




-
随着工业化进程的加快,重金属污染成为中国最主要的环境污染问题之一。据统计,受铅镉等典型重金属污染的耕地达0.1亿hm2,由此造成的经济损失超过200亿元·a-1[1];土壤资源地区分布不均,由成土和耕作方式造成的有机质含量差异及对污染物的清除降解差异,使得南方红壤区的重金属污染状况更加严重[2-3]。重金属污染具有隐蔽性和富集性,既难以被微生物降解,又会在进入食物链后放大危害效应。近年来,由于土壤重金属浓度较高引发的“重金属超标”事件频发[4-5]。目前有效清除土壤中重金属污染物的方法如物理稀释、化学和生物降解修复等措施,因费用高、操作复杂、修复时间长、随土壤环境的改变变化较大,去除效果有限而较难实施[6-8],因此寻求具有高效吸附能力、能调节土壤理化生物环境、造价相对较低、对土壤扰动小的材料逐渐成为土壤重金属污染修复领域的热点。生物质炭是有机生物质(如农林废弃物、动植物组织等)在限氧或绝氧条件下经过高温热解炭化而成的一类富碳型产物,来源广泛,稳定性能好,具有丰富的孔隙、表面官能团和表面活性物质[9],已被证实能通过静电、阳离子交换和络合等作用吸附固定土壤中的重金属,较一般的活性炭,重金属选择性更强,二次污染较小,是理想的新型土壤污染修复材料[10-12]。目前,植物源生物质炭对土壤重金属污染修复的研究较多,动物源生物质炭的研究则主要集中在对土壤性质和养分改良作用上,对其吸附土壤中重金属污染物的研究较为有限[13-15]。中国畜牧资源充足,按3%的正常死亡率,病死猪量多达2 000万头·a-1。高温热解制备成生物质炭实现了病死动物集约无害资源化处理,同时猪炭含有较高的矿质元素和磷[16],再以生物质炭形式施用于重金属污染土壤后可实现土壤品质的边修复边提高,达到提高土壤生产力的目的,符合食品安全和环境可持续发展的要求。本研究以病死猪为原料制备生物质炭,以铅离子(Pb2+)为目标重金属,考察土壤类型及不同添加量对土壤吸附Pb2+的影响,以期为动物源生物质炭修复土壤重金属污染的研究提供理论依据。
-
试验土壤采自浙江省杭州市临安区胡下塘村2块相邻菜园,土层深度为0~20 cm,成土过程相同;采集常年种植蔬菜的菜园土壤标记为熟化红壤,未被开垦种植的菜园土壤标记为新垦红壤,剔除植物残体和石子等杂物,自然风干后分别过10目和100目筛,后用于吸附试验和理化性质表征。土壤pH值和土壤电导率(EC)按m(土):v(水)=1:5混合后测定;土壤阳离子交换量(CEC)使用乙酸铵浸提法测定;土壤有机质含量用重铬酸钾容量法测定[17]。供试土壤理化性质见表 1。
表 1 供试土壤基本理化性状
Table 1. Physical and chemical properties of the soils
土壤类型 pH 电导率/(dS·m-1) 阳离子交换量/(cmol·kg-1) w有机碳/(g·kg-1) 不同粒径颗粒占土壤的质量比 砂粒/% 粉粒/% 黏粒/% 熟化红壤 6.0 0.3 5.1 65.5 38.6 45.0 16.0 新垦红壤 5.8 0.2 4.1 10.3 38.7 44.4 15.8 -
病死猪由浙江省湖州市某病死动物无害化集中处理中心提供。将整猪置于炭化炉中,650 ℃无氧、常压条件下热解4 h制备而得,标记为猪炭(pig biochar,PB);研磨粉碎后过20目和100目筛。生物质炭pH,EC和CEC采用IBI标准方法[17]测定;交换性阳离子采取乙酸钠浸提法测定[18];灰分采用ASTM D1762-84法测定[19];交换性钙、镁采用原子吸收分光光度计(FAAS-6880,日本)测定;生物质炭比表面积采用比表面分析仪(ASAP 2020M,美国)测定,并采用BET法测算生物质炭比表面积、BJH法测算生物质炭孔容和平均孔径;采用扫描电子显微镜(SEM-EDS)(S-3000N,日本)分析生物质炭的元素组成与表面形态结构;碳(C)、氢(H)、氮(N)元素质量分数采用元素分析仪(Flash EA1112 Thermo Finnigan,意大利)测定,差减法计算氧(O)元素含量;表面碱度采用返滴定法[20]测定;用氢氟酸-高氯酸-硝酸(HF-HCIO4-HNO3)法对生物质炭进行消煮,重金属质量分数采用原子吸收分光光度法[21]测定、全磷质量分数采用钼蓝比色法[17]测定。检测得到猪炭pH值为10.25;阳离子交换值为31.14 cmol·kg-1,其中交换性钾(K)、钙(Ca)、镁(Mg)分别为1.33,0.19,0.95 cmol·kg-1;灰分含量为58.42%,元素碳(C)、氢(H)、氧(O)、氮(N)、磷(P)、硫(S)质量分数分别为28.87,1.56,7.77,3.26,7.70和0.12 mg·g-1;比表面积为28.23 m2·g-1,孔容为0.12 cm3·g-1,平均孔径为3.78 nm。扫描电镜(图 1A)及能谱图(图 1B)如图 1所示。

图 1 猪炭电镜扫描图和表面某位点能量色散X射线下元素组成分析(SEM-EDS)
Figure 1. Scanning electron microscope images and elemental composition of certain loci on surface of pig biochar
-
按照m(猪炭):m(供试土壤)=0:100,1:99,5:95准确称取相应质量的生物质炭(20目筛)和土壤(10目筛),混合均匀得到试验所需吸附剂;供试吸附剂pH值和CEC值测定同1.1.1。
-
