





-
土壤是生态圈中的巨大碳库,其碳储量是陆地植被碳库和大气碳库的2~3倍[1];土壤呼吸是土壤碳的主要输出方式,对区域和全球尺度上碳收支的调配至关重要[2],其微小的变化都可能引起大气二氧化碳体积分数的较大变化[3],因此,土壤呼吸对生态系统结构和功能的影响都极其重要。土壤呼吸具有很强的时空异质性[4],受到生物和非生物因子的共同影响[5-10];不同的植被类型会形成群落结构和物种组成差异较大的生态系统,进而使得土壤呼吸受到生态系统生产力、碳分配格局、凋落物和群落小气候的影响而产生变异[11-16]。桉树Eucalyptus是中国华南地区广泛栽植的树种之一,栽植量仅次于杉木Cunninghamia lanceolata和马尾松Pinus massoniana;中国桉树人工林面积居世界第3位,仅次于巴西和印度[17]。桉树速生丰产,用途广泛,经济价值很高;与其他树种相比,桉树人工林碳汇功能更加明显,因而相关研究也颇具活力。目前,对桉树人工林土壤呼吸的研究主要集中在经营模式不同造成的土壤碳排放差异[18],林龄引起的碳转移差异[19-20]等方向,对不同种桉树人工林土壤呼吸及其影响要素的研究尚未见报道。本研究选取雷州半岛5个不同种桉树人工林和1个湿加松Pinus elliottii × caribaea林为研究对象,期望揭示不同种桉树人工林土壤呼吸速率的时空动态变化、土壤呼吸速率与影响要素的关系特征及土壤呼吸时空差异的主要影响要素等,为准确估算桉树人工林土壤碳收支状况提供数据支持。
-
研究地位于雷州半岛北部,南方国家级林木种苗示范基地境内(21°20′~21°30′N,109°22′~111°38′E)。该地区平均海拔为150.4 m,属于海洋性季风气候;土壤为玄武岩风化发育的砖红壤,土壤肥力中等;年平均气温为23.1 ℃,最热月(7月)平均气温为28.8 ℃,最冷月(1月)平均气温为15.6 ℃;年均降水量为1 567.0 mm,5-7月为雨季,多午后雷阵雨和台风带来的暴雨;年相对湿度为80%,年日照时数为1 937.0 h。2016年5-6月对尾叶桉E. urophylla(EU),粗皮桉E. pellita(EP),托里桉E. torelliana(ET),赤桉E. camaldulensis(EC),尾巨桉E. urophylla × grandis(EUG)和湿加松(PEC)6个林分开展野外调查,林地地势平坦,立地条件相似。样地概况见表 1。
表 1 样地基本概况
Table 1. Basic situation of sample plots
林分 林龄/a 平均树高/m 平均胸径/cm 林分密度/(株·hm-2) 坡度/(%) 坡向 海拔/m 林下主要物种 EU 15 25.38 ± 0.71 26.56 ± 0.81 524 0 平地 119.6 1, 2, 3, 5, 9, 10, 11 EP 10 16.80 ± 0.69 17.25 ± 0.71 970 0 平地 92.0 2:3:5:7:9 ET 10 18.00 ± 0.53 19.60 ± 0.56 810 0 平地 85.8 1, 2, 3, 4, 6, 7, 8, 12 EC 8 15.28 ± 0.64 10.82 ± 0.48 925 0 平地 98.6 3, 10, 11 EUG 8 23.15 ± 0.76 18.43 ± 0.24 690 12 南坡 113.0 1, 2, 3, 4, 5, 6, 7 PEC 15 13.73 ± 0.23 20.60 ± 0.62 599 0 平地 104.7 1, 2, 3, 4, 5, 6, 7, 12 说明:数据为平均值±标准误。物种编号:1.鹅掌柴Scheffera octophya, 2.龙船花Ixora chinensis, 3.盐肤木Rhus chinensis, 4.马樱丹Lantana carnara, 5.白背叶Mallotus apelta, 6.马唐草Digitaria sanguinalis, 7.白花鬼针草Herba bidentis, 8.胜红蓟Ageratum con-yzoides, 9.五节芒Miscanthus floridulus, 10.草芍药Paeonia obovata, 11.蟛蜞菊Wedelia chinensis, 12.荩草Arthraxon hispidus -
土壤呼吸速率测定时间为2016年3月至2017年2月。2016年1月,于6个林地中分别随机设置6个2 m × 2 m的小样方。在每个小样方的对角线交叉位置设置1个内径为20 cm,高12 cm的土壤环作为固定样点,将土壤环底端朝下垂直于地表插入土壤中,保留环顶部距离土壤表面3 cm。安装时应尽量做到一次到位,避免反复操作造成的土壤扰动;同时保证后续监测过程中土壤环位置恒定。安装2个月后,用LI-8100A土壤碳通量自动测定系统(LI-COR)连接20 cm呼吸气室,于每个月中下旬选择无降雨天气测定土壤呼吸速率。开始测定前需检查土壤环安放是否正常,并剪除土壤环内可见植物。测定频率为1次·月-1·林分-1,测定时间为从8:00至18:00,测定间隔为1 h,3次·重复-1。利用仪器自身配置的土壤温度和水分传感器探针同步测定表层土壤(10 cm)温度和土壤湿度(0~10 cm平均体积含水率),以下均称为土壤温度、土壤湿度。
-
于2016年7月,在各林地土壤呼吸样方附近分别再设置6个1 m × 1 m凋落物小样方。剪除小样方内植物活体后收集地表全部凋落物,混合均匀并标记;除去动物粪便、石砾和昆虫等杂物,用牛皮纸包裹,在65 ℃烘箱中烘干直至质量不变;机械磨碎过筛,待测。在每个凋落物样方下挖取土壤剖面,划分土壤深度0~10,10~20,20~40和40~60 cm等4层[21]进行单独取样。环刀(100 cm3)法采集自然状态下土壤样品,测定土壤容重、孔隙度和持水量等性质。同时每层另取约300 g土壤样品去除根、石头等杂物后按质量比混合均匀,装袋标记后带回实验室,风干,粉碎过100目筛。用重铬酸钾氧化-容量法测定土壤和凋落物有机碳质量。同期采用Hemiview冠层分析系统在各个凋落物样方处测定叶面积指数(LAI),3次·重复-1,测定均在晴天无风的清晨、傍晚或阴天进行,以减少散射造成的误差。
-
土壤呼吸速率与土壤温度的关系采用指数经验模型描述[22]:$f\left( R \right) = a{{\rm{e}}^{bT}} $。土壤呼吸速率与土壤湿度的关系采用二次项模型描述[23]:$ f\left( R \right) = a{h_{\rm{r}}}^2 + b{h_{\rm{r}}} + c$。土壤呼吸速率与土壤温、湿度双变量模型采用指数符合模型描述[23]:$ f\left( R \right) = a{{\rm{e}}^{bT}}{h_{\rm{r}}}^c$。各式中:f(R)为土壤呼吸速率(μmol·m-2·s-1);T为表层土壤温度(℃);hr为土壤湿度(%);a,b,c为方程拟合参数。
温度敏感性指数Q10采用指数模型描述[24-25]:Q10=e10b。
土壤呼吸速率与土壤容重、孔隙度、有机碳密度、凋落物有机碳储量和叶面积指数的相关性分析根据各桉树林分2016年7月实测数据进行处理,用于确定引起桉树林分土壤呼吸空间变异的主要因素。
观测期间各林分土壤表面碳排放累积通量(g·m-2·a-1)估算方法:以1 h为步长,通过温度单变量和温度、湿度双变量指数模型与连续动态监测的土壤温度、湿度数据建立方程,估计土壤呼吸连续数据累加获得。
数据分析利用统计软件SPSS 19.0。图或表均由Excel 2007处理获得。
-
在观测期内,6个林分土壤呼吸速率的月变化特征明显,均呈现单峰曲线趋势(图 1)。各月之间平均土壤呼吸速率均存在显著性差异(P<0.001)。不同林分土壤呼吸速率随月份逐渐上升,在2016年5-7月出现最高值;之后逐渐下降,在2016年12月至翌年2月出现最低值。由LSD多重比较可知,观测期内6个林分土壤呼吸速率均值EU为(3.46 ± 0.41)μmol·m-2·s-1,EP为(2.15 ± 0.18)μmol·m-2·s-1,ET为(3.72 ± 0.20)μmol·m-2·s-1,EC为(2.14 ± 0.15)μmol·m-2·s-1,EUG为(2.34 ± 0.21)μmol·m-2·s-1,PEC为(2.33 ± 0.17)μmol·m-2·s-1;EU和ET之间不存在显著差异(P>0.05),但两者均显著大于其他林分(P<0.05),EP,EC,EUG和PEC之间无显著差异。

图 1 不同林分土壤呼吸速率月动态变化
Figure 1. Monthy variation of soil respiration rate in different stands
-
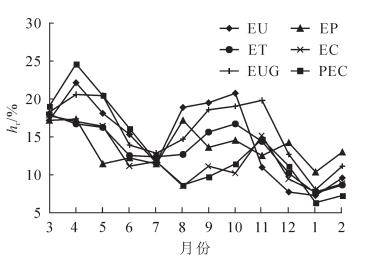
