





-
当年生枝条是木本植物进行种群空间扩展、结实和光合作用的主要部位,与种群的生长、繁殖、存活等密切相关。探讨当年生枝条的茎、叶构件之间的异速生长规律是揭示植物种群适应机制的有效途径之一[1−3]。迄今有关这方面的研究已经有很多积累,主要是通过对茎和叶回归方程斜率的显著性检验和大小的比较,开展当年生枝条茎和叶异速生长规律随环境梯度[4−6]、种间亲缘关系[7−10]或植物生活型[11−12]变化的对比研究,以验证茎对叶片光合生产的支持能力[13−17]和构件生物量的分配规律[18−19]。此类研究大多在种间水平上比较茎、叶的生长关系,进而分析其生态、生理的联系性,隐含的前提是物种的差异性和特殊性可以忽略。
对于温带落叶木本植物,难以忽略的是枝条功能的种内分化现象[20],或者称为枝条的二态性。不同类型枝条(长枝、短枝)的叶型[21]、生理生化特征[22−25]、生长特性等都可能存在差异,由此必然引起枝条不同构件间的异速生长关系发生变化,因此,不同类型枝条的茎、叶生长规律的异同,携带着进化和适应调节等重要信息。根据以往的认识,长枝功能侧重种群生长空间扩展,短枝功能侧重光合生产[26−27],具体到构件生长层面则是长枝侧重茎的生长,短枝侧重叶片的生长。但是,木本植物枝条功能分化与光照条件[28−29]以及有性繁殖过程有关,长枝、短枝的生长特征及其在种群调节中的作用可能存在环境差异性或者物种特异性,针对不同的植物种群仍需验证。
小叶白蜡Fraxinus sogdiana又名天山梣,是木樨科Oleaceae梣属Fraxinus落叶乔木,为第三纪温带阔叶林残遗树种,被2021年《国家重点保护野生植物名录》列为Ⅱ级,具有重要的科学价值和园林应用价值,目前仅在伊犁河谷喀什河河岸带尚有大面积集中分布。小叶白蜡枝条生长的独特性在于同一分枝上长枝、短枝等不同类型枝条交替发育,并且长枝和短枝都可能发育为生殖枝,呈现非常明显的枝条分化以及生长调节特征。张维等[30]和努尔买买提等[31]对小叶白蜡复叶生物量分配和表型可塑性方面进行过研究。但对小叶白蜡枝条如何发生分化、不同类型枝条的生长特征及其功能、构件生物量分配权衡等问题了解甚少。本研究分析小叶白蜡3种不同类型枝条(长枝、中枝和短枝)各构件之间的异速生长关系,通过比较异速生长方程的斜率了解不同类型枝条功能的差异性,以期为小叶白蜡种群生长调节、适应与进化等研究提供科学依据。
-
研究地点位于新疆维吾尔自治区伊犁哈萨克自治州小叶白蜡国家级自然保护区(43°42′08″~43°50′47″N,81°49′42″~82°10′37″E),该区属典型的大陆性半干旱气候,年均气温为8.5 ℃,年均降水量为330 mm,年均蒸发量为1 300~1 700 mm。保护区内地貌主要为第四纪冲积物形成的河漫滩、阶地等堆积地貌,上覆深厚的黄土层。土壤类型以灰钙土和栗钙土为主[30−31]。保护区的保护对象为原始小叶白蜡林,属河谷落叶阔叶林,乔木主要伴生密叶杨Populus talassica、天山桦Betula tianschanica等,下层草本植物以温带分布种类为主[30−31]。
-
2019年10月中旬,在保护区内小叶白蜡分布的集中地段布置样地,样地面积为50 m×50 m。取样地段的地貌为河岸带一级阶地,地势平坦,株间距约5~6 m,林木分布较为均匀。在样地内选择长势良好、均一的10株小叶白蜡成株作为目标树,胸径为(42.93±18.46) cm,树高为(12.40±1.14) m,为保证林木年龄的一致性,去除系统发育差异的影响。按不同方向在树冠外层的中、下部位随机采摘完整、健康的不同类型当年生枝条各5条,最终每个类型枝条各获得50个样品,保留其中30个相对好的枝条作为研究对象。由于小叶白蜡当年生枝条长枝和短枝既可能发生在主枝上,也可能发生在侧枝上,通过枝位无法判断枝条类型,因此,本研究通过最大叶间距判断枝条类型。小叶白蜡羽状复叶在枝条上呈螺旋状三叶轮生,具有清晰可辩的节状叶痕,可获得较准确的叶间距数据,标准如下:最大叶间距>2 cm判断为长枝,1~2 cm判断为中枝,<1 cm判断为短枝。带回室内后测量复叶片数和小叶叶面积,叶面积使用叶面积仪(LI3000C,LI-Cor,美国)测量。测定后将枝条按照叶轴、茎、叶片分开,85 ℃烘干至恒量,用0.01%电子天平称量获得生物量数据。
-
异速生长采用幂函数方程描述,并对自变量和因变量进行对数转换,获得线性方程lgy=lgb+algx,方程参数采用标准主轴回归(SMA)方法[32]进行估计,其中y为因变量,x为自变量,a为方程斜率,b为截距。本研究重点关注斜率的变化,若斜率a显著大于或小于1,则为异速生长关系,否则为同速生长关系。同时,也在3种类型枝条之间两两比较斜率差异显著性,若差异不显著,则认为斜率相同。
所有统计分析应用R语言完成,异速生长分析应用R语言smatr程序包[33]完成。所有统计分析显著水平为0.05。
-
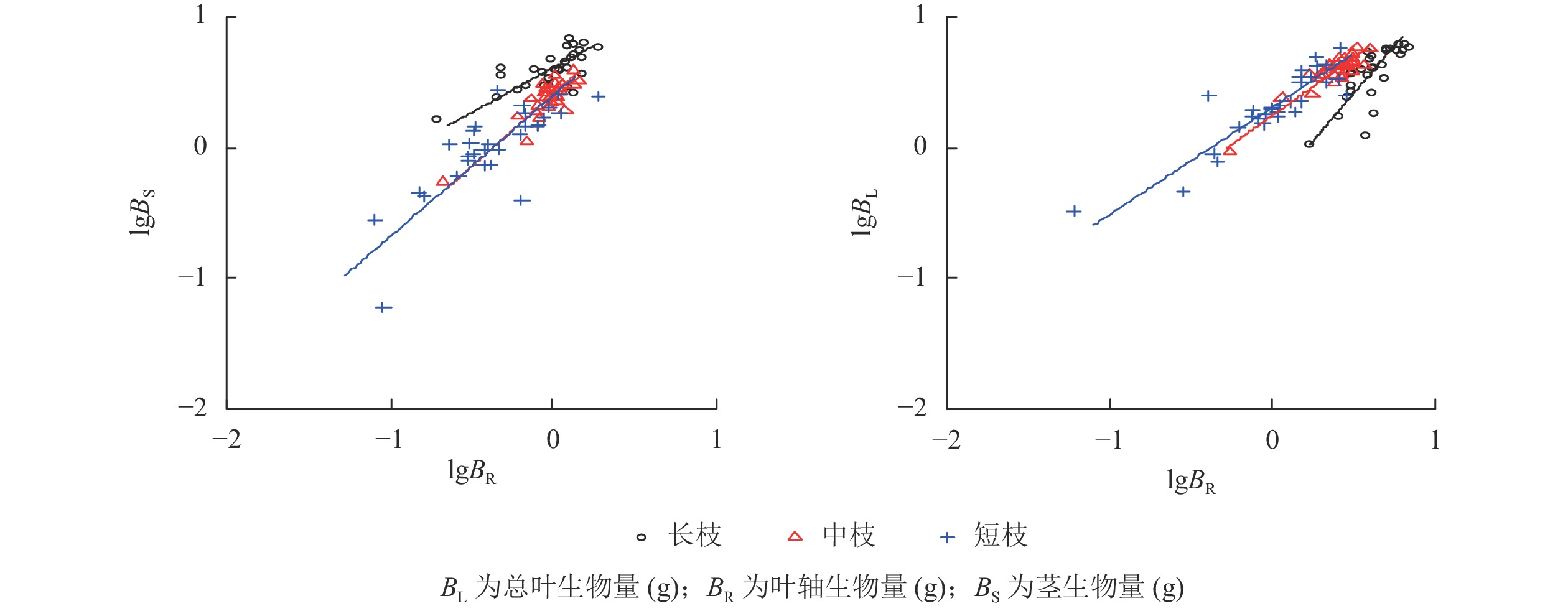
由表1和图1所示:3种类型枝条的茎生物量与总叶面积均存在显著相关性(P<0.05),长枝和中枝均为同速生长关系,斜率分别为1.170和1.135,经检验无显著差异(P>0.05),而在短枝上为异速生长关系,斜率为0.657,显著小于长枝和中枝(P<0.05)。但是,长枝和中枝与单个小叶面积无相关性(P>0.05),与复叶片数存在异速生长关系(P<0.05),而短枝则与单个小叶面积和复叶片数均存在显著的异速生长关系(P<0.05)。
表 1 小叶白蜡当年生枝条的茎与叶片回归方程参数及显著性检验
Table 1. Parameters and significance test of stem-leaf regression equations of F. sogdiana shoot
茎与叶片 变量 斜率 截距 R2 回归方程
P值异速生长
检验P值复叶片数-
茎生物量长枝 0.879 a 0.858 0.617 0.000 0.014 中枝 0.750 a 1.083 0.661 0.000 0.013 短枝 0.330 b 1.236 0.708 0.000 0.000 单个小叶面
积-茎生物量长枝 0.836 a 0.411 0.003 0.771 − 中枝 0.735 a 0.688 0.077 0.138 − 短枝 0.397 b 0.938 0.326 0.001 0.000 总叶面积-
