






-
城市污水处理过程中会产生大量剩余污泥,若不妥善处理,对环境危害较大。目前,卫生填埋仍是城市污泥处理的重要方式[1]。部分城市在早期建有简易污泥填埋池,随着使用年限增加,污泥池环境风险系数增加,需对污泥池污泥进行安全处理。污泥池加固稳定化处理的常用方法有抛石挤淤法、固化法[2]、外运等。其中外运挖出的污泥可采用资源化处置[3-4]。近年来,好氧发酵技术在污泥处理中有着极大的应用潜力,能显著提升污泥中速效态营养成分的含量以及降低污泥中的重金属的生物有效性。王舜和等[5]研究发现:对污水处理厂剩余污泥进行好氧发酵处理能有效提高污泥速效氮磷钾含量,污泥中速效钾含量在好氧发酵处理后提高了13%,而且该研究也阐述了发酵后的污泥可作为速效肥与园林土混合应用于园林绿化。刘莹等[6]通过添加外源菌剂(食苯芽孢杆菌Bacillus benzoevorans和恶臭假单胞菌Pseudomonas putida)对脱水污泥进行好氧发酵处理,发现经好氧发酵处理后,污泥样品铬、砷、铅的酸可交换态百分含量下降,表明污泥好氧发酵过程对降低重金属的生物有效性是有利的。鉴于碳氮比(C/N)是好氧发酵过程中影响微生物活性的重要因素之一[7-8],通过外加调理剂优化污泥好氧发酵过程中的C/N,有望调控发酵后的污泥中的速效氮磷钾含量及其重金属的可交换态百分含量,进而提升发酵后的污泥应用于园林绿化的前景。木屑为林业废弃物,不但具有价格低廉、易于获取等优点,而且疏松保水,是发酵常用的配料。本研究以木屑为主要调理剂,配置不同C/N填埋污泥,研究不同木屑添加量对填埋污泥发酵过程中污泥营养元素及重金属形态的影响,以期为实际资源化应用提供技术参考。
-
发酵污泥为杭州市临安区某城市污水处理厂污泥,平均堆放年限为7 a。木屑购买于杭州某木材交易市场的松树木刨花,基本性状见表1。菌剂为购买于益加益直销店的有益微生物(EM菌),主要包含枯草芽孢杆菌B. subtilis、地衣芽孢杆菌B. licheniformis等有益菌群。
表 1 发酵污泥及木屑基本性质
Table 1. Basic physical and chemical properties of composting materials
发酵原料 pH 含水率/% 有机质/(g·kg−1) 总碳
/(g·kg−1)总氮/(g·kg−1) 碳氮比 铜
/(mg·kg−1)锌
/(mg·kg−1)污泥 7.26±0.04 69.40±0.46 651.5±1.2 183.38±1.24 26.90±0.92 6.80±0.21 1793.36±29.90 3152.02±57.14 木屑 5.51±0.01 12.38±1.00 997.0±0.0 481.52±0.12 2.20±0.04 218.90±9.00 8.30±0.21 15.54±0.50 -
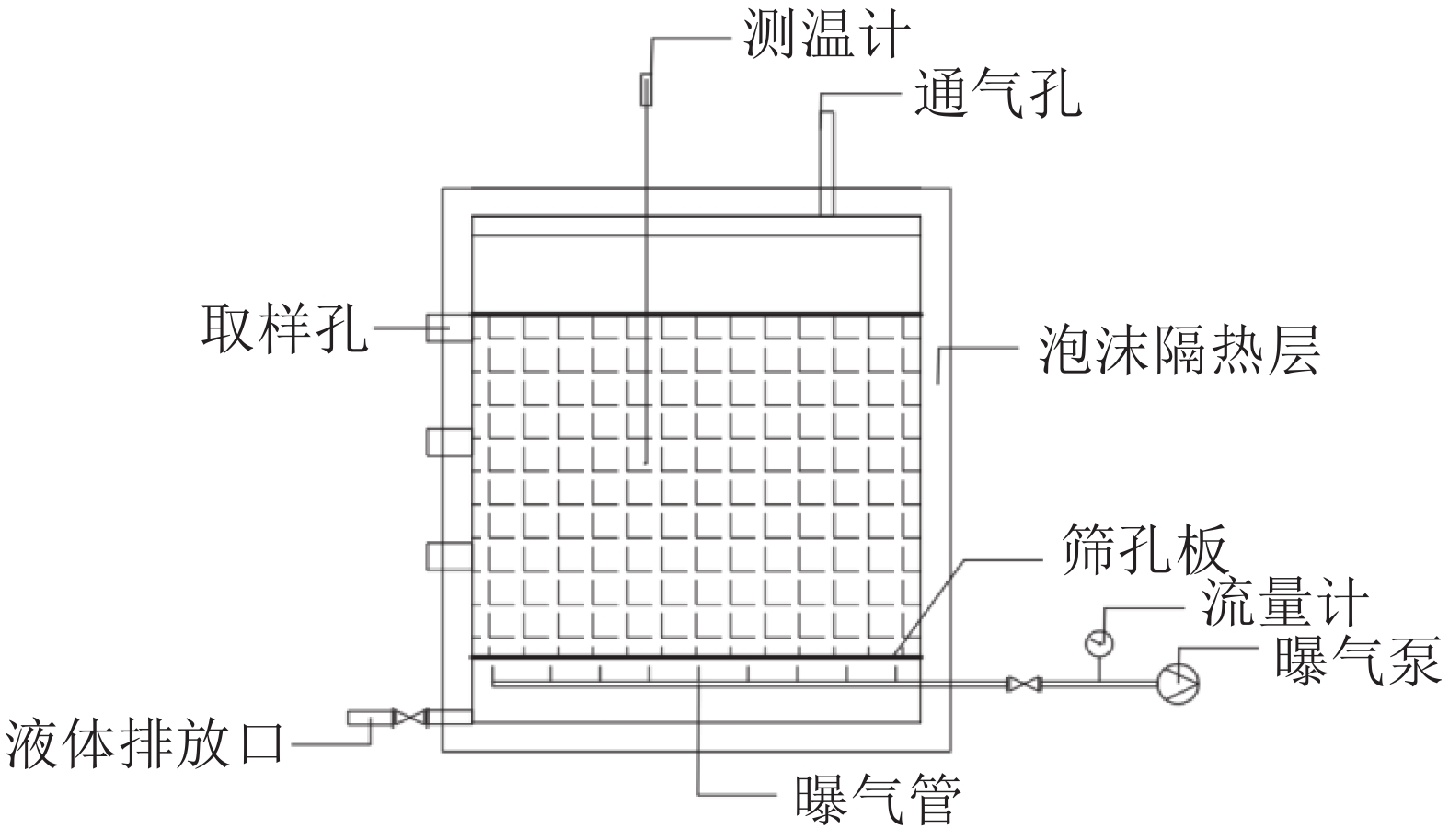
污泥好氧发酵实验在自行设计的发酵反应器(图1)中进行。发酵反应器由泡沫箱制成,规格为70 cm×40 cm×40 cm,壁厚3 cm,配顶盖。将表1所示物料分别按照污泥∶木屑质量比为3.54∶1.00(T1,C/N20),2.49∶1.00(T2,C/N25),1.91∶1.00(T3,C/N30)比例配置。各组添加质量分数为2‰的活性菌(拌100 g玉米淀粉),混合均匀后采用机械强制通风,每天分别在9:00、14:00、18:00通风3次,风速0.045 m3·min−1·m−3,每次30 min。每天20:00记录堆体温度,并于第0、3、8、14、54天 9:00取样,每组重复3次。
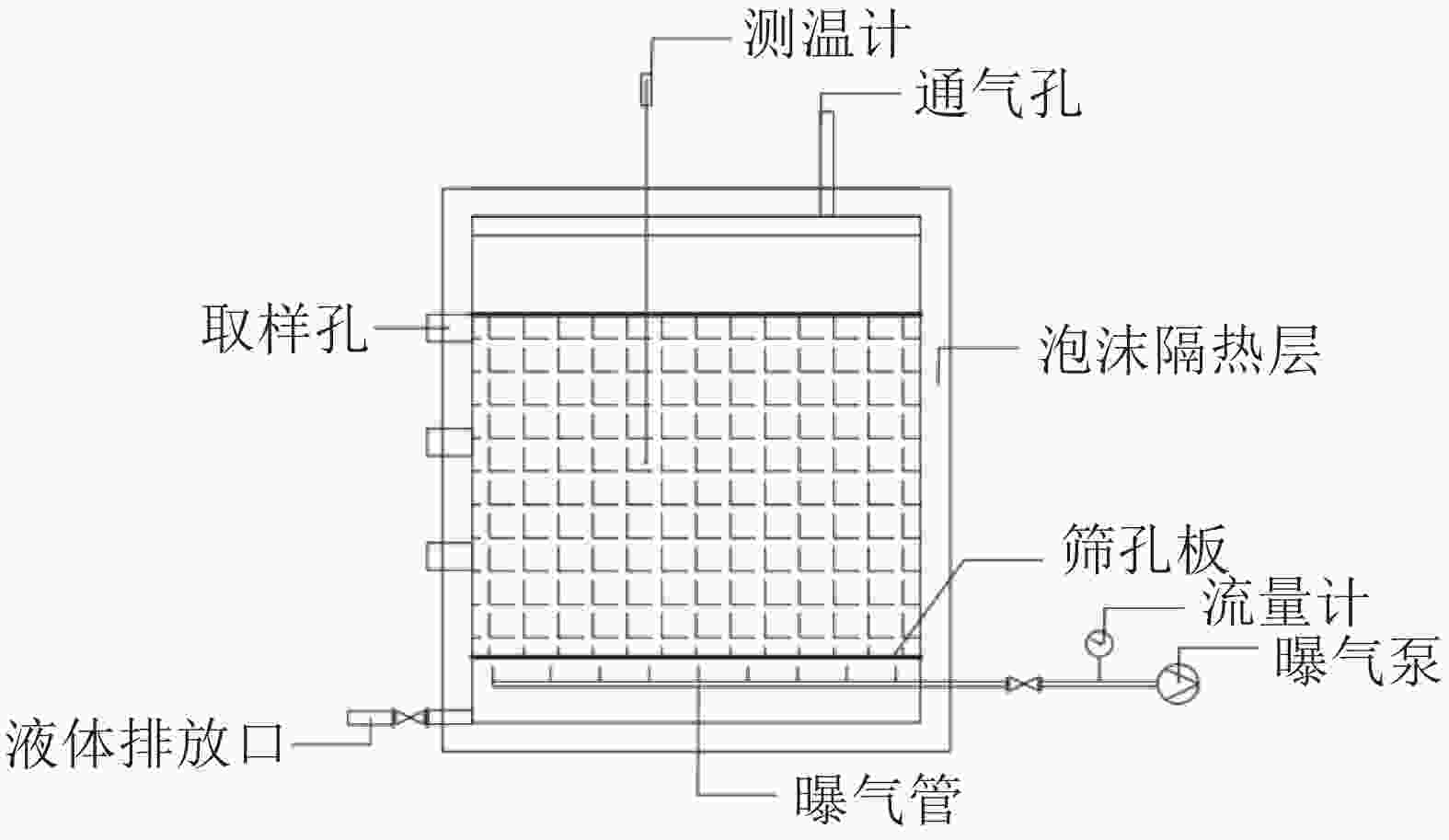
图 1 发酵反应器示意图
Figure 1. Sketch map of static fermentation reactor
-
采用探针式温度计多点测量法测量堆体温度。将鲜样与超纯水按质量体积比1∶10混合,震荡离心过滤,分别用pH计和电导率仪测定pH和电导率。含水率采用重量法测定。有机质采用差重法[9]测定。总碳、总氮质量分数由元素分析仪(S20091640 Vario max CN,德国)测定,铵态氮质量分数测定采用氯化钾浸提靛酚蓝比色法,硝态氮质量分数测定采用氯化钾浸提紫外分光光度法,总磷质量分数测定采用氢氧化钠熔融后钼锑抗分光光度法[10],有效磷质量分数测定采用双酸浸提法,总钾质量分数测定采用氢氧化钠熔融火焰光度计法[10],有效性钾质量分数测定采用冷的2 mol·L−1硝酸溶液浸提法[10]。重金属各级形态测定采用欧洲共同体标准物质局 (European Community Bureau of Reference,BCR)四步提取法[11],重金属全量测定采用样品经高氯酸-硝酸(HClO4-HNO3)消煮后[12],电感耦合等离子体发射光谱仪(ICP-OES,Prodigy 7,Leeman,美国)测定。种子发芽指数测定采用鲜样与超纯水按质量体积比1∶3,震荡1 h后离心,取5 mL浸提液于铺有滤纸并放有20粒小白菜Brassica campestris种子的培养皿中,25 ℃避光培养48 h[13]。相关公式:①各速效营养占比=各速效营养质量分数×100%/各营养元素总质量分数;②种子发芽指数IG=(浸提液种子发芽率×根长)×100%/(对照组种子发芽率×根长);③各形态重金属分配率=各形态重金属×100%/重金属全量,生物可利用态重金属分配率=醋酸溶解态重金属分配率+可还原态重金属分配率。
-
采用Excel 2020和SPSS 26.0进行数据处理和分析,采用Origin 2018 进行绘图。
-
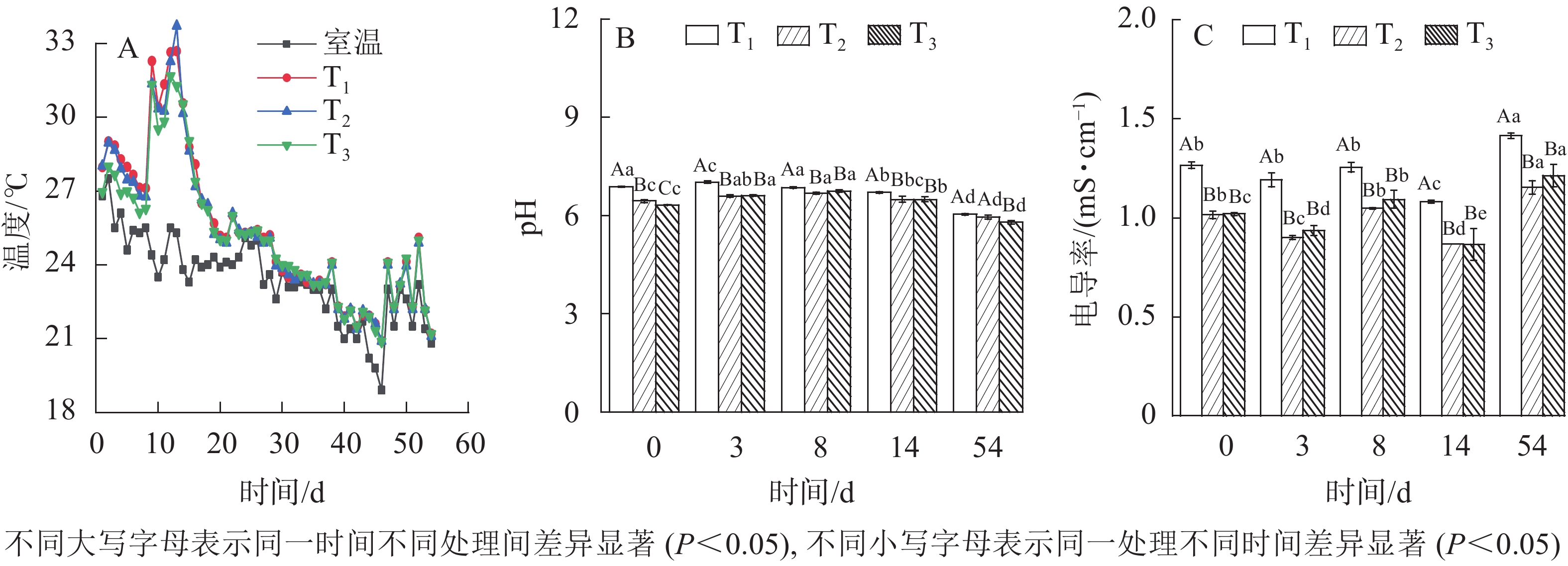
适宜的温度是促进微生物生长的重要保障[11]。如图2A所示:T1、T2、T3发酵过程温度变化均呈现3个阶段:升温阶段、中温阶段和降温阶段,所能达到的最高温度分别为32.69、33.71、31.68 ℃。这与新鲜污泥经3~5 d发酵后,温度快速升至45 ℃以上的高温发酵(45~ 60 ℃)不同,本次发酵为典型的中温发酵。可能的原因为填埋污泥在泥坑中被一定程度矿化,可生化有机物被大量消耗,不利于高温好氧微生物大量繁殖。同时T1、T2、T3发酵温度均在第9 天达30 ℃以上,保持的时间分别为6、6、4 d,其中以T2达到的温度最高,T1次之。相关性比较显示,3组发酵温度均呈现显著差异(P<0.05),然而不同堆体间最高温度与有效发酵维持时间不存在显著差异(P>0.05)。这说明不同木屑添加量对污泥中温好氧发酵影响显著,但对有效发酵(30~45 ℃)过程影响较小。
