




-
土壤碳库是森林生态系统碳库中最重要的组成部分,其微小的变化都会对森林碳库产生极大的影响[1]。如何增加土壤固碳来减缓温室效应是目前森林生态系统碳管理面临的重要挑战之一。作为森林经营管理的一项重要手段,间伐可改变林分密度、林下植被、土壤小气候和土壤养分,进而影响土壤有机碳的输入、输出和固持[2−4]。研究表明:在中国亚热带森林生态系统中,土壤异养呼吸占总呼吸的比例为60%[5−7]。因此,探究间伐后森林土壤异养呼吸的变化,对以“固碳增汇”为导向的森林碳汇经营具有重要的理论和实践意义。
土壤异养呼吸是土壤微生物分解有机质释放二氧化碳(CO2)的过程,是土壤呼吸的重要组成部分[8],也是准确估算陆地生态系统碳平衡的关键因子[9]。土壤异养呼吸大小与植被类型、土壤类型、微生物活性等生物因素有关,同时易受土壤温度和土壤水分等环境因素的影响[10]。研究表明:间伐后土壤温度升高,增强了间伐剩余物中微生物的分解作用,从而导致异养呼吸速率升高[6, 10]。温度敏感性系数(Q10)是衡量土壤异养呼吸强度的重要参数,也是衡量全球变暖与土壤碳循环之间反馈作用强度的关键指标[11]。它反映了土壤温度每增加10 ℃,土壤异养呼吸增加的程度[12]。目前,关于间伐处理引起的异养呼吸Q10的变化主要有升高[6]和降低[11]2种结论。总有机碳、微生物生物量碳、微生物区系等都是影响Q10的主要因素[12]。然而,由于土壤类型、林分类型以及测定方法等的差异,Q10对间伐的响应仍存在较大不确定性。国内外关于间伐对土壤呼吸的研究集中于纯林与人工林。如丁驰等[13]研究了间伐和施肥对亚热带杉木Cunninghamia lanceolata人工林土壤温室气体排放的影响;LEI等[6]研究发现:15%的轻度间伐显著促进了马尾松Pinus massoniana林土壤异养呼吸和总呼吸速率。但是,目前尚不清楚间伐如何影响天然针阔混交林土壤的异养呼吸。
秦岭是中国东半壁的南北气候分界线,其独特的地理位置、特殊的自然条件和复杂的地形地貌,造就了该地区丰富多样的植被类型和较高的生物多样性。松栎混交林是秦岭的主要林分类型,对整个地区的森林生态系统碳汇都有着极其重要的作用[14]。本研究选择秦岭不同间伐恢复年限的松栎混交林为研究对象,探究间伐对混交林生长季土壤异养呼吸通量及温度敏感性的影响,以期为该地区森林碳汇经营提供科学依据。
-
研究区位于陕西宁东林业局新矿林场,该区地处北亚热带北缘的秦岭山地(33°20′~33°26′N,108°32′~108°34′E),海拔为1 095.0~2 591.0 m,年均气温为8.5 ℃,年平均降水量为908.0 mm。土壤为山地黄棕壤,土层厚度为30~60 cm。研究区属于中国南方亚热带常绿阔叶林和针阔混交林经营区,该区为了恢复地带性顶级群落,采取低强度间伐和林冠下补植等保护经营作业法。林分类型为20世纪70年代末采伐后天然更新形成的次生松栎混交林,郁闭度约0.6。主要建群树种为油松Pinus tabuliformis、锐齿栎Quercus aliena var. acuteserrata、华山松Pinus armandii,伴生树种有漆树Toxicodendron vernicifluum、小叶女贞Ligustrum quihoui、青榨槭Acer davidii等。林下植被以卫矛Euonymus alatus、木姜子Litsea pungens、悬钩子属Rubus、披针叶薹草Carex lanceolata、龙牙草Agrimonia pilosa、茜草Rubia cordifolia为主。
选择该林场2010和2018年分批完成间伐强度约15%的作业林班,主要针对胸径<8 cm和受损树木进行伐除作业。同一小班或临近小班林分生长条件相对一致,完全随机设置3种处理:未间伐样地(ck)、间伐12 a样地(12 a)和间伐4 a样地(4 a)。每个处理各设置4块20 m×30 m的样方,各样地具体情况如表1所示。
表 1 样地林分信息
Table 1. Information on forest stands of sample plots
处理 主要树种 胸径/cm 林分密度/(株·hm−2) 郁闭度 坡度/(°) 海拔/m 未间伐 华山松、锐齿栎 14.6±0.9 1 420±88 0.7 18.3±5.4 1 585.0±107.1 间伐4 a 油松、锐齿栎 14.4±2.0 1 208±355 0.5 25.0±5.0 1 449.8±21.5 间伐12 a 油松、锐齿栎 13.7±1.2 1 254±207 0.6 8.8±2.2 1 757.6±34.9 说明:数值为平均值±标准差。华山松P. armandii,油松P. tabuliformis,锐齿栎 Q. aliena var. acuteserrata。 -
于2022年5月在每个样地内S型采集土壤样品(7个点),在0~20 cm土层取1 kg的土壤,将土样中的砂石和植物根系去除,混合均匀后装入无菌袋。带回实验室放置于4 ℃冰箱保存。土壤总有机碳(SOC)采用重铬酸钾-外加热法测定[15];土壤容重(BD)采用环刀法测定;土壤含水量(SWC)采用烘干法测定;土壤全碳(TC)、全氮(TN)采用元素分析仪测定;土壤微生物生物量碳(MBC)采用氯仿熏蒸法-K2SO4法提取,后用总有机碳分析仪测定;可溶性有机碳(DOC)采用总有机碳分析仪测定。
原位土壤-大气界面异养呼吸通量采用静态箱-气相色谱法测定[16]。采样前,于2021年11月采用壕沟法隔绝根部,并在样方内的垂直土壤剖面安装聚氯乙烯(PVC)呼吸环和静态箱。于2022年5—10月,在无降水的白天进行气体采样,2周采集气体样品1次(部分月份因天气原因只采集了1次)。每个箱体采气时间为1 h,采4针,2针的时间间隔约20 min。采集的气体存放于20 mL真空气瓶,使用气相色谱仪检测CO2含量。
