




-
土壤有机碳对大气温室气体的贡献主要来源于土壤活性有机碳的分解[1-3]。土壤活性碳是土壤碳库中能直接参与土壤生物化学过程、易被土壤微生物分解利用、对植物养分供应有直接作用的有机碳,虽然只占土壤总有机碳的很小比例,但是由于其易分解和周转快,能快速地反映外界干扰和环境因素对土壤有机质造成的微小变化[4-5]。因此,土壤活性有机碳常作为响应土地利用变化、植被演替过程、环境变化和气候变化等的敏感指标[6],对预测未来全球气候变化具有重要作用。微生物生物量碳(microbial biomass carbon,MBC)是活性有机碳库的一个重要组成部分,常用来表征土壤活性有机碳库的变化。土壤MBC的含量受土壤温度、pH、含水量、养分含量的共同影响,而这些因子通常随着季节变化而变化,因此土壤MBC的变化通常呈现一定的季节变化规律[7−9]。目前,关于土壤微生物生物量碳及其影响因素已开展了较多研究,主要集中在农田、森林、草地以及淡水湿地生态系统[10-13],而对于滨海湿地生态系统的相关研究还比较缺乏。在滨海湿地生态系统中,土壤MBC通常会受气候、土壤和植被等因子的影响。植被演替是滨海湿地形成和演变的重要标志,伴随着植被演替进程,物种组成发生变化,有机质来源的地表凋落物和地下根系分泌物类型也发生相应的变化,从而影响土壤中MBC含量[14-15]。江苏滨海湿地位于北亚热带向南暖温带的过渡地带,湿地面积达100 万hm2,其中淤泥质海滩面积约41.56 万hm2,是中国淤泥质滨海湿地的重要组成部分。随着滩涂的逐渐淤积抬升,水分和含盐量则逐渐降低,江苏滨海湿地的环境条件呈现出了有序变化,植被呈明显的条带分布。植被分布由海向陆分布为光滩、大米草Spartina anglica、碱蓬Suaeda glauca、芦苇Phragmites australis和刺槐Robinia pseudoacacia群落。对江苏滨海湿地不同演替阶段土壤MBC的质量分数和季节动态进行研究,对揭示滨海湿地植被演替对土壤碳库动态的影响以及科学预测滨海湿地土壤有机碳在大气碳循环中的作用都具有重要的意义。
-
研究区位于江苏北部,黄海和东海交汇处,江苏大丰国家级麋鹿Elaphurus davidianus保护区内(33°05′N,120°44′E)。该区地处北亚热带向暖温带的过渡地带,受海洋和大陆性气候的影响,具有明显的过渡性、海洋性和季风性。四季分明,气温适中,雨量充沛,年均气温为14.1 ℃,年均降水量为1 047.5 mm,降水多集中在夏季。大丰国家级麋鹿保护区为典型的淤泥质滩涂湿地,原始植被类型简单,植被演替序列相对完整,由海向陆植被演替阶段依次为光滩、大米草群落、碱蓬群落、芦苇群落和刺槐群落。
-
以空间代替时间的方法,在研究区沿与海岸线垂直的方向,自海向陆随机设置3条约30 m宽的样带,样带之间的空间分隔距离在300~500 m。在样带上自海向陆分布有光滩、大米草、碱蓬、芦苇和刺槐群落共5个演替阶段;同时,在每个对应的演替阶段分别设置3个5 m×5 m的样方,每个样方间距离不小于50 m。在每个样方内随机选取3个点挖取土壤剖面,按0~10、10~25和25~40 cm共3个土层进行分层采样。采集的土壤样品用自封袋装好标记后,迅速带回实验室,剔除肉眼可见的根系、动物、植物残体和石砾等,并将土壤样品分成2份。一份鲜土于4 ℃下冷藏,用于测定土壤MBC及土壤含水量;另一份土样经风干处理后,磨细过筛,用于土壤理化性质分析。
-
土壤MBC测定:采用氯仿熏-硫酸钾浸提法[6],熏蒸和未熏蒸的样品分别用0.5 mol·L−1的硫酸钾溶液浸提30 min,用岛津TOC-VCPH仪测定浸提液有机碳质量分数。土壤MBC质量分数wMBC
$= $ EC/0.45。其中:EC为熏蒸与未熏蒸土壤样品浸提液中有机碳质量分数的差值,mg·kg−1;0.45为MBC浸提系数。 -
采用LY/T 1210~1275−1999《森林土壤分析方法》[16]测定土样基本理化性质。重铬酸钾外加热法测定土壤总有机碳(SOC);元素分析仪(ElementarVarioEL,德国)测定总氮(TN);玻璃电极 (土水质量比为1.0∶2.5)测定土壤pH;烘干恒量法(105 ℃)测定土壤含水率;环刀法测定容重。
-
利用单因素和多因素方差分析方法分析植被演替、季节和土层因素对土壤MBC的影响;运用Pearson相关系数法分析MBC与土壤理化因子的相关性,数据分析软件为SPSS 20.0。
-
江苏滨海湿地不同演替阶段土壤MBC在0~40 cm土层的质量分数分别为:刺槐群落116.91~254.61 mg·kg−1,碱蓬群落168.12~276.19 mg·kg−1,芦苇群落219.45~290.76 mg·kg−1,大米草群落211.37~380.14 mg·kg−1,光滩117.00~326.18 mg·kg−1。平均质量分数从大到小依次为大米草、芦苇、光滩、碱蓬、刺槐群落,未表现出沿演替方向的变化趋势,其中碱蓬和刺槐群落显著低于其他3个演替阶段(图1)。0~10 cm土层,各演替阶段土壤MBC质量分数范围为157.66~380.14 mg·kg−1,在夏季、秋季和冬季从大到小均为大米草、芦苇、碱蓬、刺槐群落。其中,夏季大米草群落土壤MBC质量分数显著高于芦苇、碱蓬和刺槐群落;秋季,刺槐和碱蓬群落土壤MBC质量分数显著低于大米草群落和光滩;冬季,大米草和芦苇群落土壤MBC质量分数显著高于刺槐和碱蓬群落。10~25 cm土层,各演替阶段土壤MBC质量分数范围为140.73~357.12 mg·kg−1,土壤MBC质量分数在春季和夏季从大到小依次为芦苇、大米草、光滩、碱蓬、刺槐群落;秋季,刺槐和碱蓬群落土壤MBC质量分数均显著低于其他3个演替阶段。25~40 cm土层,各演替阶段土壤MBC质量分数范围为116.91~350.48 mg·kg−1;春季,刺槐林土壤MBC质量分数显著低于芦苇和大米草群落;秋季,刺槐群落显著低于其他演替阶段;冬季,刺槐和碱蓬群落土壤MBC质量分数显著低于大米草群落和光滩。
