan City from 2000 to 2020土地利用类型 2000年 2010年 2020年 面积/hm2 百分比/% 面积/hm2 百分比/% 面积/hm2 百分比/% 绿色空间 耕地 68 884.11 73.98 51 240.51 55.03 48 681.00 52.28 林地 122.85 0.13 112.59 0.12 49.95 0.05 草地 36.36 0.04 78.66 0.08 180.00 0.19 水域 15 156.36 16.28 17 247.87 18.52 11342.70 12.18 合计 84 199.68 90.43 68 679.63 73.75 60253.65 64.70 非绿色空间 建设用地 8 833.95 9.49 24 386.49 26.19 32828.04 35.26 未利用地 81.36 0.09 48.96 0.05 33.39 0.05 合计 8 915.31 9.58 24 435.45 26.24 32861.43 35.31 3.1.2 绿色空间面积转移矩阵分析
为了进一步揭示昆山市绿色空间用地类型的时空演变规律,本研究采用土地利用转移矩阵对昆山市各用地类型之间的转移方向和转换数量进行分析,结果如表3所示。2000—2020年昆山市各绿色空间类型转移存在明显差异:耕地净转出量最大,总量达33 918.84 hm2,主要流向建设用地,转出面积达26 327.86 hm2,转出贡献率为77.62%,反映出建设用地侵占耕地现象普遍;水域面积整体呈现先小幅增加后逐渐减少的趋势,其中2000—2010年水域面积小幅增加了2 091.51 hm2,主要由耕地转入,2010—2020年,水域面积持续减少,主要向耕地和建设用地转出,转出总面积为7 150.64 hm2。总体来看,2000—2020年昆山市绿色空间类型转移以耕地和水域的转出为主,均主要转向建设用地。这反映出昆山市在经济社会快速发展下人为开发建设活动对绿色空间侵占现象较为明显,耕地和水域等绿色空间面临较大生态压力。
表 3 2000—2020年昆山市地类转移矩阵Table 3 Land class transfer matrix in Kunshan City from 2000 to 2020时间段 土地利用类型 绿色空间/hm2 非绿色空间/hm2 转出合
计/hm2面积变化
合计/ hm2耕地 林地 草地 水域 建设用地 未利用地 2000—2010 绿色空间 耕地 46 613.56 41.62 71.57 6 168.10 15 825.59 6.48 68 726.91 −17 577.88 林地 46.23 41.28 0.07 29.54 5.62 0.00 122.74 −10.60 草地 14.15 0.00 0.16 15.64 6.29 0.11 36.36 42.30 水域 3 464.97 28.70 5.97 10 742.95 853.90 2.02 15 098.51 2 033.98 非绿色空间 建设用地 975.40 0.53 0.90 174.01 7 672.25 0.20 8 823.28 15 544.60 未利用地 34.72 0.00 0.00 2.25 4.23 40.15 81.36 −32.40 转入合计 51 149.03 112.14 78.66 17 132.48 24 367.89 48.96 92 889.16 − 时间段 土地利用类型 绿色空间/hm2 非绿色空间/hm2 转出合
计/ hm2面积变化