-
原位土壤-大气界面异养呼吸通量(F)计算如下:
$$ F=\rho \times \frac{V}{A}\times \frac{P}{{P}_{0}}\times \frac{{T}_{0}}{T}\times \frac{{\rm{d}}{C}_{t}}{{{\rm{d}}}{t}} 。 $$ (1) 式(1)中:F为单位时间单位面积内静态箱的CO2通量(g·m−2·h−1);$ \rho $为标准大气压下的气体密度;V为腔室内的气体体积;A为腔室覆盖面积;P为采样点的大气压;T为采样时的绝对温度;P0、T0分别为标准大气压和绝对温度(273.15 K);dCt/dt为采样期间气体体积分数(Ct,μL·L−1)随时间(t,min)变化的斜率,需满足回归系数(R2)>0.9。
土壤异养呼吸累计排放通量计算如下:
$$ E=\sum _{t=1}^{n}({D}_{t+1}-{D}_{t})\frac{{(F}_{t+1}+{F}_{t})}{2} 。 $$ (2) 式(2)中:$ E $为异养呼吸累计排放量(kg·m−2);$ n $为测定总次数;t为测定时间;$ {D}_{t+1}-{D}_{t} $表示相邻2次测定的时间间隔(d);$ {{F}_{t}\mathrm{和}F}_{t+1} $分别为相邻2次采样的异养呼吸速率(g·m−2·d−1)。
通过全球气候的第5代大气再分析系统ERA-5 (the fifth generation ECMWF reanalysis for the global climate and weather)得到0~7 cm的土壤温度数据。土壤异养呼吸速率与土壤温度之间的关系采用Vant’Hoff指数模型拟合,公式如下:
$$ R=a{{\rm{e}}}^{bT} 。 $$ (3) 式(3)中:R为土壤异养呼吸速率(g·m−2·h−1);T为0~7 cm土壤温度(℃);a为0 ℃时的异养呼吸速率;b为温度反应系数。
温度敏感性系数(Q10)计算如下:
$$ {Q}_{10}={{\rm{e}}}^{10b} 。 $$ (4) 式(4)中:$ {Q}_{10} $为温度升高10 ℃时土壤呼吸速率变化的倍数,可衡量土壤呼吸速率对温度的响应程度[17];b为温度反应系数。
-
采用SPSS 26 对不同处理土壤异养呼吸速率和土壤性质进行单因素方差分析和最小显著差异法检验分析;采用指数增长模型分析土壤异养呼吸通量和土壤温度间的关系;进而计算不同处理的温度敏感性系数(Q10);使用GraphPad Prism 8制图。所有数据为平均值±标准差。
-
如表2所示:间伐后土壤pH相比对照显著上升(P<0.05);微生物生物量碳均显著降低(P<0.05);间伐4 a样地的土壤总有机碳质量分数显著下降(P<0.05),间伐12 a的样地与未间伐样地无显著差异;间伐4 a样地的土壤碳氮比显著高于间伐12 a样地(P<0.05),但与未间伐样地相比皆无显著差异;间伐4 a样地的土壤含水量显著低于其他2个样地(P<0.05),间伐12 a样地与未间伐样地无显著差异。
表 2 土壤基本理化性质
Table 2. Soil basic physicochemical properties
处理 pH 容重/
(g·cm−3)含水量/% 总有机碳/
(g·kg−1)碳氮比 全氮/(g·kg−1) 可溶性有机碳/
(mg·kg−1)微生物生物量碳/
(g·kg−1)未间伐 4.9±0.1 b 1.1±0.1 a 35.9±3.6 a 39.6±6.1 a 12.6±0.6 a 3.4±0.5 a 63.0±2.5 a 1.8±0.1 a 间伐4 a 5.8±0.1 a 1.3±0.2 a 25.6±3.0 b 17.3±2.6 b 12.5±0.2 a 2.3±0.3 a 65.2±4.6 a 0.7±0.1 b 间伐12 a 5.7±0.1 a 1.2±0.1 a 37.2±1.5 a 29.2±1.3 ab 9.4±0.4 b 2.7±0.1 a 68.3±6.3 a 0.9±0.1 b 说明:不同字母表示同一理化性质在不同处理间差异显著(P<0.05)。 -
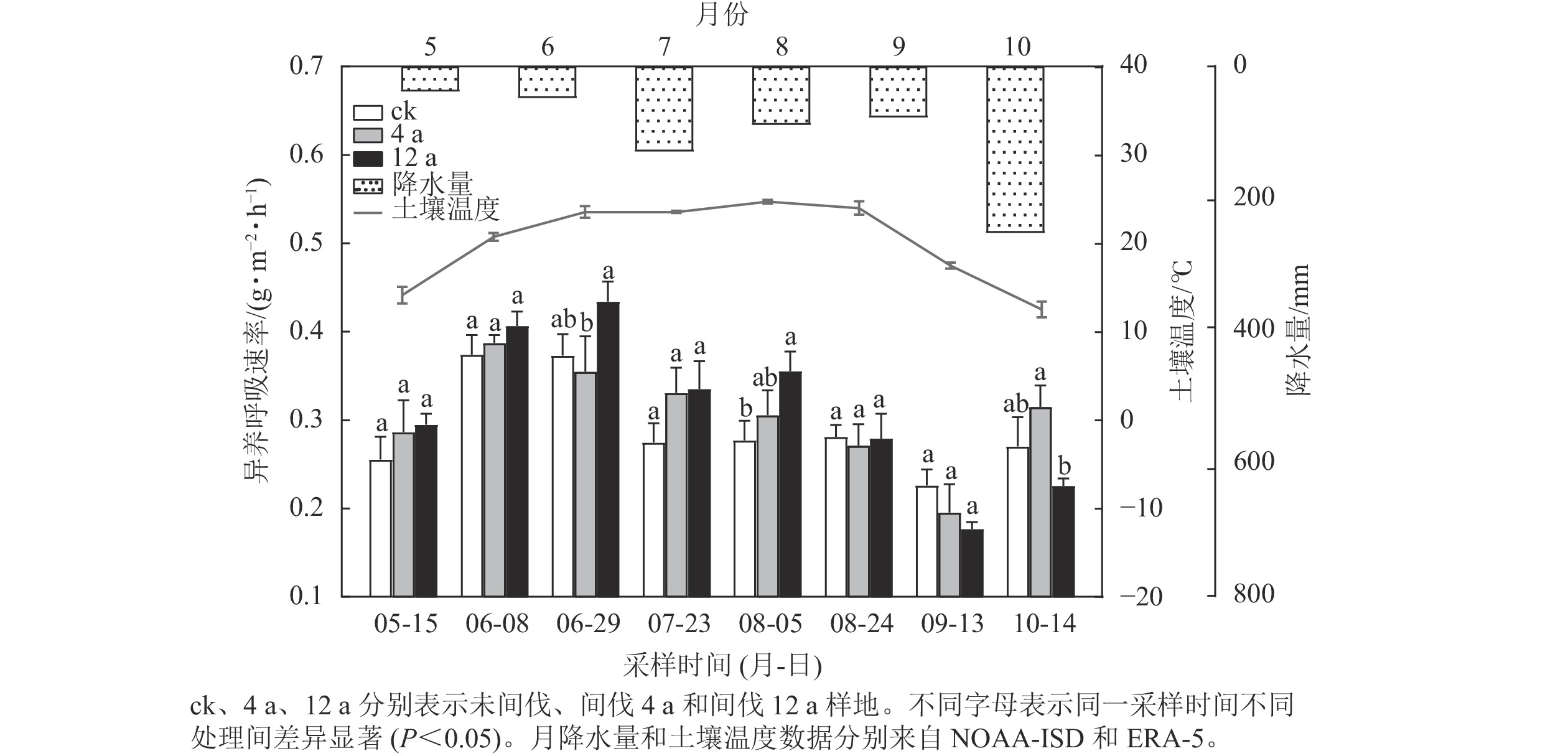
不同间伐年限处理下,秦岭松栎混交林生长季土壤异养呼吸速率呈现“双峰型”变化规律(图1),通量峰值分别出现在6和10月。6月29日(间伐4 a和间伐12 a样地)、8月5日(未间伐和间伐12 a样地)和10月14日(间伐4 a和间伐12 a样地)的通量存在显著差异(P<0.05),其余时间均无显著差异。整体来看,6月3个处理呼吸速率到达顶峰,未间伐、间伐4 a和间伐12 a样地的异养呼吸速率分别为0.38、0.39和0.44 g·m−2·h−1;9月未间伐、间伐4 a和间伐12 a样地的呼吸速率均最低,分别为0.23、0.19和0.19 g·m−2·h−1。

图 1 不同间伐恢复年限松栎混交林土壤异养呼吸速率的变化
Figure 1. Changes of soil heterotrophic respirations in oak-pine mixed forests of treatments after different thinning years
-
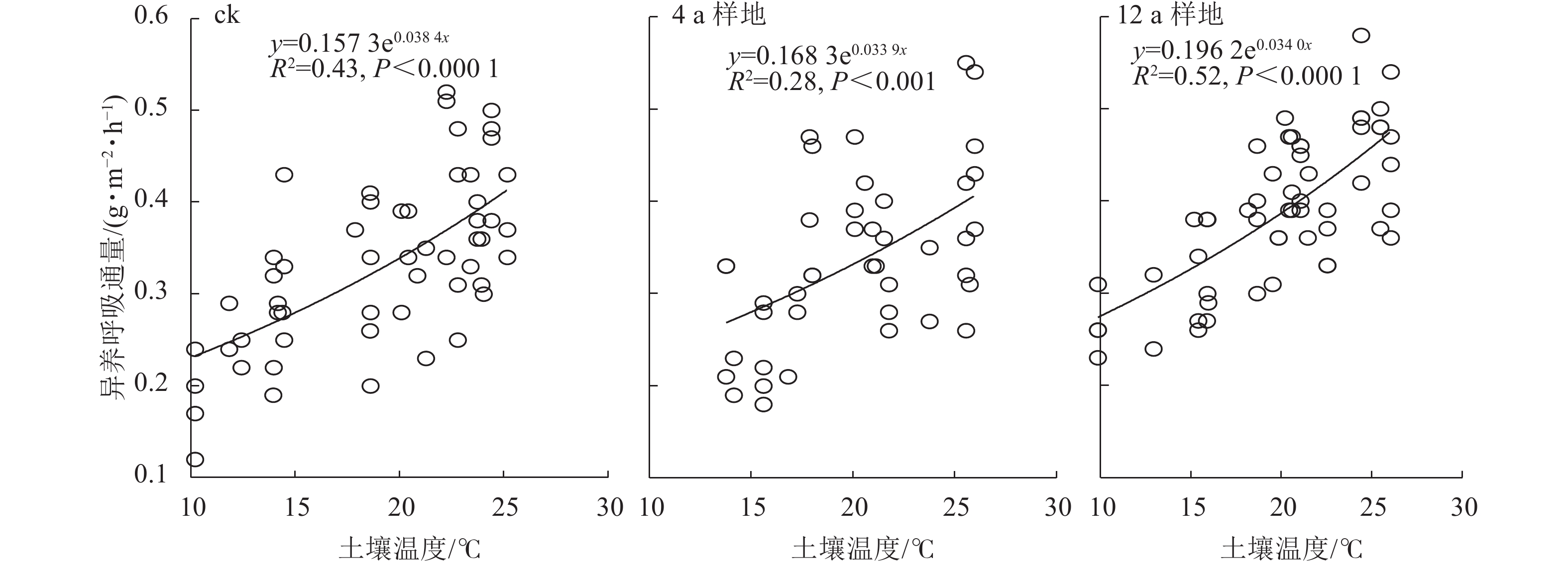