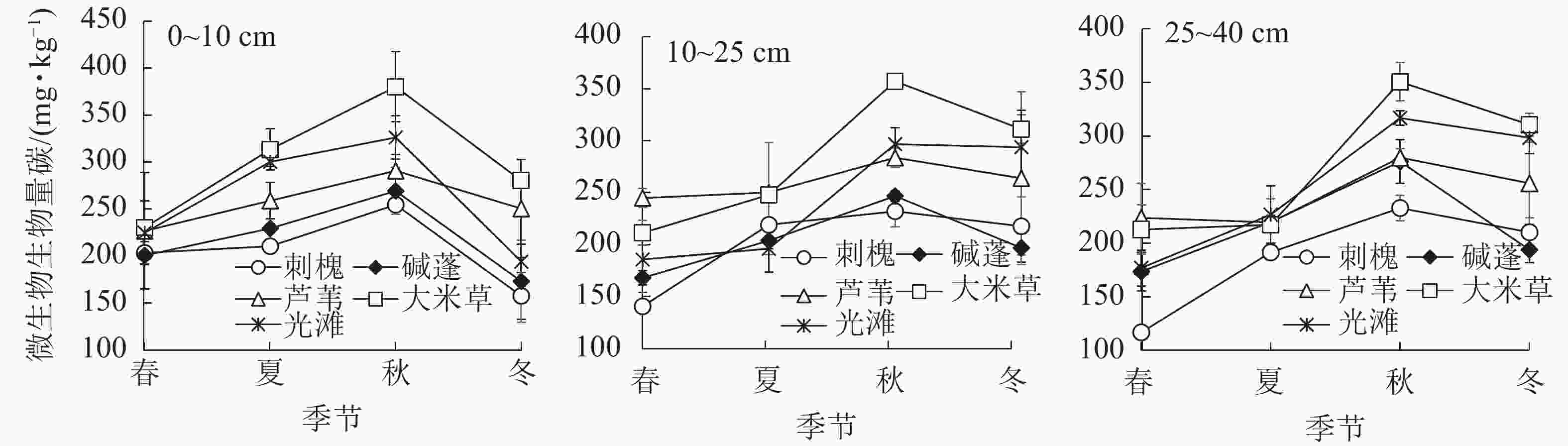
图 1 不同演替阶段土壤微生物生物量碳质量分数季节动态
Figure 1. Seasonal variations of soil MBC contents under different succession stages
-
从图1可以看出:不同演替阶段的土壤MBC质量分数在3个土层的季节变化趋势一致,均随季节先增加再下降,在秋季达到峰值;除芦苇群落外,各演替阶段土壤MBC质量分数均呈现出明显的季节变化规律。在0~10 cm土层,刺槐群落土壤MBC质量分数从大到小依次为秋季、夏季、春季、冬季;碱蓬群落土壤MBC质量分数在冬季显著低于夏季、秋季;大米草群落土壤MBC质量分数的季节变化表现为秋季最高,春季最低;光滩土壤MBC质量分数在春季、冬季显著低于夏季、秋季。10~25 cm土层,各演替阶段土壤MBC质量分数随季节的变化趋势与0~10 cm层一致,但是变化幅度不同;刺槐和碱蓬群落土壤MBC质量分数秋季较高,春季显著较低;大米草群落呈现出了明显的季节差异性,差异性从高到低依次为秋季、冬季、夏季、春季;光滩土壤MBC质量分数差异性表现为秋季和冬季显著高于春季和夏季。25~40 cm,各演替阶段土壤MBC质量分数的季节变化趋势与其他土层一致;刺槐群落土壤MBC质量分数的季节从高到低依次为秋季、冬季、夏季、春季;碱蓬群落土壤MBC质量分数在秋季最高,其他季节变化不明显;大米草群落和光滩土壤MBC质量分数在秋季和冬季显著高于春季和夏季。
-
利用多因素方差分析可得出:本研究中植被演替和季节对土壤MBC质量分数具有极显著影响(P<0.01),而土层深度对土壤MBC质量分数的影响较小且不显著(表1)。从3个影响因素间的交互作用看,季节分别与演替和土层的交互作用对土壤MBC质量分数均具有极显著影响(P<0.01)。表1中分布值可表示不同因素对土壤MBC质量分数的影响效应大小。根据分布值数据可知,季节因素对土壤MBC质量分数的影响最大,影响效应占32.29%;其次是演替因素的影响,效应占26.23%;季节和土层的交互作用对土壤MBC质量分数的影响效应占7.05%;季节与演替的交互作用对土壤MBC的影响效应相对较弱,占5.86%。
表 1 植被演替、土层和季节对土壤MBC质量分数的影响
Table 1. Effects of vegetation succession, soil layer and season on soil MBC contents
影响因素 分布值/% P 植被演替 26.23 0.000 土层 0.91 0.087 季节 32.29 0.000 植被演替×土层 0.96 0.728 植被演替×季节 5.86 0.003 土层×季节 7.05 0.000 植被演替×土层×季节 4.84 0.348 -
滨海湿地土壤理化性质也是影响土壤MBC质量分数的主要因素,本研究中滨海湿地土壤基本理化性质见文献[17]。本研究分析了不同演替阶段土壤MBC质量分数与土壤理化性质间的相关性(表2)。结果显示:土壤MBC质量分数与土壤总有机碳和总氮之间呈显著(P<0.05)的正相关关系;土壤pH与MBC质量分数之间呈负相关,且相关性达极显著水平(P<0.01);土壤MBC质量分数与土壤容重呈显著的负相关关系(P<0.05),而与土壤含水率表现出极显著的正相关关系(P<0.01)。本研究中C/N与土壤MBC质量分数的相关性不显著(P>0.05)。
表 2 土壤微生物生物量碳与土壤理化因子相关性
Table 2. Pearson correlation coefficients (r-value) between MBC and physic-chemical factors of soil
项目 土壤
有机碳总氮 C/N pH 容重 含水率 MBC 0.187* 0.173* 0.062 −0.225** −0.190* 0.461** 说明:*表示显著相关水平(P<0.05);**表示极显著相关水平 (P< 0.01) -
不同演替阶段由于植物群落不同,地表凋落物、地下根系分泌物以及土壤微生物种类也存在差异,从而影响土壤MBC的质量分数[18]。不同演替阶段土壤MBC质量分数在不同土层从高到低均依次为大米草群落、光滩或芦苇群落、碱蓬群落、刺槐群落。杨文英等[11]研究杭州湾滩涂湿地4种典型植被的活性有机碳质量分数发现:不同植被土壤MBC质量分数从高到低依次为光滩、海三棱藨草Scirpus mariqueter群落、互花米草Spartina alterniflora群落、芦苇群落;孔小琳等[19]研究胶州湾滨海湿地不同群落土壤微生物生物量碳变化得出从高到低依次为碱蓬群落、互花米草群落、芦苇群落、光滩,均与本研究的结果不一致。表明在不同区域环境下,由于土壤因子、气候因子以及凋落物等共同影响,土壤微生物有其独特的分布规律[20]。本研究中大米草群落的土壤MBC质量分数在不同土层均显著大于其他演替阶段。YANG等[21]研究了江苏盐城的互花米草、碱蓬、芦苇及光滩的土壤有机碳组分,同样发现互花米草土壤MBC质量分数较高。一方面,大米草作为一种入侵植物,具有较强的生产力和繁殖力,其大量的凋落物和地下根系分泌物促进了土壤有机碳输入,为土壤微生物提供了丰富的代谢底物,从而使土壤MBC的质量分数增加[22-24],这与本研究中土壤MBC与土壤有机碳质量分数具有正相关关系的结论一致;另一方面大米草群落所处的演替阶段容易受潮汐作用和干湿交替的影响,土壤含水率能够刺激土壤微生物活性,从而增加土壤MBC质量分数[25]。张静等[12]研究表明:土壤含水率是影响土壤MBC质量分数的一个重要因素,在一定范围内,土壤含水率高可增强植物根系的活动能力,促进大量根系分泌物的产生,从而使土壤MBC质量分数增加,这与本研究中土壤含水率较高的大米草群落具有较高的土壤MBC质量分数相符合。可见,土壤有机质、土壤含水率、土壤pH等土壤理化因子都是影响不同演替阶段土壤MBC质量分数变化的关键因子。