茎生物量长枝 1.170 a 1.601 0.398 0.000 0.922 中枝 1.135 a 1.903 0.513 0.000 0.343 短枝 0.657 b 2.174 0.591 0.000 0.001 说明:斜率后不同字母表示同一指标在不同类型枝条间差异显著(P<0.05)。–表示回归方程不显著,不进行异速生长检验 
图 1 小叶白蜡当年生枝条的茎和叶片异速生长分析
Figure 1. Allometric growth analysis of stem and leaf of F. sogdiana shoot
-
表2和图2所示:3种类型枝条总叶面积与叶轴生物量均存在显著相关性(P<0.05),其中长枝和中枝为同速生长关系(P>0.05),而短枝为异速生长关系(P<0.05)。但是,长枝和中枝的叶轴生物量与单个小叶面积并不存在相关性,而与复叶片数存在异速生长关系(P<0.05),表明长枝和中枝叶轴主要通过复叶片数增加总叶面积。短枝的叶轴与复叶片数和单个小叶面积均存在异速生长关系(P<0.05),表明短枝叶轴主要通过单个小叶面积和复叶片数增加总叶面积。
表 2 小叶白蜡当年生枝条的叶轴与叶片回归方程参数及显著性检验
Table 2. Parameters and significance test of rachis-leaf regression equations of F. sogdiana shoot
叶轴与叶片 变量 斜率 截距 R2 回归方
程P值异速生长
检验P值复叶片数-
叶轴生物量长枝 0.601 a 1.403 0.436 0.000 0.000 中枝 0.829 a 1.395 0.389 0.000 0.002 短枝 0.353 b 1.368 0.468 0.000 0.000 单个小叶面积-
叶轴生物量长枝 0.571 ab 0.929 0.024 0.423 − 中枝 0.812 a 0.994 0.002 0.154 − 短枝 0.424 b 1.097 0.273 0.003 0.000 总叶面积-
叶轴生物量长枝 0.800 b 2.326 0.368 0.000 0.107 中枝 1.255 a 2.376 0.431 0.000 0.119 短枝 0.701 b 2.437 0.436 0.000 0.016 说明:斜率后不同字母表示同一指标在不同类型枝条间差异显著(P<0.05)。−表示回归方程不显著,不进行异速生长检验 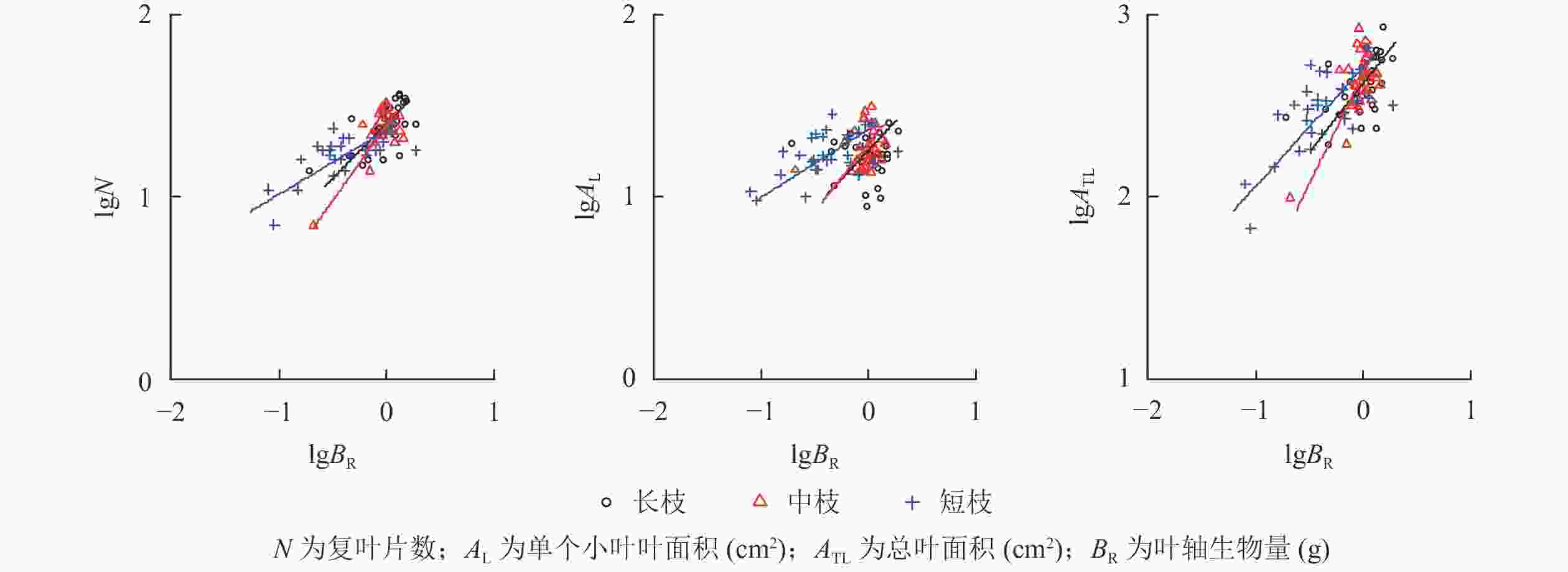
图 2 小叶白蜡当年生枝条的叶轴和叶片异速生长分析
Figure 2. Allometric growth analysis of rachis-leaf and leaf of F. sogdiana shoot
另外,在3种类型枝条之间叶的各项指标与叶轴生物量的斜率均以中枝最高,尤其总叶面积和叶轴的斜率达1.255,显著高于长枝和短枝(P<0.05),显示出最大的叶面积支持效率。
-
从表3可见:3种类型枝条的茎生物量和叶轴生物量之间均具有显著的相关性(P<0.05),其中,在长枝上表现为异速生长关系(P<0.05),斜率为0.683,在中枝和短枝上表现为同速生长关系(P>0.05),斜率分别为1.105和1.067,且均显著高于长枝(P<0.05)。
表 3 小叶白蜡当年生枝条生物量分配的回归方程参数及显著性检验
Table 3. Parameters and significance test of regression equations for biomass allocation of F. sogdiana shoot
生物量分配 变量 斜率 截距 R2 回归方程
P值异速生长
检验P值茎生物量-
叶轴生物量长枝 0.683 b 0.620 0.508 0.000 0.007 中枝 1.105 a 0.417 0.748 0.000 0.299 短枝 1.067 a 0.400 0.643 0.000 0.570 叶总生物量-
茎生物量长枝 1.459 a −0.303 0.512 0.000 0.008 中枝 0.908 b 0.243 0.817 0.000 0.242 短枝 0.840 b 0.327 0.799 0.000 0.048 说明:斜率后不同字母表示同一指标在不同类型枝条间差异显著(P<0.05) 在伊犁河谷小叶白蜡当年生枝条的生物量分配关系(图3)中发现:叶片总生物量和茎生物量之间的关系在不同类型枝条上不同。其中,在长枝上斜率为1.459,显著大于1 (P<0.05),表现为异速生长关系;在中枝上斜率为0.908 (P>0.05),表现为同速生长关系;在短枝上斜率为0.840,显著小于1 (P<0.05),表现为异速生长关系。
图 3 小叶白蜡当年生枝条的生物量分配关系
Figure 3. Biomass allocation of F. sogdiana shoots
-
植物种群在进化适应过程中总是要不断的调节构件生长,当年生枝条作为木本植物最为活跃的构件,茎和叶生长模式是其最重要的调节方式之一。但是在不同环境或不同生活型之间,茎和叶生长模式差异较大,并不存在一致性。以往在种间水平报道的茎和叶片生长关系既有异速模式[34−35],也有同速模式[36−37],这可能与不同的物种对环境梯度的进化反映差异有关。本研究在种内构件的水平上分析了不同类型枝条的生长特征,结果发现茎和叶的异速生长关系与枝条的类型有关。其中,在长枝和中枝上,茎生物量与总叶面积为同速生长关系,在短枝上,茎生物量与总叶面积为异速生长关系。因此,在解释生态过程时,有必要从不同的组织尺度上开展相关研究。
不同构件之间回归方程的斜率已经被广泛用来解释植物生长的权衡。本研究中,总叶面积与茎生物量的斜率在长枝和中枝上分别为1.170和1.135,在短枝上为0.657。说明随着茎生物量增加,长枝和中枝具有更高的叶面积支持效率(单位长度枝条的叶面积),并且主要与叶片数的增加有关,而与单个小叶叶面积无关,反之,短枝的总叶面积支持效率较低。这一规律在生物量分配中体现的尤为明显,长枝、中枝和短枝的叶片和茎生物量异速生长方程的斜率依次为1.459、0.908和0.840,叶片生物量分配比例依次降低。
