






-
壮苗培育是实现人工林优质高效培育目标的关键环节,优质苗木不仅能提高造林成活率,也是促进森林早期生长,提升森林质量的重要途径[1]。施肥管理是提高苗木质量的核心技术,深入研究苗木施肥技术不仅可以加速苗木发育进程,缩短育苗周期,同时对苗木养分积累、增产也具有显著作用,还可以提高苗木质量进而提高造林成活率,对促进林业的可持续发展具有重要的指导意义和现实意义[2−3]。苗木施肥主要是针对氮磷钾3种元素进行,氮磷是植物生长的限制元素,同时也是植物生长所必需的矿质营养元素,对苗木的生长与发育具有重要的作用[4]。研究表明:中国土壤氮磷含量低于全球水平,植物的生长容易受到氮磷元素的限制[5−6]。云南松Pinus yunnanensis分布区的土壤中,低磷少氮较明显,制约着云南松人工林的发展[7]。因此,针对氮磷矿质营养元素的施肥研究显得尤为重要。
云南松是中国西南地区的代表树种,是该地区的乡土树种及荒山绿化造林先锋树种,同时也是云南省的主要造林和用材树种,在其分布区的森林和生态系统的可持续发展中发挥着重要作用[8]。然而,云南松苗木质量并不理想。同时,近年来,大面积云南松天然林的逆向择伐,导致其森林质量降低,造成林分和林地退化等,从而形成大面积天然更新的低质低效次生林,因此,需通过人为干预有效地恢复其质量和产量[9]。在实际的造林生产中,人们也普遍缺乏对云南松苗木施肥的重视。大多凭经验施肥,导致用量过少或过多,或不注意肥料之间的配合,浪费现象普遍,甚至造成肥害或施肥不足导致生理病症[10]。这种现象严重阻碍了高质量苗木的培育,因此亟需探寻科学施肥策略,为造林提供优质的苗木。目前氮磷配施已在樟树Cinnamomum camphora[11]、马尾松Pinus massoniana和木荷Schima superba[12]等多种苗木中开展研究,针对云南松苗木的氮磷施肥研究缺乏,氮磷肥如何调控苗木质量及养分利用尚不完全清楚。鉴于此,本研究以云南松苗木为研究对象,开展不同氮磷配施对苗木质量、生物量及养分积累的影响研究,探明苗木的养分需求特征,为云南松苗期养分管理及苗木培育提供理论依据。
-
2020年6月底在西南林业大学苗圃选择2年生的云南松塑料盆栽苗,苗木长势均匀,健康无病虫害,平均苗高为8.1 cm,平均地径为9.8 mm,平均针叶长为11.6 cm,基质为V(腐殖土)∶V(红壤)=1∶3的混合土。根据情况适时对云南松进行浇水,苗木在管理措施上保持完全相同,以排除外部环境条件不同带来的干扰。
-
采用2因素3水平3×3回归设计,共组成9个处理(表1)。氮肥磷肥用量参照张跃敏等[13]的方法。外源性氮肥为尿素(CH4N2O),磷肥为过磷酸钙(CaP2H4O8)。表1数据为纯氮、纯磷用量。
表 1 外源性氮磷添加施肥试验组合
Table 1. Fertilization test combination of exogenous nitrogen and phosphorus addition
处理 氮/(g·株−1) 磷/(g·株−1) 处理 氮/(g·株−1) 磷/(g·株−1) ck 0 0 N1P1 0.4 0.8 P1 0 0.8 N1P2 0.4 1.6 P2 0 1.6 N2P1 0.8 0.8 N1 0.4 0 N2P2 0.8 1.6 N2 0.8 0 -
于2020年12月底,对指标进行测量。各处理选取3株长势一致的苗木用于生物量及养分的测定,各处理重复3次。用游标卡尺测量地径,直尺测定苗高。将每株苗木分为根、茎和叶,分别装入标记好的纸袋中,在105 ℃烘箱中杀青30 min后,调至80 ℃烘干至恒量,测量根、茎、叶的生物量。将根、茎、叶样品烘干后,研磨过筛,用H2SO4-H2O2法消煮植物样品制备待测液用于养分测定。全氮采用奈氏比色法测定,全磷采用钼锑抗比色法测定,全钾采用火焰光度法测定[14]。器官养分储量=器官养分质量分数×器官生物量;器官养分分配比=器官养分储量/单株养分储量×100%;苗木质量指数=苗木总生物量/(苗高/地径+茎生物量/根生物量)[15]
-
利用SPSS对各指标进行单因素方差分析及回归分析,采用Origin、SAS软件作图。
-
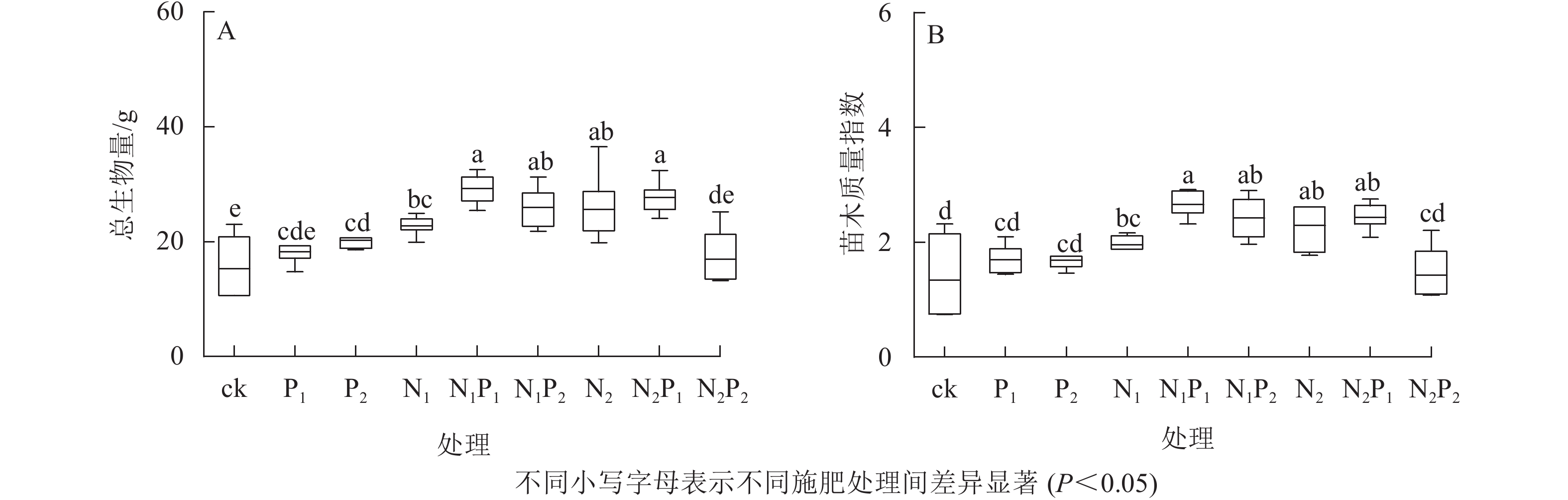
由图1A看出:除P1、N2P2处理外,其余各施肥处理的苗木总生物量均显著大于对照组(P<0.05)。苗木总生物量积累从大到小依次为N1P1、N2P1、N1P2、N2、N1、P2、P1、N2P2、ck。由图1B看出:除P1、P2、N2P2处理外,其余各施肥处理的苗木质量指数均显著大于对照组(P<0.05)。苗木质量指数从大到小依次为N1P1、N2P1、N1P2、N2、N1、P1、P2、N2P2、ck。N1P1处理下苗木具有最大的总生物量及苗木质量指数,分别是ck的1.92、1.98倍。在单施氮肥的处理中,总体上随施氮量的增加,苗木总生物量及质量指数逐渐升高;单施磷肥时,随施磷量的增加,苗木总生物量也逐渐积累,而苗木质量指数几乎没有发生变化,较为稳定。在氮磷配施的处理中,苗木质量随施氮或施磷量的增加,呈降低趋势。
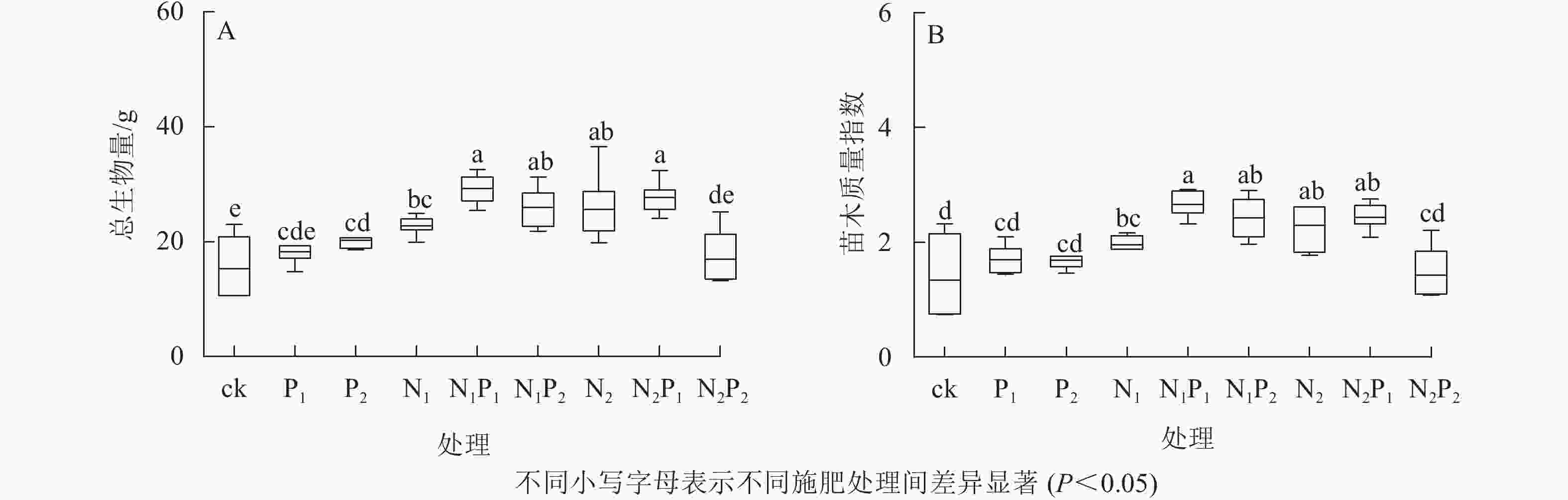
图 1 氮磷配施处理下云南松苗木生物量和质量指数的变化
Figure 1. Change of biomass and quality index of P. yunnanensis seedlings under nitrogen and phosphorus combined application treatments
-
由表2可知:各器官的氮、磷、钾储量总体从大到小依次为叶、茎、根。各施肥处理的器官及单株氮、磷、钾储量间具有显著差异(P<0.05),较对照组均有不同程度的增加。但各处理的器官及单株氮、磷、钾储量大小顺序并不唯一,N1P1处理下苗木根、茎、单株的氮、钾储量最大,根、茎、叶、单株的磷储量也最大,N2P1处理下苗木叶的氮、钾储量最大。在单施氮肥和磷肥的处理中,总体上随施氮量和施磷量的增加,各器官及单株的氮、磷、钾储量逐渐增加。在氮磷配施的处理中,养分储量随施氮或施磷量的增加,呈减小趋势。
表 2 不同施肥处理下云南松苗木养分积累情况
Table 2. Nutrient reserves of P. yunnanensis seedlings under different fertilization treatments
元素 处理 养分储量/mg 根 茎 叶 单株 氮 ck 15.92±11.32 e 33.55±11.64 e 54.94±13.16 e 104.41±35.89 g P1 32.31±18.05 cd 43.50±4.01d e 70.12±8.67 de 145.93±23.64 ef P2 33.85±9.42 cd 55.09±9.11 cd 83.72±8.85 cd 172.65±22.19 de N1 28.62±7.34 de 65.59±8.65 c 104.55±16.30 ab 198.76±16.83 cd N1P1 73.75±17.31 a 103.55±13.97 a 112.53±7.19 a 289.83±22.25 a N1P2 62.52±13.10 ab 94.83±14.29 ab 93.20±12.34 bc 250.55±33.61 ab N2 47.73±11.35 bc 85.20±26.98 b 101.10±21.08 ab 234.03±55.79 bc N2P1 56.37±10.90 b 101.48±18.57 ab 117.79±16.37 a 275.64±33.02 a N2P2 22.17±11.52 de 41.76±9.47 de 60.17±10.77 e 124.10±31.07 fg 磷 ck 3.91±2.62 e 7.15±3.25 e 9.53±2.52 d 20.60±8.27 f P1 10.65±6.78 de 10.08±0.68 de 13.09±1.74 d 33.82±6.90 def P2 13.73±4.37 cd 15.39±4.01 de 12.19±4.07 d 41.31±9.69 de N1 10.56±2.66 de 18.86±2.79 cd 19.39±4.64 cd 48.82±6.95 d N1P1 33.47±10.56 a 38.81±8.12 a 41.7±12.34 a 113.99±18.37 a N1P2 25.93±6.81 b 33.94±10.66 ab 15.98±8.46 d 75.84±13.22 c N2 20.38±7.71 bc 26.53±11.81 bc 26.64±7.72 bc 73.55±25.80 d N2P1 24.63±6.57 ab 34.50±11.03 ab 33.87±13.15 ab 93.00±14.96 b N2P2 6.04±3.14 e 9.14±2.02 e 12.97±3.49 d 28.14±7.71 ef 钾 ck 22.05±15.42 e 50.84±18.39 e 62.14±16.46 f 135.03±50.20 d P1 41.56±22.67 de 59.53±4.21 de 73.14±8.54 ef 174.23±27.31 cd P2 47.57±18.06 cd 84.21±23.45 c 90.03±16.06 de 221.82±55.86 c N1 34.43±8.15 de 79.80±9.55 cd 100.70±19.38 cd 214.94±21.10 c N1P1 102.27±18.50 a 145.46±18.96 a 131.71±9.79 ab 379.44±13.10 a N1P2 82.64±25.07 ab 127.24±20.30 ab 105.62±18.34 cd 315.50±53.39 b N2 63.79±18.56 bc 112.87±33.73 b 114.98±31.44 bc 291.64±80.58 b N2P1 75.91±19.25 b 125.61±19.41 ab 136.33±19.13 a 337.85±34.32 ab N2P2 23.97±7.92 e 58.04±8.15 de 62.89±8.74 f 144.90±22.24 d 说明:数据为平均值±标准差。同列不同小写字母表示同一养分元素同一组织在不同施肥处理间差异显著(P<0.05) -