试验以未添加生物质炭的熟化红壤(ckH)和未添加生物质炭的新垦红壤(ckL)为对照组,以添加质量分数为1%猪炭的熟化红壤(PBH1)、添加5%猪炭的熟化红壤(PBH5)、添加1%猪炭的新垦红壤(PBL1)、添加5%猪炭的新垦红壤(PBL5)为处理组,各重复3次。各样品准确称取1 g,置于50 mL塑料离心管中。
以去离子水配置0.01 mol·L-1的硝酸钠(NaNO3)背景溶液,将硝酸铅[Pb(NO3)2]稀释成100,200,400,600,800 mg·L-1的Pb2+标准溶液,用0.10 mol·L-1硝酸钠和0.10 mol·L-1氢氧化钠调节标准溶液pH值至5.5后,量取20 mL分别加入装吸附剂的离心管中。于(25±1)℃恒温震荡箱中以180 r·min-1连续振荡24 h后,取出经4 000 r·min-1离心8 min;取上清液过0.45 μm水系滤膜,通过原子吸收光谱仪(ICE3300,美国)测定Pb2+质量浓度。
-
单位质量吸附容量qe(mg·g-1)可有效表征土壤颗粒对重金属离子的吸附能力。计算公式为:qe=v(c0-c1)/m。Langmuir模型方程[22]可以表示为:ce/qe=ce/qm+1/qm×kL。其中:m为土壤质量(g);v为溶液体积(mL);c0为吸附初始溶液质量浓度(mg·L-1);c1为吸附平衡溶液质量浓度(mg·L-1);ce代表吸附平衡时溶液中重金属离子质量浓度(mg·L-1);qm代表土壤对重金属离子的最大吸附量(mg·g-1);kL与吸附强度和吸附量有关(mg·L-1),表征吸附剂对金属的单位吸附能力。
-
采用Excel 2010对数据处理并作图,利用SPSS 17.0对数据进行差异性和显著性分析,数据为平均数±标准差,不同字母表示95%以上显著差异。
-
由表 2可知:所有处理下熟化红壤pH和CEC均大于新垦红壤;施用猪炭后,土壤pH和CEC随生物质炭质量分数增大均显著升高(P<0.05)。ckL的pH为5.80,CEC为4.10 cmol·kg-1,均为各处理中最低,PBH5的pH为6.48,CEC为13.34 cmol·kg-1,均为各处理中最高。相较于对照,新垦红壤pH提高了0.11~0.42,CEC提高了2.83~7.23 cmol·kg-1,熟化红壤pH提高了0.18~0.48,CEC提高了3.23~8.26 cmol·kg-1。质量分数1%和5%的猪炭添加,使熟化红壤pH提高3.0%和8.0%,新垦红壤pH提高2.0%和7.2%;熟化红壤CEC提高69.0%和176.3%,新垦红壤CEC提高63.3%和162.0%,由此可知,猪炭可显著提高熟化红壤pH(P<0.05),显著提高新垦红壤CEC(P<0.05)。
表 2 不同质量分数猪炭对土壤pH和CEC的影响
Table 2. Effect of pig biochar on soil pH and CEC
处理 pH CEC/(cmol·kg-1) ckH 6.01±0.01 d 5.10±0.13 e BPH1 6.18±0.02 c 8.33±0.40 c BPH5 6.48±0.02 a 13.37±0.25 a ckL 5.80±0.13 f 4.10±0.05 f BPL1 5.91±0.05 e 6.93±0.15 d BPL5 6.22±0.04 b 11.33±0.25 b 说明:不同小写字母表示处理间差异显著(P < 0.05) -
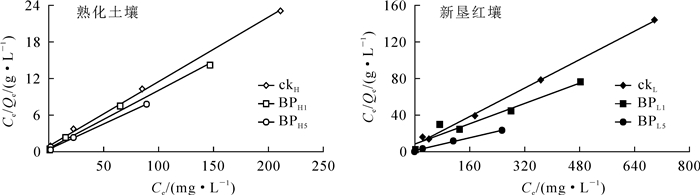
采用Langmuir等温吸附方程对土壤吸附Pb2+的数据进行线性拟合,模型假设吸附剂表层均质,分子质子间无互斥力,吸附容量随吸附点位增多而增大,与吸附剂表面特性密切相关[23]。由图 2可知:Langmuir等温吸附模型拟合相关性较好,拟合相关系数R2接近1,表明25 ℃时,土壤对Pb2+的吸附为吸附剂表层吸附点位和Pb2+相互结合的过程,以化学结合为主,表面分子质子间作用力相同;随着反应进行,吸附量逐渐增大,直至饱和吸附。相比而言,熟化红壤较新垦土壤的吸附量更大。

图 2 土壤对Pb2+的吸附等温线
Figure 2. Adsorption isotherms of Pb2+ in soil fitted with Langmuir model
从表 3可以看出:各处理下熟化红壤对Pb2+的吸附能力均大于新垦红壤。就对照而言,熟化红壤的Pb2+吸附容量是新垦红壤的2.21倍;添加猪炭后,2种土壤的吸附能力均上升,并随施用量的增加而增加,但在提高效果上呈现一定差异。当猪炭施加量为1%时,熟化红壤和新垦红壤对Pb2+吸附能力分别提高了1.21倍和1.40倍,施加量为5%时,熟化红壤和新垦红壤对Pb2+吸附能力分别提高了1.28倍和2.24倍。因此,施用质量分数5%的猪炭能显著提高土壤对Pb2+的吸附性能,相较于有机质含量较高的熟化红壤,在新垦红壤中施加猪炭,吸附Pb2+性能提升效果更明显。
表 3 土壤对Pb2+的吸附等温模型参数
Table 3. Adsorption parameters of Pb2+ in soils
处理 Langmuir参数 R2 kL/(mg·L-1) qm/(mg·g-1) ckH 0.10 11.33 ± 0.10 c 0.998 9 BPH1 0.13 12.71 ± 0.08 b 0.992 6 BPH5 0.19 14.49 ± 0.09 a 0.995 9 ckL 0.03 5.12 ± 0.02 e 0.998 1 BPL1 0.02 7.15 ± 0.03 d 0.938 5 BPL5 0.07 11.45 ± 0.11 c 0.996 6 说明:不同小写字母表示处理间差异显著P<0.05 -