综上,长枝的功能不仅体现在扩展种群生长空间上,同样体现在光合生产功能上,而短枝的功能在这2个方面相对较弱,这一点与以往的研究不同[26−27]。另外,曾凡江等[25]对新疆杨Populus alba var. pyramidalis长枝和短枝的叶水分生理特性研究表明:长枝叶片具有更高的蒸腾速率,光合速率与蒸腾速率往往存在显著的正相关关系,这与本研究小叶白蜡长枝具有较强光合功能的观点一致。
-
对于木本复叶植物,叶轴与茎共同构成枝条的支持系统,叶轴对于叶片生长也具有重要的调节作用,如努尔买买提等[31]研究表明:小叶白蜡的复叶长与叶轴长之间为异速生长关系,本研究的总叶面积和叶轴生物量之间同样存在异速生长关系。另外,将总叶面积和茎生物量的斜率与总叶面积和叶轴生物量的斜率进行对比发现:在长枝上前者为1.170,后者为0.800,表现出变小的趋势,而在中枝和短枝上,前者分别为1.135和0.657,后者分别为1.255和0.701,均表现出变大的趋势。表明长枝上茎具有更高的叶面积支持效率,而中枝和短枝上叶轴具有更高的叶面积支持效率。进一步说明叶轴对于中枝和短枝具有更为重要的作用,可以弥补茎对叶面积支持效率的不足,体现出叶轴的生态适应意义。
综上,在支撑结构上,长枝侧重于茎的生长,而叶轴的生长不足,从投资-收益权衡的角度可以理解为避免浪费过多的资源用于支持系统,而中枝和短枝则为茎和叶轴的生长并重,或者说需要增加叶轴的生长来弥补茎的生长不足,以提高对叶片的支持效率。
-
在长期进化适应过程中,小叶白蜡分化出复杂的枝条系统,长枝具有更长的茎和更多的叶片,似乎短枝无论在提高光合功能,还是拓展种群生长空间上,都不存在任何优势。而且,根据野外现场调查,在同一个一级分枝上,短枝与长枝的数量比例平均为15.1∶1.0,变异范围介于37.5∶1.0~1.5∶1.0,且分枝直径越大,短枝越多。由此产生的问题是短枝不存在任何生长优势,为什么还大量存在,对于这个问题需要结合小叶白蜡的结实生物学特征来理解。小叶白蜡当年生枝条不结实,而是2年生的枝条结实。据野外对结实植株的调查显示:如果当年生枝条为短枝,则去年生枝条结实量为12.15~12.94 g (鲜质量),当年生枝条叶片质量10.42~12.73 g (鲜质量);如果当年生枝条为长枝,则去年生枝条结实量为5.89~6.22 g (鲜质量),当年生枝条叶片质量16.28~25.92 g (鲜质量)。可见,小叶白蜡的枝条大小与结实之间存在权衡关系,短枝的大量发育是有性繁殖权衡的结果,这也符合植物种群生长和繁殖权衡的一般性规律。
-
小叶白蜡的长枝和中枝具有更高的叶片支持效率,其功能不仅体现在扩展种群生长空间方面,同样也体现在较高的光合生产能力方面,短枝的叶片支持效率较低,其功能与种群的有性繁殖过程密切相关。叶轴对叶片生长也有调节作用,在中枝和短枝上能够弥补茎对叶面积支持效率的不足,但在长枝上作用不明显。总之,小叶白蜡当年生枝条具有更为复杂的枝条功能分化,种群适应调节方式也更加多样化,研究具有二态性枝条的植物生长调节特征时需要考虑枝条类型的差异性。
Growth regulation characteristics of current-year shoots of Fraxinus sogdiana in Yili River Valley
-
摘要:
目的 当年生枝条是木本植物进行种群空间扩展、结实和光合作用的主要部位,反映种群进化和适应调节等重要生态学特征。研究当年生枝条的生长调节特征,可揭示植物种群的适应机制。 方法 通过分析国家Ⅱ级保护植物小叶白蜡Fraxinus sogdiana的长枝、中枝和短枝3种不同类型枝条各构件之间的异速生长关系,对不同类型枝条功能的差异性及其在种群适应调节中的作用进行研究。 结果 不同类型枝条的叶片和茎之间具有不同的生长规律,其中在总叶面积和茎生物量关系上,长枝和中枝上均为同速生长关系(斜率分别为1.170和1.135),而在短枝上为异速生长关系(斜率为0.657);在叶片生物量和茎生物量分配关系上,长枝、中枝和短枝分别表现为异速、同速、异速生长关系,回归方程的斜率依次为1.460、0.908和0.840。共同表明长枝和中枝具有更高的叶片生长能力,功能上侧重于扩展空间和光合生产,短枝的功能与有性繁殖过程有关。叶轴对叶片生长也有调节作用,在中枝和短枝上能够弥补茎对叶面积支持效率的不足,但在长枝上作用不明显。 结论 小叶白蜡的长枝和中枝具有更高的叶片支持效率,其功能不仅体现在扩展种群生长空间方面,同样也体现在较高的光合生产能力方面,当年生枝条具有更为复杂的功能分化,在种群适应调节中的作用更加多样化,建议在研究具有二态性枝条的植物生长调节特征时,应考虑枝条类型的差异性。图3表3参37 Abstract:Objective The current-year shoots, as the main parts of woody plants for population spatial expansion, fruiting and photosynthesis, are the most vigorous modulars and reflect the important ecological characteristics of population evolution and adaptive regulation. Therefore, this study, with an investigation into the growth regulation characteristics of current year branches, is aimed to reveal the adaptive mechanism of plant population. Method An analysis was conducted of the allometric growth relationships among the modulars of three types of shoots (long shoot, medium shoot and short shoot) of Fraxinus sogdiana, a plant of second-class protection in