6个林分土壤温度月变化表现为单峰曲线趋势(图 2),12个月份间平均土壤温度具有极显著差异(P<0.01)。观测期初期,土壤的温度不断升高,各林分土壤温度在6月达到最大值。2016年7月至翌年2月,土壤温度逐渐降低,各林分土壤温度在翌年2月达到最小值。观测期内各林分平均土壤温度分别为EU为(27.11 ± 0.37)℃,EP为(27.33 ± 0.40)℃,ET为(26.54 ± 0.38)℃,EC为(27.10 ± 0.43)℃,EUG为(26.61 ± 0.40)℃,PEC为(26.46 ± 0.42)℃,均无显著差异(P>0.05)。各林分土壤湿度月变化表现为双峰曲线趋势(图 3)。观测初期先升高后降低,在5-7月达到谷值,之后出现回升趋势。观测期内平均土壤湿度分别为EU为14.93% ± 0.45%,EP为14.46% ± 0.29%,ET为13.22% ± 0.31%,EC为11.65% ± 0.30%,EUG为15.13% ± 0.43%,PEC为12.80% ± 0.50%。EU,EP和EUG三者间差异不显著,但均显著高于其他林分(P<0.05);ET和PEC差异显著(P<0.01),且均显著高于EC(P<0.05)。

图 2 不同林分土壤温度月动态
Figure 2. Monthy variation of soil temperature in different stands

图 3 不同林分土壤湿度月动态
Figure 3. Monthy variation of soil moisture in different stands
-
由表 2可知:土壤容重仅在表层(0~10 cm)出现差异,其中EU显著大于其他林分(P<0.05);ET和PEC无显著差异,但均显著大于EP(P<0.05);EP,EC和EUG之间无显著差异。土壤总孔隙度、毛管孔隙度和非毛管孔隙度均为表层土壤不同林分间差异最大,深层土壤差异较小。土壤有机碳质量分数除10~20 cm土层外其他均存在不同程度的差异性,且均表现为ET最大,EU最小。
表 2 不同林分土壤容重、孔隙度及有机碳质量分数
Table 2. Comparison of soil bulk density and soil organic carbon content in different stands
林分 容重/(g·cm-3) 总孔隙度/% 毛管孔隙度/% 非毛管孔隙度/% w有机碳/(g·kg-1) 0~10 10~20 20~40 40~60 0~10 10~20 20~40 40~60 0~10 10~20 20~40 40~60 0~10 10~20 20~40 40~60 0~10 10~20 20~40 40~60 EU 0.84 ± 0.03 c 0.98 ± 0.02 a 0.99 ± 0.06 a 1.08 ± 0.06 a 62.38 ± 1.74 a 57.42 ± 1.29 a 57.34 ± 1.82 a 56.50 ± 21.63 ab 50.28 ± 0.68 bc 50.33 ± 1.42 a 49.01 ± 1.14 a 52.20 ± 0.99 a 5.01 ± 0.99 c 7.88 ± 2.58 a 6.02 ± 1.45 a 2.72 ± 0.15 a 22.34 ± 1.08 c 18.22 ± 1.10 a 11.36 ± 1.21 c 7.82 ± 0.76 c EP 1.07 ± 0.03 a 1.00 ± 0.07 a 1.04 ± 0.04 a 1.01 ± 0.02 a 55.29 ± 1.62 c 58.21 ± 3.39 a 55.03 ± 1.63 a 54.91 ± 0.96 ab 52.30 ± 0.87 ab 48.65 ± 1.09 a 49.86 ± 1.06 a 51.50 ± 1.79 a 10.08 ± 1.01 ab 8.77 ± 1.51 a 7.48 ± 2.54 a 5.02 ± 2.60 a 28.49 ± 1.79 bc 22.10 ± 3.00 a 15.34 ± 3.35 ab 15.34 ± 3.35 ab ET 0.89 ± 0.02 bc 0.99 ± 0.07 a 1.05 ± 0.02 a 1.01 ± 0.02 a 60.24 ± 1.10 ab 56.04 ± 1.94 a 57.12 ± 0.82 a 59.07 ± 1.15 a 50.69 ± 0.73 b 48.93 ± 1.35 a 49.54 ± 1.35 a 50.54 ± 1.30 a 6.70 ± 0.37 bc 6.76 ± 1.04 a 4.90 ± 1.37 a 3.75 ± 1.40 a 37.43 ± 3.01 a 37.43 ± 3.01 a 20.86 ± 1.28 a 20.31 ± 0.78 a 17.23 ± 2.04 a EC 0.97 ± 0.01 b 0.99 ± 0.02 a 1.04 ± 0.03 a 1.07 ± 0.01 a 59.94 ± 0.93 ab 57.98 ± 3.08 a 55.45 ± 0.32 a 57.56 ± 1.27 ab 49.52 ± 1.52 bc 47.98 ± 1.39 a 52.12 ± 1.10 a 53.31 ± 3.76 a 10.41 ± 1.74 ab 9.99 ± 2.51 a 3.34 ± 0.78 a 4.26 ± 2.49 a 27.70 ± 0.87 bc 27.70 ± 0.87 bc 22.52 ± 0.56 a 15.21 ± 1.92 abc 11.06 ± 1.14 abc EUG 0.92 ± 0.04 bc 1.02 ± 0.04 a 1.07 ± 0.03 a 1.03 ± 0.08 a 58.96 ± 1.47 abc 55.53 ± 0.30 a 55.56 ± 0.78 a 56.62 ± 1.78 ab 47.68 ± 0.16 c 47.45 ± 0.80 a 49.14 ± 1.25 a 53.56 ± 3.05 a 11.28 ± 1.50 a 8.08 ± 0.81 a 6.42 ± 1.39 a 3.07 ± 2.23 a 33.09 ± 3.77 ab 33.09 ± 3.77 ab 18.37 ± 13.72 ± 1.33 bc 10.22 ± 1.97 bc PEC 0.95 ± 0.03 b 1.07 ± 0.04 a 1.04 ± 0.01 a 0.94 ± 0.02 a 57.39 ± 0.73 bc 55.69 ± 1.01 a 54.44 ± 0.36 a 54.30 ± 1.07 b 53.44 ± 0.49 a 49.89 ± 0.87 a 50.11 ± 0.78 a 52.76 ± 1.86 a 6.80 ± 1.22 bc 6.14 ± 2.20 a 7.01 ± 0.69 a 6.31 ± 0.77 a 26.68 ± 2.02 bc 26.68 ± 2.02 bc 20.77 ± 1.85 a 14.56 ± 2.15 abc 9.70 ± 2.03 bc 说明:数值为平均值±标准误。同列不同小写字母表示不同林分间差异显著(P < 0.05) -
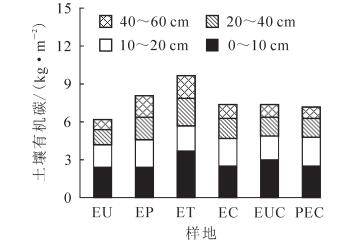
6个林分总有机碳密度(图 4)依次为EU 6.16 kg·m-2,EP 8.08 kg·m-2,ET 9.65 kg·m-2,EC 7.38 kg·m-2,EUG 7.37 kg·m-2,PEC 7.16 kg·m-2;ET与EP无显著差异,但显著高于其他4个林分(P<0.05)。表层土壤有机碳密度ET林分显著高于其他林分(P<0.05);10~20 cm土层有机碳密度6个林分间均无显著差异;20~40 cm土层ET显著高于EU(P<0.05),但两者与其他林分均无显著差异;40~60 cm土层ET与EP无显著差异,但ET显著高于其他林分(P<0.05),EP与EC,EUG无显著差异,但EP显著高于EU和PEC(P<0.05)。

图 4 不同林分土壤有机碳密度比较
Figure 4. Comparison of soil organic carbon in different stands
-
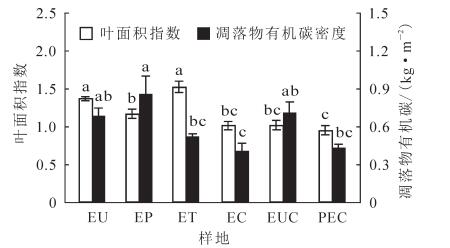
由图 5可知:6个林分叶面积指数为0.95~1.37,EU和ET差异不显著,但均显著高于其他林分(P<0.05);EP显著高于PEC(P<0.05),其他差异不显著。不同桉树林凋落物有机碳密度为0.40~0.85 kg·m-2,PEC为0.43 kg·m-2,EU,EP和EUG三者间无显著差异,但均显著高于EC(P<0.05);EP显著高于ET和PEC(P<0.05),ET与PEC差异不显著。