土壤pH和CEC是表征土壤性质的重要指标,与土壤肥力和重金属吸附能力密切相关[24]。猪炭含有较多的碱土金属元素和无机盐矿物组分,灰分含量达50%以上,且含有较多的交换性钾、钙、镁等阳离子,因此pH和CEC均较高,碱性较强。张政[25]发现:在铅镉污染土壤中施加猪肉骨炭,土壤pH和CEC显著提高,与本研究一致。土壤pH的变化与生物质炭的碱性物质含量呈线性相关,后者富含的有机官能团,有助于提高土壤对氢离子(H+)的吸附,提高土壤的碱基饱和度[26],是添加猪炭后土壤pH提高的原因。研究证实,土壤有机质及黏粒是引起土壤CEC变化的重要原因,施入土壤的猪炭,因含有能与土壤胶体和颗粒结合并相互作用的矿物,在提高土壤矿物含量的同时,也提高了土壤有机碳含量,促进了土壤对阳离子的吸附[27],从而提高土壤-炭混合物的阳离子交换总量。
-
土壤吸附Pb2+性能受土壤有机质含量、pH和阳离子交换量的影响[28]。红壤中的有机质可加速外源Pb2+的固定[29],原因在于有机质含有的腐殖酸和含氧官能团,其大量的中性和极性亲水性基团可有效提高有机质表面的吸附活性,较一般土壤胶体具有更丰富的吸附位点,更有助于土壤对重金属离子的富集。较新垦红壤,熟化红壤CEC更高[28],有机碳更加丰富,Pb2+更容易被固定,从而有利于吸附反应的进行。章明奎等[30]发现土壤有机碳吸附Pb2+的最佳pH为5~6,本研究中,熟化红壤pH为6.0,新垦红壤pH为5.8,均在有机碳的较佳吸附性能范围内;相比之下,熟化红壤pH更高,重金属元素在氧化物表面的专性吸附作用更强,有机物与重金属能形成更加稳固的络合化合物,土壤有机质-金属配合物的稳定性更好[31]。本研究熟化红壤和新垦红壤有机碳含量相差较大,也说明较高的pH和有机碳含量是熟化红壤有更高Pb2+吸附能力的原因。
-
生物质炭对生物质原材料中的矿物及灰分特征[32]保留较为完整,尤其是动物骨炭,含有的钙和磷等矿物组分较高,通常以磷酸盐矿物和羟基磷灰石等形式存在,而这些成分是吸附Pb2+的重要组分[33];其中的磷酸盐可降低Pb2+的生物有效性,形成生物有效性极低的难溶性磷矿物,在农田土壤施肥管理和吸附去除重金属污染物上被广泛应用[34]。能谱分析发现,650 ℃高温热解后,猪炭中磷、钙等矿质元素含量依然较高(图 1),猪炭施入土壤时与Pb2+形成的磷酸盐与碳酸盐沉淀增多,有利于对Pb2+的吸附固定。
猪炭较高的pH和较多交换性钾、钙、镁离子,可在其施入酸性土壤后为土壤提供更多的碱性物质,增加土壤可交换性阳离子数量,提高土壤pH和CEC(图 2)。猪炭的施入,使得土壤中可与Pb2+交换的阳离子数目增多,土壤矿物表面与金属离子间的静电作用增强;在土壤矿物以及土壤中其他组分(如土壤溶解性有机碳)作用下,土壤-生物质炭体系的电荷和金属离子产生变化,提高了土壤对Pb2+的吸附性能[35]。REES等[36]指出:随着pH的升高,吸附剂对重金属离子的吸附量明显上升,越接近临界pH,吸附量上升越缓慢甚至下降。研究发现,新垦红壤有机质含量较低,添加猪炭后,pH提高,CEC相对增加量较熟化红壤更多[27],使得表面吸附点位的增多速率更快,这可能是新垦红壤在添加猪炭后对Pb2+吸附性能提高效果大于熟化红壤的原因。
高温热裂解造成有机质和水分挥发,使电镜下观察到的猪炭外表形貌呈不均匀褶皱和粗糙块状结构;受高温影响,有机质分解生成的气体逃逸后在猪炭表面留下较多大小不等、开口不完善的孔隙,其分布、孔径及构造不仅影响重金属离子的扩散,而且扩大其内外表面积,为吸附提供了更多的吸附位点[37],可认为是施用生物质炭提高土壤吸附能力的原因之一。此外,凸起于猪炭表面的大量形状不一的白色絮状颗粒物质可能为矿质元素、灰分及其相关的无机盐矿物呈现的矿物结晶形态[38],因此添加含磷矿物组分较多的猪炭,为土壤固定Pb2+形成沉淀提供了较多的活性吸附点位[39],从而提高了土壤对重金属的吸附能力;与植物源生物炭相比,施用动物源生物质炭对土壤有机碳含量、pH均有较为显著的提升效果,为Pb2+碳酸盐结合态和有机结合态的形成和提高提供了必要条件和有利环境[40]。
-
Langmuir等温吸附模型描述了Pb2+浓度变化对土壤吸附重金属Pb2+的影响过程,经过650 ℃热解后,猪炭较高的灰分、pH、磷元素和丰富的表面离子,是施用猪炭提高土壤对Pb2+吸附性能的关键。
土壤吸附重金属Pb2+的性能同土壤类型和猪炭施用量密切相关。土壤对Pb2+吸附性能随施加量增加而增强,且对新垦红壤吸附性能提高效果更好。
施加猪炭后,土壤pH和CEC升高,对Pb2+吸附钝化效果增强,且随施用量的增加而增加。施用质量分数5%的猪炭对土壤的吸附性能提升更加有效,理论上在降低田间土壤重金属污染物浓度方面的应用价值更高。
Dead pig-derived biochar treatments and soil adsorption of Pb2+
-
摘要: 利用650℃高温炭化炉热裂解病死猪,制成生物质炭(猪炭);通过批处理恒温振荡平衡法探索施用不同质量分数(0,1%和5%)的猪炭对熟化红壤和新垦红壤吸附溶液中铅离子(Pb2+)的影响。结果表明:施用猪炭能显著提高土壤pH值和阳离子交换量(P < 0.05);土壤对Pb2+的吸附量随猪炭施用量的增加而增大,施用猪炭的熟化红壤吸附容量为12.71~14.49 mg·g-1,较未施加猪炭对照提高了12.2%~27.9%;施用猪炭的新垦红壤吸附容量为7.15~11.45 mg·g-1,较未施加猪炭对照提高了39.7%~123.8%,说明对有机质质量分数较低的新垦红壤施加猪炭,土壤吸附Pb2+性能的提高效果更明显。与未施猪炭的对照相比,施加质量分数为1%的猪炭时,熟化红壤和新垦红壤对Pb2+吸附能力分别提高1.21倍和1.40倍,施加量为5%时,熟化红壤和新垦红壤对Pb2+吸附能力分别提高1.28倍和2.24倍。