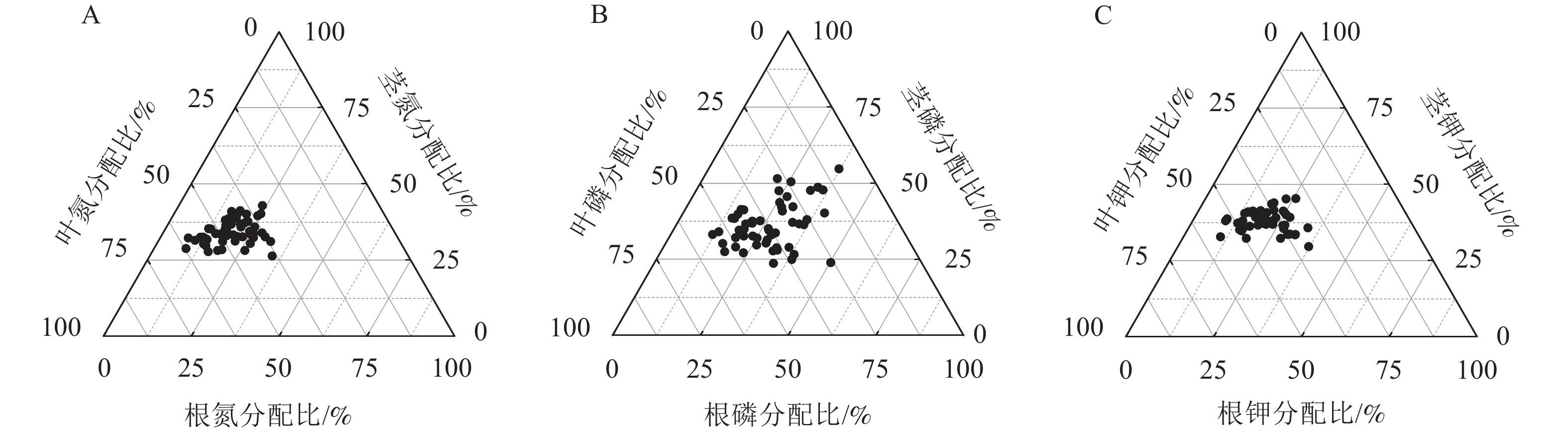
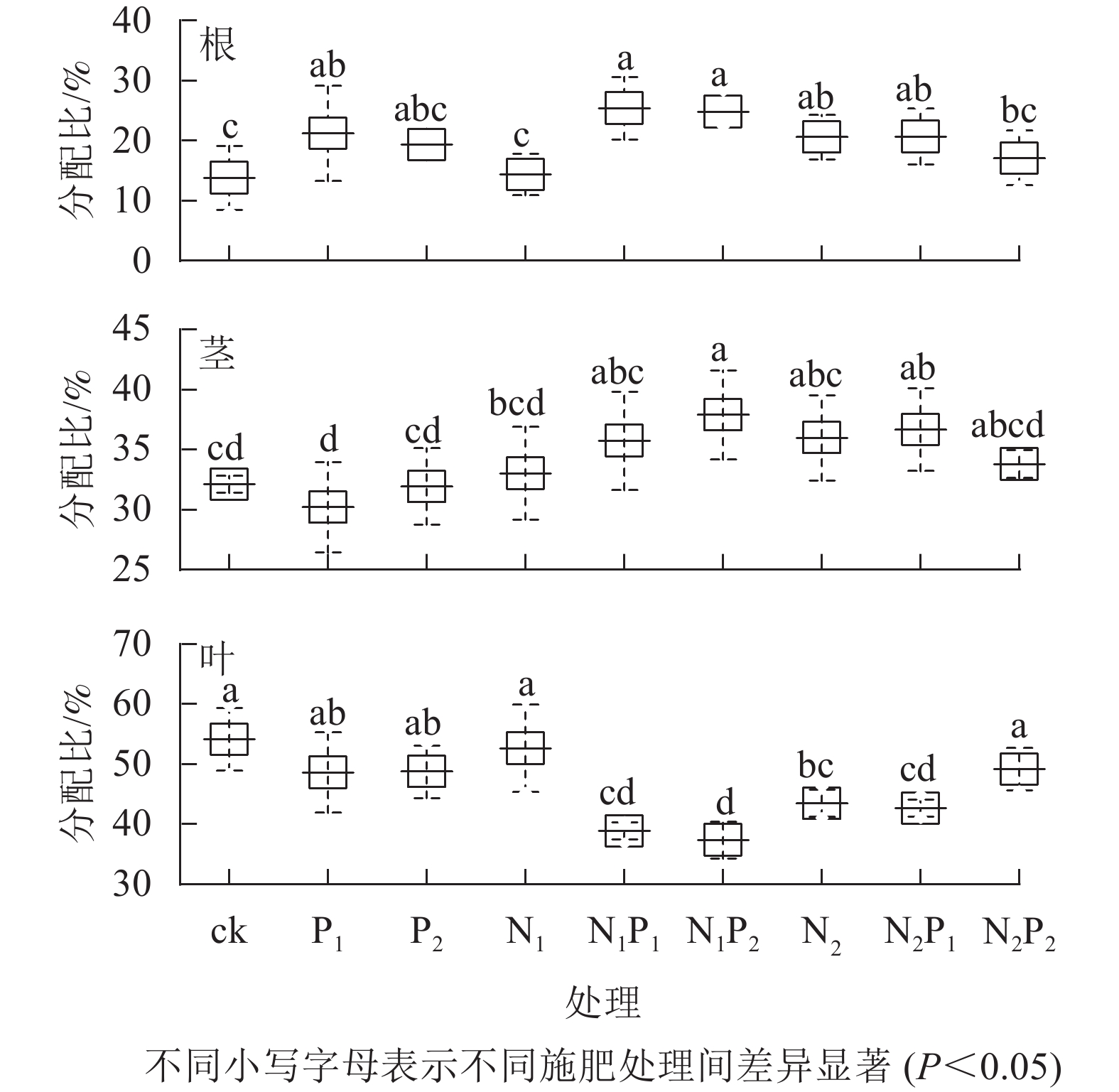
就氮分配情况而言,平均有19.68%分配到根,34.15%分配给茎,46.17%分配到叶(图2A)。就磷分配情况而言,平均有26.66%分配到根,36.17%分配给茎,37.16%分配到叶(图2B)。就钾分配情况而言,平均有20.93%分配到根,38.01%分配给茎,41.07%分配到叶(图2C)。从各处理的氮分配情况看(图3):除P2、N1、N2P2处理外,其余各施肥处理的根氮分配比与对照间差异显著(P<0.05),除P1、P2、N1、N2P2处理外,其余各施肥处理的茎、叶氮分配比与对照间差异显著(P<0.05)。各施肥处理的根氮分配比较对照均有不同程度的增加,茎氮分配比中有6个处理较对照有不同程度的增加,而叶氮分配比较对照均有不同程度的减少。
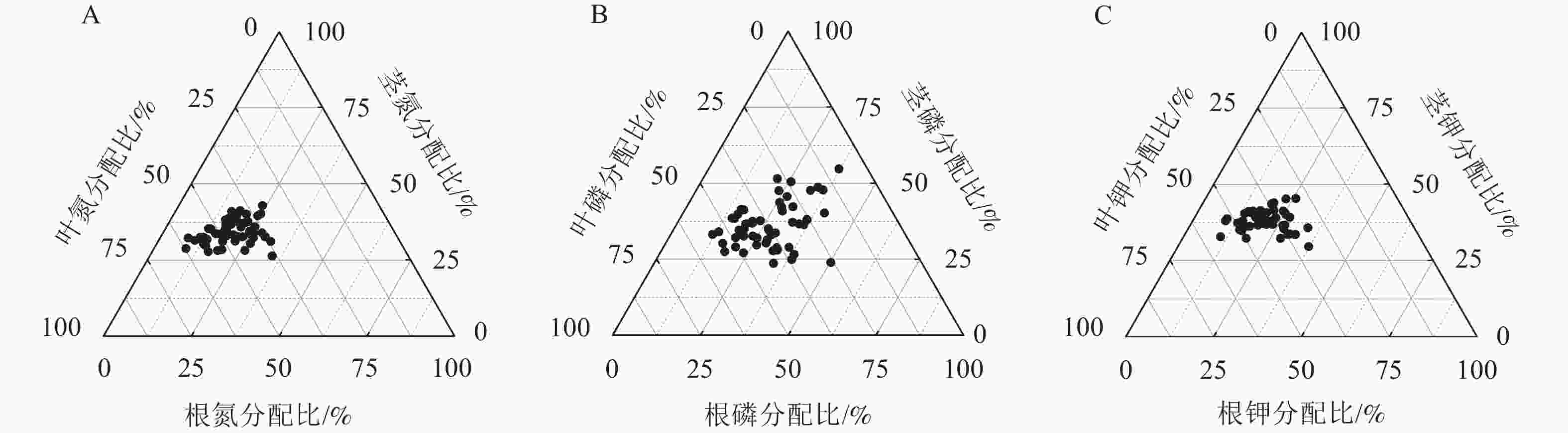
图 2 云南松苗木各器官养分分配总体情况
Figure 2. Nutrient distribution of different organs of P. yunnanensis seedlings
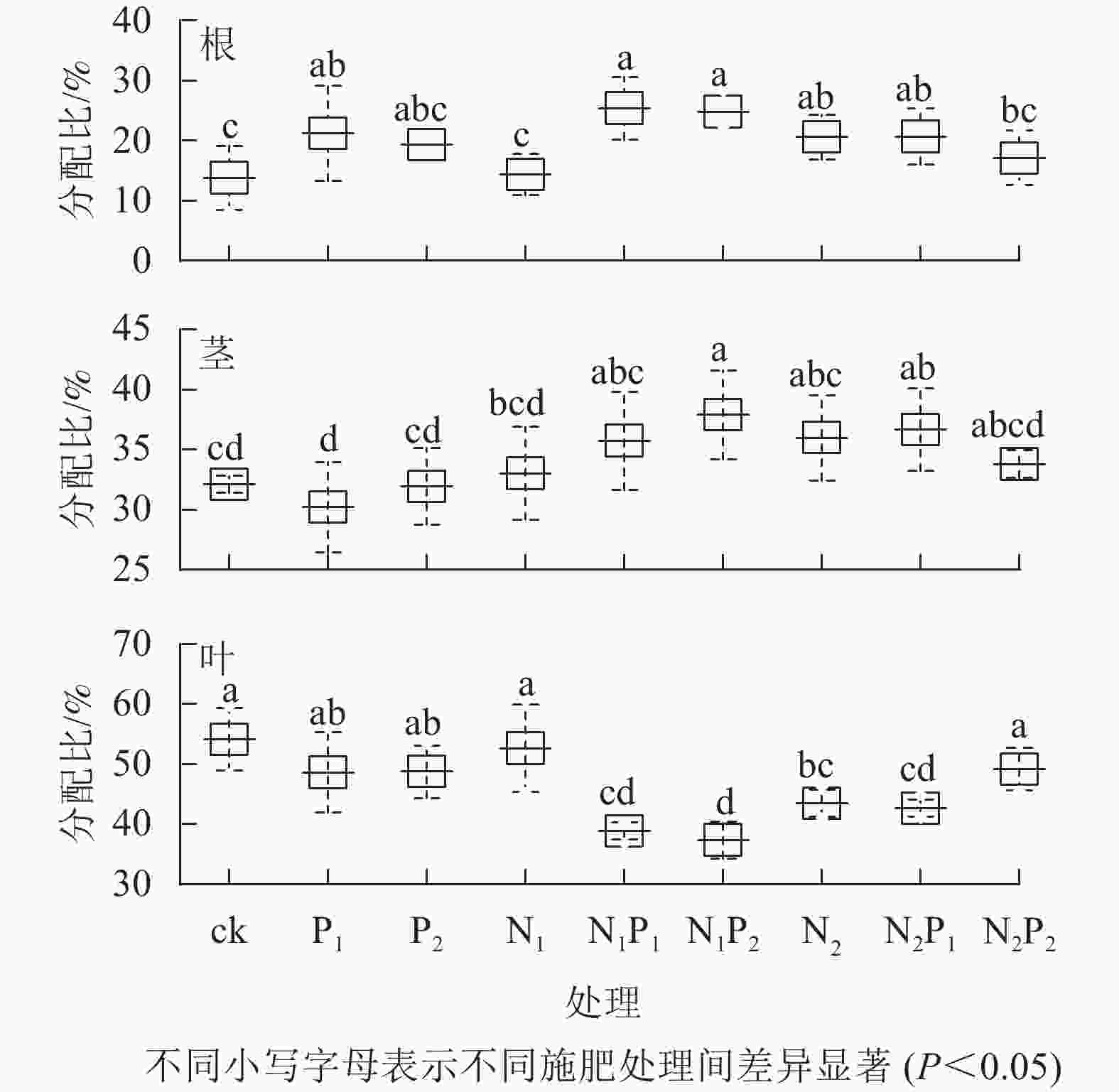
图 3 不同施肥处理云南松苗木各器官氮元素分配情况
Figure 3. Nitrogen distribution to each organ in P. yunnanensis seedlings under different fertilization treatments
由图4可知:除N2P2处理外,其余各施肥处理的根磷分配比与对照间差异显著(P<0.05)。除N1P2处理外,其余各施肥处理的茎磷分配比与对照间无显著差异(P>0.05)。除P1、N1、N2P2处理外,其余各施肥处理的叶磷分配比与对照间差异显著(P<0.05)。各施肥处理的根磷分配比较对照均有不同程度的增加,茎磷分配比中有6个处理较对照有不同程度的增加,而叶磷分配比较对照均有不同程度的减少。
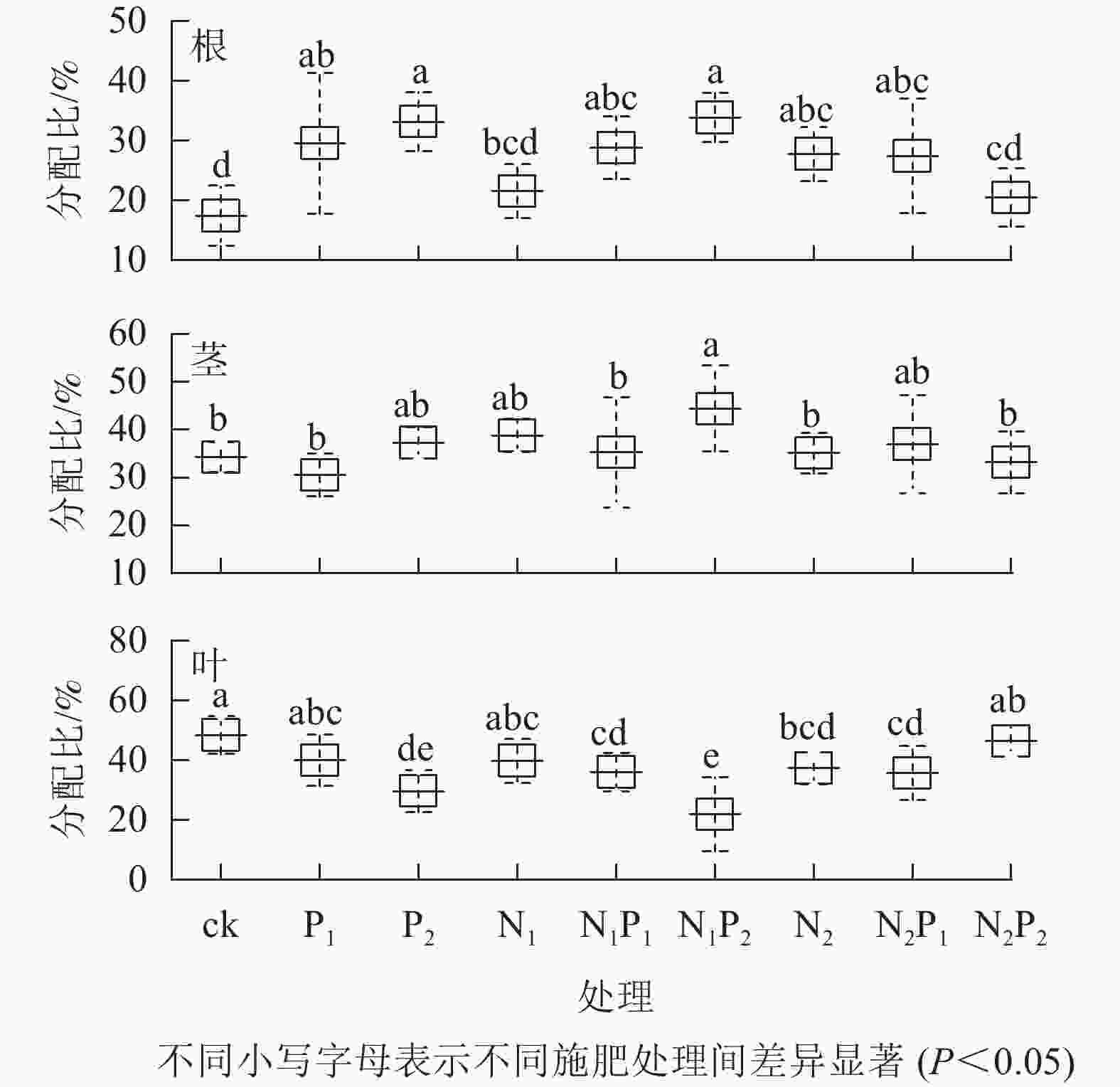
图 4 不同施肥处理云南松苗木各器官磷元素分配情况
Figure 4. Phosphorus distribution to each organ in P. yunnanensis seedlings under different fertilization treatments
由图5可知:除N1、N2P2处理外,其余各施肥处理的根钾分配比与对照间均差异显著(P<0.05)。各施肥处理的茎钾分配比与对照间无显著差异(P>0.05)。除P1、N1、N2P2处理外,其余各施肥处理的叶钾分配比与对照间差异显著(P<0.05)。各施肥处理的根钾分配比较对照均有不同程度的增加,茎钾分配比中有5个处理较对照有不同程度的增加,而叶钾分配比较对照均有不同程度的减少。
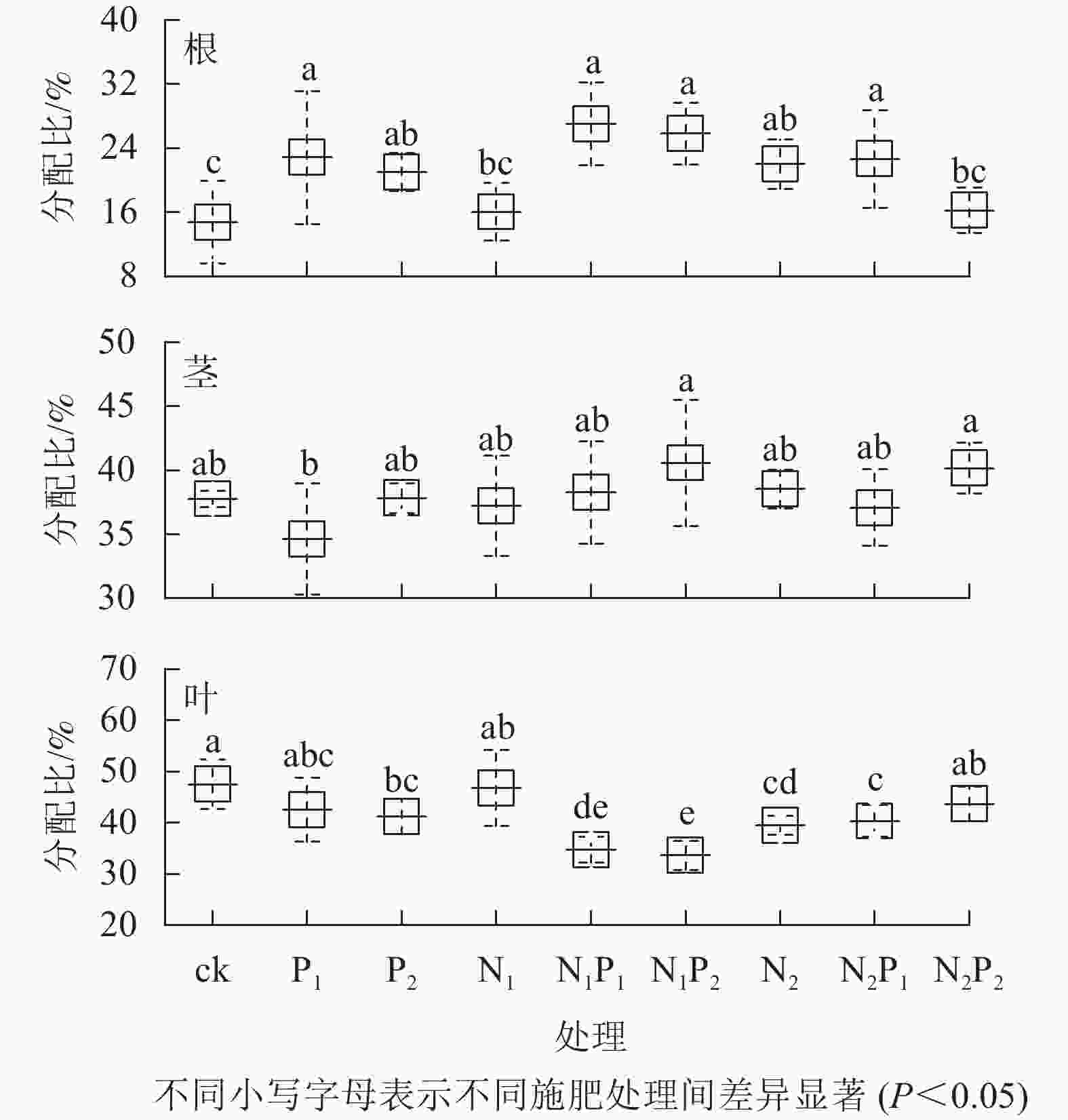
图 5 不同施肥处理云南松苗木各器官钾元素分配情况
Figure 5. Potassium distribution to each organ in P. yunnanensis seedlings under different fertilization treatments
-
由表3可知:苗木生长及养分指标与肥料用量回归方程的相关系数总体上较大,拟合程度较高,可以用来预测不同氮磷施肥处理下的云南松苗木生长及养分指标。
表 3 云南松苗木生长及养分储量指标对应的回归方程
Table 3. Regression equation of seedling growth and nutrient storage indices of P. yunnanensis seedlings
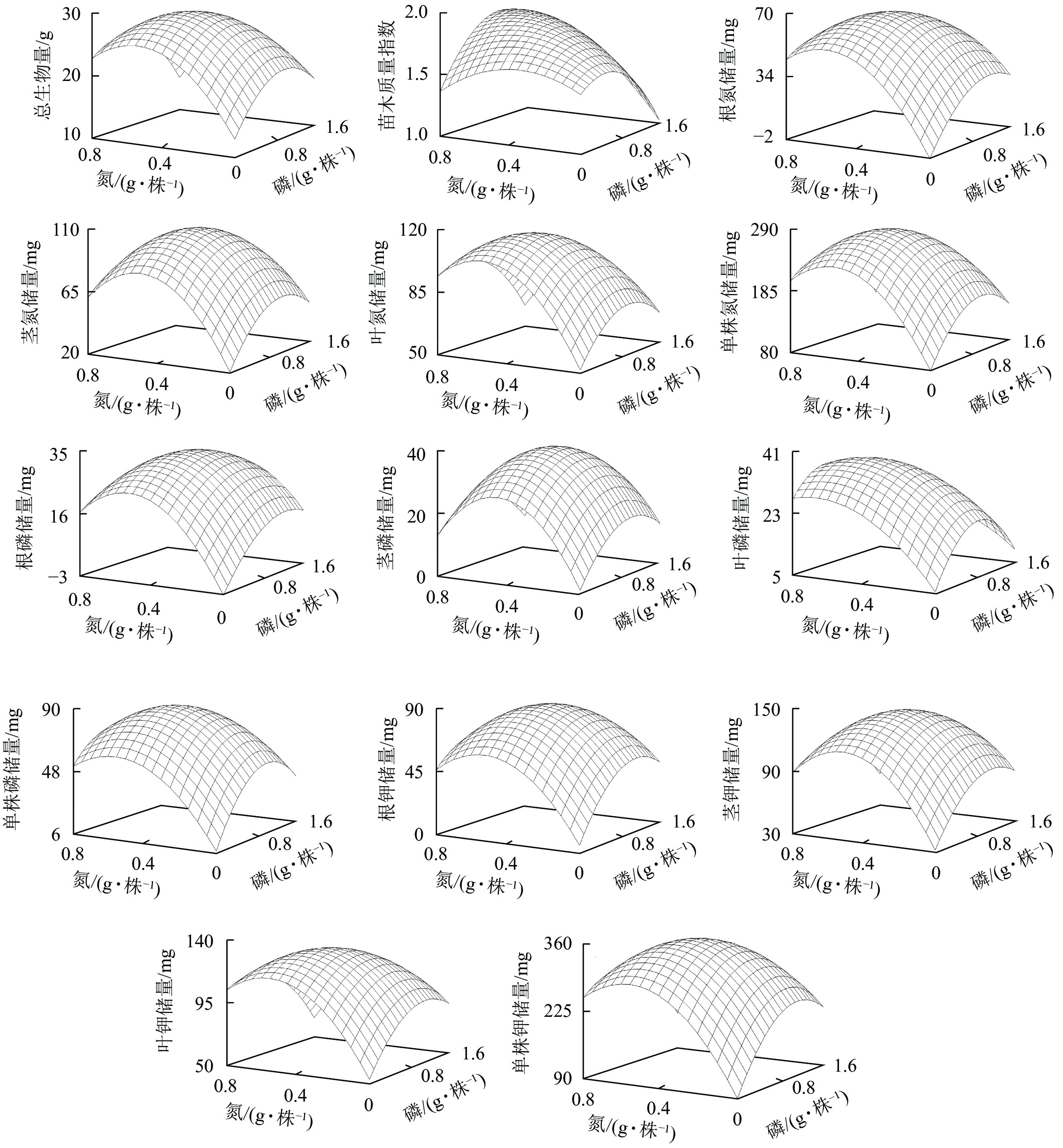
指标(Y) 回归方程 相关系数 F P 总生物量 Y=12.68+46.59XN+16.65XP−42.49XN 2−8.42XP 2−9.51XNP 0.83 5.44 <0.01 苗木质量指数 Y=1.46+0.81XN+0.43XP−1.16XN 2−0.44XP 2+0.69XNP 0.58 1.22 <0.01 根氮储量 Y=−2.05+184.14XN+57.98XP−158.49XN 2−24.09XP 2−34.70XNP 0.85 6.00 <0.01 茎氮储量 Y=20.63+243.73XN+76.14XP−241.44XN 2−36.23XP 2−32.72XNP 0.88 8.53 <0.01 叶氮储量 Y=51.79+192.21XN+54.12XP−174.42XN 2−27.80XP 2−36.28XNP 0.91 10.98 <0.01 单株氮储量 Y=79.08+555.41XN+184.83XP−502.92XN 2−90.00XP 2−99.74XNP 0.89 8.82 <0.01 根磷储量 Y=−3.27+101.64XN+29.62XP−96.23XN 2−11.87XP 2−16.89XNP 0.84 5.63 <0.01 茎磷储量 Y=1.33+111.55XN+33.07XP−121.70XN 2−16.04XP 2−10.58XNP 0.90 10.23 <0.01 叶磷储量 Y=5.45+64.52XN+35.69XP−47.76XN 2−21.00XP 2−12.75XNP 0.76 13.48 <0.01 单株磷储量 Y=7.11+189.91XN+90.68XP−168.06XN 2−44.73XP 2−38.49XNP 0.88 7.94 <0.01 根钾储量 Y=5.53+206.35XN+84.05XP−194.21XN 2−37.30XP 2−37.67XNP 0.78 3.78 <0.01 茎钾储量 Y=32.12+313.49XN+100.23XP−303.82XN 2−43.87XP 2−57.39XNP 0.84 5.55 <0.01 叶钾储量 Y=53.24+222.39XN+75.10XP−198.15XN 2−33.85XP 2−56.67XNP 0.83 5.37 <0.01 单株钾储量 Y=89.88+701.11XN+269.15XP−624.29XN 2−121.42XP 2−161.13XNP 0.83 5.45 <0.01 说明:XN、XP、XNP分别表示氮水平、磷水平、氮磷交互水平 根据表3的回归方程,可得到产量效应曲面图(图6)。由图6可知:各生长指标反应曲面均呈单峰曲面模式,即各生长指标均存在一个最大值。峰值以前,各生长指标均随氮磷用量的增加而增加。峰值以后,各生长指标随氮磷用量的增加而下降。
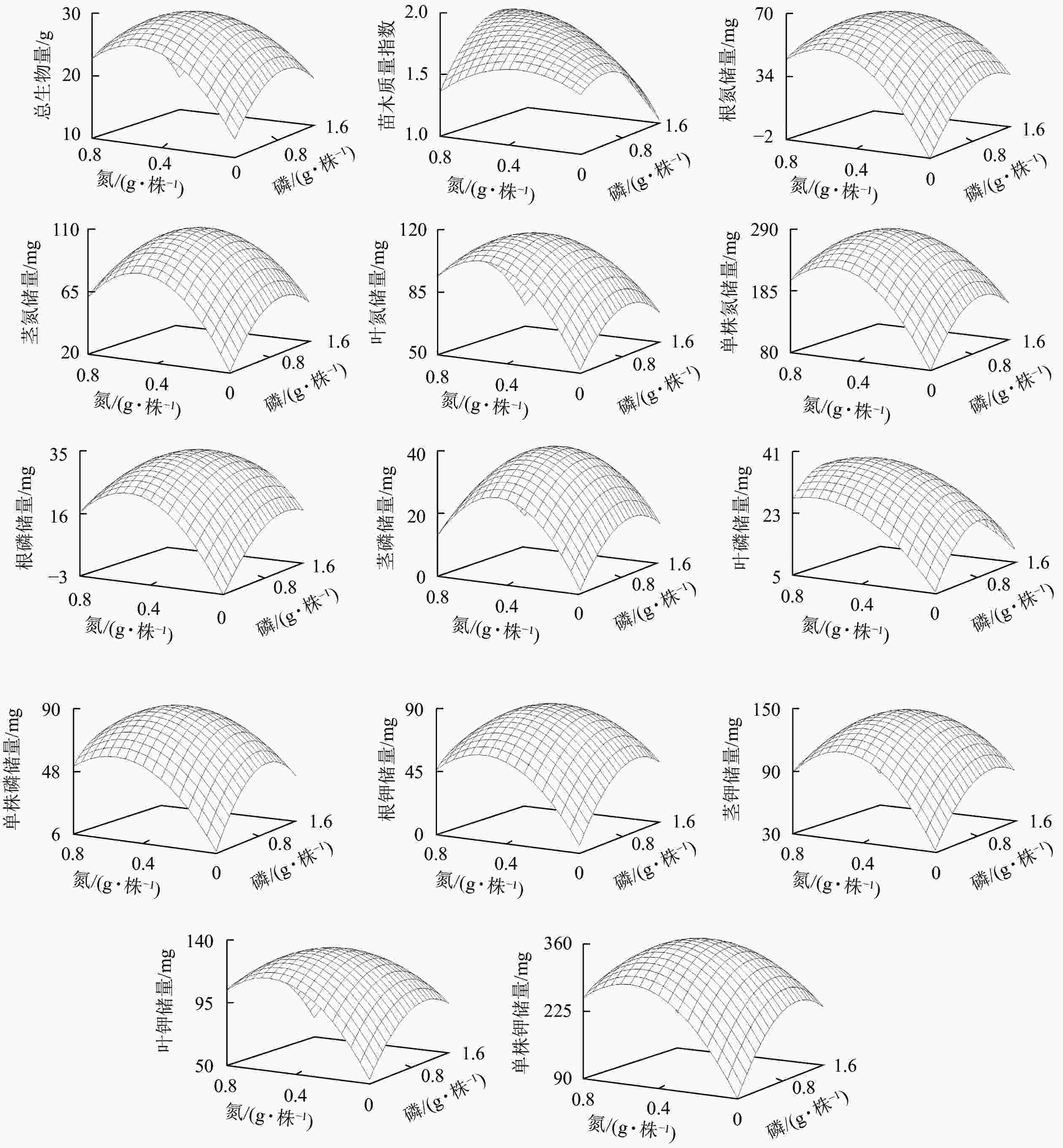
图 6 云南松苗木生物量及养分储量对氮磷配施的效应曲面
Figure 6. Effect surface of biomass and nutrient storage of P. yunnanensis seedlings on nitrogen and phosphorus combined application
由表4可知:施肥处理后,氮磷交互产量大于单施氮产量,单施氮产量优于单施磷产量。因此,在本研究中,针对2年生云南松苗木的肥料效应而言,氮磷配施的效果优于单施氮肥,单施氮肥的效果优于单施磷肥。
表 4 云南松苗木各生长指标单施氮肥、磷肥及氮磷交互的最高产量
Table 4. Highest yield in single-factor of nitrogen or phosphorus and nitrogen and phosphorus interaction with each growth index of P. yunnanensis seedlings