秦岭松栎混交林生长季3种不同间伐年限处理下,土壤异养呼吸与土壤温度呈极显著指数正相关(P<0.001,图2)。在相同的土壤温度下,秦岭不同间伐恢复年限松栎混交林生长季土壤异养呼吸Q10从大到小依次为未间伐样地(1.47)、间伐12 a样地(1.41)、间伐4 a样地(1.40)。

图 2 不同间伐恢复年限松栎混交林土壤异养呼吸与土壤温度的关系
Figure 2. Relationships between soil heterotrophic respirations and soil temperatures in oak-pine mixed forests of treatments after different thinning recovery years
-
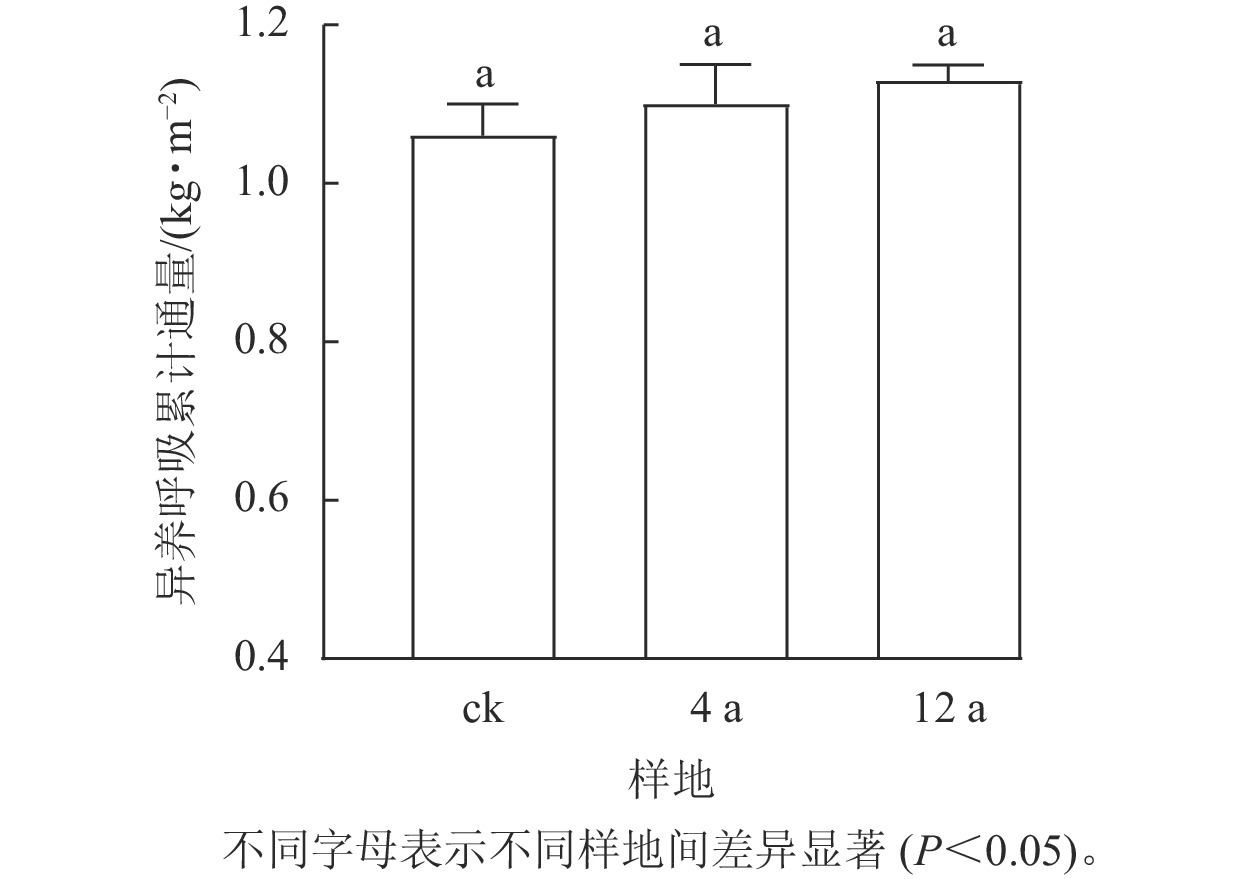
不同间伐年限下秦岭松栎混交林生长季土壤异养呼吸CO2累计排放量在3个处理间差异不显著(图3,P>0.05)。生长季各处理异养呼吸累计通量从大到小依次为间伐12 a样地 (1.13 kg·m−2)、间伐4 a样地(1.10 kg·m−2)、未间伐样地(1.06 kg·m−2)。间伐4 a和间伐12 a样地累计通量分别比未间伐样地高3.8%和6.7%。

图 3 不同间伐恢复年限松栎混交林生长季异养呼吸累计通量
Figure 3. Cumulative amount of soil heterotrophic respirations in oak-pine mixed forests after different recovery years in the growing season
-
本研究发现:生长季土壤异养呼吸通量呈现双峰曲线的变化形式。这种现象可能是由于水热条件变化导致异养呼吸出现波动,而较短的采样时间间隔(15~30 d)则正好反映了这种变化。降雨脉冲导致的土壤碳排放变异在森林生态系统中普遍存在,也是森林碳收支不确定性的重要来源,这与PAN等[18]模拟气温和降水变化对青藏高原中部土壤异养呼吸影响的研究结论一致。夏季初期在适宜的水热联合驱动下,土壤微生物代谢旺盛[19],因此土壤异养呼吸速率在此时达到高值。7—8月温度逐渐升高,太阳辐射增强,水分蒸散加快,较低的土壤含水量一定程度限制了微生物活性,从而土壤异养呼吸受限[20],因此通量相较于6月有所下降。9月土壤温度逐渐降低,异养呼吸速率下降。10月降水量增多,土壤含水量的提高导致异养呼吸速率提升,这与MAZZA等[21]认为湿季土壤CO2排放较多的结论相一致。
-
间伐由于人为的扰动使得林地内小环境发生改变,对土壤温度、pH、碳氮质量分数、有机碳组分和质量分数等产生影响,进而影响土壤微生物的呼吸作用[22−23]。本研究发现:间伐在大部分测量期间(除了6月29日、8月5日和10月14日)对异养呼吸通量没有显著影响。但间伐后土壤异养呼吸累计通量略有升高,间伐4 a和间伐12 a样地的累计通量比未间伐样地分别高3.8%和6.7%。间伐后由于林地内光辐射增强导致土壤温度升高,对土壤微生物群落结构、酶活性以及有机质分解等带来直接或间接的影响,进而促进土壤异养呼吸速率的提高[24−26]。土壤pH的显著升高为微生物活动提供了适宜的弱酸性环境,加速其繁衍和呼吸作用[27]。同时,适宜的弱酸性土壤环境使得土壤吸附的可溶性有机碳增多,为微生物提供了大量易于吸收的底物,土壤碳排放增加。间伐后总有机碳质量分数下降[28−29],但土壤可溶性有机碳与总有机碳质量分数的比值升高,即底物可利用性升高。这与传统研究[30−31]所认为的土壤有机质减少,土壤微生物因缺少呼吸底物而死亡导致异养呼吸速率下降的结果不符。然而,土壤有机碳质量分数低并不一定代表异养呼吸低,土壤有机碳质量分数、质量、可利用性以及周转的相互作用最终决定异养呼吸的时空格局[32]。
-
本研究通过指数增长模型发现:土壤温度与异养呼吸通量呈极显著正相关关系,即异养呼吸通量随着土壤温度的升高而升高。且间伐后土壤异养呼吸的温度敏感性降低,间伐4 a样地的值最低。3个处理的Q10为1.40~1.47,处于土壤Q10平均值(1.09~2.38)的较低水平[9]。研究表明:土壤温度和异养呼吸具有很好的相关性,土壤温度的变化可能会导致土壤异养呼吸季节变化规律发生偏移[10, 33]。土壤温度升高加快了微生物呼吸底物的消耗速率,降低土壤有机碳质量分数,从而导致Q10的降低[11, 34]。但由于本研究运用的是区域遥感温度,未具体到各个样地内的实测土壤温度,因而异养呼吸的温度敏感性结果仍存在一定的不确定性,有待进一步验证。
-
秦岭松栎混交林生长季土壤异养呼吸速率呈“双峰型”季节变化规律,峰值分别出现在初夏6月和雨季10月。间伐4 a和间伐12 a样地的林分异养呼吸累计通量相比未间伐样地略有增加,但3个处理间并不存在显著差异。土壤温度是异养呼吸通量的关键影响因子,并且异养呼吸温度敏感性Q10在间伐后呈现先下降后上升的趋势。
Effects of thinning on soil heterotrophic respiration of oak-pine mixed forests in Qinling Mountains
-
摘要:
目的 研究间伐对秦岭松栎混交林土壤异养呼吸的影响,可深入了解该地区间伐对森林土壤异养呼吸造成的碳损失情况,为研究区森林经营碳汇提供科学依据。 方法 以秦岭松栎混交林为研究对象,对未间伐样地(ck)、间伐4 a、间伐12 a的样地采用静态箱-气相色谱法监测生长季土壤异养呼吸速率的月变化,并计算各处理异养呼吸的温度敏感性(Q10)。 结果 ①相比ck,间伐4 a、间伐12 a的样地土壤微生物生物量碳均显著下降(P<0.05),土壤pH均显著上升(P<0.05),土壤总有机碳质量分数在间伐4 a的样地中显著下降(P<0.05)。间伐12 a样地的理化指标相比间伐4 a的样地更接近ck。②生长季土壤异养呼吸呈现“双峰型”变化规律,峰值分别出现在6和10月。异养呼吸累计通量在间伐后略有上升但不显著,不同样地从大到小依次为间伐12 a、间伐4 a、ck。③土壤温度与异养呼吸呈极显著指数正相关(P<0.001);间伐后Q10降低,不同样地Q10从大到小依次为ck、间伐12 a、间伐4 a。 结论 土壤温度是影响松栎混交林异养呼吸的关键因素。间伐没有导致秦岭松栎混交林异养呼吸通量显著增大。图3表2参34 Abstract:Objective This study, with an investigation of the effects of thinning on soil heterotrophic respiration in oak-pine mixed forests in the Qinling Mountains, is aimed to better understand the carbon loss caused by forest soil heterotrophic respiration under thinning treatments, providing a scientific basis for forest management decisions in the study area. Method The static chamber-gas chromatography technique was employed to monitor the variations of soil heterotrophic respiration flux in the growing season for the unthinned plots (4 replicates, ck) and the thinning plots (4 replicates) 4 and 12 years ago (4 a and 12 a), respectively. The temperature sensitivities (Q10) of heterotrophic respiration were also calculated. Result (1) There was a remarkable decrease in the soil microbial biomass carbon content (P<0.05), but a significant increase in the the soil pH (P<0.05) in both 4 a and 12 a compared to ck, while the decrease in the soil organic carbon content of 4 a after thinning was significant (P<0.05). Generally, the soil physical and chemical index in 12 a were closer to that in the ck than the 4 a treatment. (2) The soil heterotrophic respiration during the growing season showed a “bimodal” pattern, and the peaks appeared in June and October, respectively. However the cumulative heterotrophic respiration increased after thinning but not significantly, with the order from large to small being 12 a>4 a>ck. (3) The soil temperature was significantly and exponentially correlated with the soil heterotrophic respiration (P<0.05). The temperature sensitivities Q10 decreased after thinning, with the order from large to small being ck>12 a>4 a. Conclusion The soil temperature is a key factor affecting soil heterotrophic respiration in oak-pine mixed forests and the thinning treatment do not promote soil heterotrophic respiration of the oak-pine mixed forests in the growing season at the Qinling Mountains. [Ch, 3 fig. 2 tab. 34 ref.] -
图 1 不同间伐恢复年限松栎混交林土壤异养呼吸速率的变化
Figure 1 Changes of soil heterotrophic respirations in oak-pine mixed forests of treatments after different thinning years
图 2 不同间伐恢复年限松栎混交林土壤异养呼吸与土壤温度的关系
