





-
兰科Orchidaceae植物是单子叶植物中的第一大科[1],兰花作为中国“十大名花”之一,誉有“花中四君子”的称号,其中许多种类可作为观赏植物或药用植物加以利用,经济价值较高。在兰花栽培中,常发生多种病害导致其生长及开花品质受到较大的影响,主要有胶胞炭疽菌Colletotrichum glososporioides引起的兰花炭疽病,尖孢镰刀菌Fusarium oxysporum引起的兰花茎腐病,灰葡萄孢菌Botrytis cinerea引起的兰花灰霉病等[2]。目前,兰花病虫害的防控主要采用化学农药防治,即对病害植物发病部位进行喷施、灌根或涂抹处理,但长期使用化学农药,不仅会造成环境污染,破坏生态平衡,还会导致植物病原菌对化学农药产生抗性,影响防治效果[3]。绿色生物防治是指利用拮抗微生物对病原菌进行防治,有防病抑病和保护环境的双重功效[4]。目前,从植物中分离有益内生真菌用作生物防治已成为一大热点。研究表明:内生真菌不仅能促进植物生长发育,还能提高宿主植物抗逆性[5−6]。兰科植物是典型的内生菌根植物,其根内分布着能促进植株生长发育的菌丝体,两者互利共生是兰科植物能在生态系统中存活并发挥作用的重要因素[7]。有研究已从建兰Cymbidium ensifolium、蕙兰C. faberi、寒兰C. kanran[8−9]、带叶兜兰Paphiopedilum hirsutissimum[10]等多种野生兰科植物中分离获得的内生真菌均对植株表现出促生效应;从野生青天葵Nervilia fordii[11]、野生春兰C. goeringii[12]、铁皮石斛Dendrobium officinale[4]中分离的内生真菌对叶斑病、根腐病、炭疽病等常见植物病害具有良好的拮抗作用。
蕙兰‘红香妃’C. faberi‘Hongxiangfei’原产于四川九寨沟高海拔地区,因其叶片基部呈紫红色且花开红色而得名,拥有典雅的花色和淡雅的花香,深受大众喜爱。但由于人为过度采挖,导致蕙兰‘红香妃’野生资源处于濒危灭绝状态。本研究对蕙兰‘红香妃’内生真菌进行分离鉴定,旨在筛选出能有效控制病害的生防真菌资源,为开发兰花生物肥及绿色生物防治研究提供科学依据。
-
蕙兰‘红香妃’取自四川省九寨沟县,春兰‘绿云’C. goeringii‘Green Cloud’来自浙江农林大学花卉栽培与遗传改良实验室。抑菌试验中的指示菌菌株为该实验室保存的兰花茎腐病病原菌(尖孢镰刀菌)、兰花炭疽病病原菌(胶孢炭疽菌)、番茄早疫病病原菌(链格孢菌Alternaria alternata)、草莓根腐病病原菌(三线镰刀菌F. tricinctum)、水稻恶苗病病原菌(藤仓镰孢菌F. fujikuroi)等5种病原菌。