指标(Y) 生长指标与单施氮肥关系 生长指标与单施磷肥关系 氮磷交互最高产量 回归方程 最高产量 回归方程 最高产量 总生物量/g Y=12.68+46.59XN−42.49XN 2 25.17 Y=12.68+16.65XP−8.42XP 2 20.33 29.59 苗木质量指数 Y=1.46+0.81XN−1.16XN 2 1.50 Y=1.46+0.43XP−0.44XP 2 1.45 1.94 根氮储量/mg Y=−2.05+184.14XN−158.49XN 2 50.05 Y=−2.05+57.98XP−24.09XP 2 29.86 67.55 茎氮储量/mg Y=20.63+243.73XN−241.44XN 2 81.34 Y=20.63+76.14XP−36.23XP 2 59.15 107.04 叶氮储量/mg Y=51.79+192.21XN−174.42XN 2 103.92 Y=51.79+54.12XP−27.80XP 2 75.38 115.98 单株氮储量/mg Y=79.08+555.41XN−502.92XN 2 229.55 Y=79.08+184.83XP−90.00XP 2 167.71 281.92 根磷储量/mg Y=−3.27+101.64XN−96.23XN 2 22.93 Y=−3.27+29.62XP−11.87XP 2 14.01 33.20 茎磷储量/mg Y=1.33+111.55XN−121.70XN 2 26.71 Y=1.33+33.07XP−16.04XP 2 18.06 39.48 叶磷储量/mg Y=5.45+64.52XN−47.76XN 2 26.86 Y=5.45+35.69XP−21.00XP 2 19.95 34.47 单株磷储量/mg Y=7.11+189.91XN−168.06XN 2 59.31 Y=7.11+90.68XP−44.73XP 2 51.23 88.69 根钾储量/mg Y=5.53+206.35XN−194.21XN 2 58.85 Y=5.53+84.05XP−37.30XP 2 50.99 89.24 茎钾储量/mg Y=32.12+313.49XN−303.82XN 2 110.99 Y=32.12+100.23XP−43.87XP 2 85.81 143.27 叶钾储量/mg Y=53.24+222.39XN−198.15XN 2 113.50 Y=53.24+75.10XP−33.85XP 2 89.97 131.36 单株钾储量/mg Y=89.88+701.11XN−624.29XN 2 279.99 Y=89.88+269.15XP−121.42XP 2 227.84 358.61 说明:XN表示氮水平,XP表示磷水平 对表3构建的回归方程求偏导,确定了云南松苗木各项生长指标适宜的氮磷施肥量。由表5可知:2年生云南松苗木适宜的氮磷施肥量分别为氮0.42~0.65 g·株−1,磷0.66~1.01 g·株−1,适宜的氮磷配比为1.00∶1.15~1.00∶2.13。
表 5 2年生云南松苗木在指标最高产量下的适宜施肥量
Table 5. Suitable fertilizer amount for two years old P. yunnanensis seedlings under highest yield
指标 适宜施肥量 最佳氮磷比 氮/(g·株−1) 磷/(g·株−1) 总生物量 0.47 0.73 1.00∶1.55 苗木质量指数 0.65 1.01 1.00∶1.56 根氮储量 0.49 0.85 1.00∶1.75 茎氮储量 0.45 0.85 1.00∶1.90 叶氮储量 0.48 0.66 1.00∶1.37 单株氮储量 0.48 0.76 1.00∶1.60 根磷储量 0.45 0.93 1.00∶2.08 茎磷储量 0.42 0.89 1.00∶2.13 叶磷储量 0.59 0.67 1.00∶1.15 单株磷储量 0.48 0.81 1.00∶1.72 根钾储量 0.45 0.90 1.00∶2.03 茎钾储量 0.44 0.86 1.00∶1.97 叶钾储量 0.46 0.73 1.00∶1.59 单株钾储量 0.46 0.80 1.00∶1.76 适宜范围 0.42~0.65 0.66~1.01 1.00∶1.15~1.00∶2.13 -
生物量综合反映了苗木的各项生长指标,施肥对苗木生长的影响可以通过其对生物量的影响来反映[16]。本研究发现:施肥显著促进了苗木生物量的积累。这与施肥显著提高紫椴Tilia amurensis[17]、闽楠Phoebe bournei [3]苗木生物量的研究结果相似,说明施肥促进了苗木生长。苗木质量是衡量苗木生活力强弱的重要指标,关系到苗木成活、森林郁闭、森林经济和生态效益的发挥。造林后,苗木的存活及生长与苗木初始质量息息相关[18]。苗木质量受到施肥调控的影响,如周磊等[15]对细叶桢楠Phoebe hui容器苗进行施肥研究发现:施肥提高了细叶桢楠的苗木质量。罗仙英等[19]研究表明:施肥提高了马尾松的苗木质量。另外,苗木质量在一定程度上还受到土壤基质的影响,但有些研究却提出了不同的观点,认为土壤基质对苗木质量的影响并不大,如刘婷岩等[20]对不同基质与施肥处理下白桦Betula platyhylla的苗木质量进行研究发现:土壤基质对白桦苗木质量指数无显著影响。莎茹等[21]研究表明:在低水平施肥情况下,土壤基质对樟子松Pinus sylvestris var. mongolica苗木质量指标的影响不明显。邱琼等[22]研究表明:土壤基质对山桂花Paramichelia baillonii容器苗质量无显著影响,不同基质处理下苗木的根生物量、主根长、侧根数等苗木质量指标之间均无显著差异。而本研究在土壤基质一致的情况下,发现各施肥处理下的生物量、苗木质量均大于对照,说明施肥是提高苗木质量,促进苗木生长的有效途径之一。
施肥处理后苗木体内的养分累积有助于提高苗木的抗性和竞争力,从而提高造林成功的可能性[23]。本研究发现:云南松苗木氮磷钾元素主要储存在叶内,总体上养分储量从大到小依次为叶、茎、根。这与王光如等[24]、刘广路等[25]的研究结果类似。植物各器官基本的生理需求不同,组织结构功能的分化导致氮磷钾储量的差异[26]。植物的叶片是植物的同化器官,也是重要的养分储存器官,而茎作为植物养分的输送通道,较少储存养分,进而导致植物叶的氮磷储量高于茎[27]。本研究发现:不同施肥处理的各器官及单株氮磷钾储量总体上均高于对照,说明施肥对苗木养分积累起到明显促进作用。王益明等[28]对山核桃Carya cathayensis幼苗进行施肥研究表明:苗木各器官的氮积累量均显著高于对照组,常笑超等[29]对毛白杨Populus tomentosa的苗木施肥研究也得出:硝态氮能显著促进毛白杨的养分积累,可见氮磷添加促进了植物对营养元素的吸收[30]。在N1P1处理下,云南松苗木器官与单株储量总体最大,说明氮磷配施处理对养分积累的促进效果最佳。同时,也表明在适宜的施肥范围内,各项生长指标随施肥量的增加而增长,而超过一定的施肥量范围,则会抑制苗木生长。
营养元素在不同器官的分配是遗传差异与环境共同作用的结果,植物器官结构和功能差异影响到氮磷钾等营养元素在植物体内的分配[31],而养分元素在不同器官的合理分配是植物对外界环境变化响应与适应的重要机制。对于植物来说,叶及根分别是地上和地下的主要营养器官,茎则是植物各种物质的转运器官,叶、根与茎将植物的地上和地下部分联系起来[32]。本研究发现:苗木分配给叶的营养元素最多,其次为茎,分配给根的较少,这与王维奇等[33]的研究结果相似。这阐释了资源在各功能器官之间的分配权衡,将更多的营养物质分配给叶和茎,以保证物质充分生产维持苗木正常生长,同时增强苗木对资源以及空间的竞争能力[34]。施肥会使植物的养分分配格局发生改变,影响幼苗的养分平衡。施侃侃等[35]对红花玉兰Magnolia wufengensis幼苗养分分配格局研究发现:不同的氮磷钾配施会改变红花玉兰幼苗各营养器官内养分的分配。常云妮等[36]对华润楠Machilus chinensis苗木养分分配格局研究发现:氮添加改变了氮磷在各器官中的分配比例。本研究也得到类似的结论,即施肥改变了苗木各器官的营养元素分配格局,增大了对根和茎的养分分配,减小了对叶的养分分配。根系是植物吸收水分和养分的主要器官,担负着供给地上部分养分的任务[37]。当苗木生存的养分环境改变时,将更多的资源分配到根和茎,以扩大营养吸收范围,获取更多的水分、养分资源及增强对空间的竞争。同时,在根充分吸收养分和生长后,又可为相对较小的地上部分生长提供所需要的营养与能量,加大对环境资源的分享。这样不仅使有限的环境资源得到最大化的利用,也保证个体的存活与个体的生长,减少营养缺失对云南松幼苗生长的限制,增大了对不良环境的抵抗能力。因此,根系高比例的养分投资是云南松苗木的生态对策之一。
构建苗木养分指标与肥料用量的回归方程发现:氮磷配施的效果优于单施氮肥,单施氮肥的效果优于单施磷肥。林婉奇等[11]对樟树幼苗进行施肥发现:氮磷配施对幼苗生长的促进效果最好。刘斌等[38]对九连山次生阔叶林幼苗生长进行施肥发现:与对照相比,氮磷配施对苗木的冠幅增长率促进效果最佳,氮添加次之,磷添加抑制幼苗生长,这与本研究结果类似。根据回归方程求得2年生云南松苗木适宜的氮磷施用量分别为氮0.42~0.65 g·株−1,磷0.66~1.01 g·株−1,适宜的氮磷配比为1.00∶1.15~1.00∶2.13。从苗木的质量、生物量及养分积累量考虑,N1P1处理为本研究最优氮磷配施组合。