-
不同演替阶段的土壤MBC质量分数在3个土层均在秋季和冬季达到显著性差异。表明土壤MBC的动态变化是一个复杂过程,即使气候条件相同,不同植被下土壤MBC的季节变化也有差异。本研究中,除芦苇群落外,其他演替阶段土壤MBC质量分数均具有明显的季节变化规律,说明季节因素对不同演替阶段土壤MBC质量分数具有显著的影响。不同演替阶段不同土层土壤MBC质量分数的季节变化幅度不一致,但是均在秋季达到最大峰值,这与田舒怡等[18]的研究结果一致。土壤MBC质量分数在秋季最高,可能是由于秋季大量凋落物、衰老根系及碳水化合物由地上向地下转移,为微生物提供更多的能量[24];土壤MBC质量分数在植物开始生长的春季最低,可能是由于春季微生物活性以及凋落物分解速率较慢[26]。
-
植被演替、季节,以及植被演替和季节的交互作用均显著影响土壤MBC质量分数。植被演替对土壤MBC质量分数的影响主要是由于各演替阶段的植被组成不同,其凋落物质量和数量的差异会引起输入土壤的养分不同,进而影响土壤微生物活性;凋落物分解速率快,为土壤微生物提供的养分多,土壤微生物生物量则较大[27]。土壤MBC质量分数受季节与演替的交互影响,是由于季节变化通常会引起不同演替阶段植被生长节律和土壤微生物对养分吸收能力的改变,从而影响土壤MBC的质量分数[28]。本研究中土壤深度虽然对土壤MBC质量分数的影响不明显,但是土层与季节的交互作用却对土壤MBC质量分数存在显著影响,表明季节是影响土壤MBC的主导因子,且其影响在不同土层间存在差异。刘平等[26]研究渤海泥质海岸典型防护林土壤MBC季节动态,同样得到季节因素是影响土壤MBC质量分数变化的重要因素,且不同林分类型和不同土层,季节因素的影响作用存在差异。从植被演替、季节与土层及其之间的交互作用对土壤MBC质量分数的影响效应看,季节因素的影响效应所占比例最大,表明季节因素是影响滨海湿地土壤MBC质量分数最关键的因素。土壤MBC质量分数的季节动态是一个复杂的生物化学过程,可能受不同机制的驱动,季节变化通常能够引起地表凋落物、地下根系分泌物以及土壤有机质、温度、湿度、pH等理化因子的改变,从而综合影响土壤微生物活性,使土壤MBC质量分数表现出季节差异性[29]。本研究中土壤MBC质量分数随季节变化先增加后减少并在秋季达到最大值,这是由于冬季过渡到春季这段时间,温度上升缓慢,凋落物分解速率较慢,土壤中微生物可利用的有机质仍然较少,土壤MBC仍然较低[29];夏季雨水充足,土壤温湿度适宜,植物进入生长季,微生物活性逐渐增加,使土壤MBC增加[30];从夏季到秋季,虽然降雨量减少,土壤含水率下降,导致土壤微生物活性减弱,但是由于大量植物凋落物分解,为微生物提供了丰富的营养物质,使土壤中MBC继续增加[29];从秋季到冬季,随着气温的降低,微生物活性收到抑制,土壤中MBC减少[30]。
-
土壤性质是影响土壤MBC质量分数和分布的直接因素[26]。相关性分析显示:土壤MBC质量分数与土壤总氮和土壤有机碳均呈显著正相关,这与ZHOU等[31]的研究结果一致,表明土壤有机质是影响土壤MBC的重要因素,有机质含量高,能够为土壤微生物提供进行自身合成和代谢所需的碳、氮物质来源以及能量来源[32]。刘平等[26]同样得到渤海泥质海岸土壤MBC质量分数与土壤总氮和土壤有机碳具有极显著正相关关系的结论。也有部分研究结果显示:土壤MBC与土壤有机碳、全氮质量分数间的相关性不显著[32-33],研究结果的差异性可能是由于研究区空间尺度较大,受区域气候、土壤类型等因素的综合影响导致的[32]。本研究中土壤MBC质量分数与土壤pH、含水率成呈极显著相关,表明土壤pH和含水率都是影响滨海湿地土壤MBC质量分数的限制性因子[12]。土壤pH、含水率可直接影响土壤微生物活性,是调节土壤微生物活性的主要因子[34]。范志平等[35]研究辽东山地不同森林类型土壤有机碳季节动态及其驱动因子发现,土壤MBC质量分数与土壤pH呈显著线性正相关;张静等[12]和刘明慧等[24]研究发现:土壤含水率是土壤MBC等活性有机碳变化的主要驱动因子。
-
江苏滨海湿地土壤MBC质量分数在不同演替阶段的分布从高到低依次为大米草群落、芦苇群落、光滩、碱蓬群落、刺槐群落,其分布规律与演替方向不一致。不同演替阶段不同土层土壤MBC质量分数的季节动态均表现为随季节先增加再下降的趋势,且在秋季达到最大峰值。江苏滨海湿地土壤MBC质量分数在不同土层无显著差异,但是受季节和植被演替的影响较显著,其中季节因素的影响效应占较大比例。土壤理化因子是影响滨海湿地土壤MBC质量分数的直接因素,其中土壤有机碳、土壤总氮、土壤含水率与土壤MBC质量分数具有显著正相关关系,而土壤pH与土壤MBC呈显著负相关,表明滨海湿地土壤有机质、土壤含水率、土壤pH都是影响土壤MBC质量分数的限制性因子。
Characteristics and influencing factors of soil microbial biomass carbon content at different succession stages of coastal wetlands in Jiangsu Province
-
摘要:
目的 研究江苏滨海湿地土壤微生物生物量碳(MBC)质量分数在不同演替阶段的分布特征和季节动态,揭示其主要影响因素。 方法 选择江苏滨海湿地典型演替序列的不同阶段,光滩、大米草Spartina anglica群落、碱蓬Suaeda glauca群落、芦苇Phragmites australis群落和刺槐Robinia pseucdoacacia群落为研究对象,分析植被演替、土层和季节因素对土壤微生物生物量碳质量分数的影响,以及土壤微生物生物量碳质量分数与土壤理化性质的关系,探讨影响滨海湿地土壤微生物生物量碳质量分数的关键因素。 结果 土壤微生物生物量碳质量分数范围为116.91~326.18 mg·kg−1,不同演替阶段之间差异显著,但分布趋势与演替方向不一致,从高到低依次为大米草群落、芦苇群落、光滩、碱蓬群落、刺槐群落。在0~10 cm土层,大米草群落土壤微生物生物量碳质量分数在4个季节均显著大于其他演替阶段。不同演替阶段的土壤微生物生物量碳质量分数在3个土层均表现出随季节先增加再下降的季节变化趋势,在秋季达到峰值,而在春季或冬季则较低。滨海湿地土壤微生物生物量碳受季节和演替阶段影响较显著,其中季节因素的影响力比例最大,占32.29%。土壤微生物生物量碳质量分数与土壤总有机碳、总氮和土壤含水率之间呈显著正相关关系,而与土壤pH呈极显著的负相关关系。 结论 植被演替和季节是影响江苏滨海湿地土壤微生物生物量碳分布和动态特征的主要因素,其中季节因素的影响力最大,土壤有机质、含水率和pH是直接影响土壤微生物活性的关键性因子。图1表2参35 Abstract:Objective The objective is to explore distribution characteristics and seasonal variations of soil microbial biomass carbon (MBC) content at different succession stages of coastal wetlands in Jiangsu Province, and to reveal its main influencing factors. Method Five typical succession stages of coastal wetlands in Jiangsu were selected as the research objects, including coastal mudflats, Spartina auglica wetland, Suaeda glauca wetland, Phragmites australis wetland, and Robinia pseucdoacacia forest. The distribution characteristics of soil MBC at the different succession stages, the effects of vegetation succession, soil layer and season on MBC, the relationship between soil MBC and soil physical and chemical properties were analyzed. The key factors affecting soil MBC in coastal wetlands were discussed. Result Soil MBC content ranged from 116.91 to 326.18 mg·kg−1, with a significant difference between different succession stages, but the distribution trend was not consistent with the succession direction. The highest was Sp. auglica, followed by P. australis, coastal mudflats, Su. glauca, and R. pseucdoacacia. In 0−10 cm soil depth, soil MBC content in Sp. auglica wetland was significantly higher than that of other succession stages in four seasons. Soil MBC content at different succession stages first increased and then decreased with variation of seasons in the three soil layers, reaching its peak in autumn but lower in spring or winter. Soil MBC in coastal wetlands was significantly affected by seasons and succession stages, among which the seasonal factors had the largest impact, accounting for 32.29%. There existed a significant positive correlation between soil MBC content and total organic carbon, total nitrogen and soil moisture content, but a significant negative correlation with soil pH. Conclusion Vegetation succession and seasons are the main factors affecting the distribution and dynamic characteristics of soil MBC in coastal wetlands, among which seasonal factors have the greatest influence, and soil organic matter, moisture content and pH are the key factors that directly affect soil microbial activity. [Ch, 1 fig. 2 tab. 35 ref.] -