-
马铃薯葡萄糖琼脂培养基(PDA)、马铃薯葡萄糖肉汤培养基(PDB)[6]。
-
采用组织分离法分离内生真菌[13]。取蕙兰‘红香妃’新鲜健康根段,先用洗洁精洗去根表面的污垢,并用流水冲洗30 min。将洗净后的根段置于超净工作台,于体积分数为75%乙醇溶液中浸泡30 s,用无菌水漂洗2次,体积分数为2%次氯酸钠溶液表面灭菌5 min,再用无菌水漂洗3~5次;将已表面灭菌的根切成0.5 cm的根段,切面朝向PDA培养基,在28 ℃下恒温培养,每天观察菌丝生长情况。为验证表面灭菌是否彻底,取适量最后一次漂洗组织的无菌水,涂布于PDA培养基上,同等条件培养,观察有无菌落产生。
-
对已分离的不同形态的菌落进行梯度稀释纯化[14]。取5个2 mL无菌离心管,依次加入1 000、900、900、900、900 μL无菌水;用无菌牙签刮取适量菌落于1号离心管中,充分摇匀后吸取100 μL至2号离心管中,摇匀后再吸取100 μL至3号离心管中,以此类推,将5管稀释成10、100、1 000、10 000、100 000倍等;每管取50 μL菌落悬浮液均匀涂在PDA培养基上,每个倍数重复3次,在28 ℃恒温培养箱中暗培养5~7 d后,取最边缘的菌落接到新的PDA培养基上培养,多次转接直至菌落呈单一,将纯化的真菌于4 ℃冰箱中保存备用。
-
使用5 mm打孔器在已纯化菌饼边缘打孔,将菌饼投入到事先制备好的PDB培养基中。PDB培养基使用500 mL锥形瓶盛装,装液量为200 mL,在180 r·min−1、28 ℃摇床中培养7 d,后用无菌水稀释至1×109 CFU·L−1以备用[15]。
-
菌落宏观形态观察:取已纯化的真菌接种于新的PDA培养基上,28 ℃恒温培养7 d后观察菌落颜色、质地、形状等特征。菌落微观形态观察:用无菌牙签挑取微量菌丝于载玻片上染色制成临时装片,于光学显微镜下,观察菌丝体、孢子的大小及形状等显微特征[16]。
-
采用CTAB法提取真菌DNA[17],采用通用引物ITS1 (5′-TCCGTAGGTGAACCTGCGG-3′)和ITS4 (5′-TCCTCCGCTTATTGATATGC-3′)进行PCR扩增真菌ITS序列。PCR反应体系(20 μL):ddH2O 7 μL,ITS1 1 μL,ITS4 1 μL,Easy Taq Mix酶10 μL,DNA模板1 μL。PCR反应程序:94 ℃预变性4 min,94 ℃变性30 s,55 ℃退火30 s,72 ℃延伸45 s,72 ℃延伸10 min,共35个循环。切胶回收目的片段,送擎科生物科技有限公司测序。