Figure 2 Relationships between soil heterotrophic respirations and soil temperatures in oak-pine mixed forests of treatments after different thinning recovery years
图 3 不同间伐恢复年限松栎混交林生长季异养呼吸累计通量
Figure 3 Cumulative amount of soil heterotrophic respirations in oak-pine mixed forests after different recovery years in the growing season
表 1 样地林分信息
Table 1. Information on forest stands of sample plots
处理 主要树种 胸径/cm 林分密度/(株·hm−2) 郁闭度 坡度/(°) 海拔/m 未间伐 华山松、锐齿栎 14.6±0.9 1 420±88 0.7 18.3±5.4 1 585.0±107.1 间伐4 a 油松、锐齿栎 14.4±2.0 1 208±355 0.5 25.0±5.0 1 449.8±21.5 间伐12 a 油松、锐齿栎 13.7±1.2 1 254±207 0.6 8.8±2.2 1 757.6±34.9 说明:数值为平均值±标准差。华山松P. armandii,油松P. tabuliformis,锐齿栎 Q. aliena var. acuteserrata。  下载: 导出CSV
下载: 导出CSV
表 2 土壤基本理化性质
Table 2. Soil basic physicochemical properties
处理 pH 容重/
(g·cm−3)含水量/% 总有机碳/
(g·kg−1)碳氮比 全氮/(g·kg−1) 可溶性有机碳/
(mg·kg−1)微生物生物量碳/
(g·kg−1)未间伐 4.9±0.1 b 1.1±0.1 a 35.9±3.6 a 39.6±6.1 a 12.6±0.6 a 3.4±0.5 a 63.0±2.5 a 1.8±0.1 a 间伐4 a 5.8±0.1 a 1.3±0.2 a 25.6±3.0 b 17.3±2.6 b 12.5±0.2 a 2.3±0.3 a 65.2±4.6 a 0.7±0.1 b 间伐12 a 5.7±0.1 a 1.2±0.1 a 37.2±1.5 a 29.2±1.3 ab 9.4±0.4 b 2.7±0.1 a 68.3±6.3 a 0.9±0.1 b 说明:不同字母表示同一理化性质在不同处理间差异显著(P<0.05)。
下载: 导出CSV
-
[1] GEORGIOU K, JACKSON R B, VINDUSKOVA O, et al. Global stocks and capacity of mineral-associated soil organic carbon[J/OL]. Nature Communications, 2022, 13(1): 3797[2023-03-01]. doi: 0.1038/s41467-022-31540-9. [2] WANG Dong, CHEN Xinli, CHEN Hongyi, et al. Contrasting effects of thinning on soil CO2 emission and above-and belowground carbon regime under a subtropical Chinese fir plantation [J]. Science of the Total Environment, 2019, 690: 361 − 369. [3] CHEN Xinli, WANG Dong, CHEN Xin, et al. Soil microbial functional diversity and biomass as affected by different thinning intensities in a Chinese fir plantation [J]. Applied Soil Ecology, 2015, 92: 35 − 44. [4] ZHOU Lili, CAI Liping, HE Zongming, et al. Thinning increases understory diversity and biomass, and improves soil properties without decreasing growth of Chinese fir in southern China [J]. Environmental Science and Pollution Research, 2016, 23(23): 24135 − 24150. [5] LI Renshan, ZHENG Wenhui, YANG Qingpeng. The response of soil respiration to thinning was not affected by understory removal in a Chinese fir (Cunninghamia lanceolata) plantation [J]. Geoderma, 2019, 353: 47 − 54. [6] LEI Lei, XIAO Wenfa, ZENG Lixiong, et al. Thinning but not understory removal increased