Effects of different nitrogen and phosphorus combinations on seedling growth and nutrient accumulation of Pinus yunnanensis seedlings
-
摘要:
目的 开展氮磷配施,揭示云南松Pinus yunnanensis苗木生长及养分积累对氮磷配施的响应,为优化云南松苗期养分管理及苗木培育提供理论依据。 方法 以2年生云南松盆栽苗为研究对象,设置对照(不施肥)、磷肥0.8 g·株−1(P1)、磷肥1.6 g·株−1(P2)、氮肥0.4 g·株−1 (N1)、氮肥0.8 g·株−1(N2)、氮肥0.4 g·株−1和磷肥0.8 g·株−1(N1P1)、氮肥0.4 g·株−1和磷肥1.6 g·株−1 (N1P2)、氮肥0.8 g·株−1和磷肥0.8 g·株−1 (N2P1)、氮肥0.8 g·株−1和磷肥1.6 g·株−1 (N2P2)等9个不同施肥处理,分析不同施肥对云南松苗木生长及养分积累的影响。 结果 各施肥处理的总生物量、苗木质量均大于对照组,N1P1处理下苗木总生物量和苗木质量最大。各器官的养分积累从大到小依次为叶、茎、根。各施肥处理的器官及单株氮、磷、钾储量与对照组之间总体上均具有显著差异(P<0.05),较对照组均有不同程度增加。N1P1处理下苗木根、茎、单株的氮、钾储量最大,根、茎、叶、单株的磷储量最大,N2P1处理下叶的氮、钾储量最大。氮、磷、钾养分分配格局从大到小依次为叶、茎、根。 结论 施肥促进了云南松苗木的生物量积累,提高了苗木质量,促进了苗木各器官的养分积累,并增大了根、茎的养分分配,减小了对叶的养分分配。氮磷配施的效果优于单施氮肥,单施氮肥的效果优于单施磷肥,N1P1处理为最优配施组合。图6表5参38 Abstract:Objective This study tries to reveal the response of Pinus yunnanensis seedling growth and nutrient accumulation to exogenous N and P combined application, and to provide a theoretical basis for optimizing nutrient management and seedling cultivation of P. yunnanensis. Method 2-year-old potted seedlings of P. yunnanensis were taken as the research objects. 9 different fertilization treatments were set to analyze the effects of different fertilization treatments on the growth and nutrient accumulation of P. yunnanensis seedlings, including ck (no fertilization), P1 (P, 0.8 g per seedling), P2 (P, 1.6 g per seedling), N1 (N, 0.4 g per seedling), N2 (N, 0.8 g per seedling), N1P1 (N, 0.4 g per seedling, P, 0.8 g per seedling), N1P2 (N, 0.4 g per seedling, P, 1.6 g per seedling), N2P1 (N, 0.8 g per seedling, P, 0.8 g per seedling), N2P2 (N, 0.8 g per seedling, P, 1.6 g per seedling). Result The total biomass and seedling quality of each fertilization treatment were higher than those of ck, and those of N1P1 treatment were the highest. The nutrient accumulation of each organ ranging from large to small was leaf, stem and root. The N, P and K reserves of organs and individual plants of different fertilization treatments were significantly different from those of ck (P<0.05), and increased in different degrees compared with ck. The seedlings under N1P1 treatment had the largest N and K reserves in root, stem and individual plant, and the largest P reserves in root, stem, leaf and individual plant, while the seedlings under N2P1 treatment had the largest N and K reserves in leaf. The distribution pattern of N, P and K nutrients was leaf, stem and root from large to small. Conclusion N and P combined application significantly improves biomass accumulation, seedling quality, as well as nutrient accumulation in each organ. It also increases the nutrient distribution of root and stem, and decreases the nutrient distribution of leaf. The combined application of N and P is superior to the single application of either N or P. Therefore, N1P1 is the optimal treatment. [Ch, 6 fig. 5 tab. 38 ref.] -
Key words:
- Pinus yunnanensis /
- proportional fertilization /
- biomass /
- nutrient accumulation
-
图 1 氮磷配施处理下云南松苗木生物量和质量指数的变化
Figure 1 Change of biomass and quality index of P. yunnanensis seedlings under nitrogen and phosphorus combined application treatments
图 2 云南松苗木各器官养分分配总体情况
Figure 2 Nutrient distribution of different organs of P. yunnanensis seedlings
图 3 不同施肥处理云南松苗木各器官氮元素分配情况
Figure 3 Nitrogen distribution to each organ in P. yunnanensis seedlings under different fertilization treatments
图 4 不同施肥处理云南松苗木各器官磷元素分配情况
Figure 4 Phosphorus distribution to each organ in P. yunnanensis seedlings under different fertilization treatments
图 5 不同施肥处理云南松苗木各器官钾元素分配情况
Figure 5 Potassium distribution to each organ in P. yunnanensis seedlings under different fertilization treatments
图 6 云南松苗木生物量及养分储量对氮磷配施的效应曲面
Figure 6 Effect surface of biomass and nutrient storage of P. yunnanensis seedlings on nitrogen and phosphorus combined application
表 1 外源性氮磷添加施肥试验组合
Table 1. Fertilization test combination of exogenous nitrogen and phosphorus addition