China, so as to explore the functional differences of different types of branches and their roles in population adaptation and regulation. Result Leaves and stems of different types of shoots are featured with different growth rules with allometric growth on long shoot and medium shoot (slope was 1.170 and 1.135 respectively), and isokinetic growth on short shoot (slope was 0.657) in terms of total leaf area and stem biomass. Isokinetic, allometric and isokinetic growth rules apply on long shoot, medium shoot and short shoot when it comes to the relationship between leaf biomass and stem biomass with the slope of regression equation being 1.460, 0.908 and 0.840 respectively. The long shoot and the medium shoot had higher leaf growth ability and mainly functioned in the expansion of space and photosynthetic production whereas the short shoot was more involved in the sexual reproduction process. Leaf rachis can also regulate leaf growth, making up for the lack of stem support efficiency for leaf area on medium shoot and short shoots, yet with no obvious effect on long shoots. Conclusion The long and medium shoots of F. sogdiana have higher leaf support efficiency, and their functions are not only reflected in expanding the population growth space, but also in higher photosynthetic production capacity. The current year shoots of F. sogdiana demonstrate more complex functional differentiation, with their roles in population adaptation regulation being more diverse, implying that shoot types should be considered in future studies dealing with the characteristics of plant growth regulation with dimorphic shoots. [Ch, 3 fig. 3 tab. 37 ref.] -
Key words:
- Fraxinus sogdiana /
- biomass allocation /
- population regulation /
- allometric growth /
- modular /
- Yili River Valley
-
图 1 小叶白蜡当年生枝条的茎和叶片异速生长分析
Figure 1 Allometric growth analysis of stem and leaf of F. sogdiana shoot
图 2 小叶白蜡当年生枝条的叶轴和叶片异速生长分析
Figure 2 Allometric growth analysis of rachis-leaf and leaf of F. sogdiana shoot
表 1 小叶白蜡当年生枝条的茎与叶片回归方程参数及显著性检验
Table 1. Parameters and significance test of stem-leaf regression equations of F. sogdiana shoot
茎与叶片 变量 斜率 截距 R2 回归方程
P值异速生长
检验P值复叶片数-
茎生物量长枝 0.879 a 0.858 0.617 0.000 0.014 中枝 0.750 a 1.083 0.661 0.000 0.013 短枝 0.330 b 1.236 0.708 0.000 0.000 单个小叶面
积-茎生物量长枝 0.836 a 0.411 0.003 0.771 − 中枝 0.735 a 0.688 0.077 0.138 − 短枝 0.397 b 0.938 0.326 0.001 0.000 总叶面积-
茎生物量长枝 1.170 a 1.601 0.398 0.000 0.922 中枝 1.135 a 1.903 0.513 0.000 0.343 短枝 0.657 b 2.174 0.591 0.000 0.001 说明:斜率后不同字母表示同一指标在不同类型枝条间差异显著(P<0.05)。–表示回归方程不显著,不进行异速生长检验  下载: 导出CSV
下载: 导出CSV
表 2 小叶白蜡当年生枝条的叶轴与叶片回归方程参数及显著性检验
Table 2. Parameters and significance test of rachis-leaf regression equations of F. sogdiana shoot