图 1 不同演替阶段土壤微生物生物量碳质量分数季节动态
Figure 1 Seasonal variations of soil MBC contents under different succession stages
表 1 植被演替、土层和季节对土壤MBC质量分数的影响
Table 1. Effects of vegetation succession, soil layer and season on soil MBC contents
影响因素 分布值/% P 植被演替 26.23 0.000 土层 0.91 0.087 季节 32.29 0.000 植被演替×土层 0.96 0.728 植被演替×季节 5.86 0.003 土层×季节 7.05 0.000 植被演替×土层×季节 4.84 0.348  下载: 导出CSV
下载: 导出CSV
表 2 土壤微生物生物量碳与土壤理化因子相关性
Table 2. Pearson correlation coefficients (r-value) between MBC and physic-chemical factors of soil
项目 土壤
有机碳总氮 C/N pH 容重 含水率 MBC 0.187* 0.173* 0.062 −0.225** −0.190* 0.461** 说明:*表示显著相关水平(P<0.05);**表示极显著相关水平 (P< 0.01)
下载: 导出CSV
-
[1] D’ANDRILLI J, COOPER W T, FOREMAN C M, et al. An ultrahigh-resolution mass spectrometry index to estimate natural organic matter lability [J]. Mass Spectrom, 2015, 29(24): 2385 − 2400. [2] HU Yufu, JIANG Shuanglong, YUAN Shu, et al. Changes in soil organic carbon and its active fractions in different desertification stages of alpine-cold grassland in the eastern Qinghai-Tibet Plateau [J]. Environ Earth Sci, 2017, 76(1): 348. [3] LU Xin, HU Haiqing, SUN Long. Effect of fire disturbance on active organic carbon of Larix gmelinii forest soil in Northeastern China [J]. J For Res, 2017, 28(4): 1 − 12. [4] ZHANG Shuai, XU Mingxiang, ZHANG Yengya, et al. Effects of land use change on soil active organic carbon in deep soils in Hilly Loess Plateau region of northwest China [J]. Environ Sci, 2015, 36: 661 − 668. [5] KABIRI V, RAIESI F, GHAZAVI M A. Tillage effects on soil microbial biomass, SOM mineralization and enzyme activity in a semi-arid Calcixerepts [J]. Agric Ecosyst Environ, 2016, 232: 73 − 84. [6] XIAO Ye, HUANG Zhigang, LU Xianguo. Changes of soil labile organic carbon fractions and their relation to soil microbial characteristics in four typical wetlands of Sanjiang Plain, northeast China [J]. Ecol Eng, 2015, 82: 381 − 389. [7] 李红琴, 徐海燕, 马小亮, 等. 马衔山多年冻土与季节冻土区土壤微生物量及酶活性的季节动态[J]. 冰川冻土, 2017, 39(2): 421 − 428. LI Hongqin, XU Haiyan, MA Xiaoliang, et al. The seasonal dynamics of soil microbial biomass and enzyme activities in permafrost area and seasonally frozen ground area of the Maxian Mountain [J]. J Glaciol Geocryol, 2017, 39(2): 421 − 428. [8] 周世兴, 邹秤, 肖永翔, 等. 模拟氮沉降对华西雨屏区天然常绿阔叶林土壤微生物生物量碳和氮的影响[J]. 应用生态学报, 2017, 28(1): 12 − 18. ZHOU Shixing, ZOU Cheng, XIAO Yongxiang, et al. Effects of simulated nitrogen deposition on soil microbial biomass carbon and nitrogen in natural ever- green broad-leaved forest in the rainy area of west China [J]. Chin J Appl Ecol, 2017, 28(1): 12 − 18. [9] 杨帆, 杨万勤, 吴福忠, 等. 