图 5 不同林分叶面积指数及凋落物有机碳储量
Figure 5. Comparison of leaf area index (LAI) and litter organic carbon storage in different stands
-
Person相关性分析表明(表 3),在观测期内各林分土壤呼吸速率与土壤温度均为极显著正相关(P<0.01);与土壤湿度均为负相关关系,除EC为显著外其他林分均为极显著。各林分一致表现出土壤呼吸速率与土壤温度的相关性高于他与土壤湿度的相关性,说明土壤温度对土壤呼吸速率的影响比湿度更大。土壤呼吸速率与土壤温度、湿度的指数关系、二次多项式关系均达到极显著水平(P<0.001)。土壤温度、湿度双因子模型在EU,EP,EC和EUG林分中拟合度较单一因子高,说明土壤呼吸速率受到土壤温度、湿度的综合作用。决定系数R2越大表明模型的拟合程度越好,其百分数可以用来解释影响因子对土壤呼吸速率的变异程度。由表 4可知:土壤温度解释不同种桉树林分土壤呼吸速率43.7%~75.8%的变异,均低于PEC(79.8%)。土壤湿度解释不同种桉树林分土壤呼吸速率18.3%~50.9%的变异,而PEC为41.2%。土壤温度、湿度双因子对桉树林分土壤呼吸变异的解释能力为44.8%~83.9%,对照PEC为78.7%。桉树林分土壤呼吸的温度敏感性Q10值为1.42~1.99,PEC为1.60。
表 3 土壤呼吸速率与土壤温度、湿度的关系
Table 3. Relationship between soil respiration rate and soil temperature and moisture
林分 $f\left( R \right) = a{{\rm{e}}^{bT}} $ $f\left( R \right) = a{h_{\rm{r}}}^2 + b{h_{\rm{r}}} + c $ $f\left( R \right) = a{{\rm{e}}^{bT}}{h_{\rm{r}}}^c $ Q10 a b R2 r a b c R2 r a b c R2 EU 0.487 0.069 0.437 0.628** -0.030 1.019 -4.251 0.443 0.507** 0.421 0.042 0.358 0.448 1.99 EP 0.488 0.053 0.630 0.817** -0.010 0.375 -1.069 0.378 0.568** 0.294 0.044 0.288 0.751 1.70 ET 1.442 0.035 0.638 0.781** -0.022 0.678 -1.009 0.387 0.432** 1.269 0.029 0.114 0.614 1.42 EC 0.635 0.044 0.758 0.851** -0.018 0.491 -0.923 0.183 0.173* 0.407 0.042 0.201 0.761 1.55 EUG 0.381 0.066 0.754 0.893** -0.014 0.480 -1.441 0.509 0.531** 0.270 0.054 0.256 0.839 1.93 PEC 0.631 0.047 0.798 0.884** -0.013 0.418 -0.769 0.412 0.315** 0.569 0.045 0.067 0.787 1.60 说明:R2为方程拟合优度, 即决定系数; r为土壤呼吸与土壤温度、湿度经Pearson相关性分析所得相关系数; *表示P<0.05;**表示P<0.01 表 4 土壤呼吸速率与土壤性质、凋落物及叶面积指数的相关性
Table 4. Correlation between soil respiration rate and soil properties, litter organic carbon storage and leaf area index (LAI)
土层/cm 相关性分析 容重 总孔隙度 毛管孔隙度 非毛管孔隙度 有机碳密度 凋落物有机碳密度 叶面积指数 土壤温度 土壤湿度 0-10 -0.752** 0.622** 0.024 0.611** 0.434 -0.109 0.681** -0.054 0.211 10~20 -0.149 0.165 0.434 -0.105 0.327 20-40 0.161 -0.360 -0.161 -0.149 0.174 40-60 0.164 -0.453 -0.071 -0.304 0.133 说明: **表示P<0.01, *表示P<0.05 -
对5个桉树林分土壤呼吸速率与土壤性质、凋落物有机碳密度、叶面积指数和土壤温度、湿度的相关性分析表明,土壤呼吸速率与表层土壤容重、总孔隙度和非毛管孔隙度呈极显著相关(P<0.01),说明土壤呼吸速率与表层土壤物理性质关系密切,与土壤有机碳质量分数、凋落物有机碳质量分数均无显著相关性,与叶面积指数呈极显著正相关(P<0.01),且随叶面积指数呈线性增加趋势,模型拟合度达到R2=0.468,与土壤温度、湿度相关性均不显著。
-
各林分土壤表面碳排放通量:EU为(1 333.42 ± 8.19)g·m-2·a-1,EP为(804.18 ± 3.82)g·m-2·a-1,ET为(1 408.94 ± 5.23)g·m-2·a-1,EC为(804.98 ± 4.55)g·m-2·a-1,EUG为(893.31 ± 5.97)g·m-2·a-1和PEC为(891.15 ± 4.95)g·m-2·a-1。其中EU和ET无显著差异,但均显著高于其他林分(P<0.05)。相关性分析表明:各林分7月土壤表面碳排放通量与表层土壤容重存在极显著负相关关系(P<0.01),与表层土壤总孔隙度和非毛管孔隙度存在极显著正相关关系(P<0.01),与表层土壤有机碳密度呈显著正相关关系(P<0.05),相关系数分别为0.743,0.693,0.644和0.484。
-
土壤二氧化碳主要来源于微生物分解土壤有机质和植物根系呼吸,土壤温度会影响土壤微生物活性和植物呼吸酶的活性,进而影响土壤呼吸速率[25-26];而土壤微生物和植物根系的生命活动直接受到土壤含水率的调控[27],因而土壤含水率对土壤呼吸的影响也是十分重要的。本研究中6个林分的土壤呼吸速率月变化一致,均表现为单峰曲线格局,与土壤温度月变化符合极显著的指数关系,与土壤体积含水率月变化符合极显著的二次多项式关系(P<0.001)。6个林分土壤呼吸最大值出现在5-7月,比其他亚热带林分[17, 23, 28-29]的7-8月要早,可能是由于研究区土壤温度从5月即开始接近年均最大值,升温早于其他地区,导致土壤呼吸变化提前;最小值出现在土壤温度、湿度均接近年均最小值的12月和翌年1-2月,与其他亚热带林分相似。
Q10可以反映土壤呼吸对土壤温度的敏感程度。陈光水等[30]的研究表明:中国森林土壤呼吸Q10值为1.33~5.53,平均值为2.65。本研究结果为1.42~1.99,与之相符合,但处于偏低水平。吴蒙[28]对桂林尧山桉树的研究发现:20年生桉树Q10值为2.53,4年生桉树为2.41,均比本研究结果高;原因可能是研究区年均温度较高,土壤呼吸温度敏感性与土壤温度存在显著负相关关系[31],导致土壤呼吸温度敏感性低于其他地区。
-
土壤呼吸作用的物质基础源于光合作用,植物根系呼吸作用要依赖于植物地上部分光合产物的分配;研究发现土壤呼吸速率与植被净初级生产力存在显著正相关关系[32],叶面积的大小及分布直接影响着林分对光能的截获和利用,进而影响林分生产力[33]。本研究发现:5个桉树林分的土壤呼吸速率随叶面积指数的增大呈线性增加趋势,与BOND-LAMBERTY等[34]研究结果相同,认为两者之间存在极显著正相关关系。REICHSTEIN等[35]也证实了标准条件下的土壤呼吸速率与叶面积指数之间也存在很强的相关性。
土壤孔隙度的大小与土壤通透性有关,会影响土壤有机质的分解速率和规模,进而影响土壤呼吸速率[36],是指示土壤结构优劣的一个重要指标。土壤容重反映了土壤呼吸排放通道的顺畅程度[37]。本研究中5个桉树林分的土壤呼吸速率与土壤表层(0~10 cm)容重呈极显著负相关,这是因为土壤是多孔系统,植物根系和土壤微生物呼吸释放的二氧化碳多聚集在这些空隙中,容重越大,孔隙度会相应减小,进而阻碍了气体扩散的物理学进程[38]。本研究中,土壤呼吸速率与土壤总孔隙度呈极显著正相关关系,与宋启亮等[39]、王建国[40]的研究结果相似。土壤呼吸速率与土壤非毛管孔隙度呈极显著正相关,推测原因是非毛管孔隙具有通气透水性,有利于土壤微生物的呼吸作用,从而有助于土壤有机质的分解。