叶轴与叶片 变量 斜率 截距 R2 回归方
程P值异速生长
检验P值复叶片数-
叶轴生物量长枝 0.601 a 1.403 0.436 0.000 0.000 中枝 0.829 a 1.395 0.389 0.000 0.002 短枝 0.353 b 1.368 0.468 0.000 0.000 单个小叶面积-
叶轴生物量长枝 0.571 ab 0.929 0.024 0.423 − 中枝 0.812 a 0.994 0.002 0.154 − 短枝 0.424 b 1.097 0.273 0.003 0.000 总叶面积-
叶轴生物量长枝 0.800 b 2.326 0.368 0.000 0.107 中枝 1.255 a 2.376 0.431 0.000 0.119 短枝 0.701 b 2.437 0.436 0.000 0.016 说明:斜率后不同字母表示同一指标在不同类型枝条间差异显著(P<0.05)。−表示回归方程不显著,不进行异速生长检验
下载: 导出CSV
表 3 小叶白蜡当年生枝条生物量分配的回归方程参数及显著性检验
Table 3. Parameters and significance test of regression equations for biomass allocation of F. sogdiana shoot
生物量分配 变量 斜率 截距 R2 回归方程
P值异速生长
检验P值茎生物量-
叶轴生物量长枝 0.683 b 0.620 0.508 0.000 0.007 中枝 1.105 a 0.417 0.748 0.000 0.299 短枝 1.067 a 0.400 0.643 0.000 0.570 叶总生物量-
茎生物量长枝 1.459 a −0.303 0.512 0.000 0.008 中枝 0.908 b 0.243 0.817 0.000 0.242 短枝 0.840 b 0.327 0.799 0.000 0.048 说明:斜率后不同字母表示同一指标在不同类型枝条间差异显著(P<0.05)
下载: 导出CSV
-
[1] 宋海燕, 张静, 赵雅洁, 等. 不同程度石漠化对金山荚蒾末端小枝的生长和生物量积累及分配的影响[J]. 植物科学学报, 2018, 36(1): 103 − 111. SONG Haiyan, ZHANG Jing, ZHAO Yajie, et al. Effects of rocky desertification on growth and biomass accumulation and distribution of terminal twigs in Viburnum chishanense Graedn [J]. Plant Science Journal, 2018, 36(1): 103 − 111. [2] 彭希, 赵安玖, 陈智超, 等. 雅安周公山不同发育阶段峨眉含笑的枝叶性状[J]. 浙江农林大学学报, 2021, 38(1): 65 − 73. PENG Xi, ZHAO Anjiu, CHEN Zhichao, et al. Twig and leaf traits of Michelia wilsonii at different developmental stages in Zhougong Mountain, Ya’an [J]. Journal of Zhejiang A&F University, 2021, 38(1): 65 − 73. [3] 兰洁, 肖中琪, 李吉玫, 等. 天山雪岭云杉生物量分配格局及异速生长模型[J]. 浙江农林大学学报, 2020, 37(3): 416 − 423. LAN Jie, XIAO Zhongqi, LI Jimei, et al. Biomass allocation and allometric growth of Picea schrenkiana in Tianshan Mountains [J]. Journal of Zhejiang A&F University, 2020, 37(3): 416 − 423. [4] 耿梦娅, 陈芳清, 吕坤, 等. 长柄双花木叶性状异速生长关系随发育阶段和海拔梯度的变化[J]. 广西植物, 2019, 39(10): 1387 − 1397. GENG Mengya, CHEN Fangqing, LÜ Kun, et al. Changes of allometric relationships among leaf traits of Dissanthus cercidifolius var. longipes in different ontogenetic stages and altitude gradients [J]. Guihaia, 2019, 39(10): 1387 − 1397. [5] 孙俊, 王满堂, 程林, 等. 不同海拔典型竹种枝叶大小异速生长关系[J]. 应用生态学报, 2019, 30(1): 165 − 172. SUN Jun, WANG Mantang, CHENG Lin, et al. Allometry between twig size and leaf size of typical bamboo species along an altitudinal gradient [J]. Chinese Journal of Applied Ecology, 2019, 30(1): 165 − 172. [6] 龙嘉翼, 赵宇萌, 孔祥琦, 等. 观赏灌木枝条和叶性状在林下庇荫环境中的权衡关系[J]. 生态学报, 2018, 38(22): 8022 − 8030. LONG Jiayi, ZHAO Yumeng, KONG Xiangqi, et al. Trade-offs between twig and leaf traits of ornamental shrubs grown in shade [J]. Acta Ecologica Sinica, 2018, 38(22): 8022 − 8030. [7] 杨冬梅, 毛林灿, 彭国全. 常绿和落叶阔叶木本植物枝条内生物量分配关系研究: 异速生长分析[J]. 