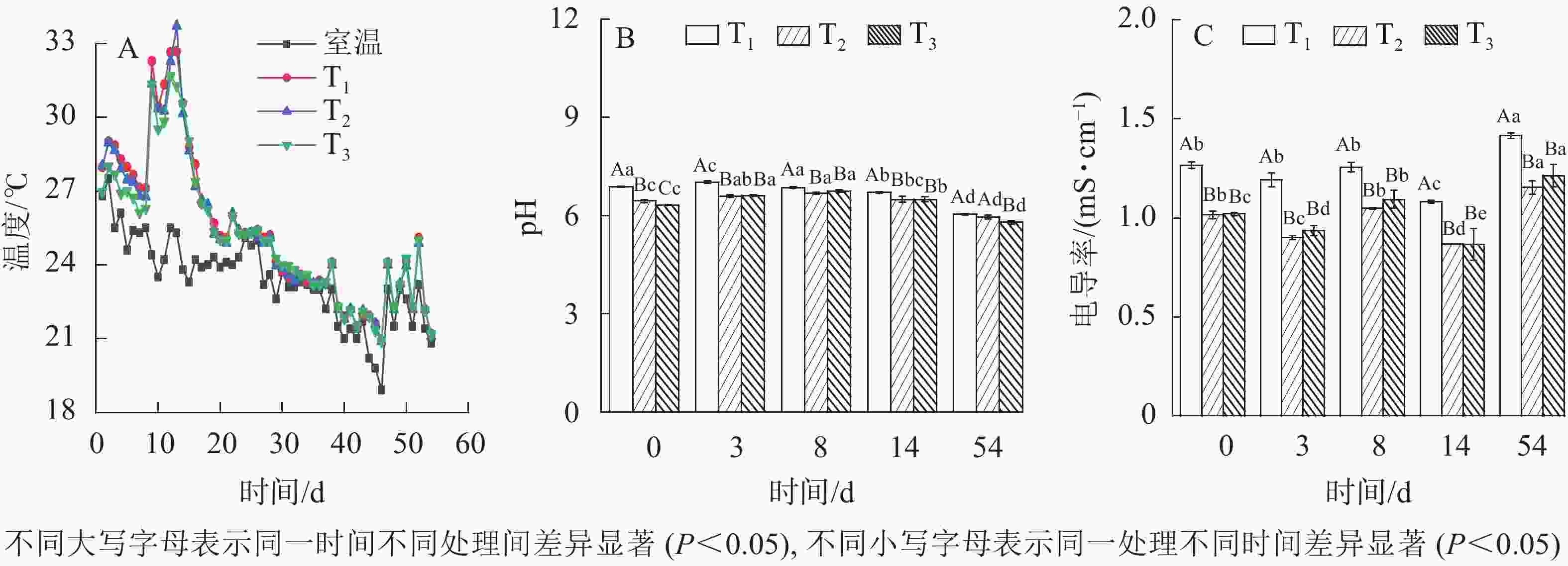
图 2 不同木屑添加量下发酵温度、pH及电导率
Figure 2. Temperature, pH values and electrical conductance values during sludge composting process with different amount of sawdust
YUAN等[12]研究表明:发酵过程pH的变化会影响发酵后污泥性质及用途。如图2B:各处理pH呈现先升高后降低。发酵过程中通过补充超纯水保持污泥含水率为50%,因此排除稀释作用,而极大原因是由于发酵前期好氧分解对有机氮矿化[13],产生一定量的铵态氮致pH增大。可随着发酵的深入,有机物被分解生成有机酸和二氧化碳,硝化作用也产生部分H+[14],引起系统pH降低。木屑不同添加量的污泥发酵过程pH变化显著(
$P_{{{\rm{T}}_1}{\text{-}}{{\rm{T}}_2}}$ =0.02、$P_{{{\rm{T}}_1}{\text{-}}{{\rm{T}}_3}} $ =0.04、$P_{{{\rm{T}}_2}{\text{-}}{{\rm{T}}_3}} $ =0.01)。至发酵54 d结束,T1、T2、T3的 pH分别降低至6.04、5.95和5.79,符合GB/T 23486—2009《城镇污水处理厂污泥处理 园林绿化用泥质》碱性土壤理化指标。过高的电导率不利于微生物发酵与作物生长[15-17]。如图2C所示:与初始值相比,发酵结束后,T1、T2、T3电导率增长率分别为12%、14%、19%,增至1.42、1.15、1.21 mS·cm−1,均可满足GB/T 23486—2009耐盐植物标准限值2.0 mS·cm−1。
-
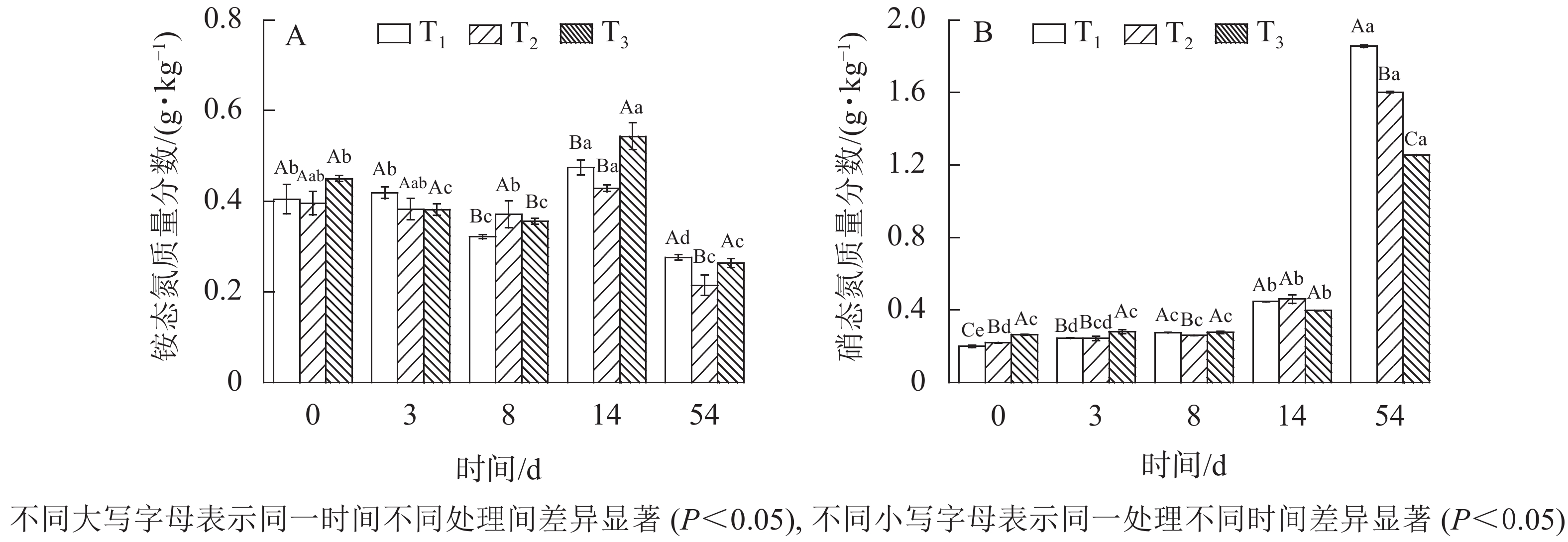
由图3A可知:铵态氮质量分数呈现降低—升高—降低的变化趋势。0~8 d铵态氮降低,可能是由于发酵堆体曝气和升温导致氨挥发,造成铵态氮流失。随着有机氮被微生物不断降解,铵态氮质量分数逐渐回升。至发酵14 d后,硝化细菌逐渐成为发酵优势菌,铵态氮经硝化细菌作用转化为硝态氮,其质量分数缓慢下降。这种变化受木屑添加量的影响,3组数据之间铵态氮差异显著(
$P_{{{\rm{T}}_1}{\text{-}}{{\rm{T}}_3}} $ =0.03、$P_{{{\rm{T}}_2}{\text{-}}{{\rm{T}}_3}} $ =0.05、$P_{{{\rm{T}}_1}{\text{-}}{{\rm{T}}_2}} $ =0.06)表明:木屑添加量及差值越大差异越显著。至发酵结束,T1、T2、T3质量分数铵态氮分别为0.28、0.21、0.26 g·kg−1,相比较堆肥初期,损失率分别为30%、47%、42%。铵态氮质量分数降低被认为是堆体腐熟的标志之一[18],pH也相应降低[19]。这与堆体温度和pH变化趋势相一致。如图3B所示:硝态氮变化呈增加趋势。发酵结束后,T1、T2、T3硝态氮质量分数分别增至1.85、1.60、1.26 g·kg−1,是发酵原始物料的8.25、6.27、3.85倍。主要原因为随着发酵的进行,物料进入腐熟期,硝化作用逐渐增强,促进铵态氮向硝态氮转化[14]。木屑不同添加量对发酵污泥硝态氮有显著的影响 (P<0.05),木屑添加量越小硝态氮增量越大。
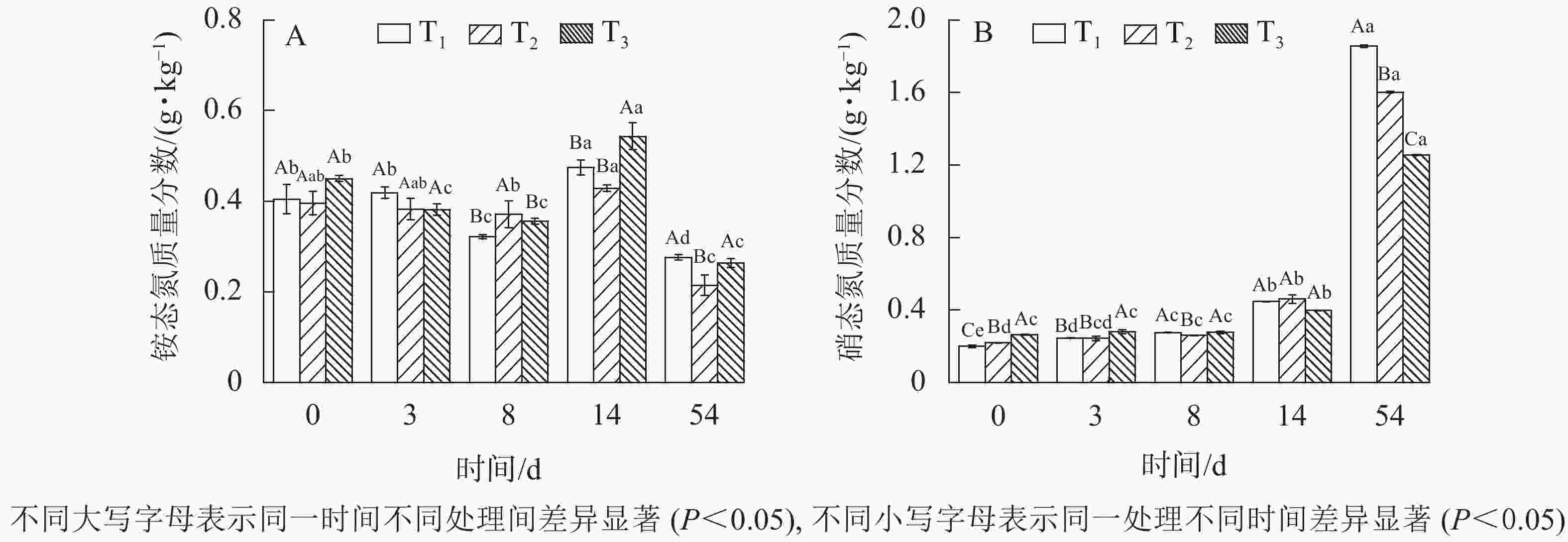
图 3 不同木屑添加量下铵态氮和硝态氮质量分数
Figure 3. Nitrogen contents during sludge composting process with different amount of sawdust
铵态氮、硝态氮是园林植物氮素的主要利用形式[20],可以直观表示氮素养分。0 d时,T3铵态氮和硝态氮质量分数较T1和T2高,可能的原因为木屑呈酸性且对铵态氮和硝态氮具有较好的吸附作用,污泥与木屑混合过程,增加了单位质量铵态氮和硝态氮,且与木屑添加量成正比,这与0 d 时三者中T3 pH最低及木屑添加量最高相对应。然而经过54 d发酵,T1铵态氮、硝态氮质量分数之和显著高于T2、T3,氮素营养价值较高,各处理间均存在显著差异(
$P_{{{\rm{T}}_1}{\text{-}}{{\rm{T}}_2}} $ =0、$P_{{{\rm{T}}_1}{\text{-}}{{\rm{T}}_3}} $ =0.01、$P_{{{\rm{T}}_2}{\text{-}}{{\rm{T}}_3}} $ =0.02)。分析认为:污泥含氮量远高于木屑,在C/N较低的情况下,污泥相比木屑比例高,转化为速效营养的潜力更大。说明木屑添加量对污泥发酵后氮素质量分数有直接显著影响,同时因为发酵过程存在浓缩效应,实际影响大于此结果。 -
由图4A可知:发酵过程总磷质量分数有一定程度增加,与初始质量分数相比分别增加了7.0%、11.8%、10.8%,但木屑不同添加量之间总磷质量分数变化不存在显著性差异(
$P_{{{\rm{T}}_1}{\text{-}}{{\rm{T}}_2}} $ =0.10、$P_{{{\rm{T}}_1}{\text{-}}{{\rm{T}}_3}} $ =0.21、$P_{{{\rm{T}}_2}{\text{-}}{{\rm{T}}_3}} $ =0.38)。可能的原因为磷素较为稳定,无挥发形式逸出,因此总磷在整个发酵过程中脱离反应系统的途径极少[21],反而因浓缩效应,有一定程度的增加。有效磷质量分数是衡量发酵污泥磷素养分高低的指标之一。如图4B所示:发酵过程中有效磷质量分数整体呈增加趋势。经过54 d发酵,T1、T2、T3分别增至2.01、1.94、1.86 g·kg−1,分别占总磷质量分数的18%、22%、25%,与发酵原料相比分别增加8%、9%、9%。显著性检验表明差异不显著(