萘对川西亚高山森林土壤呼吸、可溶性有机质和微生物生物量的影响[J]. 应用生态学报, 2018, 29(3): 757 − 764. YANG Fan, YANG Wangqin, WU Fuzhong, et al. Effects of naphthalene on soil respiration, dissolved organic matter and microbial biomass in the subalpine forest of western Sichuan, China [J]. Chin J Appl Ecol, 2018, 29(3): 757 − 764. [10] 蒋永梅, 姚拓, 田永亮, 等. 不同管理措施对高寒草甸土壤微生物量季节性变化的影响[J]. 草原与草坪, 2016, 36(5): 105 − 110. JIANG Yongmei, YAO Tuo, TIAN Yongliang, et al. Effect of different management patterns on seasonal dynamics of soil microbial biomass in the alpine meadow [J]. Grassland Turf, 2016, 36(5): 105 − 110. [11] 杨文英, 邵学新, 梁威, 等. 杭州湾湿地土壤酶活性分布特征及其与活性有机碳组分的关系[J]. 湿地科学与管理, 2011, 7(2): 54 − 58. YANG Wenying, SHAO Xuexin, LIANG Wei, et al. Vertical dynamics of soil enzyme activities and its relationship with active organic carbon fractions in Hangzhou bay wetland [J]. Wetland Sci Manage, 2011, 7(2): 54 − 58. [12] 张静, 马玲, 丁新华, 等. 扎龙湿地不同生境土壤微生物生物量碳氮的季节变化[J]. 生态学报, 2014, 34(13): 3712 − 3719. ZHANG Jing, MA Ling, DING Xinhua, et al. Seasonal dynamics of soil microbial biomass C and N in different habitats in Zhalong Wetland [J]. Acta Ecol Sin, 2014, 34(13): 3712 − 3719. [13] 石思博, 王旭东, 叶正钱, 等. 菌渣化肥配施对稻田土壤微生物量碳氮和可溶性碳氮的影响[J]. 生态学报, 2018, 38(23): 8612 − 8620. SHI Sibo, WANG Xudong, YE Zhengqian, et al. Effects of the combination of fungal residue and chemical fertilizer on soil microbial biomass carbon and nitrogen and dissolved organic carbon and nitrogen in paddy soil [J]. Acta Ecol Sin, 2018, 38(23): 8612 − 8620. [14] XIONG Xiong, GRUNWALD S, MYERS D B, et al. Interaction effects of climate and land use/land cover change on soil organic carbon sequestration [J]. Sci Total Environ, 2014, 493: 974 − 982. [15] HUANG Wei, McDOEELL W H, ZOU Xiaoming, et al. Qualitative differences in headwater stream dissolved organic matter and riparian water-extractable soil organic matter under four different vegetation types along an altitudinal gradient in the Wuyi Mountains of China [J]. Appl Geochem, 2015, 52: 67 − 75. [16] 国家林业局. 森林土壤分析方法: LY/T 1210~1275−1999[S]. 北京: 中国标准出版社, 1999: 1 − 108. [17] 王磊, 何冬梅, 刘华, 等. 江苏滨海湿地不同植被演替阶段土壤有机碳分布特征[J]. 安徽农业大学学报, 2017, 44(6): 1064 − 1069. WANG Lei, HE Dongmei, LIU Hua, et al. Distribution of soil organic carbon under different vegetation successions in the coastal wetland of Jiangsu [J]. J Anhui Agric Univ, 2017, 44(6): 1064 − 1069. [18] 田舒怡, 满秀玲. 大兴安岭北部森林土壤微生物量碳和水溶性有机碳特征研究[J]. 