由此认为猪炭有助于土壤对Pb2+的吸附和固定,施用质量分数5%的猪炭能有效提高土壤对Pb2+的吸附。Abstract: A laboratory study was conducted to evaluate the effects of dead pig biochar (PB) on adsorption of lead ions (Pb2+) in two soils with contrasting properties. A batch equilibration method[(25±1)℃, 180 r·min-1, 24 h] was used to investigate the adsorption of Pb2+ on soils amended with biochar derived from dead pigs pyrolyzed at a peak temperature of 650℃. Two soils, including a vegetable garden soil with high organic matter content and an uncultivated soil with low organic matter content, which were adjacent in village in Lin'an, the experimental samples(0-20 cm) were chosen randomly, natural air drying and sifting, were treated with biochars mixed at three application rates (0, 1%, and 5%, w/w), 6 treatments in all, 3 repetitions in each treatment, soils without biochar were taken as the control. ANOVA and LSD (least significant difference) were used to statistic analysis. Langmuir isotherms were used to describe the results of adsorption processes. Results showed that both soil pH and cation exchange capacity values significantly (P < 0.05) increased with the biochar treatments. The Pb2+ adsorption capacity(Qm) increased in both soils when they were amended with biochars, and significantly increased (P < 0.05) with the increase of biochar dosage. In comparison to the control, Qm values of the soil with high organic matter content ranged from 12.71 to 14.49 mg·g-1 and increased 12.2%-27.9%; whereas, the Qm values of the soil with low organic matter content ranged from 7.15 to 11.45 mg·g-1 and increased 39.7%-123.8%. When 1% pig biochar was applied to soils, adsorption capacity of Pb2+ in the high organic matter content soil increased 1.21 times and in the low organic matter soil increased 1.40 times; when 5% of pig biochar was applied in the two types of soils, the Qm increased 1.28 times in the high organic matter content soil and 2.24 times in the low organic matter soil. Therefore, pig biochar was a suitable adsorption material for Pb2+ immobilization in soils, particularly at the 5% application rate.
-
Key words:
- soil science /
- pig biochar /
- heavy metal /
- contaminated soil /
- immobilization
-
图 1 猪炭电镜扫描图和表面某位点能量色散X射线下元素组成分析(SEM-EDS)
Figure 1 Scanning electron microscope images and elemental composition of certain loci on surface of pig biochar
图 2 土壤对Pb2+的吸附等温线