$P_{{{\rm{T}}_1}-{{\rm{T}}_3}} $ =0.16、$P_{{{\rm{T}}_1}-{{\rm{T}}_2}} $ =0.20、$P_{{{\rm{T}}_2}-{{\rm{T}}_3}} $ =0.29),证明此发酵过程有效磷变化受木屑添加量影响较小。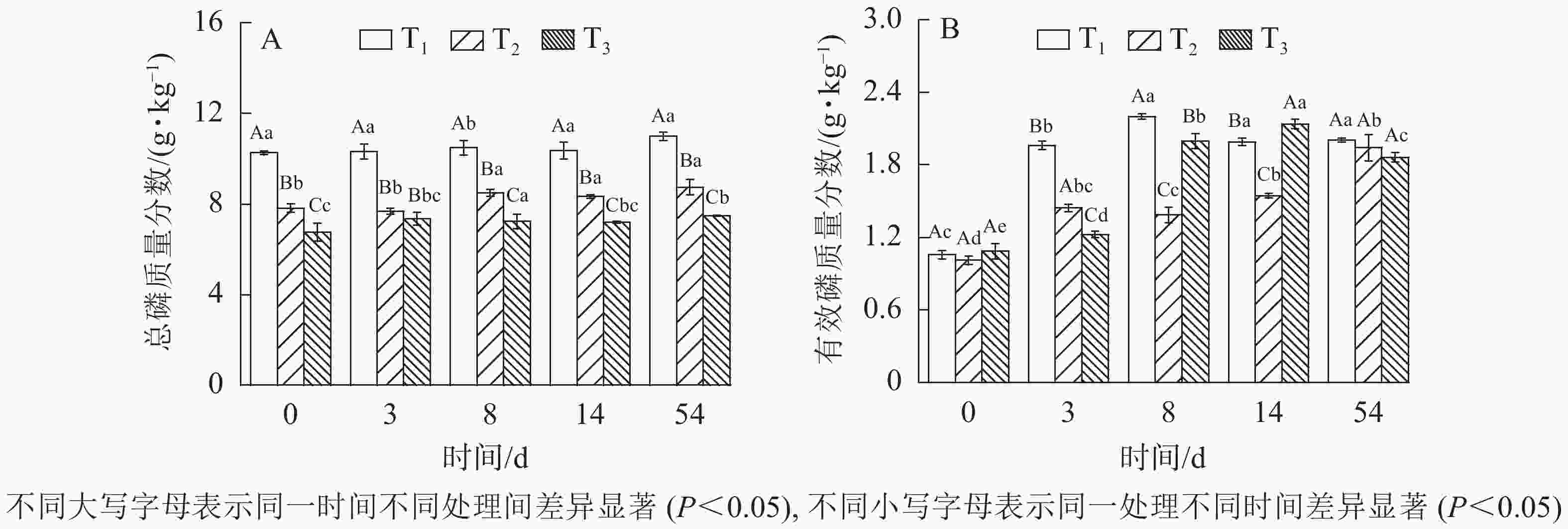
图 4 不同木屑添加量下磷元素质量分数
Figure 4. Phosphorus contents during sludge composting process with different amount of sawdust
-
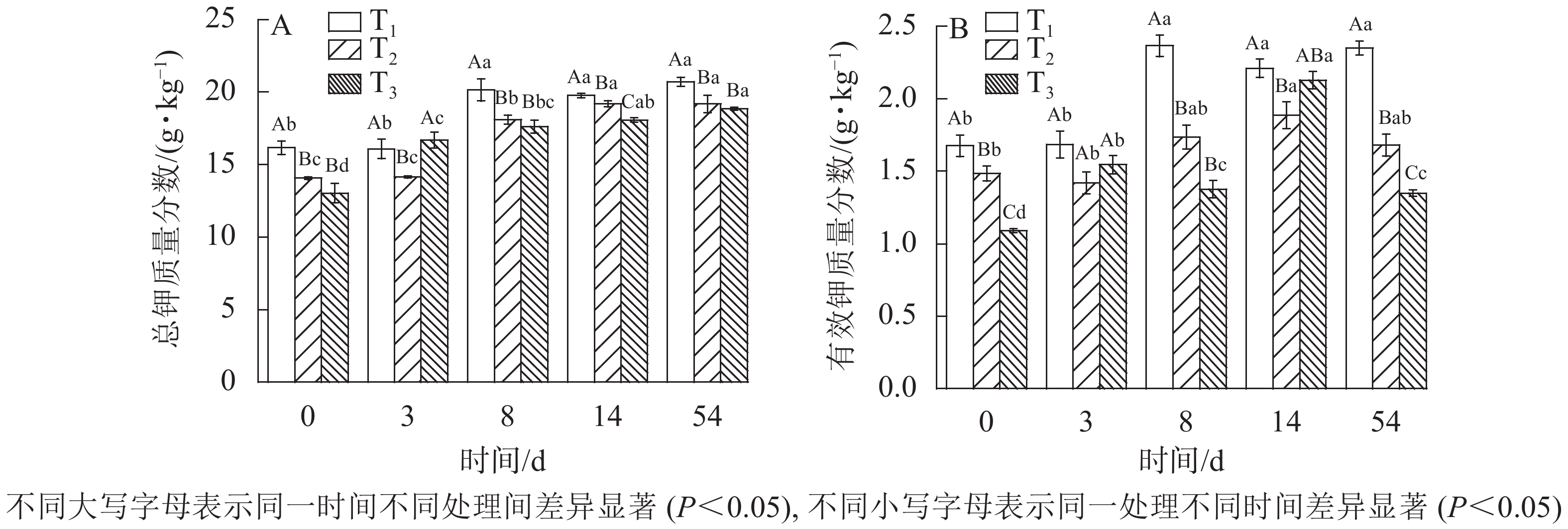
发酵过程中总钾质量分数的变化趋势如图5A所示。随着发酵的进行,物料体积及质量不断减小,营养物质等元素不断被浓缩,与磷相同,钾无逸出形式,因此总钾质量分数也在不断增加。发酵后T1、T2、T3总钾质量分数分别由初始16.17、14.07、13.04 g·kg−1增加至发酵结束后的20.72、19.19、18.83 g·kg−1,其中T1总钾质量分数变化与T2呈显著差异(
$P_{{{\rm{T}}_1}{\text{-}}{{\rm{T}}_2}} $ =0)。总钾质量分数增加量T3最大,T2次之,T1最小,因此3组处理中木屑添加量越高,浓缩效应越明显。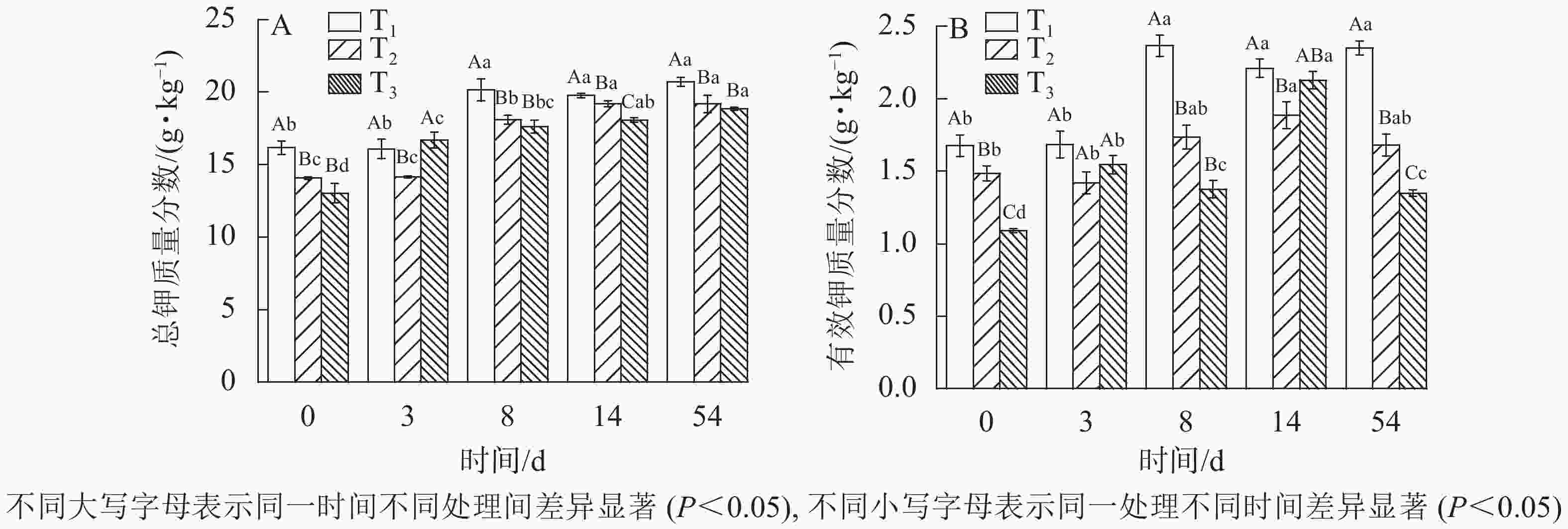
图 5 不同木屑添加量下钾元素质量分数
Figure 5. Potassium contents during sludge composting process with different amount of sawdust
图5B所示:T1、T2、T3的有效钾质量分数呈现先升高后降低的趋势,这与唐淦海[22]的研究实验结果一致。至发酵结束,T1、T2、T3有效钾质量分数分别增至2.35、1.21、1.35 g·kg−1,相比发酵原料分别增加40%、13%、24%,除与堆体浓缩效应有关外还与微生物活动将含钾物质中的钾转化为可溶性钾盐有关[13, 23],但木屑添加量对有效钾质量分数增加幅度的影响不显著 (P>0.05)。
-
分别对添加不同量木屑的污泥样品中的铜、锌等2种重金属元素质量分数进行分析,结果如表2。未经调理的污泥中,铜、锌质量分数高于GB/T 23486—2009《城镇污水处理厂污泥处置 园林绿化用泥质》中碱性土壤污染物限值,其中铜质量分数超标19.5%。经木屑调理发酵后,因物料浓缩作用,铜、锌质量分数较发酵前均有增加,但仍在标准范围内。重金属总量与木屑添加量呈正相关。
表 2 木屑不同添加量发酵后污泥铜锌质量分数
Table 2. Cu and Zn contents after fermentation with different amount of sawdust
处理 铜/(mg·kg−1) 锌/(mg·kg−1) 发酵前 发酵后 发酵前 发酵后 T1 1 075.30±7.23 1 295.06±80.62 1 734.76±45.26 1 985.38±21.49 T2 970.76±6.03 1024.01±3.18 1 514.27±17.16 1 615.77±4.75 T3 816.38±4.70 916.96±11.66 1 240.61±20.82 1 346.57±16.14 重金属在污泥中一般以醋酸溶解态、可还原态、可氧化态、残渣态等4种形态存在[24],其中由醋酸溶解态、可还原态组成的生物有效态与重金属的生物有效性密切相关[18]。由图6可知:在发酵初始阶段,铜以可氧化态为主,其生物可利用态分配率不足50%。发酵处理后,铜的可氧化态和残渣态分配率逐渐降低,醋酸溶解态和可还原态分配率逐渐增加,即生物有效态增加。T1、T2、T3中,生物有效态铜分别从初始的32%、38%、40%增加至72%、74%、74%。这主要由微生物将络合态铜转化为生物可利用态[19]。
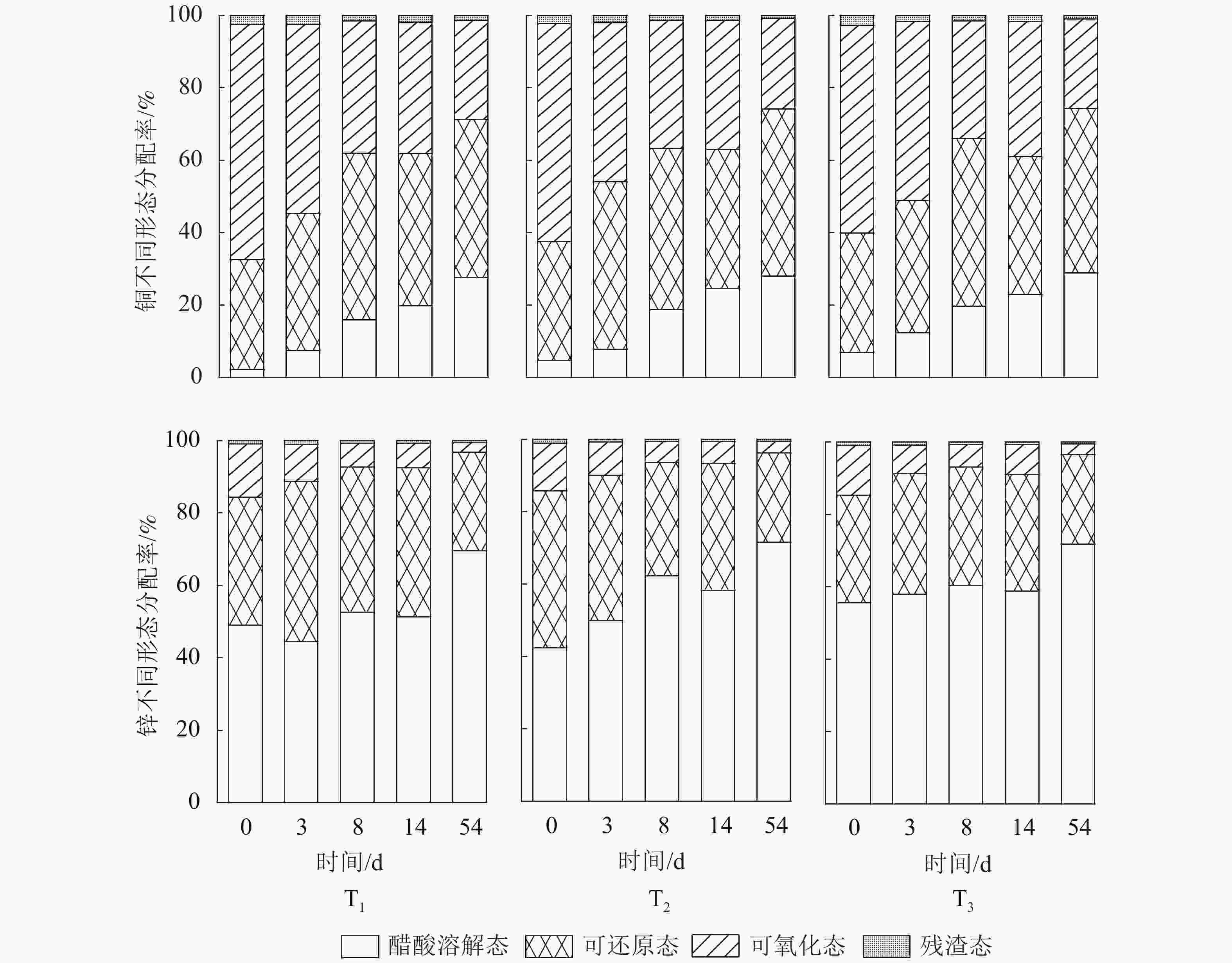
图 6 不同木屑添加量下铜、锌化学形态分配率
Figure 6. Distribution of Cu and Zn during composting process with different amount of sawdust
从图6可知:污泥中的各形态锌所占比例从大到小依次为醋酸溶解态、可还原态、可氧化态、残渣态,表明锌的生物可利用态占比较大。这可能是锌与有机质亲和性较弱[25]。发酵初期,T1、T2、T3生物可利用态分配率分别为84%、85%、86%。发酵过程中锌的可还原态和可氧化态分配率降低,醋酸溶解态分配率增加,增加幅度高于可还原态的降低幅度。发酵结束后,处理T1、T2、T3的生物可利用态锌分配率分别为96%、98%、97%,表明发酵使锌向易于生物利用的形态转化。这与岳建芝等[21]、曾正中等[26]的研究结果相一致。
木屑添加量对发酵过程铜、锌质量分数变化幅度的影响如表3所示。可见木屑添加量对发酵污泥铜、锌金属各形态质量分数均有一定影响。其中对醋酸溶解态铜、可氧化态铜和可氧化态锌影响显著(P<0.05)。对于生物可利用态铜有显著影响(P<0.05),木屑添加量高可降低污泥中铜质量分数。锌稳定态受木屑添加量影响显著,实际应用过程应对这种影响予以考虑。
表 3 木屑不同添加量对铜锌化学形态分配率变化的影响(单变量方差分析)
Table 3. Effects of different amount of sawdust on distribution of Cu and Zn during composting process(univariate)
项目 不同形态铜差异显著性 不同形态锌差异显著性 醋酸溶
解态可还原态 可氧化态 残渣态 生物可
利用态稳定态 醋酸溶
解态可还原态 可氧化态 残渣态 生物可