-
目的序列于美国国家生物技术信息中心(NCBI)数据库(
https://blast.ncbi.nlm.nih.gov/Blast.cgi )进行Nucleotide BLAST比对,选取并下载同源性较高的序列。先用Clustal X软件进行序列比对,再用MEGA 6.0软件中的邻接法(neighbor-joining method)构建系统进化树,其中Bootstrap检验重复次数为1 000次。 -
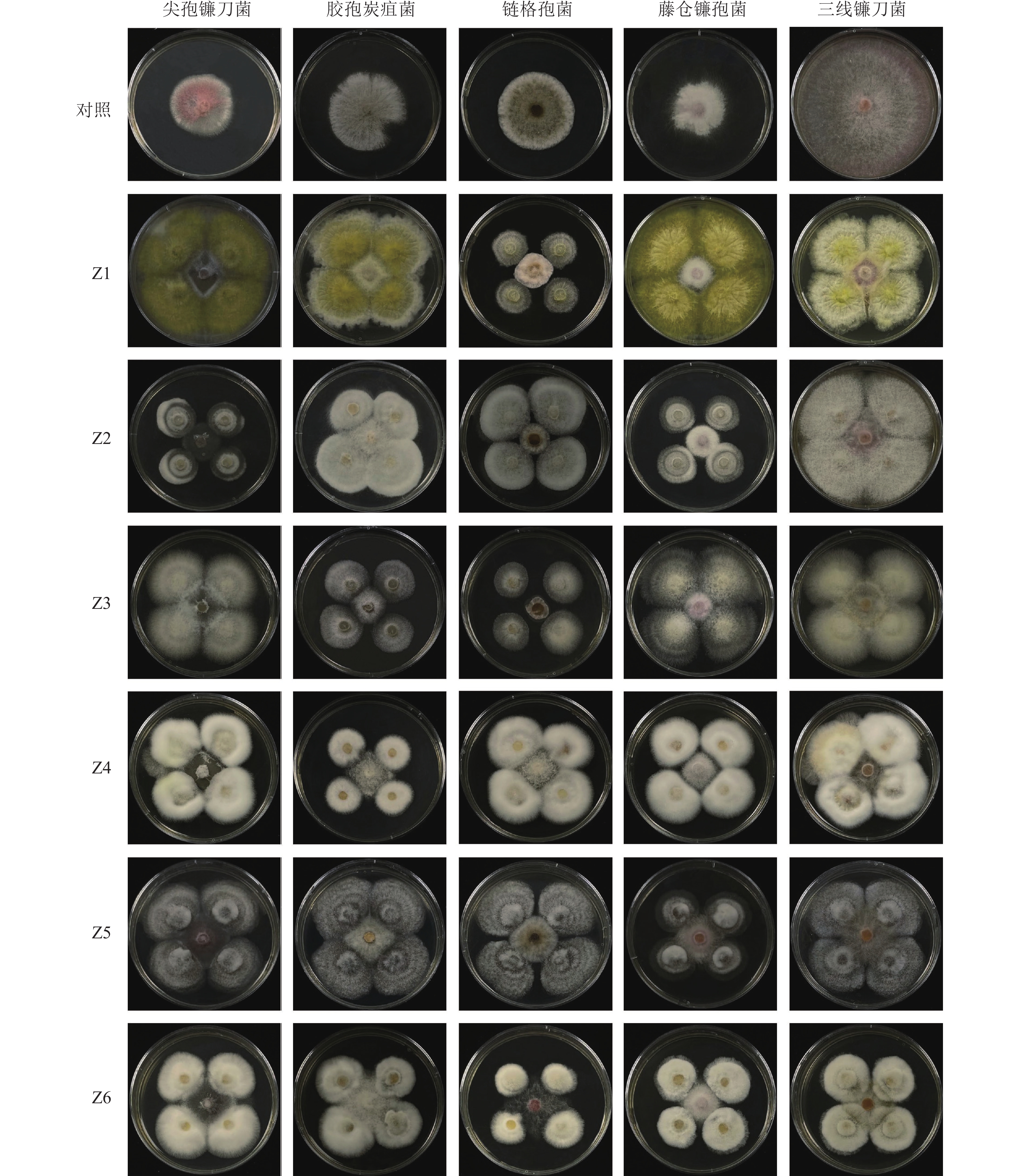
采用平板对峙法初步筛选出具有拮抗作用的内生真菌。在PDA平板中间位置放置直径为5 mm的指示菌菌饼,离指示菌菌饼上下左右各2 cm处放置相同大小的内生真菌,每个处理3次重复。以5 mm中间只接指示菌为对照,在28 ℃恒温培养箱中培养7 d后,用十字交叉法测量病原菌的菌落直径。计算内生真菌对病原菌的生长抑制率:抑制率=[(对照菌落直径−处理菌落直径)/对照菌落直径]×100%[18]。
-
选择胶孢炭疽菌初筛抑菌效果较好的内生真菌,针刺接种病菌于春兰‘绿云’进行活体试验[19]。设定2组:对照组为清水+病原菌(胶孢炭疽菌);处理组为内生真菌+病原菌(胶孢炭疽菌)。5株为1组,重复3组,用清水和内生真菌发酵液进行灌根和喷施处理30 d,再用病原菌胶孢炭疽菌发酵液进行灌根和喷施处理30 d。病情指数设定为6级,具体分级标准见表1。统计计算病叶率、病情指数、防治效果。病叶率= 病叶数/调查总叶数×100%;病情指数=∑(各级病株数×相对级数值)/(调查总株数×最高级值)×100%;防治效果=(对照病情指数−处理病情指数)/对照病情指数×100%。
表 1 病害程度分级标准
Table 1. Classification standard of disease
病情等级 代表值 病叶数量/片 0 0 0 1 1 1~3 2 3 4~5 3 5 6~7 4 7 8~9 5 9 ≥10 -
数据采用Excel 2010和SPSS 22.0软件的Duncan检验法对各处理组数据进行0.05或0.01水平下的差异显著性分析。
-
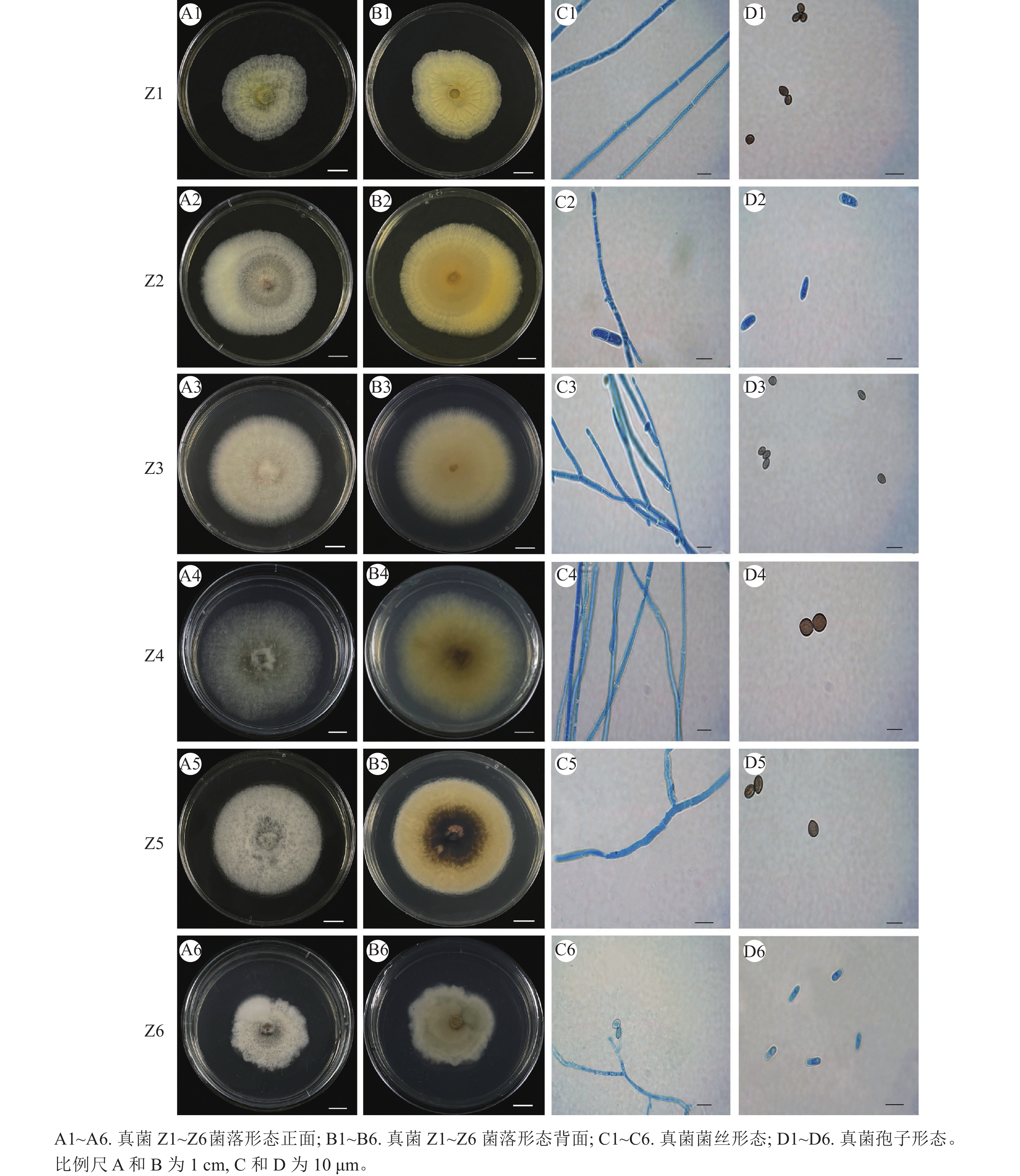
从来源于四川省九寨沟县的蕙兰‘红香妃’根段培养中共分离出13个真菌菌株,经纯化得到6个真菌菌株,其在PDA培养基上培养10 d后的菌落形态特征如图1所示,根据《真菌鉴定手册》[20]和《病原真菌鉴定》[21]进行形态学初步鉴定。
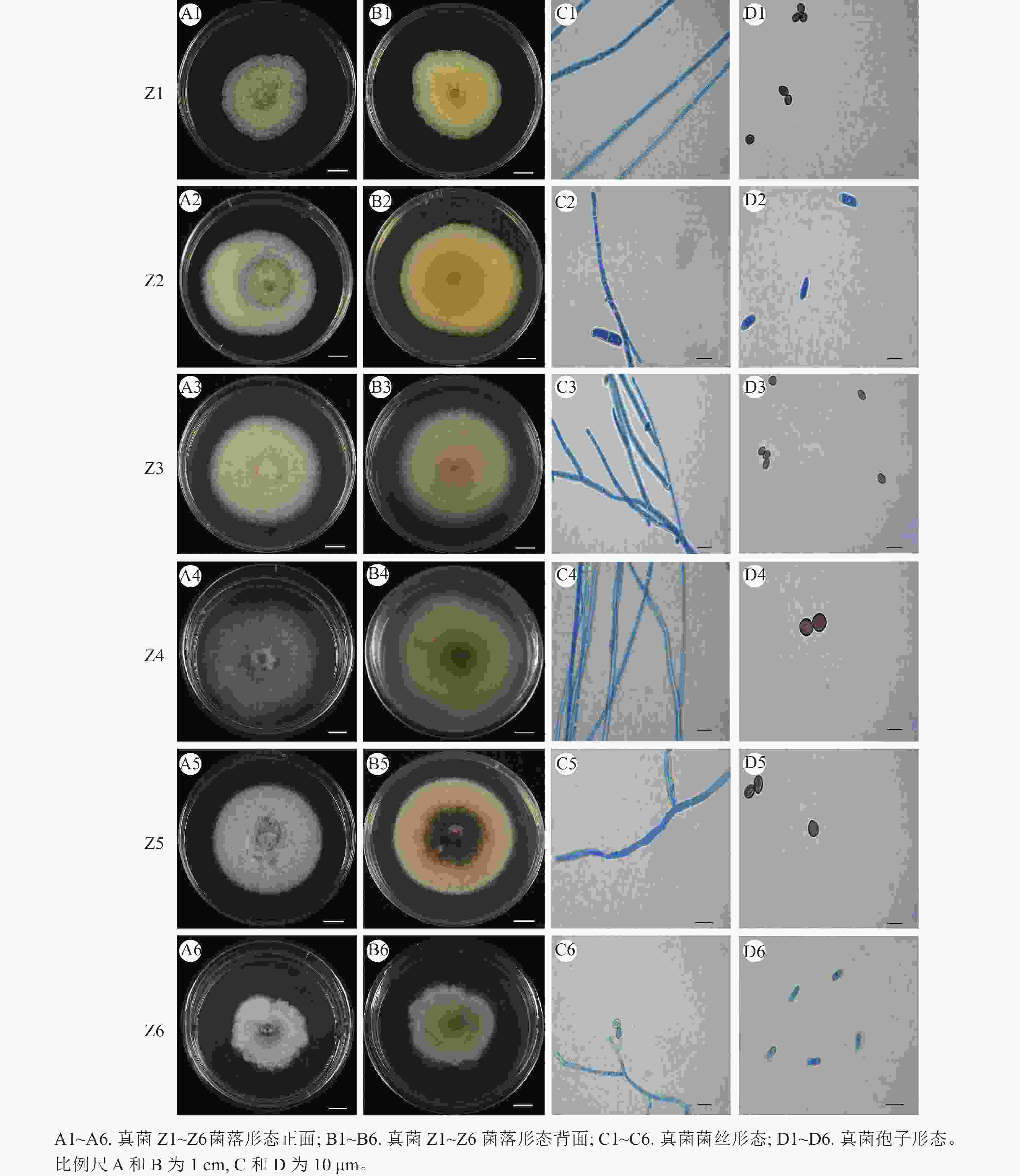