土壤通报, 2016, 47(4): 838 − 845. TIAN Shuyi, MAN Xiuling. Characteristics of soil microbial biomass carbon and dissolved organic carbon in northern forest region of Daxing’an montains [J]. Chin J Soil Sci, 2016, 47(4): 838 − 845. [19] 孙小琳, 孔范龙, 李悦, 等. 胶州湾滨海湿地枯落物分解对土壤活性有机碳含量及其三维荧光特性的影响[J]. 应用生态学报, 2019, 30(2): 563 − 572. KONG Xiaolin, KONG Fanlong, LI Yue, et al. Effects of litter decomposition on contents and three-dimensional fluorescence spectroscopy characteristics of soil labile organic carbon in coastal wetlands of Jiaozhou Bay, China [J]. Chin J Appl Ecol, 2019, 30(2): 563 − 572. [20] 管海英, 王权, 赵鑫, 等. 2种典型荒漠植被区土壤微生物量碳的季节变化及影响因素分析[J]. 干旱区地理, 2015, 38(1): 67 − 75. GUAN Haiying, WANG Quan, ZHAO Xin, et al. Seasonal patterns of soil microbial biomass C and impacting factors in two typical arid desert vegetation regions [J]. Arid Land Geogr, 2015, 38(1): 67 − 75. [21] YANG Wen, ZHAO Hui, CHEN Xinglong, et al. Consequences of short-term C4 plan Spartina alerniflora invasions for soil organic carbon dynamics in a coastal wetland of eastern China [J]. Ecol Eng, 2013, 61(12): 50 − 57. [22] 张祥霖, 石盛莉, 潘根兴, 等. 互花米草入侵下福建漳江口红树林湿地土壤生态化学变化[J]. 地球科学进展, 2008, 23(9): 974 − 981. ZHANG Xianglin, SHI Shengli, PAN Genxing, et al. Changes in ecochemical properties of a mangrove wetland under spartina invasion from Zhangjiangkou, Fujian, China [J]. Adv Earth Sci, 2008, 23(9): 974 − 981. [23] 周义贵, 郝凯婕, 李贤伟, 等. 川西亚高山不同土地利用类型对土壤微生物量碳动态特征的影响[J]. 自然资源学报, 2014, 29(11): 1944 − 1956. ZHOU Yigui, HAO Kaijie, LI Xianwei, et al. Effect of different land use patterns on seasonal dynamic of soil microbial biomass carbon in the subalpine forest of western Sichuan, China [J]. J Nat Resour, 2014, 29(11): 1944 − 1956. [24] 刘明慧, 孙雪, 于文杰, 等. 长白山不同海拔原始红松林土壤活性有机碳含量的生长季动态[J]. 南京林业大学学报(自然科学版), 2018, 42(2): 67 − 74. LIU Minghui, SUN Xue, YU Wenjie, et al. Seasonal dynamics of soil active organic carbon content in the original Pinus koraiensis forest in Changbai Mountains, China [J]. J Nanjing For Univ Nat Sci Ed, 2018, 42(2): 67 − 74. [25] 韩广轩. 潮汐作用和干湿交替对盐沼湿地碳交换的影响机制研究进展[J]. 生态学报, 2017, 37(24): 8170 − 8178. HAN Guangxuan. Effect of tidal action and drying-wetting cycles on carbon exchange in a salt marsh: progress and prospects [J]. Acta Ecol Sin, 2017, 37(24): 8170 − 8178. [26] 刘平, 邱月, 王玉涛, 等. 渤海泥质海岸典型防护林土壤微生物量季节动态变化[J]. 生态学报, 2019, 39(1): 363 − 370. LIU Ping, QIU Yue, WANG Yutao, et al. Seasonal dynamics of soil microbial biomass in typical shelterbelts on the Bohai muddy coast [J]. Acta Ecol Sin, 2019, 39(1): 363 − 370. [27] 涂志华, 尉永键, 范志平, 等. 