处理 氮/(g·株−1) 磷/(g·株−1) 处理 氮/(g·株−1) 磷/(g·株−1) ck 0 0 N1P1 0.4 0.8 P1 0 0.8 N1P2 0.4 1.6 P2 0 1.6 N2P1 0.8 0.8 N1 0.4 0 N2P2 0.8 1.6 N2 0.8 0  下载: 导出CSV
下载: 导出CSV
表 2 不同施肥处理下云南松苗木养分积累情况
Table 2. Nutrient reserves of P. yunnanensis seedlings under different fertilization treatments
元素 处理 养分储量/mg 根 茎 叶 单株 氮 ck 15.92±11.32 e 33.55±11.64 e 54.94±13.16 e 104.41±35.89 g P1 32.31±18.05 cd 43.50±4.01d e 70.12±8.67 de 145.93±23.64 ef P2 33.85±9.42 cd 55.09±9.11 cd 83.72±8.85 cd 172.65±22.19 de N1 28.62±7.34 de 65.59±8.65 c 104.55±16.30 ab 198.76±16.83 cd N1P1 73.75±17.31 a 103.55±13.97 a 112.53±7.19 a 289.83±22.25 a N1P2 62.52±13.10 ab 94.83±14.29 ab 93.20±12.34 bc 250.55±33.61 ab N2 47.73±11.35 bc 85.20±26.98 b 101.10±21.08 ab 234.03±55.79 bc N2P1 56.37±10.90 b 101.48±18.57 ab 117.79±16.37 a 275.64±33.02 a N2P2 22.17±11.52 de 41.76±9.47 de 60.17±10.77 e 124.10±31.07 fg 磷 ck 3.91±2.62 e 7.15±3.25 e 9.53±2.52 d 20.60±8.27 f P1 10.65±6.78 de 10.08±0.68 de 13.09±1.74 d 33.82±6.90 def P2 13.73±4.37 cd 15.39±4.01 de 12.19±4.07 d 41.31±9.69 de N1 10.56±2.66 de 18.86±2.79 cd 19.39±4.64 cd 48.82±6.95 d N1P1 33.47±10.56 a 38.81±8.12 a 41.7±12.34 a 113.99±18.37 a N1P2 25.93±6.81 b 33.94±10.66 ab 15.98±8.46 d 75.84±13.22 c N2 20.38±7.71 bc 26.53±11.81 bc 26.64±7.72 bc 73.55±25.80 d N2P1 24.63±6.57 ab 34.50±11.03 ab 33.87±13.15 ab 93.00±14.96 b N2P2 6.04±3.14 e 9.14±2.02 e 12.97±3.49 d 28.14±7.71 ef 钾 ck 22.05±15.42 e 50.84±18.39 e 62.14±16.46 f 135.03±50.20 d P1 41.56±22.67 de 59.53±4.21 de 73.14±8.54 ef 174.23±27.31 cd P2 47.57±18.06 cd 84.21±23.45 c 90.03±16.06 de 221.82±55.86 c N1 34.43±8.15 de 79.80±9.55 cd 100.70±19.38 cd 214.94±21.10 c N1P1 102.27±18.50 a 145.46±18.96 a 131.71±9.79 ab 379.44±13.10 a N1P2 82.64±25.07 ab 127.24±20.30 ab 105.62±18.34 cd 315.50±53.39 b N2 63.79±18.56 bc 112.87±33.73 b 114.98±31.44 bc 291.64±80.58 b N2P1 75.91±19.25 b 125.61±19.41 ab 136.33±19.13 a 337.85±34.32 ab N2P2 23.97±7.92 e 58.04±8.15 de 62.89±8.74 f 144.90±22.24 d 说明:数据为平均值±标准差。同列不同小写字母表示同一养分元素同一组织在不同施肥处理间差异显著(P<0.05)
下载: 导出CSV
表 3 云南松苗木生长及养分储量指标对应的回归方程
Table 3. Regression equation of seedling growth and nutrient storage indices of P. yunnanensis seedlings
指标(Y) 回归方程 相关系数 F P 总生物量 Y=12.68+46.59XN+16.65XP−42.49XN 2−8.42XP 2−9.51XNP 0.83 5.44 <0.01 苗木质量指数 Y=1.46+0.81XN+0.43XP−1.16XN 2−0.44XP 2+0.69XNP 0.58 1.22 <0.01 根氮储量 Y=−2.05+184.14XN+57.98XP−158.49XN 2−24.09XP 2−34.70XNP 0.85 6.00 <0.01 茎氮储量 Y=20.63+243.73XN+76.14XP−241.44XN 2−36.23XP 2−32.72XNP 0.88 8.53 <0.01 叶氮储量 Y=51.79+192.21XN+54.12XP−174.42XN 2−27.80XP 2−36.28XNP 0.91 10.98 <0.01 单株氮储量 Y=79.08+555.41XN+184.83XP−502.92XN 2−90.00XP 2−99.74XNP 0.89 8.82 <0.01 根磷储量 Y=−3.27+101.64XN+29.62XP−96.23XN 2−11.87XP 2−16.89XNP 0.84 5.63 <0.01 茎磷储量 Y=1.33+111.55XN+33.07XP−121.70XN 2−16.04XP 2−10.58XNP 0.90 10.23 <0.01 叶磷储量 Y=5.45+64.52XN+35.69XP−47.76XN 2−21.00XP 2−12.75XNP 0.76 13.48 <0.01 单株磷储量 Y=7.11+189.91XN+90.68XP−168.06XN 2−44.73XP 2−38.49XNP 0.88 7.94 <0.01 根钾储量 Y=5.53+206.35XN+84.05XP−194.21XN 2−37.30XP 2−37.67XNP 0.78 3.78 <0.01 茎钾储量 Y=32.12+313.49XN+100.23XP−303.82XN 2−43.87XP 2−57.39XNP 0.84 5.55 <0.01 叶钾储量 Y=53.24+222.39XN+75.10XP−198.15XN 2−33.85XP 2−56.67XNP 0.83 5.37 <0.01 单株钾储量 Y=89.88+701.11XN+269.15XP−624.29XN 2−121.42XP 2−161.13XNP 0.83 5.45 <0.01 说明:XN、XP、XNP分别表示氮水平、磷水平、氮磷交互水平
下载: 导出CSV
表 4 云南松苗木各生长指标单施氮肥、磷肥及氮磷交互的最高产量
Table 4. Highest yield in single-factor of nitrogen or phosphorus and nitrogen and phosphorus interaction with each growth index of P. yunnanensis seedlings