合计/ hm2耕地 林地 草地 水域 建设用地 未利用地 2010—2020 绿色空间 耕地 39 356.15 8.54 127.00 1 161.04 10 502.27 6.64 51 161.64 −2 650.12 林地 65.13 25.78 0.13 17.69 3.57 0.00 112.30 −62.47 草地 2.17 0.00 1.46 0.00 74.73 0.30 78.66 101.34 水域 6 482.98 14.97 37.18 9 906.54 667.66 6.89 17 116.24 −5 812.93 非绿色空间 建设用地 2 592.59 0.53 14.22 217.65 21 538.87 1.19 24 365.05 8 439.75 未利用地 12.50 0.00 0.00 0.38 17.71 18.37 48.96 −15.57 转入合计 48 511.52 49.82 180.00 11 303.31 32 804.80 33.39 92 882.85 − 说明:−表示无此项。 3.2 昆山市绿色空间景观生态风险时空演变分析
3.2.1 绿色空间景观格局指数时序变化
运用Fragstats软件计算得到昆山市2000、2010、2020年各绿色空间类型景观格局指数。统计结果表明:2000—2020年昆山市绿色空间景观格局发生了较大变化(表4)。①研究期间耕地破碎度和分离度指数显著上升,表明建设用地快速扩张,促使耕地空间分布趋于离散,破碎化程度加剧,景观优势度不断降低,受外界干扰程度增加。景观损失度逐年上升。②水域破碎度指数先下降后上升,总体呈上升趋势,景观优势度降低,且水域周边城镇较为密集,易受人为活动干扰,使景观脆弱程度不断增加,损失度上升。③林地破碎度、干扰度、脆弱度指数均先下降后上升,总体呈下降趋势,表明林地斑块分布逐渐聚集,景观结构稳定性提升。④草地破碎度指数先上升后下降,表明草地斑块在空间上趋于集聚与整合,抗外界干扰能力提高,景观脆弱度与损失度有所降低。
表 4 2000—2020年昆山市绿色空间景观格局指数变化Table 4 Change of green space landscape pattern index in Kunshan City from 2000 to 2020土地利用类型 年份 斑块数量 斑块面积/hm2 破碎度 分离度 优势度 干扰度 脆弱度 损失度 耕地 2000 1378 68 884.11 0.020 0.973 0.647 0.431 0.082 0.035 2010 4401 51 240.51 0.086 0.987 0.602 0.459 0.087 0.040 2020 4667 48 681.00 0.096 0.992 0.597 0.465 0.088 0.041 林地 2000 494 122.85 4.021 1.000 0.092 2.329 0.222 0.516 2010 355 112.59 3.153 1.000 0.076 1.892 0.180 0.340 2020 172 49.95 3.443 1.000 0.046 2.031 0.193 0.392 草地 2000 56 36.36 1.540 1.000 0.025 1.075 0.153 0.165 2010 195 78.66 2.479 1.000 0.017 1.551 0.221 0.343 2020 123 180.00 0.683 1.000 0.047 0.651 0.093 0.060 水域 2000 4128 15 156.36 0.272 1.000 0.417 0.520 0.124 0.064 2010 3566 17 247.87 0.207 1.000 0.399 0.483 0.115 0.056 2020 3770 11 342.70 0.332 1.000 0.365 0.539 0.128 0.069 3.2.2 绿色空间景观生态风险时空分布格局
基于景观生态风险评价指标计算结果,在ArcGIS 10.2中利用克里金插值法对昆山市生态风险值进行空间插值,得到昆山市绿色空间景观生态风险空间分布图,使用自然断点法将景观生态风险值(IERk)划分为5个等级:低生态风险(0<IERk≤0.026)、较低生态风险(0.026<IERk≤0.031)、中生态风险(0.031<IERk≤0.037)、较高生态风险(0.037<IERk≤0.041)和高生态风险(IERk>0.041),结果如图3,并统计得到不同景观生态风险等级的面积及占比(表5)。