Figure 2 Adsorption isotherms of Pb2+ in soil fitted with Langmuir model
表 1 供试土壤基本理化性状
Table 1. Physical and chemical properties of the soils
土壤类型 pH 电导率/(dS·m-1) 阳离子交换量/(cmol·kg-1) w有机碳/(g·kg-1) 不同粒径颗粒占土壤的质量比 砂粒/% 粉粒/% 黏粒/% 熟化红壤 6.0 0.3 5.1 65.5 38.6 45.0 16.0 新垦红壤 5.8 0.2 4.1 10.3 38.7 44.4 15.8  下载: 导出CSV
下载: 导出CSV
表 2 不同质量分数猪炭对土壤pH和CEC的影响
Table 2. Effect of pig biochar on soil pH and CEC
处理 pH CEC/(cmol·kg-1) ckH 6.01±0.01 d 5.10±0.13 e BPH1 6.18±0.02 c 8.33±0.40 c BPH5 6.48±0.02 a 13.37±0.25 a ckL 5.80±0.13 f 4.10±0.05 f BPL1 5.91±0.05 e 6.93±0.15 d BPL5 6.22±0.04 b 11.33±0.25 b 说明:不同小写字母表示处理间差异显著(P < 0.05)
下载: 导出CSV
表 3 土壤对Pb2+的吸附等温模型参数
Table 3. Adsorption parameters of Pb2+ in soils
处理 Langmuir参数 R2 kL/(mg·L-1) qm/(mg·g-1) ckH 0.10 11.33 ± 0.10 c 0.998 9 BPH1 0.13 12.71 ± 0.08 b 0.992 6 BPH5 0.19 14.49 ± 0.09 a 0.995 9 ckL 0.03 5.12 ± 0.02 e 0.998 1 BPL1 0.02 7.15 ± 0.03 d 0.938 5 BPL5 0.07 11.45 ± 0.11 c 0.996 6 说明:不同小写字母表示处理间差异显著P<0.05
下载: 导出CSV
-
[1] 周建军, 周桔, 冯仁国.我国土壤重金属污染现状及治理战略[J].中国科学院院刊, 2014, 29(3):315-320. ZHOU Jianjun, ZHOU Ju, FENG Renguo. Status of China's heavy metal contamination in soil and its remediation strategy[J]. Bull Chin Acad Sci, 2014, 29(3):315-320. [2] 赵其国, 黄国勤, 马艳芹.中国南方红壤生态系统面临的问题及对策[J].生态学报, 2013, 33(24):7615-7622. ZHAO Qiguo, HUANG Guoqin, MA Yanqin. The problems in red soil ecosystem in southern of China and its countermeasures[J]. Acta Ecol Sci, 2013, 33(24):7615-7622. [3] 陈世宝, 华珞, 白玲玉, 等.有机质在土壤重金属污染治理中的应用[J].农业环境与发展, 1997, 53(3):26-29, 48. CHEN Shibao, HUA Luo, BAI Lingyu, et al. Application of organic matters in the improvement of soil polluted by heavy metals[J]. Agro-Environ Dev, 1997, 53(3):26-29, 48. [4] 李小平, 刘洁, 夏舒楠, 等.我国城市土壤铅与儿童血铅空间分布格局[J].土壤通报, 2015, 46(1):226-232. LI Xiaoping, LIU Jie, XIA Shunan, et al. Spatial distribution pattern of lead in urban soil and in children's blood, Chin[J]. Chin J Soil Sci, 2015, 46(1):226-232. [5] 夏星辉, 陈静生.土壤重金属污染治理方法研究进展[J].环境科学, 1997, 18(3):72-76, 95. XIA Xinghui, CHEN Jingsheng. Advances in the study of remediation methods of heavy metal-contaminated soil[J]. Chin J Environ Sci, 1997, 18(3):72-76, 95. [6] 徐东昱, 周怀东, 高博.生物炭吸附重金属污染物的研究进展[J].中国水利水电科学研究院学报, 2016, 14(1):7-15. XU Dongyu, ZHOU Huaidong, GAO Bo. Review of sorption of heavy metal contaminant on biochar[J]. J Chin Ins Water Conserv Hydroelectric Sci, 2016, 14(1):7-15. [7] 刘少文, 焦如珍, 董玉红, 等.土壤重金属污染的生物修复研究进展[J].