异氧呼吸在土壤呼吸中所占的比例范围因地域不同变异较大,热带和温带森林为30%~83%,寒冷地区为7%~50%[41]。土壤有机碳作为异氧呼吸的物质基础,其对土壤呼吸的影响不容忽视。本研究中5个桉树林分土壤呼吸速率与土壤有机碳质量分数呈正相关性,与王国兵等[42]研究结果相似。土壤表面碳排放通量与土壤表层有机碳密度呈显著正相关,与韩营营等[43]对白桦Betula platyphylla天然次生林的研究结果一致;推测其原因可能是研究区位于热带地区,表层土壤温度较高,适宜土壤微生物的代谢活动,且表层土壤有机碳密度较高,微生物异氧呼吸有更多的物质来源,因而促进了土壤二氧化碳的释放[44-46]。
本研究中所有林分土壤呼吸速率年均值为2.14~3.72 μmol·m-2·s-1,比华南地区杉木(2.13 μmol·m-2·s-1)[47]和马尾松人工林(2.07 μmol·m-2·s-1)要高;除EU(3.46 μmol·m-2·s-1)和ET(3.72 μmol·m-2·s-1)外,其他4个林分均低于雷蕾等[48]、梁国华等[49]对马尾松人工林的研究结果亦低于中国森林土壤呼吸年通量平均值(约2.58 μmol·m-2·s-1)[30]。由于EU和ET林分具较高的叶面积指数,叶面积指数表征林分生产力,因此光合作用强度高于其他4个林分,根呼吸强度亦高于其他4个林分,故叶面积指数可能是EU和ET林分土壤呼吸高于其他林分的主导因素。研究发现:5个桉树林分土壤呼吸速率与表层土壤容重、总孔隙度及非毛管孔隙度与土壤呼吸均存在极显著相关关系,故可认为土壤呼吸速率空间异质性受到多因素综合影响。
Temporal and spatial dynamics of soil respiration and influencing factors in Eucalyptus plantations
-
摘要: 为研究不同种桉树Eucalyptus人工林土壤呼吸速率时空变异特征及其影响要素,估算桉树人工林土壤碳排放通量,测定2016年3月-2017年2月时段内5个不同种桉树林及1个湿加松Pinus elliottii×caribaea林土壤呼吸速率,分析桉树人工林土壤呼吸速率时空变化及其与影响要素的相关关系。结果表明:6个林分土壤呼吸速率时间变化明显,均呈单峰曲线格局;土壤呼吸速率与表层土壤温度符合指数模型,与平均体积含水率符合二次多项式模型(P < 0.001),土壤呼吸速率时间变化受土壤温度和体积含水率共同驱动,温、湿度双因素模型可以解释土壤呼吸速率44.8%~83.9%的变异。土壤呼吸速率的空间变异主要受表层土壤容重、叶面积指数、总孔隙度和非毛管孔隙度的影响,相关性均为极显著(P < 0.01);土壤表面二氧化碳累积通量还受到土壤表层有机碳密度影响,相关性显著(P < 0.05)。尾叶桉E. urophylla林和托里桉E. torelliana林的土壤呼吸速率年均值及土壤表面碳排放年累积通量均显著大于其他林分(P < 0.05),两者之间差异不显著。Abstract: To contribute to a further understanding of soil respiration dynamics and to help to provide an accurate calculation of carbon emissions, the relationship between soil respiration dynamics and influencing factors for six plantation types:Eucalyptus urophylla, E. pellita, E. torelliana, E. camaldulensis, E. urophylla×E. grandis and Pinus elliottii×P. caribaea, along the Leizhou Peninsula was field continuously using the LI-8100A (LI-COR, USA) for spatial and temporal variability in the soil respiration rate from March 2016 to February 2017. With this data, analyses (including correlation analyses) of spatial and temporal variation and analyses of the relative importance of key factors influencing these parameters were undertaken. Results showed that soil respiration rates for the six plantations studied, in relation to soil temperature, all had a single peak in their pattern with both index and quadratic models able to simulate relationships between soil respiration and soil temperature or moisture (0-10 cm). Temporal variation of the soil respiration rate was driven by both soil temperature and moisture with a two-factor equation (soil temperature and moisture at 10 cm) explaining 44.8% to 83.9% of the variation in soil respiration. Spatial variation of the soil respiration rate was mainly affected by soil bulk density, leaf area index, total soil porosity, and non-capillary soil porosity, with correlations between these parameters and soil respiration rates highly significant (P < 0.01). Soil surface carbon flux was also correlated to soil organic carbon density (P < 0.05) except for soil bulk density, total soil porosity and non-capillary soil porosity. The annual average of soil respiration and soil surface carbon flux in both E. urophylla and E. torelliana plantations were all significantly higher than E. pellita, E. camaldulensis, E. urophylla×E. grandis plantations and P. elliottii×P. caribaea plantation (P < 0.05), and with no significant differences between E. urophylla and E. torelliana plantations. In total, the spatial and temporal heterogeneity of soil respiration was influenced by various factors, which is the most important consideration in estimating soil respiration flux.
-
图 1 不同林分土壤呼吸速率月动态变化
Figure 1 Monthy variation of soil respiration rate in different stands
图 5 不同林分叶面积指数及凋落物有机碳储量
Figure 5 Comparison of leaf area index (LAI) and litter organic carbon storage in different stands
表 1 样地基本概况
Table 1. Basic situation of sample plots