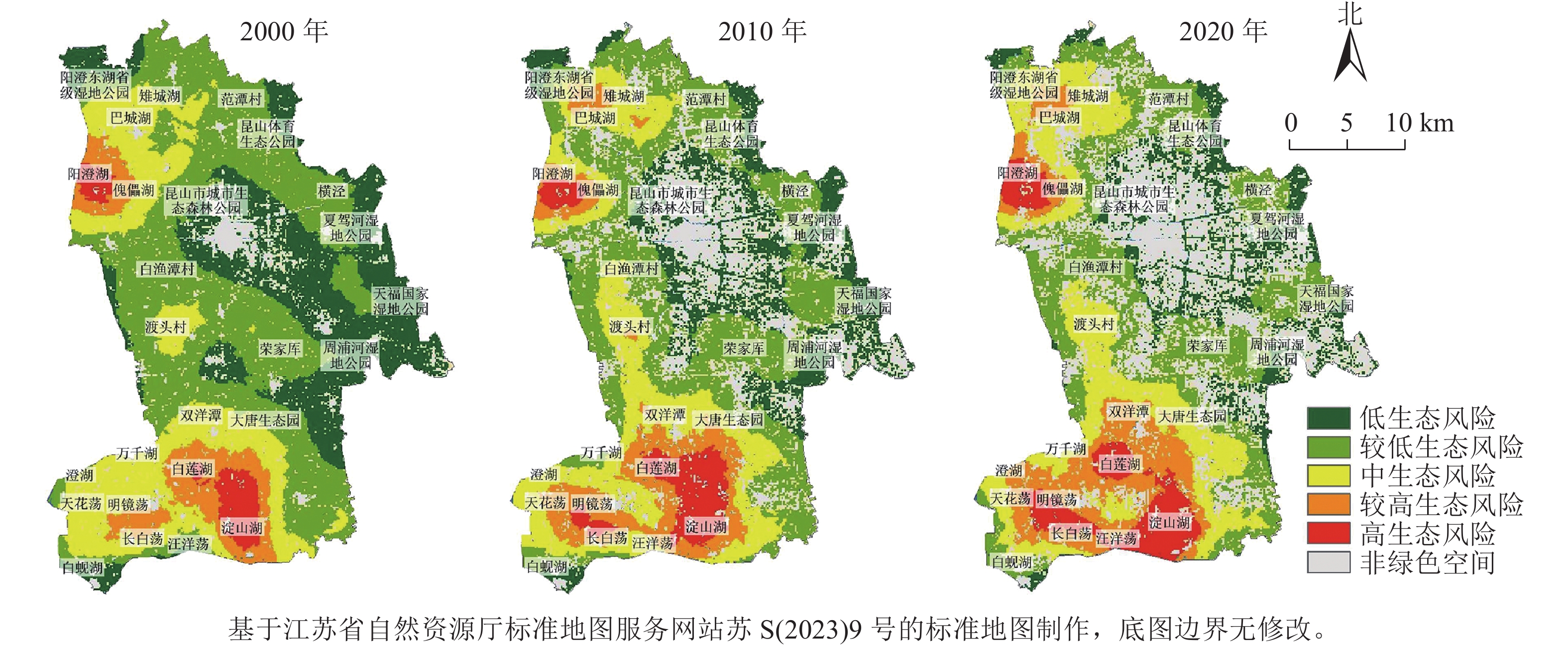 图 3 2000—2020年昆山市绿色空间景观生态风险空间分布示意图Figure 3 Spatial distribution of ecological risks in green space landscape of Kunshan City from 2000 to 2020表 5 2000—2020年昆山市绿色空间景观生态风险等级面积及比例Table 5 Area and proportion of landscape ecological risk level of green space in Kunshan City from 2000 to 2020
图 3 2000—2020年昆山市绿色空间景观生态风险空间分布示意图Figure 3 Spatial distribution of ecological risks in green space landscape of Kunshan City from 2000 to 2020表 5 2000—2020年昆山市绿色空间景观生态风险等级面积及比例Table 5 Area and proportion of landscape ecological risk level of green space in Kunshan City from 2000 to 2020年份 低风险区 较低风险区 中等风险区 较高风险区 高风险区 面积/hm2 比例/% 面积/hm2 比例/% 面积/hm2 比例/% 面积/hm2 比例/% 面积/hm2 比例/% 2000 1 116.90 25.66 1 830.87 42.06 918.09 21.09 345.15 7.93 141.48 3.25 2010 642.78 19.94 1 210.41 37.55 745.56 23.13 409.32 12.70 215.82 6.69 2020 452.70 15.93 961.02 33.81 692.64 24.37 455.58 16.03 280.26 9.86 2000—2020年昆山市绿色空间景观生态风险整体呈上升趋势,呈现“南北高,中间低”的空间分布特征。高、较高风险区面积明显上升,面积占比分别增加8.10%、6.61%,主要分布于淀山湖、白莲湖等湖荡密集地区,且不断向湖荡周围辐射扩张。该区域绿色空间类型以水域为主,由于围网养殖等人为活动对水域干扰程度加大,景观损失度逐年增加,使区域风险等级不断升高。中风险区面积小幅上升,面积占比增加3.28%,集中分布于渡头村、双洋潭等地区,并逐步沿较高风险区外围向四周扩散,区域内耕地、水域交错分布,受人为活动干扰较大,生态稳定性下降。较低、低风险区面积明显缩减,占比分别减少8.25%和9.73%,主要分布于研究区中部白渔潭村、荣家厍及北部范潭村、横泾等地区,且分布逐渐变得零散破碎,人为开发建设活动频繁,绿色空间不断减少,抗干扰能力减弱,景观生态风险值有增强趋势。
3.2.3 绿色空间景观生态风险等级空间变化
借助景观生态风险等级变化分布(图4)分析2000—2020年期间研究区各风险等级的变化情况。①风险等级升高区域的面积为21 503.12 hm2,占绿色空间总面积的36.69%,其中较低风险区域上升为中风险的区域面积最大,为6 413.09 hm2,其次为中风险区域上升为较高风险区域。主要分布在白莲湖、明镜荡、汪洋荡等地区,区域内湖荡、耕地镶嵌分布,城镇建设用地的扩张使生态斑块破碎化程度加剧,生态结构和功能受到损害,生态系统稳定性和恢复力下降。②风险等级基本不变区域的面积为31 026.25 hm2,占绿色空间总面积的52.61%,其中较低风险区域面积最大,为13 102.74 hm2。主要分布在白渔潭村、荣家厍、范潭村片区等。该区域生态环境相对较好,生态系统结构和整体格局较为完整,对外界干扰具备一定的抵御能力,可维持基本的生态功能。③风险等级降低区域的面积为5 241.88 hm2,占绿色空间总面积的10.70%,其中中风险区域下降为较低风险区域面积最大,为2 925.90 hm2,其次为较低风险区域下降为低风险区域。在空间上集中在大渔新村、朱家湾村、黄家埭等地区。区域内具有较好的生态基底,生态斑块间连续性较强且受经济建设活动干扰较小,生态系统稳定性提高,能够提供较好的生态服务效益。