植物研究, 2011, 31(4): 472 − 477. YANG Dongmei, MAO Lincan, PENG Guoquan. Within-twig biomass allocation in evergreen and deciduous broad-leaved species: allometric scaling analyses [J]. Bulletin of Botanical Research, 2011, 31(4): 472 − 477. [8] LIU Zhiguo, CAI Yongli, LI Kai, et al. The leaf size-twig size spectrum in evergreen broadleaved forest of subtropical China [J]. African Journal of Biotechnology, 2010, 9(23): 3382 − 3387. [9] SUN Shucun, JIN Dongmei, SHI Peili. The leaf size-twig size spectrum of temperate woody species along an altitudinal gradient: an invariant allometric scaling relationship [J]. Annals of Botany, 2006, 97(1): 97 − 107. [10] WESTOBY M, WRIGHT I J. The leaf size-twig size spectrum and its relationship to other important spectra of variation among species [J]. Oecologia, 2003, 135(4): 621 − 628. [11] 郭庆学, 柴捷, 钱凤, 等. 不同木本植物功能型当年生枝条功能性状差异[J]. 生态学杂志, 2013, 32(6): 1465 − 1470. GUO Qingxue, CHAI Jie, QIAN Feng, et al. Leaf and stem traits of current-year twigs vary with different functional types of woody plant [J]. Chinese Journal of Ecology, 2013, 32(6): 1465 − 1470. [12] LI Guoyong, YANG Dongmei, SUN Shucun. Allometric relationships between lamina area, lamina mass and petiole mass of 93 temperate woody species vary with leaf habit, leaf form and altitude [J]. Functional Ecology, 2010, 22(4): 557 − 564. [13] SMITH D D, SPERRY J S, ADLER F R. Convergence in leaf size versus twig leaf area scaling: do plants optimize leaf area partitioning? [J]. Annals of Botany, 2016, 119(3): 447 − 456. [14] 宫浩, 郭伟, 王存国, 等. 帽儿山常见树种枝叶功能性状特征及相关性[J]. 北华大学学报(自然科学版), 2016, 17(4): 455 − 462. GONG Hao, GUO Wei, WANG Cunguo, et al. Function traits and correlation characteristics of common tree species in Maoer Mountain [J]. Journal of Beihua University (Natural Science), 2016, 17(4): 455 − 462. [15] 章建红, 史青茹, 许洺山, 等. 浙江天童木本植物Corner法则的检验: 个体密度的影响[J]. 植物生态学报, 2014, 38(7): 655 − 664. ZHANG Jianhong, SHI Qingru, XU Mingshan, et al. Testing of Corner’s rules across woody plants in Tiantong region, Zhejiang Province: effects of individual density [J]. Chinese Journal of Plant Ecology, 2014, 38(7): 655 − 664. [16] 史青茹, 许洺山, 赵延涛, 等. 浙江天童木本植物Corner法则的检验: 微地形的影响[J]. 植物生态学报, 2014, 38(7): 665 − 674. SHI Qingru, XU Mingshan, ZHAO Yantao, et al. Testing of Corner’s rules across woody plants in Tiantong region, Zhejiang Province: effects of micro-topography [J]. Chinese Journal of Plant Ecology, 2014, 38(7): 665 − 674. [17] 李亚男, 杨冬梅, 孙书存, 等. 杜鹃花属植物枝条大小对枝条生物量分配及叶面积支持效率的影响: 异速生长分析[J]. 植物生态学报, 2008, 32(5): 1175 − 1183. LI Ya’nan, YANG Dongmei, SUN Shucun, et al. Effects of twig size on biomass allocation within twigs and on lamina area supporting efficiency in Rhododendron: allometirc scaling analysis [J]. Chinese Journal of Plant Ecology, 2008, 32(5): 1175 − 1183. [18] 孙蒙柯, 程林, 王满堂, 等. 