太子河源流域不同类型水源涵养林土壤微生物生物量碳、氮的季节动态[J]. 生态学杂志, 2018, 37(7): 2139 − 2147. TU Zhihua, WEI Yongjian, FAN Zhiping, et al. Seasonal variations of soil microbial biomass C and N in different types of water conservation forest in the headstream of Taizi River watershed [J]. Chin J Ecol, 2018, 37(7): 2139 − 2147. [28] 刘纯, 刘延坤, 金光泽. 小兴安岭6种森林类型土壤微生物量的季节变化特征[J]. 生态学报, 2014, 34(2): 451 − 459. LIU Chun, LIU Yankun, JIN Guangze. Seasonal dynamics of soil microbial biomass in six forest types in Xiaoxing’an Mountains, China [J]. Acta Ecol Sin, 2014, 34(2): 451 − 459. [29] 杨凯, 朱教君, 张金鑫, 等. 不同林龄落叶松人工林土壤微生物生物量碳氮的季节变化[J]. 生态学报, 2009, 29(10): 5500 − 5507. YANG Kai, ZHU Jiaojun, ZHANG Jinxin, et al. Seasonal dynamics of soil microbial biomass C and N in two larch plantation forests with different ages in northeastern China [J]. Acta Ecol Sin, 2009, 29(10): 5500 − 5507. [30] 张剑, 汪思龙, 王清奎, 等. 不同森林植被下土壤活性有机碳含量及其季节变化[J]. 中国生态农业学报, 2009, 17(1): 41 − 47. ZHANG Jian, WANG Silong, WANG Qingkui, et al. Content and seasonal change in soil labile organic carbon under different forest covers [J]. Chin J Eco-Agric, 2009, 17(1): 41 − 47. [31] ZHOU Zhenghu WANG C K. Reviews and syntheses: Soil resources and climate jointly drive variations in microbial biomass carbon and nitrogen in China’s forest ecosystems [J]. Biogeosci Discuss, 2015, 12(14): 6751 − 6760. [32] 王薪琪, 韩轶, 王传宽. 帽儿山不同林龄落叶阔叶林土壤微生物生物量及其季节动态[J]. 植物生态学报, 2017, 41(6): 597 − 609. WANG Xinqi, HAN Yi, WANG Chuankuan. Soil microbial biomass and its seasonality in deciduous broadleaved forests with different stand ages in the Mao’ershan region, northeast China [J]. Chin J Plant Ecol, 2017, 41(6): 597 − 609. [33] SONG Piao, REN Haibao, JIA Qi, et al. Effects of historical logging on soil microbial communities in a subtropical forest in southern China [J]. Plant Soil, 2015, 397: 115 − 126. [34] 张雨洁, 王斌, 李正才, 等. 施肥措施对古香榧林地土壤活性有机碳和养分的影响[J]. 林业科学研究, 2019, 32(2): 87 − 93. ZHANG Yujie, WANG Bin, LI Zhengcai, et al. Effects of fertilization measures on soil labile organic carbon and nutrient of old Torreya grandis [J]. For Res, 2019, 32(2): 87 − 93. [35] 范志平, 王琼, 李法云. 辽东山地不同森林类型土壤有机碳季节动态及其驱动因子[J]. 生态学杂志, 2018, 37(11): 3220 − 3230. FAN Zhiping, WANG Qiong, LI Fayun. Seasonal dynamics of soil organic carbon in different forest types and its driving factors in mountainous region of eastern Liaoning [J]. Chin J Ecol, 2018, 37(11): 3220 − 3230. -

-
链接本文:
https://zlxb.zafu.edu.cn/article/doi/10.11833/j.issn.2095-0756.20190565
 点击查看大图
点击查看大图
计量
- 文章访问数: 2701
- HTML全文浏览量: 756
- PDF下载量: 373
- 被引次数: 0