指标(Y) 生长指标与单施氮肥关系 生长指标与单施磷肥关系 氮磷交互最高产量 回归方程 最高产量 回归方程 最高产量 总生物量/g Y=12.68+46.59XN−42.49XN 2 25.17 Y=12.68+16.65XP−8.42XP 2 20.33 29.59 苗木质量指数 Y=1.46+0.81XN−1.16XN 2 1.50 Y=1.46+0.43XP−0.44XP 2 1.45 1.94 根氮储量/mg Y=−2.05+184.14XN−158.49XN 2 50.05 Y=−2.05+57.98XP−24.09XP 2 29.86 67.55 茎氮储量/mg Y=20.63+243.73XN−241.44XN 2 81.34 Y=20.63+76.14XP−36.23XP 2 59.15 107.04 叶氮储量/mg Y=51.79+192.21XN−174.42XN 2 103.92 Y=51.79+54.12XP−27.80XP 2 75.38 115.98 单株氮储量/mg Y=79.08+555.41XN−502.92XN 2 229.55 Y=79.08+184.83XP−90.00XP 2 167.71 281.92 根磷储量/mg Y=−3.27+101.64XN−96.23XN 2 22.93 Y=−3.27+29.62XP−11.87XP 2 14.01 33.20 茎磷储量/mg Y=1.33+111.55XN−121.70XN 2 26.71 Y=1.33+33.07XP−16.04XP 2 18.06 39.48 叶磷储量/mg Y=5.45+64.52XN−47.76XN 2 26.86 Y=5.45+35.69XP−21.00XP 2 19.95 34.47 单株磷储量/mg Y=7.11+189.91XN−168.06XN 2 59.31 Y=7.11+90.68XP−44.73XP 2 51.23 88.69 根钾储量/mg Y=5.53+206.35XN−194.21XN 2 58.85 Y=5.53+84.05XP−37.30XP 2 50.99 89.24 茎钾储量/mg Y=32.12+313.49XN−303.82XN 2 110.99 Y=32.12+100.23XP−43.87XP 2 85.81 143.27 叶钾储量/mg Y=53.24+222.39XN−198.15XN 2 113.50 Y=53.24+75.10XP−33.85XP 2 89.97 131.36 单株钾储量/mg Y=89.88+701.11XN−624.29XN 2 279.99 Y=89.88+269.15XP−121.42XP 2 227.84 358.61 说明:XN表示氮水平,XP表示磷水平
下载: 导出CSV
表 5 2年生云南松苗木在指标最高产量下的适宜施肥量
Table 5. Suitable fertilizer amount for two years old P. yunnanensis seedlings under highest yield
指标 适宜施肥量 最佳氮磷比 氮/(g·株−1) 磷/(g·株−1) 总生物量 0.47 0.73 1.00∶1.55 苗木质量指数 0.65 1.01 1.00∶1.56 根氮储量 0.49 0.85 1.00∶1.75 茎氮储量 0.45 0.85 1.00∶1.90 叶氮储量 0.48 0.66 1.00∶1.37 单株氮储量 0.48 0.76 1.00∶1.60 根磷储量 0.45 0.93 1.00∶2.08 茎磷储量 0.42 0.89 1.00∶2.13 叶磷储量 0.59 0.67 1.00∶1.15 单株磷储量 0.48 0.81 1.00∶1.72 根钾储量 0.45 0.90 1.00∶2.03 茎钾储量 0.44 0.86 1.00∶1.97 叶钾储量 0.46 0.73 1.00∶1.59 单株钾储量 0.46 0.80 1.00∶1.76 适宜范围 0.42~0.65 0.66~1.01 1.00∶1.15~1.00∶2.13
下载: 导出CSV
-
[1] 孙慧彦, 刘勇, 马履一, 等. 长白落叶松苗木质量与造林效果关系的比较[J]. 北京林业大学学报, 2009, 31(6): 176 − 180. SUN Huiyan, LIU Yong, MA Lüyi, et al. Relationship between seedling quality and outplanting performance of Larix olgensis [J]. Journal of Beijing Forestry University, 2009, 31(6): 176 − 180. [2] 荣俊冬, 凡莉莉, 陈礼光, 等. 不同施氮模式和施氮量对福建柏幼苗生物量分配和根系生长的影响[J]. 林业科学, 2020, 56(7): 175 − 184. RONG Jundong, FAN Lili, CHEN Liguang, et al. Impacts on biomass allocation and root growth of Fokienia hodginsii seedlings of different patterns and quantities of nitrogen application [J]. Scientia Silvae Sinicae, 2020, 56(7): 175 − 184. [3] 王晓, 王樱琳, 韦小丽, 等. 不同指数施氮量对闽楠幼苗生长生理及养分积累的影响[J]. 浙江农林大学学报, 2020, 37(3): 514 − 521. WANG Xiao, WANG Yinlin, WEI Xiaoli, et al. Growth, physiology and nutrient accumulation of Phoebe bournei seedlings under different amount of exponential nitrogen fertilization [J]. Journal of Zhejiang A&F University, 2020, 37(3): 514 − 521. [4] 陈亚南, 马露莎, 张向茹, 等. 陕西黄土高原刺槐枯落叶生态化学计量学特征[J]. 生态学报, 2014, 34(15): 4412 − 4422. CHEN Ya’nan, MA Lusha, ZHANG Xiangru, et al. Ecological stoichiometry characteristics of leaf litter of Robinia pseudoacacia in the Loess Plateau of Shaanxi Province [J]. Acta Ecologica Sinica, 2014, 34(15): 4412 − 4422. [5] HAN Wenxuan, FANG Jingyun, GUO Dali, et al. Leaf nitrogen and phosphorus stoichiometry across 753 terrestrial plant species in China [J]. New Phytologist, 2005, 168: 377 − 385. [6] 任书杰, 于贵瑞, 陶波, 等. 中国东部南北样带654种植物叶片氮和磷的华北计量学特征研究[J]. 环境科学, 2007, 28(12): 2665 − 2673. REN Shujie, YU Guirui, TAO Bo, et al. Leaf nitrogen and phosphorus stoichiometry across 654 terrestrial plant species in NSTEC [J]. Environmental Science, 2007, 28(12): 2665 − 2673. [7] 李莲芳, 王慷林, 段安安, 等. 昆明宜良尖山云南松林的土壤理化特性[J]. 云南农业大学学报(自然科学), 2017, 32(1): 146 − 152. LI Lianfang, WANG Kanglin, DUAN Anan, et al. Physical and chemical properties of Pinus yunnanensis forest soil in Jianshan, Yiliang, Kunming [J]. Journal of Yunnan Agricultural University, 2017, 32(1): 146 − 152. [8] 李亚麒, 孙继伟, 李江飞, 等. 云南松不同家系苗木生物量分配及其异速生长[J]. 北京林业大学学报, 2021, 43(8): 18 − 28. LI Yaqi, SUN Jiwei, LI Jiangfei, et al. Biomass allocation and its allometric growth of Pinus yunnanensis seedlings of different families [J]. Journal of Beijing Forestry University, 2021, 43(8): 18 − 28. [9] 杨文君, 李莲芳, 汪亚愈, 等. 云南松种子发芽对温度和生长调节剂的响应[J]. 种子, 2017, 36(8): 10 − 15, 19. YANG Wenjun, LI Lianfang, WANG Yayu, et al. Effects of temperature and exogenous hormone presoaking seeds on seed germination of Pinus yunnanensis [J]. Seed, 2017, 36(8): 10 − 15, 19. [10] 陈琳, 曾杰, 贾宏炎, 等. 林木苗期营养诊断与施肥研究进展[J]. 世界林业研究, 2012, 25(3): 26 − 31. CHEN Lin, ZENG Jie, JIA Hongyan, et al. Advances in nutrient diagnosis and fertilization study of seedlings [J]. World Forestry Research, 2012, 25(3): 26 − 31. [11] 林婉奇, 蔡金桓, 薛立. 