武夷山常绿阔叶林木本植物枝条生物量分配[J]. 生态学杂志, 2018, 37(6): 1815 − 1823. SUN Mengke, CHENG Lin, WANG Mantang, et al. Twig biomass allocation of woody species in evergreen broad-leaf forest’ Wuyi Mountain [J]. Chinese Journal of Ecology, 2018, 37(6): 1815 − 1823. [19] 许月, 杨晓东, 谢一鸣, 等. 浙江天童木本植物小枝的“大小-数量”权衡[J]. 植物生态学报, 2012, 36(12): 56 − 64. XU Yue, YANG Xiaodong, XIE Yiming, et al. Twig size-number trade-off among woody plants in Tiantong region, Zhejiang Province of China [J]. Chinese Journal of Plant Ecology, 2012, 36(12): 56 − 64. [20] DÖRKEN V M, STÜTZEL T. The adaptive value of shoot differentiation in deciduous trees and its evolutionary relevance [J]. Boletín De La Sociedad Argentina De Botánica, 2009, 4(3/4): 421 − 439. [21] 冯梅, 黄文娟, 李志军. 胡杨叶形变化与叶片养分间的关系[J]. 生态学杂志, 2014, 33(6): 1467 − 1473. FENG Mei, HUANG Wenjuan, LI Zhijun. Relationship of leaf shape change and leaf nutrient contents in Populus euphratica [J]. Chinese Journal of Ecology, 2014, 33(6): 1467 − 1473. [22] KITAOKA S, QU Laiye, WATANABE Y, et al. Heterophyllous shoots of Japanese larch trees: the seasonal and yearly variation in CO2 assimilation capacity of the canopy top with changing environment[J/OL]. Plants, 2020, 9(10): 1278[2022-02-15]. doi:10.3390/plants9101278. [23] ANTTI T, SARITA K S, JARKKO S, et al. Differences in growth and gas exchange between southern and northern provenances of silver birch (Betula pendula Roth) in northern Europe [J]. Tree Physiology, 2019, 40(2): 198 − 214. [24] 高照全, 程建军, 李静, 等. 白海棠不同类型枝条光合能力比较研究[J]. 中国农学通报, 2015, 31(25): 90 − 95. GAO Zhaoquan, CHENG Jianjun, LI Jing, et al. Study on photosynthetic ability of different shoot types of white crabapple [J]. Chinese Agricultural Science Bulletin, 2015, 31(25): 90 − 95. [25] 曾凡江, 李小明, 张希明. 塔克拉玛干沙漠南缘新疆杨长枝叶与短枝叶水分生理特性的初步研究[J]. 干旱区研究, 2002, 19(1): 44 − 48. ZENG Fanjiang, LI Xiaoming, ZHANG Ximing. A preliminary study on the moisture physiological characteristics of long and short shoot leaves of Populus alba var. pyramidalis in the south marginal zone of Taklamakan desert [J]. Arid Zone Research, 2002, 19(1): 44 − 48. [26] JARČUŠKA B, MILLA R. Shoot-level biomass allocation is affected by shoot type in Fagus sylvatica [J]. Journal of Plant Ecology, 2012, 5(4): 422 − 428. [27] 王开, 刘建军, 康博文, 等. 8种落叶树种的当年枝变异格局的研究[J]. 西北林学院学报, 2008, 23(4): 45 − 51. WANG Kai, LIU Jianjun, KANG Bowen, et al. Variation patterns in current shoot structure in eight deciduous tree species [J]. Journal of Northwest Forestry University, 2008, 23(4): 45 − 51. [28] SAOUAB F E, AMRAOUI M B. Short shoot growth and reproduction response to light conditions vary with order shooting in the proximal part of C. atlantica crown [J]. International Journal of Forestry Research, 2020, 2020(9): 1 − 9. [29] 何丙辉, 钟章成. 