利用态稳定态 PT1-T2 0.02 0.08 0.02 0.09 0.01 0.02 0.24 0.20 0.00 0.34 0.47 0.00 PT1-T3 0.01 0.00 0.01 0.19 0.00 0.01 0.02 0.04 0.03 0.05 0.05 0.03 PT2-T3 0.02 0.06 0.05 0.01 0.00 0.05 0.27 0.27 0.04 0.07 0.62 0.03 -
种子发芽指数是表征经发酵后的污泥对植物的毒害作用指标之一。当污泥用作园林用泥基质时, IG值应大于70%[25]。本研究污泥原样IG为34.7%,对种子发芽抑制作用较强。T1、T2、T3发酵过程中, IG统计如表4所示。0 d时T1、T2、T3的IG均超过70%。发酵初始阶段, IG短暂上升后出现抑制期,经过抑制期后回升。分析认为,在第8天时,堆体中有机物降解产生大量有机酸、多酚、醛类等毒性物质,对植物生长产生抑制作用。随着发酵的进行,毒性物质分解转化,可利用营养元素质量分数增加,对植物生长的毒害作用减弱,促进作用增强[27]。发酵结束后,处理T1、T2、T3的IG值分别为164.2%、186.4%、160.2%,显示T2对种子毒性最小,而且后两者IG降低呈现显著差异 (P=0.04),说明种子发芽指数受木屑添加量影响大。
表 4 发酵过程中种子发芽指数(IG)值统计
Table 4. Statistics of different amount of sawdust on seed germination index (IG) during composting process
时间/d IG/% T1 T2 T3 0 82.7±9.5 Ce 86.7±9.9 Be 103.2±5.8 Ad 3 103.7±7.4 Cd 133.9±2.2 Ac 127.5±6.9 Bc 8 162.8±10.1 Cb 184.8±6.6 Ab 172.6±4.3 Ba 14 142.6±23.7 Ac 105.8±8.7 Bd 96.8±10.0 Ce 54 164.2±18.4 Ba 186.4±7.5 Aa 160.2±13.6 Cb 说明:不同大写字母表示同一时间不同处理间差异显著(P<0.05),不同小写字母表示同一处理不同时间差异显著(P<0.05) -
好氧发酵可分为升温阶段、中温阶段和降温阶段等3个阶段,木屑添加量对发酵温度、pH、氮、钾、可生物利用态铜、稳定态锌及IG均有显著性影响(P<0.05)。
不同木屑添加量处理的填埋污泥,经发酵后pH、电导率、重金属铜和锌及IG均符合GB/T 23486—2009《城镇污水处理厂污泥处理 园林绿化用泥质》碱性土壤用泥标准,且氮、磷、钾等营养元素的总量及其速效营养质量分数均有不同程度增加,木屑添加量在C/N为20时营养潜力最高,为填埋污泥中温好氧发酵制作园林绿化碱性土壤调理剂提供了数据参考。
木屑添加量控制在C/N为25的T2,在发酵前后pH、电导率波动变化幅度小,有效重金属铜降幅大,种子发芽指数最高,实际应用时对木屑添加量控制最具参考价值。
Optimal addition of aerobic fermentation sawdust in landfill sludge reuse
-
摘要:
目的 为了提升污泥在园林绿化方面的应用潜力,探讨填埋污泥发酵时最佳的木屑添加量,以期为工程应用提供参考。 方法 以杭州地区某填埋池污泥为主要原料,基于不同碳氮比(20、25、30)控制木屑添加量,分别配制了T1、T2、T3共3组发酵物料处理,通过间歇式好氧发酵,分析了发酵过程中温度、pH、电导率、氮、磷、钾和重金属形态等理化指标的变化。 结果 发酵为典型中温发酵(30~45 ℃),发酵过程中不同木屑添加量之间温度呈现差异显著(P<0.05),pH呈先升高后降低趋势,受木屑添加量影响显著(P<0.05);电导率受木屑添加量影响较小。发酵结束后,3组处理铵态氮损失率分别为30%、47%、42%,硝态氮增加了8.25、6.27、3.85倍,说明木屑添加量对氮素影响大。总磷增长率分别为7.0%、11.8%、10.8%,有效磷占比为8%、9%、9%,总钾占比为27%、36%、44%,有效钾占比为8%、12%、17%,其中仅T1和T2之间总钾呈现显著差异(P<0.05)。铜的生物有效态分配率分别从初始的32%、38%、40%增加至72%、74%、74%,生物可利用态锌分别从84%、85%、86%增加至96%、98%、97%;不同木屑添加量下,生物有效态、可氧化态铜、锌分别呈显著差异(P<0.05);3组发酵物对植物生长均无毒性[种子发芽指数(IG>80%)],但T2毒性显著大于T3毒性(P<0.05)。 结论 发酵过程pH、氮、钾、生物可利用态铜、稳定态锌和种子发芽指数受木屑添加量显著影响(P<0.05),3组处理所检测的指标均符合GB/T 23486—2009《城镇污水处理厂污泥处置 园林绿化用泥质》碱性土壤用泥限值标准,其中T2处理的木屑添加量对发酵过程控制最具参考价值。图6表4参27 Abstract:Objective The objective is to explore the optimal amount of sawdust during landfill sludge fermentation so as to improve the potential of sludge in landscaping. Method Three groups of fermentation materials, i.e. C/N20(T1), C/N25(T2), and C/N30(T3), were prepared by using sludge from a landfill in Hangzhou as the main raw material and sawdust as a conditioning agent. Through batch aerobic fermentation, the changes of physical and chemical indicators such as temperature, pH, conductivity, nitrogen, phosphorus, potassium and heavy metal