林分 林龄/a 平均树高/m 平均胸径/cm 林分密度/(株·hm-2) 坡度/(%) 坡向 海拔/m 林下主要物种 EU 15 25.38 ± 0.71 26.56 ± 0.81 524 0 平地 119.6 1, 2, 3, 5, 9, 10, 11 EP 10 16.80 ± 0.69 17.25 ± 0.71 970 0 平地 92.0 2:3:5:7:9 ET 10 18.00 ± 0.53 19.60 ± 0.56 810 0 平地 85.8 1, 2, 3, 4, 6, 7, 8, 12 EC 8 15.28 ± 0.64 10.82 ± 0.48 925 0 平地 98.6 3, 10, 11 EUG 8 23.15 ± 0.76 18.43 ± 0.24 690 12 南坡 113.0 1, 2, 3, 4, 5, 6, 7 PEC 15 13.73 ± 0.23 20.60 ± 0.62 599 0 平地 104.7 1, 2, 3, 4, 5, 6, 7, 12 说明:数据为平均值±标准误。物种编号:1.鹅掌柴Scheffera octophya, 2.龙船花Ixora chinensis, 3.盐肤木Rhus chinensis, 4.马樱丹Lantana carnara, 5.白背叶Mallotus apelta, 6.马唐草Digitaria sanguinalis, 7.白花鬼针草Herba bidentis, 8.胜红蓟Ageratum con-yzoides, 9.五节芒Miscanthus floridulus, 10.草芍药Paeonia obovata, 11.蟛蜞菊Wedelia chinensis, 12.荩草Arthraxon hispidus  下载: 导出CSV
下载: 导出CSV
表 2 不同林分土壤容重、孔隙度及有机碳质量分数
Table 2. Comparison of soil bulk density and soil organic carbon content in different stands
林分 容重/(g·cm-3) 总孔隙度/% 毛管孔隙度/% 非毛管孔隙度/% w有机碳/(g·kg-1) 0~10 10~20 20~40 40~60 0~10 10~20 20~40 40~60 0~10 10~20 20~40 40~60 0~10 10~20 20~40 40~60 0~10 10~20 20~40 40~60 EU 0.84 ± 0.03 c 0.98 ± 0.02 a 0.99 ± 0.06 a 1.08 ± 0.06 a 62.38 ± 1.74 a 57.42 ± 1.29 a 57.34 ± 1.82 a 56.50 ± 21.63 ab 50.28 ± 0.68 bc 50.33 ± 1.42 a 49.01 ± 1.14 a 52.20 ± 0.99 a 5.01 ± 0.99 c 7.88 ± 2.58 a 6.02 ± 1.45 a 2.72 ± 0.15 a 22.34 ± 1.08 c 18.22 ± 1.10 a 11.36 ± 1.21 c 7.82 ± 0.76 c EP 1.07 ± 0.03 a 1.00 ± 0.07 a 1.04 ± 0.04 a 1.01 ± 0.02 a 55.29 ± 1.62 c 58.21 ± 3.39 a 55.03 ± 1.63 a 54.91 ± 0.96 ab 52.30 ± 0.87 ab 48.65 ± 1.09 a 49.86 ± 1.06 a 51.50 ± 1.79 a 10.08 ± 1.01 ab 8.77 ± 1.51 a 7.48 ± 2.54 a 5.02 ± 2.60 a 28.49 ± 1.79 bc 22.10 ± 3.00 a 15.34 ± 3.35 ab 15.34 ± 3.35 ab ET 0.89 ± 0.02 bc 0.99 ± 0.07 a 1.05 ± 0.02 a 1.01 ± 0.02 a 60.24 ± 1.10 ab 56.04 ± 1.94 a 57.12 ± 0.82 a 59.07 ± 1.15 a 50.69 ± 0.73 b 48.93 ± 1.35 a 49.54 ± 1.35 a 50.54 ± 1.30 a 6.70 ± 0.37 bc 6.76 ± 1.04 a 4.90 ± 1.37 a 3.75 ± 1.40 a 37.43 ± 3.01 a 37.43 ± 3.01 a 20.86 ± 1.28 a 20.31 ± 0.78 a 17.23 ± 2.04 a EC 0.97 ± 0.01 b 0.99 ± 0.02 a 1.04 ± 0.03 a 1.07 ± 0.01 a 59.94 ± 0.93 ab 57.98 ± 3.08 a 55.45 ± 0.32 a 57.56 ± 1.27 ab 49.52 ± 1.52 bc 47.98 ± 1.39 a 52.12 ± 1.10 a 53.31 ± 3.76 a 10.41 ± 1.74 ab 9.99 ± 2.51 a 3.34 ± 0.78 a 4.26 ± 2.49 a 27.70 ± 0.87 bc 27.70 ± 0.87 bc 22.52 ± 0.56 a 15.21 ± 1.92 abc 11.06 ± 1.14 abc EUG 0.92 ± 0.04 bc 1.02 ± 0.04 a 1.07 ± 0.03 a 1.03 ± 0.08 a 58.96 ± 1.47 abc 55.53 ± 0.30 a 55.56 ± 0.78 a 56.62 ± 1.78 ab 47.68 ± 0.16 c 47.45 ± 0.80 a 49.14 ± 1.25 a 53.56 ± 3.05 a 11.28 ± 1.50 a 8.08 ± 0.81 a 6.42 ± 1.39 a 3.07 ± 2.23 a 33.09 ± 3.77 ab 33.09 ± 3.77 ab 18.37 ± 13.72 ± 1.33 bc 10.22 ± 1.97 bc PEC 0.95 ± 0.03 b 1.07 ± 0.04 a 1.04 ± 0.01 a 0.94 ± 0.02 a 57.39 ± 0.73 bc 55.69 ± 1.01 a 54.44 ± 0.36 a 54.30 ± 1.07 b 53.44 ± 0.49 a 49.89 ± 0.87 a 50.11 ± 0.78 a 52.76 ± 1.86 a 6.80 ± 1.22 bc 6.14 ± 2.20 a 7.01 ± 0.69 a 6.31 ± 0.77 a 26.68 ± 2.02 bc 26.68 ± 2.02 bc 20.77 ± 1.85 a 14.56 ± 2.15 abc 9.70 ± 2.03 bc 说明:数值为平均值±标准误。同列不同小写字母表示不同林分间差异显著(P < 0.05)
下载: 导出CSV
表 3 土壤呼吸速率与土壤温度、湿度的关系
Table 3. Relationship between soil respiration rate and soil temperature and moisture
林分 $f\left( R \right) = a{{\rm{e}}^{bT}} $ $f\left( R \right) = a{h_{\rm{r}}}^2 + b{h_{\rm{r}}} + c $ $f\left( R \right) = a{{\rm{e}}^{bT}}{h_{\rm{r}}}^c $ Q10 a b R2 r a b c R2 r a b c R2 EU 0.487 0.069 0.437 0.628** -0.030 1.019 -4.251 0.443 0.507** 0.421 0.042 0.358 0.448 1.99 EP 0.488 0.053 0.630 0.817** -0.010 0.375 -1.069 0.378 0.568** 0.294 0.044 0.288 0.751 1.70 ET 1.442 0.035 0.638 0.781** -0.022 0.678 -1.009 0.387 0.432** 1.269 0.029 0.114 0.614 1.42 EC 0.635 0.044 0.758 0.851** -0.018 0.491 -0.923 0.183 0.173* 0.407 0.042 0.201 0.761 1.55 EUG 0.381 0.066 0.754 0.893** -0.014 0.480 -1.441 0.509 0.531** 0.270 0.054 0.256 0.839 1.93 PEC 0.631 0.047 0.798 0.884** -0.013 0.418 -0.769 0.412 0.315** 0.569 0.045 0.067 0.787 1.60 说明:R2为方程拟合优度, 即决定系数; r为土壤呼吸与土壤温度、湿度经Pearson相关性分析所得相关系数; *表示P<0.05;**表示P<0.01