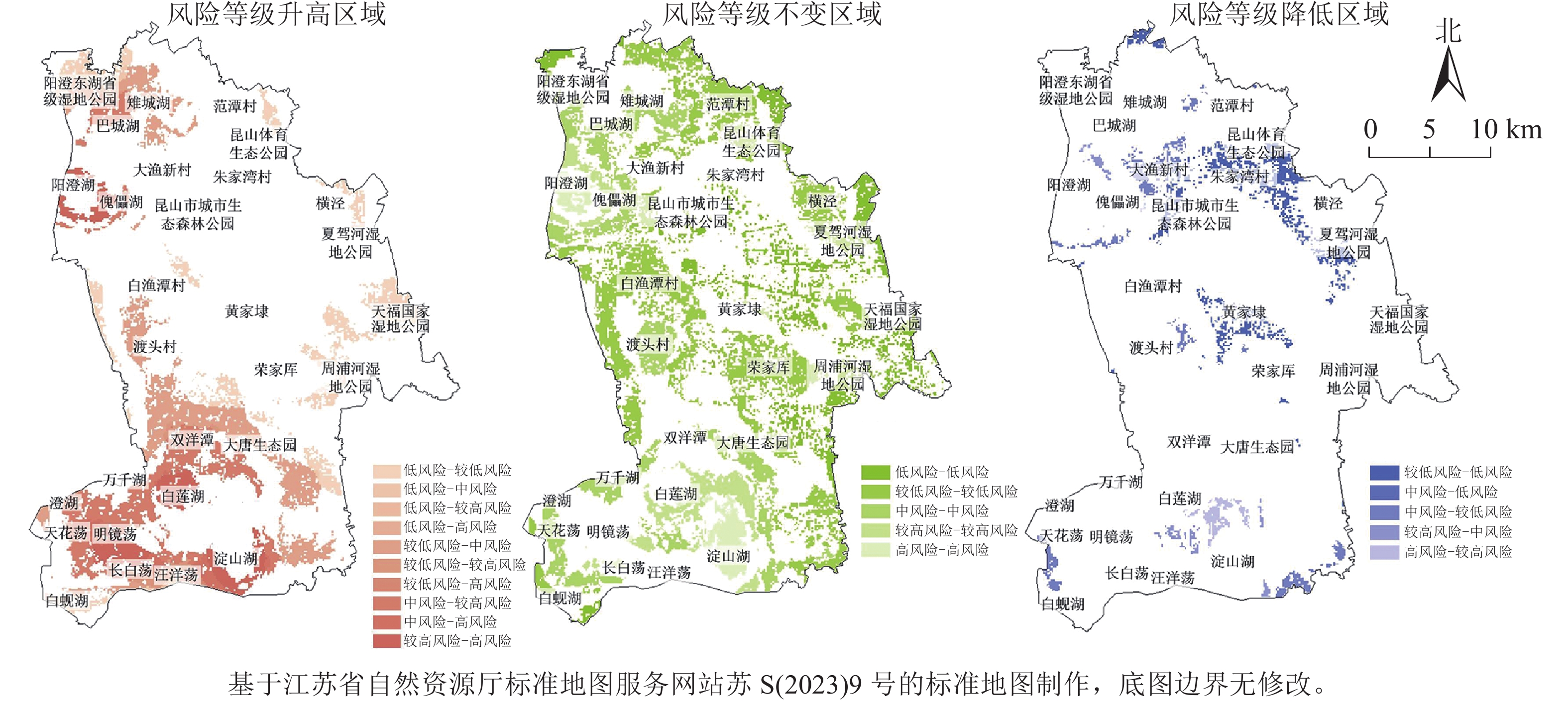 图 4 2000—2020年昆山市绿色空间景观生态风险等级变化示意图Figure 4 Change of landscape ecological risk level of green space in Kunshan City from 2000 to 2020
图 4 2000—2020年昆山市绿色空间景观生态风险等级变化示意图Figure 4 Change of landscape ecological risk level of green space in Kunshan City from 2000 to 20203.3 基于景观生态风险评价的昆山市绿色空间分区调控策略
基于2000—2020年昆山市绿色空间景观生态风险等级变化特征,将风险等级升高、不变和降低的区域分别划定为重点修复区、协调缓冲区和优化利用区。依据《苏州市“十四五”生态环境保护规划》《昆山市生态环境保护“十四五”规划》《昆山市国土空间总体规划(2021—2035)》等规划政策,结合调控分区的景观生态风险水平,提出有针对性的空间分区调控策略。
3.3.1 重点修复区实施生态保育,降低绿色空间生态风险
重点修复区为景观生态风险等级升高的区域,主要表现为较低风险向中风险、中风险向较高风险转移。片区内绿色空间破碎度增加,生态系统稳定性下降,景观生态风险水平不断上升。应加强生态保育与生态修复,对淀山湖、白莲湖等主要核心水域开展生态治理与修复工程,提升水域生态涵养功能;系统梳理、串通河网水系,在河网沿线严格管控开发强度大的建设活动;对破碎的绿色空间斑块进行整合,特别是南部长白荡、明镜荡等水域密集地区,着力提升水网景观的连通性和抗干扰能力,维护绿色空间的完整性与稳定性。
3.3.2 协调缓冲区加强缓冲区建设,筑牢绿色空间生态安全屏障
协调缓冲区为景观生态风险等级基本不变的区域,片区内绿色空间生态稳定性较强,能够抵御一定程度的外界干扰,景观生态风险维持在稳定水平。这些区域可作为生态缓冲地,提升绿色空间抗风险能力。通过强化河流水系、滨水绿带等生态廊道结构连通性[25],串联湖荡、农田大型生态斑块,构建水陆联动的网络化生态空间格局;加强傀儡湖、阳澄湖等生境敏感区的缓冲区建设,构建区域生态安全屏障,维护生态保护网络边界,增强区域景观生态风险缓冲能力。
3.3.3 优化利用区优化生态建设,发挥绿色空间生态效益
优化利用区为景观生态风险等级降低的区域,主要表现为中风险向较低风险、较低风险向低风险转换。片区内绿色空间生态系统结构较为完整,对外界干扰具有较强的适应能力,景观生态风险水平有所下降。应依托片区内良好的生态优势,适度优化建设,提升水网空间活力,维护生态系统的稳定性。首先明确生态保护红线边界,保护绿色空间健康稳定发展;其次对绿色空间进行分级分类管控,加强对城市生态森林公园、夏驾河湿地公园等核心生态资源的保护与管理,定期监测与评估生态用地的环境状况;同时在生态保护基础上优化建设,结合黄家埭等地区独特的水网空间优势开展科普教育、休闲游憩等服务,提升绿色空间的生态效益。
4. 讨论与结论
4.1 讨论
本研究表明:绿色空间用地类型转变与景观生态风险具有关联性。研究期间昆山市南部水域及周边地区由于城镇用地扩张,耕地、水域等绿色空间面积持续减少,生态系统结构稳定性下降,景观生态风险等级呈上升趋势。这与于淑会等[26]、陈斌等[27]的研究结论一致。水网地区以纵横交错的河流、湖荡为主体,水域面积较大,易受外界城镇建设用地扩张的干扰而破碎化,景观脆弱度高。本研究结果表明:水域范围内的景观生态风险指数普遍较高。这与何钊全等[28]对延安市的研究存在一定差异。延安市地处黄土丘陵区,林地和耕地是优势景观类型,受经济发展和建设用地扩张影响较大,林地、耕地破碎化程度加剧,抗干扰能力下降,景观损失度增加,使林地与耕地的景观生态风险值较高。