forms were analyzed. Result The fermentation was a typical medium temperature fermentation (30~45 ℃). During the fermentation process, there was a significant difference in temperature between different sawdust additions (P<0.05), and the pH increased first and then decreased, which was significantly affected by sawdust amount while the conductivity was less affected. The ammonium nitrogen loss rates for T1, T2 and T3 after the fermentation were 30%, 47%, and 42%, respectively, and the nitrate nitrogen rates increased by 8.25, 6.27, and 3.85 times, indicating that the addition of sawdust had a great impact on nitrogen. The growth rates of total phosphorus for T1, T2 and T3 were 7.0%, 11.8%, and 10.8%, respectively, and the proportions of available phosphorus were 8%, 9%, and 9%, respectively. The growth rates of total potassium were 27%, 36%, and 44%, and the proportions of available potassium were 8%, 12%, and 17%, respectively, among which there was a significant difference in total potassium between T1 and T2 (P<0.05). The bioavailability of Cu for T1, T2 and T3 increased from 32%, 38%, and 40% to 72%, 74%, and 74%, and the bioavailability of Zn for T1, T2 and T3 increased from 84%, 85%, and 86% to 96%, 98%, and 97%, respectively. There were significant differences in bioavailable and oxidized Cu and Zn under different sawdust addition levels (P<0.05). The fermentation products of the three groups had no toxicity to plant growth (IG>80%), but the toxicity of T2 was significantly greater than that of T3 (P<0.05). Conclusion The pH, nitrogen, potassium, bioavailable Cu, stable Zn, and seed germination index in the fermentation process are significantly affected by the amount of sawdust (P<0.05). The indicators of the three treatments are all in line with the GB/T 23486−2009 limit standard for argillaceous alkaline soil for landscaping, of which T2 has the most reference value for sawdust control in fermentation process. [Ch, 6 fig. 4 tab. 27 ref.] -
Key words:
- landfill sludge /
- sawdust /
- C/N ratio /
- aerobic fermentation
-
图 2 不同木屑添加量下发酵温度、pH及电导率
Figure 2 Temperature, pH values and electrical conductance values during sludge composting process with different amount of sawdust
图 3 不同木屑添加量下铵态氮和硝态氮质量分数
Figure 3 Nitrogen contents during sludge composting process with different amount of sawdust
图 4 不同木屑添加量下磷元素质量分数
Figure 4 Phosphorus contents during sludge composting process with different amount of sawdust
图 5 不同木屑添加量下钾元素质量分数
Figure 5 Potassium contents during sludge composting process with different amount of sawdust
图 6 不同木屑添加量下铜、锌化学形态分配率
Figure 6 Distribution of Cu and Zn during composting process with different amount of sawdust