林业科学, 2017, 53(5):146-155. LIU Shaowen, JIAO Ruzhen, DONG Yuhong, et al. Research progress in bioremediation of heavy-metal contaminated soil[J]. For Sci, 2017, 53(5):146-155. [8] ZHANG Xiaokai, WANG Hailong, HE Lizhi, et al. Using biochar for remediation of soils contaminated with heavy metals and organic pollutants[J]. Environ Sci Pollut Res, 2013, 20(12):8472-8483. [9] SOHI S P, KRULL E, LOPEZ-CAPEL E. A review of biochar and its use and function in soil[C]//SPARKS D L. Advances in Agronomy. London: Elsevier, 2010: 47-82. [10] NOVAK J M, BUSSCHER W J, LAIRD D L, et al. Impact of biochar amendment on fertility of a southeastern coastal plain soil[J]. Soil Sci, 2009, 174(2):105-112. [11] 肖瑶, 葛成军, 张丽, 等.木薯渣基生物质炭对水中Cd2+、Cu2+的吸附行为研究[J].农业环境科学学报, 2016, 35(8):1587-1594. XIAO Yao, GE Chengjun, ZHANG Li, et al. Adsorption performance of Cd2+ and Cu2+ in aqueous solution by biochars prepared from manioc wastes[J]. J Agro-Environ Sci, 2016, 35(8):1587-1594. [12] 刘晶晶, 杨兴, 陆扣萍, 等.生物质炭对土壤重金属形态转化及其有效性的影响[J].环境科学学报, 2015, 35(11):3679-3687. LIU Jingjing, YANG Xing, LU Kouping, et al. Effect of bamboo and rice straw biochars on the transformation and bioavailability of heavy metals in soil[J]. Acta Sci Circumstantiae, 2015, 35(11):3679-3687. [13] 林雪原, 荆延德, 巩晨, 等.生物炭吸附重金属的研究进展[J].环境污染与防治, 2014, 36(5):83-87. LIN Xueyuan, JING Yande, GONG Chen, et al. Research progress on the sorption of heavy metals by biochar[J]. Environ Pollut Control, 2014, 36(5):83-87. [14] 郭茜, 陆扣萍, 胡国涛, 等.死猪炭和竹炭对菜地土壤理化性质和蔬菜产量的影响[J].浙江农林大学学报, 2017, 34(2):244-252. GUO Xi, LU Kouping, HU Guotao, et al. Greenhouse soil properties and vegetable yield with dead pig and bamboo biochars[J]. J Zhejiang A&F Univ, 2017, 34(2):244-252. [15] 王秋月, 杨兴, 何丽芝, 等.酸化猪炭对土壤中不同形态磷质量分数及相互转化的影响[J].浙江农林大学学报, 2018, 35(2):387-397. WANG Qiuyue, YANG Xing, HE Lizhi, et al. Concentration and fractionation of phosphorous in soils with acidulated pig biochar[J]. J Zhejiang A&F Univ, 2018, 35(2):387-397. [16] 金姝兰, 黄益宗, 胡莹, 等.骨炭、磷矿粉和生物调理剂对赣南稀土矿区土壤稀土元素淋溶的影响[J].环境科学学报, 2016, 36(10):3818-3825. JIN Shulan, HUANG Yizong, HU Ying, et al. Effects of bone char, phosphate rock and modifying agent on leaching of rare earth elements in soil[J]. J Environ Sci, 2016, 36(10):3818-3825. [17] 鲁如坤.土壤农业化学分析方法[M].北京:中国农业科技出版社, 2000. [18] International Biochar Initiative. Standardized product definition and product testing guidelines for biochar that is used in soil: 2012[S/OL].