下载: 导出CSV
表 4 土壤呼吸速率与土壤性质、凋落物及叶面积指数的相关性
Table 4. Correlation between soil respiration rate and soil properties, litter organic carbon storage and leaf area index (LAI)
土层/cm 相关性分析 容重 总孔隙度 毛管孔隙度 非毛管孔隙度 有机碳密度 凋落物有机碳密度 叶面积指数 土壤温度 土壤湿度 0-10 -0.752** 0.622** 0.024 0.611** 0.434 -0.109 0.681** -0.054 0.211 10~20 -0.149 0.165 0.434 -0.105 0.327 20-40 0.161 -0.360 -0.161 -0.149 0.174 40-60 0.164 -0.453 -0.071 -0.304 0.133 说明: **表示P<0.01, *表示P<0.05
下载: 导出CSV
-
[1] 唐罗忠, 葛晓敏, 吴麟, 等.南方型杨树人工林土壤呼吸及其组分分析[J].生态学报, 2012, 32(22):7000-7008. TANG Luozhong, GE Xiaomin, WU Lin, et al. Partitoning of autotrophic and heterotrophic soil respiration in southern type poplar plantations[J]. Acta Ecol Sin, 2012, 32(22):7000-7008. [2] ZHOU Lingyan, ZHOU Xuhui, SHAO Junjiong, et al. Interactive effects of global change factors on soil respiration and its components:a meta-analysis[J]. Global Change Biol, 2016, 22(9):3157-3169. [3] ANDREWS J A, HARRISON K G, MATAMALA R, et al. Separation of root respiration from total soil respiration using carbon-13 labeling during free-air carbon dioxide enrichment (FACE)[J]. Soil Sci Soc Am J, 1999, 63(2/4):1429-1435. [4] ARCHMILLER A A, SAMUELSON L J, LI Yingru. Spatial variability of soil respiration in a 64-year-old longleaf pine forest[J]. Plant Soil, 2016, 403(1/2):419-435. [5] MANDE H K, ABDULLAH A M, ARIS A Z, et al. Factors responsible for spatial and temporal variation of soil CO2 efflux in a 50 year recovering tropical forest, Peninsular Malaysia[J]. Environ Earth Sci, 2015, 73(9):5559-5569. [6] FANG Chu, MONCRIEFF J B, GHOLZ H L, et al. Soil CO2 efflux and its spatial variation in a Florida slash pine plantation[J]. Plant Soil, 1998, 205(2):135-146. [7] GAUMONT-GUAY D, BLACK T A, GRIFFIS T J, et al. Influence of temperature and drought on seasonal and interannual variations of soil, bole and ecosystem respiration in a boreal aspen stand[J]. Agric For Meteorol, 2006, 140(1):203-219. [8] HARTLEY I P, HEINEMEYER A, EVANS S P, et al. The effect of soil warming on bulk soil vs. rhizosphere respiration[J]. Global Change Biol, 2008, 13(12):2654-2667. [9] FERRÉC, CASTRIGNANÒA, COMOLLI R. Assessment of multi-scale soil-plant interactions in a poplar plantation using geostatistical data fusion techniques:relationships to soil respiration[J]. Plant Soil, 2015, 390(1/2):95-109. [10] SAIZ G, GREEN C, BUTTERBACH-BAHL K, et al. Seasonal and spatial variability of soil respiration in four Sitka spruce stands[J]. Plant Soil, 2006, 287(1/2):161-176. [11] RAICH J W, SCHLESINGER W H. The global dioxide carbon flux in respiration and its relationship to vegetation and climate[J]. Tellus Ser B-Chem Phys Meteorol, 1992, 44(2):81-99. [12] ZHAO Ningning, GUGGENBERGER G, SHIBISTOVA O, et al. Aspect-vegetation complex effects on biochemical characteristics and decomposability of soil organic carbon on the eastern Qinghai-Tibetan Plateau[J]. Plant Soil, 2014, 384(1/2):289-301. [13] CHEN Dima, ZHANG Chenlu, WU Jianping, et al. Subtropical plantations are large carbon sinks:evidence from two monoculture plantations in South China[J]. Agric For Meteorol, 2011, 151(9):1214-1225. [14] CORNWELL W K, CORNELISSEN J H C, AMATANGELO K, et al. Plant species traits are the predominant control on litter decomposition rates within biomes worldwide[J]. Ecol Lett, 2008, 11(10):1065-1071. [15] DAVIDSON E A, JANSSENS I A. Temperature sensitivity of soil carbon decomposition and feedbacks to climate change[J]. Nature, 2006, 440(7081):165-173. [16] LITTON C M, RAICH J W, RYAN M G. Carbon allocation in forest ecosystems[J]. Global Change Biol, 2007, 13(10):2089-2109. [17] 黄雪蔓, 刘世荣, 尤业明.第2代桉树人工纯林和混交林土壤呼吸及其组分研究[J].