不同密度樟树(Cinnamomum camphora)幼苗生长和叶片性状对氮磷添加的响应[J]. 生态学报, 2019, 39(18): 6738 − 6744. LIN Wanqi, CAI Jinhuan, XUE Li. Responses of Cinnamomum camphora seedling growth and leaf traits to additions of nitrogen and phosphorous under different planting densities [J]. Acta Ecologica Sinica, 2019, 39(18): 6738 − 6744. [12] 江佳, 盛笑羚, 邵炯妮, 等. 马尾松和木荷幼苗主要功能性状对氮磷和石灰添加的响应[J]. 应用与环境生物学报, 2020, 26(2): 410 − 416. JIANG Jia, SHENG Xiaolin, SHAO Jiongni, et al. Responses of the major functional traits of Pinus massoniana and Schima superba seedlings to nitrogen, phosphorus, and lime addition [J]. Chinese Journal of Applied &Environmental Biology, 2020, 26(2): 410 − 416. [13] 张跃敏, 李根前, 李莲芳, 等. 云南松实生苗生长对N、P配施的响应[J]. 东北林业大学学报, 2009, 37(8): 5 − 8. ZHANG Yuemin, LI Genqian, LI Lianfang, et al. Fertilization effect of nitrogen combination with phosphorus on seedling growth of Pinus yunnanensis [J]. Journal of Northeast Forestry University, 2009, 37(8): 5 − 8. [14] 鲍士旦. 土壤农化分析[M]. 3版. 北京: 中国农业出版社, 2013: 13 − 54. BAO Shidan. Soil Agrochemical Analysis [M]. 3rd ed. Beijing: China Agriculture Press, 2013: 13 − 54. [15] 周磊, 刘美玲, 李铁华, 等. 施肥对细叶桢楠容器苗生长与光合的影响[J]. 中南林业科技大学学报, 2021, 41(7): 80 − 87. ZHOU Lei, LIU Meiling, LI Tiehua, et al. Effects of fertilization on growth and photosynthetic of Phoebe hui seedlings [J]. Journal of Central South University of Forestry &Technology, 2021, 41(7): 80 − 87. [16] SINGH B, SINGH G. Effects of controlled irrigation on water potential, nitrogen uptake and biomass production in Dalbergia sissoo seedlings [J]. Environmental &Experimental Botany, 2006, 55(1): 209 − 219. [17] 杨阳, 张德鹏, 及利, 等. 配比施肥对紫椴播种苗生长、养分积累及根系形态的影响[J]. 中南林业科技大学学报, 2021, 41(9): 63 − 70. YANG Yang, ZHANG Depeng, JI Li, et al. Efects of formula fertilization on growth, nutrient accumulation and root morphology of Tilia amurensis seedlings [J]. Journal of Central South University of Forestry &Technology, 2021, 41(9): 63 − 70. [18] 王益明, 卢艺, 张慧, 等. 指数施肥对美国山核桃幼苗生长及叶片养分含量的影响[J]. 中国土壤与肥料, 2018(6): 136 − 140. WANG Yiming, LU Yi, ZHANG Hui, et al. Effects of exponential fertilization on growth and foliar nutrient status of pecan seedlings [J]. Soils and Fertilizers Sciences in China, 2018(6): 136 − 140. [19] 罗仙英, 莫荣海, 丁贵杰, 等. 不同配比施肥对马尾松幼苗生长特征的影响[J]. 广西植物, 2022, 42(4): 608 − 616. LUO Xianying, MO Ronghai, DING Guijie, et al. Effect of different fertilization ratio on the growth characteristics of Pinus massoniana seedlings [J]. Guihaia, 2022, 42(4): 608 − 616. [20] 刘婷岩, 郝龙飞, 王庆成, 等. 不同轻基质和施肥处理对白桦苗木养分承载的影响[J]. 东北林业大学学报, 2019, 47(10): 16 − 19, 29. LIU Tingyan, HAO Longfei, WANG Qingcheng, et al. Nutrient loading of Betula platyhylla seedlings under different light medium and fertilization treatments [J]. Journal of Northeast Forestry University, 2019, 47(10): 16 − 19, 29. [21] 莎茹, 王洪体. 基质与施肥对樟子松幼苗培育的影响[J]. 内蒙古林业科技, 2013, 39(1): 40 − 43. SHA Ru, WANG Hongti. Effects of matrix and fertilization on seedling culture of Pinus sylvestris var. mongolica [J]. Journal of Inner Mongolia Forestry Science and Technology, 2013, 39(1): 40 − 43. [22] 邱琼, 杨德军, 钟萍, 等. 育苗基质、容器规格和施肥量对山桂花幼苗生长的影响[J]. 西部林业科学, 2018, 47(6): 131 − 135. QIU Qiong, YANG Dejun, ZHONG Ping, et al. Effects of seedling substrate, container size and fertilizer amount on the seedling growth of Paramichelia baillonii [J]. Journal of West China Forestry Science, 2018, 47(6): 131 − 135. [23] 刘欢, 王超琦, 吴家森, 等. 氮素指数施肥对1年生杉木苗生长及氮素积累的影响[J]. 浙江农林大学学报, 2017, 34(3): 459 − 464. LIU Huan, WANG Chaoqi, WU Jiasen, et al. Growth and nitrogen accumulation in seedlings of Cunninghamia lanceolata clones with nitrogen exponential fertilization [J]. Journal of Zhejiang A&F University, 2017, 34(3): 459 − 464. [24] 王光如, 冯火炬, 樊燕, 等. 江西瑞昌石灰岩山地淡竹林植物元素分布与吸收特征[J]. 广西植物, 2019, 39(2): 152 − 160. WANG Guangru, FENG Huoju, FAN Yan, et al. Element distribution pattern in Phyllostachys glauca forest and plant absorption traits in a limestone mountain of Ruichang, Jiangxi Province [J]. Guihaia, 2019, 39(2): 152 − 160. [25] 刘广路, 范少辉, 官凤英, 等. 