[2018-10-22]. http://walkingpointfarms.com/wp-content/uploads/2012/05/Guidelines_for_Biochar_That_Is_Used_in_Soil_Final.pdf. [19] ASTM International. Standard practice for coding plastic manufactured articles for resin identification: 2013[S/OL]. https://www.astm.org/COMMIT/d7611.pdf. [20] YUAN Jinhua, XU Renkou, ZHANG Hong. The forms of alkalis in the biochar produced from crop residues at different temperatures[J]. Bioresour Technol, 2011, 102(3):3488-3497. [21] 罗毅.地表水环境质量监测实用分析方法[M].北京:中国环境科学出版社, 2009. [22] HAGHSERESHT F, LU G Q. Adsorption characteristics of phenolic compounds onto coal-reject-derived adsorbents[J]. Energy Fuels, 1998, 12(6):1100-1107. [23] CLEMENTE R, BERNAL M P. Fractionation of heavy metals and distribution of organic carbon in two contaminated soils amended with humic acids[J]. Chemosphere, 2006, 64(8):1264-1273. [24] 郭鹏, 郭平, 康春莉, 等.城市土壤吸附重金属动力学特征及其与土壤理化性质的关系[J].环境保护科学, 2008, 34(6):23-26. GUO Peng, GUO Ping, KANG Chunli, et al. Adsorption kinetic characteristics of heavy metals in urban soils and the relationship with the physicochemical properties of soils[J]. Environ Prot Sci, 2008, 34(6):23-26. [25] 张政.肉骨生物炭对水体和土壤环境中铅镉修复效果的研究[D].上海: 华东理工大学, 2016. ZHANG Zheng. Immobilization of Pb2+ and Cd2+ in Contaminated Waters and Soils by Biochars Derived from Meat and Bone Meal[D]. Shanghai: East China University of Science and Technology, 2016. [26] XU Renkou, ZHAO Anzhen, YUAN Jinhua, et al. pH buffering capacity of acid soils from tropical and subtropical regions of China as influenced by incorporation of crop straw biochars[J]. J Soils Sediments, 2012, 12(4):494-502. [27] 黄超, 刘丽君, 章明奎.生物质炭对红壤性质和黑麦草生长的影响[J].浙江大学学报(农业与生命科学版), 2011, 37(4):439-445. HUANG Chao, LIU Lijun, ZHANG Mingkui. Effects of biochar on properties of red soil and ryegrass growth[J]. J Zhejiang Univ Agric Life Sci, 2011, 37(4):439-445. [28] PHILLIPS I R. Copper, lead, cadmium, and zinc sorption by waterlogged and air-dry soil[J]. J Soil Contam, 1999, 8(3):343-364. [29] 吴曼, 徐明岗, 徐绍辉, 等.有机质对红壤和黑土中外源铅镉稳定化过程的影响[J].农业环境科学学报, 2011, 30(3):461-467. WU Man, XU Minggang, XU Shaohui, et al. Effects of organic matter on the stabilization process of added cadmium and lead in red soil and black soil[J]. J Agro-Environ Sci, 2011, 30(3):461-467. [30] 章明奎, 郑顺安, 王丽平.土壤中颗粒状有机质对重金属的吸附作用[J].土壤通报, 2007, 38(6):1100-1104. ZHANG Mingkui, ZHENG Shun'an, WANG Liping. Adsorption of heavy metals by soil particulate organic matter[J]. Chin J Soil Sci, 2007, 38(6):1100-1104. [31] 李福燕, 李许明, 吴鹏飞, 等.