林业科学研究, 2014, 27(5):575-582. HUANG Xueman, LIU Shirong, YOU Yeming. Study on the soil respiration and its components of the second rotation Eucalyptus plantations in subtropical China[J]. For Res, 2014, 27(5):575-582. [18] 黄雪蔓. 南亚热带桉树人工林不同经营模式土壤碳动态变化及其调控机制[D]. 北京: 中国林业科学研究院, 2013. HUANG Xueman. The Dynamic and Regulation Mechanisms of Soil Carbon in Eucalyptus Plantations in Southern China[D]. Beijing: Chinese Academy of Forestry, 2013. [19] 袁渭阳. 短轮伐期巨桉人工林土壤碳转移研究[D]. 雅安: 四川农业大学, 2008. YUAN Weiyang. Studies on Soil Carbon Flux in the Short-Rotation Plantation of Eucalyptus grandis[D]. Ya'an: Sichuan Agricultural University, 2008. [20] 吴蒙, 马姜明, 梁士楚, 等.桂林尧山桉树及马尾松林秋季土壤碳通量特征[J].广西植物, 2014, 34(6):780-787. WU Meng, MA Jiangming, LIANG Shichu, et al. Characteristics on soil carbon flux of Eucalyptus spp. and Pinus massoniana forest in autumn in Yaoshan Mountain of Guilin[J]. Guihaia, 2014, 34(6):780-787. [21] 梁启鹏, 余新晓, 庞卓, 等.不同林分土壤有机碳密度研究[J].生态环境学报, 2010, 19(4):889-893. LIANG Qipeng, YU Xinxiao, PANG Zhuo, et al. Study on soil organic carbon density of different forest types[J]. Ecol Environ Sci, 2010, 19(4):889-893. [22] LUO Yiqi, WAN Shiqiang, HUI Dafeng, et al. Acclimatization of soil respiration to warming in a tall grass prairie[J]. Nature, 2001, 413(6856):622-625. [23] 姜艳, 王兵, 汪玉如, 等.亚热带林分土壤呼吸及其与土壤温湿度关系的模型模拟[J].应用生态学报, 2010, 21(7):1641-1648. JIANG Yan, WANG Bing, WANG Yuru, et al. Soil respiration in subtropical forests and model simulation of its relationship with soil temperature and moisture content[J]. Chin J Appl Ecol, 2010, 21(7):1641-1648. [24] BOWDEN R D, NADELHOFFER K J, BOONE R D, et al. Contributions of aboveground litter, belowground litter, and root respiration to total soil respiration in a temperate mixed hardwood forest[J]. Can J For Res, 1993, 23(7):1402-1407. [25] REY A, PEGORARO E, TEDESCHI V, et al. Annual variation in soil respiration and its components in a coppice oak forest in Central Italy[J]. Global Change Biol, 2002, 8(9):851-866. [26] VOSE J M, RYAN M G. Seasonal respiration of foliage, fine roots, and woody tissues in relation to growth, tissue N, and photosynthesis[J]. Glob Change Biol, 2002, 8(2):182-193. [27] 邓东周, 范志平, 王红, 等.土壤水分对土壤呼吸的影响[J].林业科学研究, 2009, 22(5):722-727. DENG Dongzhou, FAN Zhiping, WANG Hong, et al. Influences of soil moisture on soil respiration[J]. For Res, 2009, 22(5):722-727. [28] 吴蒙. 广西桉树及其他几种林分土壤呼吸特征[D]. 桂林: 广西师范大学, 2014. WU Meng. Characteristics on Soil Respiration of Eucalyptus spp. Plantations and Several Other Forests in Guangxi[D]. Guilin: Guangxi Normal University, 2014. [29] 王光军, 田大伦, 闫文德, 等.亚热带杉木和马尾松群落土壤系统呼吸及其影响因子[J].植物生态学报, 2009, 33(1):53-62. WANG Guangjun, TIAN Dalun, YAN Wende, et al. Soil system respiration and its controlling factors in Cunninghamia lanceolata and Pinus massoniana communities of subtropical China[J]. Chin J Plant Ecol, 2009, 33(1):53-62. [30] 陈光水, 杨玉盛, 吕萍萍, 等.中国森林土壤呼吸模式[J].生态学报, 2008, 28(4):1748-1761. CHEN Guangshui, YANG Yusheng, LÜ Pingping, et al. Regional patterns of soil respiration in China's forests[J]. Acta Ecol Sin, 2008, 28(4):1748-1761. [31] XU Ming, QI Ye. Spatial and seasonal variations of Q10 determined by soil respiration measurements at a Sierra nevadan Forest[J]. Glob Biogeochem Cycl, 2001, 15(3):687-696. [32] MOYANO F E, KUTSCH W L, SCHULZE E D. Response of mycorrhizal, rhizosphere and soil basal respiration to temperature and photosynthesis in a barley field[J]. Soil Biol Biochem, 2007, 39(4):843-853. [33] 王希群, 马履一, 贾忠奎, 等.叶面积指数的研究和应用进展[J].生态学杂志, 2005, 24(5):537-541. WANG Xiqun, MA Lüyi, JIA Zhongkui, et al. Research and application advances in leaf area index (LAI)[J]. Chin J Ecol, 2005, 24(5):537-541. [34] BOND-LAMBERTY B, THOMSON A. Temperature-associated increases in the global soil respiration record[J]. Nature, 2010, 464(7288):579-582. [35] REICHSTEIN M, REY A, FREIBAUER A, et al. Modeling temporal and large-scale spatial variability of soil respiration from soil water availability, temperature and vegetation productivity indices[J]. Glob Biogeochem Cycl, 2003, 17(4):1104. doi:10.1029/2003GB002035. [36] 王娓, 周晓梅, 郭继勋.