本研究在快速城镇化背景下,基于景观生态风险评价,加强绿色空间分区规划调控,对提升区域生态安全水平,优化国土空间结构,促进区域可持续发展具有一定理论指导意义。但研究仍存在一定局限性:①研究侧重从景观空间结构变化视角来评价绿色空间景观生态风险状况,对社会、经济等层面影响因素研究不足,还需进一步完善景观生态风险影响因素和驱动机制研究。②生态过程具有复杂性和抽象性,其具体演变过程很难做到定量表述。需要对生态风险展开多尺度分析,深入探讨景观格局生态风险和生态过程的耦合关系,为区域风险管理提供更加科学的依据。
4.2 结论
①2000—2020年昆山市绿色空间总面积持续减少,其中耕地面积缩减最多;水域面积先小幅增加后持续减少,总体呈减少趋势;林地、草地面积占比较小,维持相对平稳。研究区用地类型转换主要表现为耕地和水域转向建设用地。②2000—2020年昆山市绿色空间景观格局变化特征明显,耕地空间分布在建设用地扩张影响下趋于分散,破碎化程度加大,损失度增加;水域破碎度指数先下降后上升,总体破碎度呈增大趋势,景观受外界干扰增加;林地破碎度、干扰度和脆弱度呈下降趋势,斑块分布呈集聚态势;草地破碎度指数先上升后下降,总体破碎度呈下降趋势,空间分布趋于集聚,景观损失度降低。③2000—2020年昆山市绿色空间景观生态风险等级总体呈上升趋势,其中高风险区、较高风险区面积显著扩大,占比分别增加8.10%、6.61%,空间分布上主要集中在南部淀山湖、白莲湖等水域密集地区,并有进一步向外围蔓延发展的趋势;较低风险区、低风险区面积缩减明显,占比分别下降8.25%和9.73%;景观生态风险以低风险等级向更高一级转变为主,绿色空间受人工建设干扰生态风险不断增强。④依据景观生态风险等级变化特征将研究区划分为重点修复区、协调缓冲区和优化利用区。
-
图 1 高温胁迫下喷施不同浓度水杨酸对叶片叶绿素质量分数的影响
Figure 1 Chlorophyll content in response to different contents of SA under high temperature stress
图 2 高温胁迫下喷施不同浓度水杨酸对叶片丙二醛质量摩尔浓度的影响
Figure 2 MDA content in response to different contents of SA under high temperature stress
图 3 喷施不同浓度水杨酸在高温胁迫下菊花叶片脯氨酸、可溶性糖和可溶性蛋白质量分数的变化
Figure 3 roline, soluble sugar and soluble content in leaf in response to different contents of SA under high temperature stress
-
[1] 杨华庚, 陈慧娟. 高温胁迫对蝴蝶兰幼苗叶片形态和生理特性的影响[J]. 中国农学通报, 2009, 25(11): 123-127. YANG Huageng, CHEN Huijuan. Effect of high temperature stress on morphological and physiological characteristics in Phalaenopsis seedlings [J]. Chin Agric Sci Bull, 2009, 25(11): 123-127. [2] 李天来, 李淼, 孙周平. 钙和水杨酸对亚高温胁迫下番茄叶片保护酶活性的调控作用[J]. 