图 1 蕙兰‘红香妃’根部内生真菌菌株Z1~Z6的菌落形态特征
Figure 1. Colony morphological characteristics of endophytic fungi strains Z1-Z6 from the roots of C. faberi ‘Hongxiangfei’
菌株Z1:菌落大小为3.0~4.8 cm,菌落正面中间浅黄色,边缘白色,背面浅黄色,呈辐射沟纹,菌落质地绒毡状,边缘整齐。菌丝有隔膜,孢子柠檬形或近球形,两端突起,两侧平滑,大小为(3.9~8.4) μm×(5.0~6.5) μm。
菌株Z2:菌落大小为4.8~7.7 cm,正面白色,背面黄色,菌落质地棉絮状,一侧菌丝生长较多,边缘全缘。气生菌丝丰富,菌丝有隔膜,大型分生孢子镰刀形,多数为3个分隔,大小为(12.9~22.5) μm×(3.6~6.5) μm;小型分生孢子椭圆形或卵圆形,0~1个分隔,大小约为(4.8~12.7) μm×(2.5~5.8) μm。
菌株Z3:菌落大小为5.4~7.2 cm,正面白色,背面中间淡黄色边缘白色,菌落质地绒毡状,菌落形状圆形,边缘整齐。菌丝有隔,内含物颗粒多。孢子卵形或柠檬形,两端稍尖,大小为(4.2~7.5) μm×(4.0~6.5) μm。
菌株Z4:菌落大小为7.1~7.8 cm,正面灰色,背面中间深灰色,外圈由淡黄色至白色,菌落质地绒毡状,菌落形状接近圆形,边缘整齐。菌丝有隔,子囊具有8个子囊孢子,簇生,呈棍棒状。孢子呈褐色,卵圆形或柠檬形,两端突起,两侧平滑,大小为(8.2~11.2) μm×(5.1~8.6) μm。
菌株Z5:菌落大小为3.5~5.4 cm,正面烟灰色,背面中间深褐色,菌落质地绒毡状,菌落形状圆形,边缘整齐。菌丝有隔膜,孢子柠檬形,幼时无色,成熟后呈褐色或橄榄色,大小为(5.6~8.5) μm×(3.8~6.7) μm。
菌株Z6:菌落大小为3.2~4.6 cm,正面深灰色至烟灰色,背面淡黄色至浅灰色,菌落质地绒毛状,边缘不规则。菌丝有隔膜,子囊具有8个子囊孢子,呈椭圆形。孢子卵形,大小为(4.5~9.6) μm×(5.2~8.4) μm。
结合菌落与菌丝、孢子形态进行形态学初步鉴定可知:菌株Z1、Z3、Z4、Z5与毛壳属Chaetomium 特征相似,初步推断这4株菌株为毛壳属真菌;菌株Z2与镰刀属Fusarium 描述相似,推测菌株Z2为镰刀属真菌;菌株Z6初步判断为赛多孢属Scedosporium 真菌。需结合分子鉴定确定到种。
-