heterotrophic respiration and total soil respiration in Pinus massoniana stands [J]. Science of the Total Environment, 2018, 621: 1360 − 1369. [7] ZHANG Hui, YING Binbin, HU Yanjing, et al. Response of soil respiration to thinning is altered by thinning residue treatment in Cunninghamia lanceolata plantations[J/OL]. Agricultural and Forest Meteorology, 2022, 324: 109089[2023-03-01]. doi: 10.1016/j.agrformet.2022.109089. [8] 李小宇, 李勇, 于寒青, 等. 退耕还林坡地土壤CO2排放的空间变化: 地形的控制作用[J]. 植物营养与肥料学报, 2015, 21(5): 1217 − 1224. LI Xiaoyu, LI Yong, YU Hanqing, et al. Spatial changes in soil CO2 emission from re-forested hill slopes on the Loess Plateau: a geomorphic control [J]. Journal of Plant Nutrition and Fertilizer, 2015, 21(5): 1217 − 1224. [9] 周涛, 史培军, 惠大丰, 等. 中国土壤呼吸温度敏感性空间格局的反演[J]. 中国科学(生命科学), 2009, 39(3): 315 − 322. ZHOU Tao, SHI Peijun, HUI Dafeng, et al. Inversion of spatial pattern of soil respiration temperature sensitivity in China [J]. Scientia Sinica (Vitae), 2009, 39(3): 315 − 322. [10] ZEESHAN M, ZHOU Wenjun, WU Chuansheng, et al. Soil heterotrophic respiration in response to rising temperature and moisture along an altitudinal gradient in a subtropical forest ecosystem, southwest China[J/OL]. Science of the Total Environment, 2022, 816: 151643[2023-03-01]. doi: 10.1016/j.scitotenv.2021.151643. [11] HAN Mengguang, GAO Weifeng, SHI Baoku, et al. Long-term (42 years) effect of thinning on soil CO2 emission in a mixed broadleaved-Korean pine (Pinus koraiensis) forest in Northeast China [J]. Pedosphere, 2021, 31(2): 353 − 362. [12] 王陈里, 张利杰, 张仲富, 等. 基于室内试验的土壤呼吸温度敏感性影响因素整合分析[J]. 西南林业大学学报(自然科学), 2022, 42(3): 128 − 137. WANG Chenli, ZHANG Lijie, ZHANG Zhongfu, et al. Meta-analysis of the factors affecting soil respiration temperature sensitivity based on indoor measurements [J]. Journal of Southwest Forestry University (Natural Science), 2022, 42(3): 128 − 137. [13] 丁驰, 雷梅, 甘子莹, 等. 间伐和施肥对杉木人工林土壤温室气体排放的影响[J]. 生态学杂志, 2022, 41(6): 1056 − 1065. DING Chi, LEI Mei, GAN Ziying, et al. Effects of thinning and fertilization on GHGs emissions in Chinese fir plantation soil [J]. Chinese Journal of Ecology, 2022, 41(6): 1056 − 1065. [14] 刘畅. 陕西省秦岭南北气候的差异性及其变化[D]. 