海南省农用地土壤重金属含量与土壤有机质及pH的相关性[J].土壤, 2009, 41(1):49-53. LI Fuyan, LI Xuming, WU Pengfei, et al. Correlation between heavy metal pollution and basic properties of agricultural soils in Hainan Province[J]. Soils, 2009, 41(1):49-53. [32] 玛依拉·麦麦提热依木.羊骨炭制备条件的优化及其对酚类有机物吸附性能研究[D].乌鲁木齐: 新疆大学, 2015. Mahira Mamatriyim. Preparation and Modification of Sheep Bone Char and Its Adsorption Capacity for Phenols[D]. Urumqi: Xinjiang University, 2015. [33] 张金利, 刘大伟, 杨庆.骨炭对Pb(Ⅱ)的吸附特性[J].环境工程学报, 2014, 8(5):1784-1790. ZHANG Jinli, LIU Dawei, YANG Qing. Adsorption behavior of bone char to heavy metal Pb(Ⅱ)[J]. J Environ Eng, 2014, 8(5):1784-1790. [34] ZWONITZER J C, PIERZYNSKI G M, HETTIARACHCHI G M. Effects of phosphorus additions on lead, cadmium, and zinc bioavailabilities in a metal contaminated soil[J]. Water Air Soil Pollut, 2003, 143(1-4):193-209. [35] 刘荣琴, 钱林波, 晏井春, 等. pH及共存金属离子对生物质炭吸附铅稳定性的影响[J].土壤, 2017, 49(3):467-475. LIU Rongqin, QIAN Linbo, YAN Jingchun, et al. Effects of coexisting metal ions and pH on adsorption stability of lead on biochars[J]. Soils, 2017, 49(3):467-475. [36] REES F, SIMONNOT M O, MOREL J L. Short-term effects of biochar on soil heavy metal mobility are controlled by intra-particle diffusion and soil pH increase[J]. Eur J Soil Sci, 2014, 65(1):149-161. [37] 张玲, 詹肇麟, 张天栋, 等.高比表面积多孔炭质材料的孔结构特性及对苯酚的吸附[J].化工新型材料, 2017, 45(2):117-119. ZHANG Ling, ZHAN Zhaolin, ZHANG Tiandong, et al. Pore structure and phenol adsorption property of high specific surface area porous carbon materials[J]. New Chem Mater, 2017, 45(2):117-119. [38] ZHENG Hao, WANG Zhenyu, XIA Deng, et al. Characteristics and nutrient value of biochars produced from giant reed at different temperatures[J]. Bioresour Technol, 2013, 130(2):463-471. [39] YANG Hong, XU Ran, XUE Xiaoming, et al. Hybrid surfactant-templated mesoporous silica formed in ethanol and its application for heavy metal removal[J]. J Hazardous Mater, 2008, 152(2):690-698. [40] 林爱军, 张旭红, 苏玉红, 等.骨炭修复重金属污染土壤和降低基因毒性的研究[J].环境科学, 2007, 28(2):232-237. LIN Aijun, ZHANG Xuhong, SU Yuhong, et al. Chemical fixation of metals in soil using bone char and assessment of the soil genotoxicity[J]. Environ Sci, 2007, 28(2):232-237. -

-
链接本文:
https://zlxb.zafu.edu.cn/article/doi/10.11833/j.issn.2095-0756.2019.03.019
 点击查看大图
点击查看大图

计量
- 文章访问数: 4343
- HTML全文浏览量: 857
- PDF下载量: 62
- 被引次数: 0