表 1 发酵污泥及木屑基本性质
Table 1. Basic physical and chemical properties of composting materials
发酵原料 pH 含水率/% 有机质/(g·kg−1) 总碳
/(g·kg−1)总氮/(g·kg−1) 碳氮比 铜
/(mg·kg−1)锌
/(mg·kg−1)污泥 7.26±0.04 69.40±0.46 651.5±1.2 183.38±1.24 26.90±0.92 6.80±0.21 1793.36±29.90 3152.02±57.14 木屑 5.51±0.01 12.38±1.00 997.0±0.0 481.52±0.12 2.20±0.04 218.90±9.00 8.30±0.21 15.54±0.50  下载: 导出CSV
下载: 导出CSV
表 2 木屑不同添加量发酵后污泥铜锌质量分数
Table 2. Cu and Zn contents after fermentation with different amount of sawdust
处理 铜/(mg·kg−1) 锌/(mg·kg−1) 发酵前 发酵后 发酵前 发酵后 T1 1 075.30±7.23 1 295.06±80.62 1 734.76±45.26 1 985.38±21.49 T2 970.76±6.03 1024.01±3.18 1 514.27±17.16 1 615.77±4.75 T3 816.38±4.70 916.96±11.66 1 240.61±20.82 1 346.57±16.14
下载: 导出CSV
表 3 木屑不同添加量对铜锌化学形态分配率变化的影响(单变量方差分析)
Table 3. Effects of different amount of sawdust on distribution of Cu and Zn during composting process(univariate)
项目 不同形态铜差异显著性 不同形态锌差异显著性 醋酸溶
解态可还原态 可氧化态 残渣态 生物可
利用态稳定态 醋酸溶
解态可还原态 可氧化态 残渣态 生物可
利用态稳定态 PT1-T2 0.02 0.08 0.02 0.09 0.01 0.02 0.24 0.20 0.00 0.34 0.47 0.00 PT1-T3 0.01 0.00 0.01 0.19 0.00 0.01 0.02 0.04 0.03 0.05 0.05 0.03 PT2-T3 0.02 0.06 0.05 0.01 0.00 0.05 0.27 0.27 0.04 0.07 0.62 0.03
下载: 导出CSV
表 4 发酵过程中种子发芽指数(IG)值统计
Table 4. Statistics of different amount of sawdust on seed germination index (IG) during composting process
时间/d IG/% T1 T2 T3 0 82.7±9.5 Ce 86.7±9.9 Be 103.2±5.8 Ad 3 103.7±7.4 Cd 133.9±2.2 Ac 127.5±6.9 Bc 8 162.8±10.1 Cb 184.8±6.6 Ab 172.6±4.3 Ba 14 142.6±23.7 Ac 105.8±8.7 Bd 96.8±10.0 Ce 54 164.2±18.4 Ba 186.4±7.5 Aa 160.2±13.6 Cb 说明:不同大写字母表示同一时间不同处理间差异显著(P<0.05),不同小写字母表示同一处理不同时间差异显著(P<0.05)
下载: 导出CSV
-
[1] YANG Kai, ZHU Ying, SHAN Ranran, et al. Heavy metals in sludge during anaerobic sanitary landfill: speciation transformation and phytotoxicity [J]. J Environ Manage, 2017, 45(6): 58 − 66. [2] 罗小勇, 王艳明, 熊建英, 等. 垃圾填埋场污泥坑原位修复工程实践[J]. 环境工程学报, 2018, 12(9): 2707 − 2716. LUO Xiaoyong, WANG Yanming, XIONG Jianying, et al. Practice of in-situ remediation of sludge pits inlandfill [J]. Chin J Environ Eng, 2018, 12(9): 2707 − 2716. [3] BIAN Yongning, YUAN Qian, ZHU Guocheng, et al. Recycling of waste sludge: preparation and application of sludge-based activated carbon [J]. Int J Polym Sci, 2018, 2018(3): 1 − 17. [4] CHENG Fangwei, LUO Hongxi, HU Lei, et al. Sludge carbonization and activation: from hazardous waste to functional materials for water treatment [J]. J Environ Chem Eng, 2016, 4(4): 4574 − 4786. [5] 王舜和, 张月勇, 李晓晶, 等. 好氧发酵污泥特性及园林绿化应用的可行性[J]. 环境工程, 2015, 33(增刊1): 595 − 597, 618. WANG Shunhe, ZHANG Yueyong, LI Xiaojing, et al. Characteristics of aerobic fermented sludge and the feasibility of landscaping[J]. Environ Eng, 2015, 33(suppl 1): 595 − 597, 618. [6] 刘莹, 苏青青, 于梦琦, 等. 