不同年龄毛竹营养器官主要养分元素分布及与土壤环境的关系[J]. 林业科学研究, 2010, 23(2): 252 − 258. LIU Guanglu, FAN Shaohui, GUAN Fengying, et al. Distribution pattern of nutrient elements and its relationship with soil environment in different aged Phyllostachys edulis [J]. Forest Research, 2010, 23(2): 252 − 258. [26] 马任甜. 黄土高原刺槐、柠条人工林土壤-植物生态化学计量特征研究[D]. 杨凌: 西北农林科技大学, 2017. MA Rentian. Study on Soil-Plant Ecological Stoichiometry in Robinia pseudoacacia and Caragana korshinskii Plantations on the Loess Plateau[D]. Yangling: Northwest A&F University, 2017. [27] 赵亚芳, 徐福利, 王渭玲, 等. 华北落叶松根茎叶碳氮磷含量及其化学计量学特征的季节变化[J]. 植物学报, 2014, 49(5): 560 − 568. ZHAO Yafang, XU Fuli, WANG Weiling, et al. Seasonal variation in contents of C, N and P and stoichiometry characteristics in fine roots, stems and needles of Larix principis-rupprechtii [J]. Bulletin of Botany, 2014, 49(5): 560 − 568. [28] 王益明, 李瑞瑞, 张慧, 等. 指数施肥对美国山核桃幼苗生物量及氮积累的影响[J]. 生态学杂志, 2018, 37(10): 2920 − 2926. WANG Yiming, LI Ruirui, ZHANG Hui, et al. Effects of exponential fertilization on biomass and nitrogen accumulation of Carya illinoensis seedlings [J]. Chinese Journal of Ecology, 2018, 37(10): 2920 − 2926. [29] 常笑超, 刘勇, 李进宇, 等. 不同形态氮素配比对雄性毛白杨苗木生长的影响[J]. 北京林业大学学报, 2018, 40(9): 63 − 71. CHANG Xiaochao, LIU Yong, LI Jinyu, et al. Effects of different nitrogen forms and ratios on growth of male Populus tomentosa seedlings [J]. Journal of Beijing Forestry University, 2018, 40(9): 63 − 71. [30] ROWE E C, SMART S M, KENNEDY V H, et al. Nitrogen deposition increases the acquisition of phosphorus and potassium by heather Calluna vulgaris [J]. Environmental Pollution, 2008, 155(2): 201 − 207. [31] AERTS R, CHAPIN F S. The mineral nutrition of wild plants revisited: a re-evaluation of processes and patterns [J]. Advances in Ecological Research, 2000, 30(8): 1 − 67. [32] ZHAO Ning, YU Guirui, HE Nianpeng, et al. Invariant allometric scaling of nitrogen and phosphorus in leaves, stems, and fine roots of woody plants along an altitudinal gradient [J]. Journal of Plant Research, 2016, 129(4): 647 − 657. [33] 王维奇, 徐玲琳, 曾从盛, 等. 闽江河口湿地互花米草入侵机制[J]. 自然资源学报, 2011, 26(11): 1900 − 1907. WANG Weiqi, XU Linglin, ZENG Congsheng, et al. Invasion mechanism of Spartina alterniflora in Minjiang River estuarine wetland [J]. Journal of Natural Resources, 2011, 26(11): 1900 − 1907. [34] SCHENK H J, JACKSON R B. Rooting depths, lateral root spreads and below ground above ground allometries of plants in waterlimited ecosystems [J]. Journal of Ecology, 2002, 90(3): 480 − 494. [35] 施侃侃, 梁栋, 贾忠奎. 不同氮磷钾配比对红花玉兰幼苗生长及养分转运的影响[J]. 中南林业科技大学学报, 2018, 38(9): 58 − 64. SHI Kankan, LIANG Dong, JIA Zhongkui, et al. Effects of different ratio of N, P, K on nutrient accumulation and translocation of Magnolia wufengensis [J]. Journal of Central South University of Forestry &Technology, 2018, 38(9): 58 − 64. [36] 常云妮, 钟全林, 程栋梁, 等. 氮添加对华润楠苗木碳氮磷化学计量特征的影响[J]. 甘肃农业大学学报, 2013, 48(5): 94 − 99. CHANG Yunni, ZHONG Quanlin, CHENG Dongliang, et al. Effects of nitrogen addition on ecological stoichiometry characteristics of Machilus chinensis [J]. Journal of Gansu Agricultural University, 2013, 48(5): 94 − 99. [37] 邱权, 潘昕, 李吉跃, 等. 速生树种尾巨桉和竹柳幼苗的光合特性和根系特征比较[J]. 中南林业科技大学学报, 2014, 34(1): 53 − 59. QIU Quan, PAN Xin, LI Jiyue, et al. Comparison on photosynthetic and roots characteristics of Eucalyptus urophylla × Eucalyptus grandis and bamboo-willow seedlings [J]. Journal of Central South University of Forestry &Technology, 2014, 34(1): 53 − 59. [38] 刘斌, 陈维, 陈伏生, 等. 九连山次生阔叶林幼苗生长对氮磷添加的响应[J]. 应用生态学报, 2020, 31(8): 2533 − 2540. LIU Bin, CHEN Wei, CHEN Fusheng, et al. Responses of seedling growth in subtropical secondary broad leaved forest to nitrogen and phosphorus addition in Jiulian Mountain, China [J]. Chinese Journal of Applied Ecology, 2020, 31(8): 2533 − 2540. -

-
链接本文:
https://zlxb.zafu.edu.cn/article/doi/10.11833/j.issn.2095-0756.20220223
 点击查看大图
点击查看大图
计量
- 文章访问数: 1816
- HTML全文浏览量: 364
- PDF下载量: 104
- 被引次数: 0