应用生态学报, 2009, 20(3): 586-590. LI Tianlai, LI Miao, SUN Zhouping. Regulation effect of calcium and salicylic acid on defense enzyme activities in tomato leaves under sub-high temperature stress [J]. Chin J Appl Ecol, 2009, 20(3): 586-590. [3] 曹淑红, 李宁毅. 水杨酸对高温胁迫下百日草幼苗耐热性的影响[J]. 沈阳农业大学学报, 2014, 45(1): 91-94. CAO Shuhong, LI Ningyi. Effects of salicylic acid on heat resistance of Zinnia elegans seedlings under high temperature stress [J]. J Shenyang Agric Univ, 2014, 45(1): 91-94. [4] 贾思振, 房伟民, 陈发棣, 等. 夏菊耐热性指标筛选和综合评价[J]. 浙江林学院学报, 2009, 26(1): 52-57. JIA Sizhen, FANG Weimin, CHEN Fadi, et al. Heat tolerance for summer blooming of chrysanthemum [J]. J Zhejiang For Coll, 2009, 26(1): 52-57. [5] 贾思振, 房伟民, 陈发棣, 等. 高温下5个夏菊品种开花特性、叶片组织结构与光合特性的比较[J]. 南京农业大学学报, 2009, 32(3): 151-156. JIA Sizhen, FANG Weimin, CHEN Fadi, et al. Comparison of blooming traits, leaf anatomic structure and photosynthetic characteristics of five summer chrysanthemum varieties under high temperature [J]. J Nanjing Agric Univ, 2009, 32(3): 151-156. [6] 李合生. 植物生理生化实验原理和技术[M]. 北京: 高等教育出版社, 2001. [7] 水德聚, 石瑜, 曹亮亮, 等. 外源水杨酸预处理对高温胁迫下白菜耐热性和光合特性的影响[J]. 植物生理学报, 2012, 48(4): 386-392. SHUI Deju, SHI Yu, CAO Liangliang, et al. Effects of exogenous SA pretreatment on thermotolerance and photosynthesis in pakchoi under high temperature stress [J]. Plant Physiol J, 2012, 48(4): 386-392. [8] 杨岚, 师帅, 王红娟, 等. 水杨酸对高温胁迫下铁皮石斛幼苗耐热性的影响[J]. 西北植物学报, 2013, 33(3): 534-540. YANG Lan, SHI Shuai, WANG Hongjuan, et al. Effects of salicylic acid on heat-resistance of Dendrobium officinale seeding under high temperature stress [J]. Acta Bot Boreal-Occident Sin, 2013, 33(3): 534-540. [9] 孙军利, 赵宝龙, 郁松林. 外源水杨酸(SA)对高温胁迫下葡萄幼苗耐热性诱导研究[J]. 