城市污泥好氧发酵前后重金属形态及有效性变化[J]. 广州化工, 2016, 44(22): 114 − 117. LIU Ying, SU Qingqing, YU Mengqi, et al. Changes in speciation and bio-availability of heavy metals before and after municipal sludge aerobic fermentation [J]. Guangzhou Chem Ind, 2016, 44(22): 114 − 117. [7] TAKAHASHI N, MOCHIZUKI S, MASUDA K, et al. Influence of temperature, water content and C/N ratio on the aerobic fermentation rate of woody biomass [J]. Kagaku Kogaku Ronbunshu, 2017, 43(4): 231 − 237. [8] WANG Bo, PENG Yongzhen, GUO Yuanyuan, et al. Impact of partial nitritation degree and C/N ratio on simultaneous sludge fermentation, denitrification and anammox process [J]. Bioresour Technol, 2016, 219: 411 − 419. [9] 中华人民共和国环境保护部. 固体废物有机质的测定灼烧减量法: HJ 761—2015[S]. 北京: 中国环境科学出版社, 2015. Ministry of Environmental Protection of the People’s Republic of China. Solid Waste-determination of Organic Matter- Ignition Loss Method: HJ 761−2015[S]. Beijing: China Environmental Science Press, 2015. [10] 鲍士旦. 土壤农化分析[M]. 3版. 北京: 中国农业出版社, 2000. BAO Shidan. Soil Agrochemical Analysis[M]. 3rd ed. Beijing: China Agricultural Press, 2000. [11] RAURET G, RUBIO R, LÓPEZ-SÁNCHEZ J. Optimization of tessier procedure for metal solid speciation in river sediments [J]. Int J Environ Anal Chem, 1989, 36(2): 69 − 83. [12] YUAN Yue, PENG Yongzhen, LIU Ye, et al. Change of pH during excess sludge fermentation under alkaline, acidic and neutral conditions [J]. Bioresour Technol, 2014, 174: 1 − 5. [13] 李荣华, 张广杰, 张增强, 等. 添加木炭改善猪粪稻壳好氧堆肥工艺及质量[J]. 农业工程学报, 2014, 30(16): 230 − 238. LI Ronghua, ZHANG Guangjie, ZHANG Zengqiang, et al. Improving pig manure and rice husk compost technology and quality by wood charcoal addition [J]. Transac Chin Soc Agric Eng, 2014, 30(16): 230 − 238. [14] BRITO L M, COUTINHO J, SMITH S R. Methods to improve the composting process of the solid fraction of dairy cattle slurry [J]. Bioresour Technol, 2008, 99(18): 8955 − 8960. [15] REID E, LIU Xingrong, JUDD S. Effect of high salinity on activated sludge characteristics and membrane permeability in an immersed membrane bioreactor [J]. J Membrane Sci, 2006, 283(1/2): 164 − 171. [16] ŞEVIK F, TOSUN İ, EKINCI K. Composting of olive processing wastes and tomato stalks together with sewage sludge or dairy manure [J]. Int J Environ Sci Technol, 2016, 13(5): 1207 − 1218. [17] 王琳, 李素艳, 孙向阳, 等. 不同配比园林绿化废弃物和蘑菇渣混合蚯蚓堆肥的效果[J]. 浙江农林大学学报, 2019, 36(2): 326 − 334. WANG Lin, LI Suyan, SUN Xiangyang, et al. Mixing garden wastes and spent mushroom compost of different ratios for vermicomposting [J]. J Zhejiang A&F Univ, 2019, 36(2): 326 − 334. [18] 马利民, 陈玲, 吕彦, 等. 