兰州: 兰州大学, 2016. LIU Chang. Climate Change and its Difference Between Southern and Northern Qinling in Shaanxi Province[D]. Lanzhou: Lanzhou University, 2016. [15] SPARKS D L, PAGE A L, HELMKE P A, et al. Methods of Soil Analysis: Part 3 Chemical Methods[M]. Madison: Soil Science Society of America, American Society of Agronomy, 1996. [16] MOSIER A R, MORGAN J A, KING J Y, et al. Soil-atmosphere exchange of CH4, CO2, NOx, and N2O in the Colorado shortgrass steppe under elevated CO2 [J]. Plant and Soil, 2002, 240(2): 201 − 211. [17] 胡文沛, 张闯, 胡春胜, 等. 长期增温和施氮对华北平原农田土壤呼吸及其温度敏感性的影响[J]. 中国生态农业学报, 2022, 30(5): 761 − 768. HU Wenpei, ZHANG Chuang, HU Chunsheng, et al. Effects of long-term warming and nitrogen fertilization on soil respiration and temperature sensitivity in the North China Plain [J]. Chinese Journal of Eco-Agriculture, 2022, 30(5): 761 − 768. [18] PAN Yongjie, LI Suosuo, LI Xia, et al. Modeling the effects of changes in air temperature and precipitation on heterotrophic respiration in the central Tibetan Plateau [J]. Theoretical and Applied Climatology, 2022, 149(1/2): 451 − 464. [19] 陈蕾, 董希斌. 抚育间伐强度对大兴安岭落叶松天然次生林冻融后期土壤呼吸及性质的影响[J]. 东北林业大学学报, 2020, 48(6): 152 − 156, 162. CHEN Lei, DONG Xibin. Effect of tending thinning intensity on soil respiration and physicochemical properties of Larix gmelinii forest during early winter [J]. Journal of Northeast Forestry University, 2020, 48(6): 152 − 156, 162. [20] DI Tong, LI Zhongwu, XIAO Haibin, et al. How do soil microbes exert impact on soil respiration and its temperature sensitivity? [J]. Environmental Microbiology, 2021, 23(6): 3048 − 3058. [21] MAZZA G, AGNELLI A E, CANTIANI P, et al. Short-term effects of thinning on soil CO2, N2O and CH4 fluxes in Mediterranean forest ecosystems [J]. Science of the Total Environment, 2019, 651: 713 − 724. [22] AKBURAK S, MAKINECI E. Thinning effects on soil and microbial respiration in a coppice-originated Carpinus betulus L. stand in Turkey [J]. Iforest-Biogeosciences and Forestry, 2016, 9: 783 − 790. [23] ZHAO Bo, CAO Jing, GENG Yan, et al. Inconsistent responses of soil respiration and its components to thinning intensity in a Pinus tabuliformis plantation in northern China [J]. Agricultural and Forest Meteorology, 2019, 265: 370 − 380. [24] CHENG Xiaoqin, HAN Hairong, ZHU Jiang, et al. Forest thinning and organic matter manipulation drives changes in soil respiration in a Larix principis-rupprechtii plantation in China[J/OL]. Soil and Tillage Research, 2021, 211: 104996[2023-03-01]. doi: 10.1016/j.still.2021.104996. [25] CHENG Xiaoqin, KANG Fengfeng, HAN Hairong, et al. Effect of thinning on partitioned soil respiration in a young Pinus tabulaeformis plantation during growing season [J]. Agricultural and Forest Meteorology, 2015, 214: 473 − 482. [26] TEMPLETON B S, SEILER J R, PETERSON J A, et al. Environmental and stand management influences on soil CO2 efflux across the range of loblolly pine [J]. Forest Ecology and Management, 2015, 355: 15 − 23. [27] 高明, 朱玉杰, 董希斌. 