水土保持学报, 2014, 28(3): 290-299. SUN Junli, ZHAO Baolong, YU Songlin. Study of exogenous salicylic acid (SA) on the heat tolerance in grape seedlings under high temperature stress [J]. J Soil Water Conserv, 2014, 28(3): 290-299. [10] 叶陈亮, 柯玉琴, 陈伟. 大白菜耐热性的生理研究(Ⅲ)酶性和非酶性活性氧清除能力与耐热性[J]. 福建农业大学学报, 1996, 26(4): 498-501. YE Chenliang, KE Yuqin, CHEN Wei. A study on the physiology of heat tolerance in Chinese cabbage (Ⅲ) ability to scavenge active oxygen of emzyme and non-emzyme system and heat tolerance [J]. J Fujian Agric Univ, 1996, 26(4): 498-501. [11] 孟焕文, 张延峰, 程智慧, 等. 黄瓜幼苗对热胁迫的生理反应及耐热鉴定指标筛选[J]. 西北农业学报, 2000, 9(1): 96-99. MENG Huanwen, ZHANG Yanfeng, CHENG Zhihui, et al. The physiological reaction to hot stress and screening of hot tolerance index in cucumber [J]. Acta Agric Boreal-Occident Sin, 2000, 9(1): 96-99. [12] 叶凡, 侯喜林, 袁建玉. 高温胁迫对不结球白菜幼苗抗氧化酶活性和膜脂过氧化作用的影响[J]. 江苏农业学报, 2007, 23(2): 154-156. YE Fan, HOU Xilin, YUAN Jianyu. Effects of heat stress on antioxidant enzyme activity and lipid peroxidation of non-heading Chinese cabbage seedlings [J]. Jiangsu J Agric Sci, 2007, 23(2): 154-156. [13] 杜朝昆, 李忠光, 龚明. 水杨酸诱导的玉米幼苗适应高温和低温胁迫的能力与抗氧化酶系统的关系[J]. 植物生理学通讯, 2005, 41(1): 19-23. DU Chaokun, LI Zhongguang, GONG Ming. The adaptations to heat and chilling stresses and relation to antioxidant enzymes of maize seedlings induced by salicylic acid [J]. Plant Physiol Commun, 2005, 41(1): 19-23. [14] KUMARA G D K, XIA Yiping, ZHU Zhujun, et al. Effects of exogenous salicylic acid on antioxidative enzyme activities and physiological characteristics in gerbera (Gerbera jamesonii L.) grown under NaCl stress [J]. J Zhejiang Univ Agric & Life Sci, 2010, 36(6): 591-601. -

-
链接本文:
https://zlxb.zafu.edu.cn/article/doi/10.11833/j.issn.2095-0756.2016.03.011
 点击查看大图
点击查看大图
计量
- 文章访问数: 3306
- HTML全文浏览量: 503
- PDF下载量: 578
- 被引次数: 0