不同光强与干旱胁迫对银杏枝叶构件生长的影响[J]. 广西师范大学学报(自然科学版), 2005, 23(3): 66 − 69. HE Binghui, ZHONG Zhangcheng. Effects of environmental stress on modular growth of leaf and branch population in Ginkgo biloba [J]. Journal of Guangxi Normal University (Natural Science Edition), 2005, 23(3): 66 − 69. [30] 张维, 杨晓绒, 贾娜尔·阿汗, 等. 新疆伊犁河岸小叶白蜡复叶构件生物量分配[J]. 干旱区研究, 2016, 33(1): 114 − 119. ZHANG Wei, YANG Xiaorong, Jianaer Ahan, et al. Biomass allocation among components of compound leaves of Fraxinus sogdiana in Yili River Reaches’ Xinjiang [J]. Arid Zone Research, 2016, 33(1): 114 − 119. [31] 努尔买买提, 张维, 杨晓绒, 等. 新疆伊犁河岸小叶白蜡复叶构件的表型可塑性[J]. 东北林业大学学报, 2014, 42(11): 31 − 34, 61. Nuer Maimaiti, ZHANG Wei, YANG Xiaorong, et al. Phenotype plasticity on component size of compound leaves of Fraxinus sogdiana in Yili’ Xinjiang [J]. Journal of Northeast Forestry University, 2014, 42(11): 31 − 34, 61. [32] WARTON D I, WRIGHT I J, FALSTER D S, et al. Bivariate line-fitting methods for allometry [J]. Biological Reviews, 2006, 81(2): 259 − 291. [33] WARTON D I, DUURSMA R A, FALSTER D S, et al. Smatr 3-an R package for estimation and inference about allometric lines [J]. Methods in Ecology and Evolution, 2012, 3(2): 257 − 259. [34] 李曼, 郑媛, 郭英荣, 等. 武夷山不同海拔黄山松枝叶大小关系[J]. 应用生态学报, 2017, 28(2): 537 − 544. LI Man, ZHENG Yuan, GUO Yingrong, et al. Scaling relationships between twig size and leaf size of Pinus hwangshanensis along an altitudinal gradient in Wuyi Mountains, China [J]. Chinese Journal of Applied Ecology, 2017, 28(2): 537 − 544. [35] 李钰, 赵成章, 侯兆疆, 等. 高寒退化草地狼毒种群个体大小与茎、叶的异速生长[J]. 生态学杂志, 2013, 32(2): 241 − 246. LI Yu, ZHAO Chengzhang, HOU Zhaojiang, et al. Body size and stem- and leaf allometry of Stellera chanaejasme in degraded alpine grassland [J]. Chinese Journal of Ecology, 2013, 32(2): 241 − 246. [36] 商侃侃, 张希金, 宋坤. 上海辰山植物园不同生活型木本植物枝叶大小关系的比较[J]. 植物研究, 2020, 40(5): 641 − 647. SHANG Kankan, ZHANG Xijin, SONG Kun. Variation of stem-leaf size relationship of woody plants among different lifeforms in Shanghai Chenshan Botanical Garden [J]. Bulletin of Botanical Research, 2020, 40(5): 641 − 647. [37] 刘志国, 蔡永立, 李恺. 亚热带常绿阔叶林植物叶-小枝的异速生长[J]. 植物生态学报, 2008, 32(2): 363 − 369. LIU Zhiguo, CAI Yongli, LI Kai. Studies on the leaf size-twig size spectrum of subtropical evergreen broad-leaved woody species [J]. Journal of Plant Ecology, 2008, 32(2): 363 − 369. -

-
链接本文:
https://zlxb.zafu.edu.cn/article/doi/10.11833/j.issn.2095-0756.20220243
 点击查看大图
点击查看大图
计量
- 文章访问数: 1504
- HTML全文浏览量: 324
- PDF下载量: 53
- 被引次数: 0