序列于NCBI数据库中进行BLAST比对,选取相似度最高序列,构建进化树(图2)。由系统发育树结果可知:菌株Z1与旋丝毛壳菌Collariella bostychodes (MK419025.1)亲缘关系最近,同源率为99%;菌株Z2与腐皮镰刀菌F. solani (FJ874633.1)亲缘关系最近,同源率为99%;菌株Z3与蝇生二叉毛壳菌D. funicola (KR909159.1)位于同一分支,同源率为100%;菌株Z4与球毛壳菌C. globosum (MT341778.1)位于同一分支,同源率为100%;菌株Z5与粪闭毛壳菌C. fimeti (MH001466.1)亲缘关系最近,同源率为100%;菌株Z6与波氏假阿利什霉Pseudallescheria boydii (AY213683.1)位于同一分支,同源率为100%。菌株Z1、Z3、Z4、Z5在系统发育树中聚为一个大分支,同属于毛壳属真菌。

图 2 基于菌株及其相似真菌的rDNA ITS序列构建的系统发育树
Figure 2. Phylogenetic tree based on rDNA ITS sequences of mycorrhizal fungi and similar fungi
-
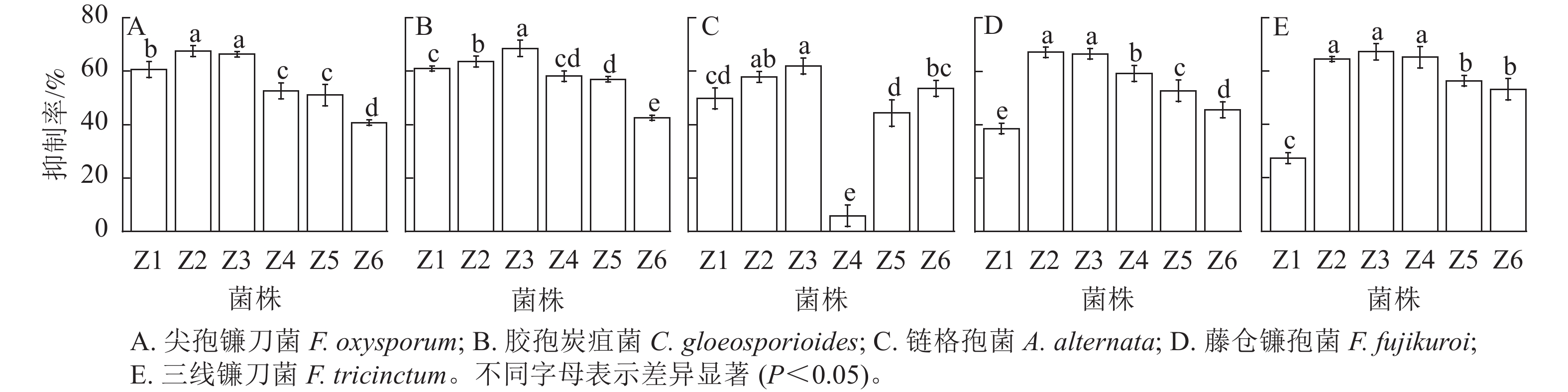
由图3和图4可知:6株内生真菌对5株病原菌的生长均具有一定的抑制作用,其中菌株Z2和Z3菌丝生长速度较快,对尖孢镰刀菌和藤仓镰孢菌的抑制效果显著优于其他菌株(P<0.05),而两者之间无显著差异;菌株Z3对胶孢炭疽菌的抑制效果较其他菌株存在显著性差异(P<0.05);菌株Z2和Z3对链格孢菌的抑制效果显著优于其他菌株(P<0.05);菌株Z2、Z3和Z4较其他菌株对三线镰刀菌的抑制效果显著(P<0.05),且三者之间无显著差异。综上,菌株Z2和Z3具有较大的生防潜力,可用于田间防控验证。
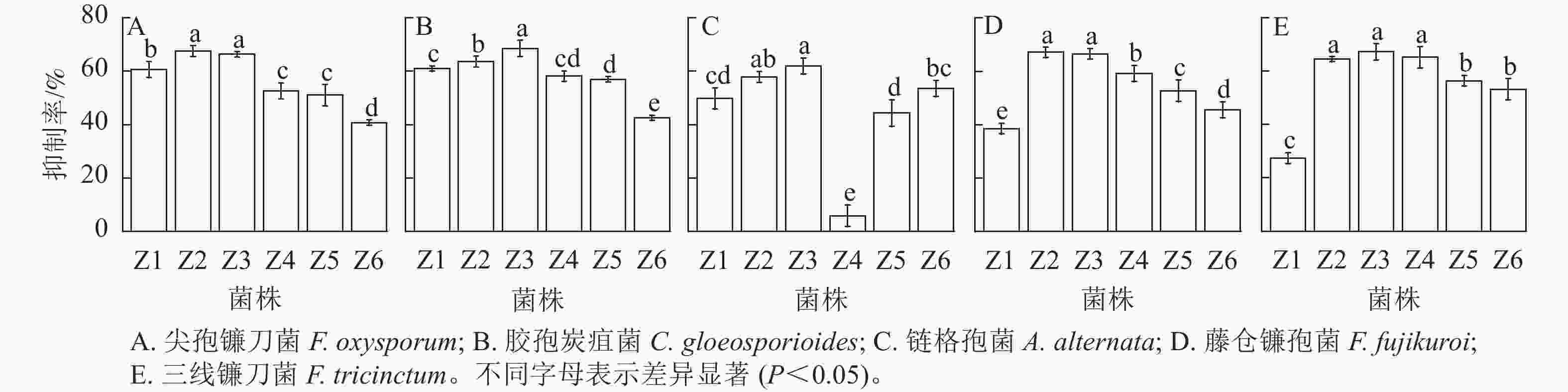
图 3 菌株Z1~Z6对5种主要病原真菌的拮抗抑制率
Figure 3. Antagonistic inhibition of Z1-Z6 against five pathogen fungi

图 4 菌株Z1~Z6对5种主要病原真菌的拮抗效果
Figure 4. Antagonistic effects of Z1-Z6 against five pathogen fungi
-
浇灌和喷施炭疽病菌后,春兰‘绿云’发病初期,叶片上呈现圆形、椭圆型红褐色小斑点,后期扩大成深褐色病斑。经重分离鉴定确认:该病害为胶孢炭疽菌所引起。由表2可知:P2组病斑明显少于对照组,且病叶率和病情指数极显著低于对照组(P<0.01),菌株Z3蝇生二叉毛壳菌对胶孢炭疽菌防治效果达41.41%,表明菌株Z3蝇生二叉毛壳菌对炭疽病菌具有一定的抑菌作用。