污泥土地利用对土壤中重金属形态的影响[J]. 生态环境, 2004, 13(2): 151 − 153. MA Limin, CHEN Ling, LÜ Yan, et al. Effects of sewage sludge application to land on species of heavy metals in soil [J]. Ecol Environ, 2004, 13(2): 151 − 153. [19] 生骏, 陆文静, 王洪涛. 粉煤灰对污泥堆肥过程和土地施用后交换态重金属 (Cu, Zn, Pb)的影响[J]. 环境科学, 2007, 28(6): 1367 − 1371. SHENG Jun, LU Wenjing, WANG Hongtao. Effects of fly ash on the exchangeable heavy metals (Cu, Zn, Pb) during sewage sludge composting and land utilization [J]. J Environ Sci, 2007, 28(6): 1367 − 1371. [20] 莫江明, 郁梦德, 孔国辉. 鼎湖山马尾松人工林土壤硝态氮和铵态氮动态研究[J]. 植物生态学报, 1997, 21(4): 335 − 341. MO Jiangming, YU Mengde, KONG Guohui. The dynamics of soil NH4 +-N and NO3 −-N in a pine forest of Dinghushan, as assesses by ion exchange resin bag method [J]. J Phytoecol Sin, 1997, 21(4): 335 − 341. [21] 岳建芝, 常兴涛, 李辉, 等. 花生壳炭对污泥堆肥营养元素和重金属动态变化的影响[J]. 河南农业大学学报, 2019, 53(6): 950 − 958. YUE Jianzhi, CHANG Xingtao, LI Hui, et al. Effects of peanut shell biochar on dynamic changes of nutrient elements and heavy metals during sewage sludge composting [J]. J Henan Agric Univ, 2019, 53(6): 950 − 958. [22] 唐淦海. 不同秸秆污泥堆肥施用土壤-植物效应研究[D]. 重庆: 西南大学, 2011. TANG Ganhai. Studyon Effect of Sludge Compost Application on Soil-Plant Ecosystems[D]. Chongqing: Southwest University, 2011. [23] 杨雪, 连宾, 朱晓玲, 等. 添加钾矿粉对鸡粪堆肥中N、P和K含量的影响[J]. 地球与环境, 2012, 40(2): 286 − 292. YANG Xue, LIAN Bin, ZHU Xiaoling, et al. Effects of adding potassium-bearing mineral powder on nutrogen, potassium and potassium contents of chicken manure compost [J]. Earth Environ, 2012, 40(2): 286 − 292. [24] NEMATI K, BAKAR N K A, ABAS M R, et al. Speciation of heavy metals by modified BCR sequential extraction procedure in different depths of sediments from Sungai Buloh, Selangor, Malaysia [J]. J Hazardous Mater, 2011, 192(1): 402 − 410. [25] HERNÁNDEZ D, PLAZA C, SENESI N, et al. Detection of copper (Ⅱ) and zinc (Ⅱ) binding to humic acids from pig slurry and amended soils by fluorescence spectroscopy [J]. Environ Pollut, 2006, 143(2): 212 − 220. [26] 曾正中, 张贺飞, 潘玉, 等. 污泥添加粉煤灰堆肥化对Cu、Zn、Pb形态的影响[J]. 水土保持学报, 2011, 25(5): 144 − 153. ZENG Zhengzhong, ZHANG Hefei, PAN Yu, et al. Effect of fly ash to status on Cu, Zn, Pb, during composting of sewage sludge [J]. J Soil Water Conserv, 2011, 25(5): 144 − 153. [27] 李思敏, 赵阳悦, 唐锋兵. 辅料配比对市政污泥堆肥效果的影响[J]. 当代化工, 2020, 49(4): 564 − 567, 571. LI Simin, ZHAO Yangyue, TANG Fengbing. Influence of additive ratio on municipal sludge composting effect [J]. Contemp Chem Ind, 2020, 49(4): 564 − 567, 571. -

-
链接本文:
https://zlxb.zafu.edu.cn/article/doi/10.11833/j.issn.2095-0756.20210335
 点击查看大图
点击查看大图
计量
- 文章访问数: 1763
- HTML全文浏览量: 664
- PDF下载量: 20
- 被引次数: 0
