采伐强度对大兴安岭用材林土壤化学性质的影响[J]. 东北林业大学学报, 2013, 41(12): 39 − 41,76. GAO Ming, ZHU Yujie, DONG Xibin. Effects of tending felling on soil chemical property of timber forest in Daxing’an Mountain [J]. Journal of Northeast Forestry University, 2013, 41(12): 39 − 41,76. [28] ZHANG Xinzhong, GUAN Dexin, LI Weibin, et al. The effects of forest thinning on soil carbon stocks and dynamics: a meta-analysis [J]. Forest Ecology and Management, 2018, 429: 36 − 43. [29] LEI Lei, XIAO Wenfa, ZENG Lixiong, et al. Effects of thinning intensity and understory removal on soil microbial community in Pinus massoniana plantations of subtropical China [J/OL]. Applied Soil Ecology, 2021, 167: 104055[2023-03-01]. doi: 10.1016/j.apsoil.2021.104055. [30] XU Ming, SHANG Hua. Contribution of soil respiration to the global carbon equation [J]. Journal of Plant Physiology, 2016, 203: 16 − 28. [31] HU Shuaidong, LI Yongfu, CHANG S X, et al. Soil autotrophic and heterotrophic respiration respond differently to land-use change and variations in environmental factors[J]. Agricultural and Forest Meteorology, 2018, 250/251: 290 − 298. [32] TANG Xiaolu, DU Jie, SHI Yuehong, et al. Global patterns of soil heterotrophic respiration: a meta-analysis of available dataset[J/OL]. Catena, 2020, 191: 104574[2023-03-01]. doi: 10.1016/j.catena.2020.104574. [33] SUN Yarong, WANG Yajuan, ZHAO Min, et al. Response of soil respiration rates to soil temperature and moisture at different soil depths of Caragana korshinskii plantation in the Loess-Hilly region [J]. Environmental Science, 2022, 43(10): 4648 − 4657. [34] 范志平, 王红, 邓东周, 等. 土壤异养呼吸的测定及其温度敏感性影响因子[J]. 生态学杂志, 2008, 27(7): 1221 − 1226. FAN Zhiping, WANG Hong, DENG Dongzhou, et al. Measurement methods of soil heterotrophic respiration and key factors affecting the temperature sensitivity of the soil heterotrophic respiration [J]. Chinese Journal of Ecology, 2008, 27(7): 1221 − 1226. -

-
链接本文:
https://zlxb.zafu.edu.cn/article/doi/10.11833/j.issn.2095-0756.20230193
 点击查看大图
点击查看大图
计量
- 文章访问数: 1745
- HTML全文浏览量: 528
- PDF下载量: 56
- 被引次数: 0