表 2 Z3菌株对春兰‘绿云’炭疽病的防治效果
Table 2. Control effect of Z3 strain on C. gloeosporioides of C. goeringii ‘Green Cloud’
处理 病叶率/% 病情指数 防治效果/% 对照组 26.43 17.53 ± 2.61 a 处理组 14.81 10.27 ± 2.17 b 41.41 说明:病情指数中“±”后为标准差;同列不同小写字母表示差异极显著(P<0.01)。 -
近年来,在园艺植物栽培中,生物防治作为一种环境友好型防治手段,越来越受到人们的关注。生防菌主要通过产生活性物质、重寄生或竞争营养来抑制病原微生物的生长及危害[2]。据研究报道:引起兰花茎腐病的病原菌为镰刀菌,镰刀属真菌多为土传类病原菌,主要侵染植物根茎部,导致根腐、茎腐、枯萎等[22]。镰刀属真菌虽被广泛认为是引起病害的主要致病菌[23−24],但会以致病菌或非致病菌的形式与兰花共生[25],而非致病性镰刀属真菌通常会以分解者或共生者的身份与兰花共生[26],有益于兰花的生长发育[27]。本研究从蕙兰‘红香妃’根部分离、纯化、鉴定出6株内生真菌,其中4株属于毛壳属,1株属于镰刀属,另1株属于赛多孢属;同时其与5株植物病原指示菌进行对峙培养发现:6株内生真菌对5株病原菌均有抑制效果,其中腐皮镰刀菌对尖孢镰刀菌抑制效果最佳,说明腐皮镰刀菌可能与蕙兰‘红香妃’建立共生关系,并有益于其生长和抗病性。李梅蓉等[4]发现:镰刀菌对铁皮石斛圆斑病具有良好的抑制作用。陈大为等[28]在对黄瓜Cucumis sativus白粉病的研究中也验证了腐皮镰刀菌可增强黄瓜植株抗性,抑制白粉病发病率。
炭疽病是园艺作物中寄主十分广泛的一种病害,引起兰花炭疽病的病原菌是炭疽菌属Colletotrichum spp.真菌,主要危害植物叶片,导致叶枯,其中胶孢炭疽菌是最为常见的病原菌[29−30]。本研究从蕙兰‘红香妃’中分离得到的蝇生二叉毛壳菌,对胶孢炭疽菌抑制效果显著。而毛壳属是极具生防潜力的真菌,在防治植物病害方面尤为突出[31−32]。PHONG等[33]探究了毛壳属真菌对茶树Camellia sinensis枯萎病的防治效果,发现毛壳属真菌均能显著抑制枯萎病菌菌丝的生长和产孢。SONG等[34]发现:毛壳属真菌对水稻Oryza sativa稻瘟病病菌具有拮抗作用。