东北羊草草原两种主要群落环境因素对土壤呼吸贡献量的影响[J].草业学报, 2002, 11(1):12-16. WANG Wei, ZHOU Xiaomei, GUO Jixun. Effect of environmental factors on CO2 release of soil respiration of two main communities in Lymus chinensis grassland in northeastern China[J]. Acta Pratac Sin, 2002, 11(1):12-16. [37] 徐洪灵, 张宏, 张伟.川西北高寒草甸土壤理化性质对土壤呼吸速率影响研究[J].四川师范大学学报(自然科学版), 2012, 35(6):835-841. XU Hongling, ZHANG Hong, ZHANG Wei. Influence of soil physical and chemical properties on soil respiration rate of alpine meadow in the northwestern plateau of Sichuan Province[J]. J Sichuan Norm Univ Nat Sci, 2012, 35(6):835-841. [38] RYAN M G, LAW B E. Interpreting, measuring, and modeling soil respiration[J]. Biogeochemistry, 2005, 73(1):3-27. [39] 宋启亮, 董希斌.大兴安岭5种类型低质林土壤呼吸日变化及影响因素[J].东北林业大学学报, 2014, 42(9):77-82. SONG Qiliang, DONG Xibin. Diurnal variation and influenced factors of soil respiration in five typical low-quality forest in Daxing'an Mountains[J]. J Northeast For Univ, 2014, 42(9):77-82. [40] 王建国. 黄土高原水蚀风蚀交错区土壤呼吸及其影响因素研究[D]. 杨凌: 西北农林科技大学, 2011. WANG Jianguo. Affecting Factors of Soil Respiration in the Water-wind Erosion Crisscross Region[D]. Yangling: Northwest A&F University, 2011. [41] 杨玉盛, 董彬, 谢锦升, 等.森林土壤呼吸及其对全球变化的响应[J].生态学报, 2004, 24(3):583-591. YANG Yusheng, DONG Bin, XIE Jinsheng, et al. Soil respiration of forest ecosystems and its respondence to global change[J]. Acta Ecol Sin, 2004, 24(3):583-591. [42] 王国兵, 郝岩松, 王兵, 等.土地利用方式的改变对土壤呼吸及土壤微生物生物量的影响[J].北京林业大学学报, 2006, 28(增刊2):73-79. WANG Guobing, HAO Yansong, WANG Bing, et al. Influence of land-use change on soil respiration and soil microbial biomass[J]. J Beijing For Univ, 2006, 28(suppl 2):73-79. [43] 韩营营, 黄唯, 孙涛, 等.不同林龄白桦天然次生林土壤碳通量和有机碳储量[J].生态学报, 2015, 35(5):1460-1469. HAN Yingying, HUANG Wei, SUN Tao, et al. Soil organic carbon stocks and fluxes in different age stands of secondary Betula platyphylla in Xiaoxing'an Mountain, China[J]. Acta Ecol Sin, 2015, 35(5):1460-1469. [44] 刘绍辉, 方精云.土壤呼吸的影响因素及全球尺度下温度的影响[J].生态学报, 1997, 17(5):469-476. LIU Shaohui, FANG Jingyun. Effect factors of soil respiration and the temperature's effects on soil respiration in the global scale[J]. Acta Ecol Sin, 1997, 17(5):469-476. [45] XU Ming, QI Ye. Soil-surface CO2 efflux and its spatial and temporal variations in a young ponderosa pine plantation in northern California[J]. Glob Change Biol, 2001, 7(6):667-677. [46] XU Xia, ZHOU Yan, RUAN Honghua, et al. Temperature sensitivity increases with soil organic carbon recalcitrance along an elevational gradient in the Wuyi Mountains, China[J]. Soil Biol Biochem, 2010, 42(10):1811-1815. [47] 陈灿, 江灿, 范海兰, 等.凋落物去除/保留对杉木人工林林窗和林内土壤呼吸的影响[J].生态学报, 2017, 37(1):102-109. CHEN Can, JIANG Can, FAN Hailan, et al. Effects of removing/keeping litter on soil respiration in and outside the gaps in Chinese fir plantation[J]. Acta Ecol Sin, 2017, 37(1):102-109. [48] 雷蕾, 肖文发, 曾立雄, 等.马尾松林土壤呼吸组分对不同营林措施的响应[J].生态学报, 2016, 36(17):5360-5370. LEI Lei, XIAO Wenfa, ZENG Lixiong, et al. Responses of soil respiration and its components to forest management in Pinus massoniana stands[J]. Acta Ecol Sin, 2016, 36(17):5360-5370. [49] 梁国华, 吴建平, 熊鑫, 等.南亚热带不同演替阶段森林土壤呼吸对模拟酸雨的响应[J].生态学杂志, 2016, 35(1):125-134. LIANG Guohua, WU Jianping, XIONG Xin, et al. Response of soil respiration to simulated acid rain in three successional subtropical forests in southern China[J]. Chin J Ecol, 2016, 35(1):125-134. -

-
链接本文:
https://zlxb.zafu.edu.cn/article/doi/10.11833/j.issn.2095-0756.2018.03.004
 点击查看大图
点击查看大图

计量
- 文章访问数: 3511
- HTML全文浏览量: 741
- PDF下载量: 346
- 被引次数: 0