徐姣等[35]将5种人参Panax ginseng内生真菌与6种人参病原菌进行拮抗试验,发现毛壳属菌株对人参病害的抗菌谱较广,说明毛壳属真菌对植物不同病原菌均存在一定的拮抗作用,且同属不同种真菌发挥作用因寄主而异。为进一步验证抑菌效果,本研究选择菌株Z3蝇生二叉毛壳菌对春兰‘绿云’开展活体植株试验。结果表明:处理组病叶率、病情指数均极显著低于对照组,说明菌株Z3蝇生二叉毛壳菌具有较优的抑菌效果,有望开发为兰科植物生防菌,为兰科植物栽培及绿色生防提供科学依据。
-
本研究通过对四川省九寨沟县健康的蕙兰‘红香妃’根部内生真菌进行分离纯化、形态学鉴定及构建系统发育树等,确定内生真菌共6株。对峙试验明确6株内生真菌的抑菌谱较广,其中菌株Z2 (F. solani)和菌株Z3 (D. funicola)对5株病原菌均具有显著的抑制效果,具有生防潜力;通过菌株Z3对春兰‘绿云’活体植株进行浇灌喷施试验,证实菌株Z3具有一定的生防功效,但其抑菌相关机制有待进一步研究。
Isolation and identification of endophytic fungi from Cymbidium faberi ‘Hongxiangfei’ and their bacteriostatic effect in vitro
-
摘要:
目的 从蕙兰‘红香妃’Cymbidium faberi ‘Hongxiangfei’中筛选获得兰科Orchidaceae植物病害生防菌,为蕙兰菌根化栽培及绿色生物防治提供科学依据。 方法 通过蕙兰‘红香妃’健康根段培养、分离、纯化、鉴定内生真菌;以5株常见植物病原菌为指示菌,通过平板对峙培养法筛选出生物防治效果最优的内生真菌,并用活体接种法验证其抑菌效果。 结果 在蕙兰‘红香妃’健康植株根段培养中共分离筛选出6株内生真菌,经鉴定4株属于毛壳属Chaetomium spp.,1株属于镰刀属Fusarium spp.,另1株属于赛多孢属Scedosporium spp.。在平板对峙培养中发现:6株内生真菌对5株病原菌均具有一定的抑菌效果,其中Z2菌株腐皮镰刀菌Fusarium solani对兰花茎腐病致病菌——尖孢镰刀菌 F. oxysporum的抑制效果明显,抑制率为67.51%;Z3菌株蝇生二叉毛壳菌Dichotomopilus funicola对兰花炭疽病致病菌——胶孢炭疽菌Colletotrichum gloeosporioides的抑制率达68.56%。活体接种研究表明:经Z3菌株处理,春兰‘绿云’ C. goeringii ‘Green Cloud’的病叶率、病情指数极显著(P<0.01)低于对照。 结论 Z3菌株是筛选出的防治兰科植物病害的优势拮抗生物防治菌。图4表2参35 Abstract:Objective The objective is to screen and obtain the biocontrol fungi against orchid plant diseases from Cymbidium faberi ‘Hongxiangfei’ , so as to provide scientific basis for mycorrhizal cultivation and green biological control of C. faberi. Method The endophytic fungi were isolated, purified and identified from the healthy root segments of C. faberi ‘Hongxiangfei’ . With 5 common plant pathogens as indicator fungi, the endophytic fungi with the best biocontrol effect were screened by plate confrontation culture method, and their bacteriostatic effect was verified by in vivo inoculation. Result 6 endophytic fungi were isolated from the root segment culture of healthy plants of C. faberi ‘Hongxiangfei’ , and 4 strains belonged to Chaetomium spp., 1 strain belonged to Fusarium spp. and 1 belonged to Scedosporium spp. It was found in the plate confrontation culture that all 6 endophytic fungi had certain inhibitory effect on 5 pathogenic strains, among which Z2 strain (Fusarium solani) had an obvious inhibitory effect on F. oxysporum (the causative agent of orchid stem rot), with an inhibition rate of 67.51%. The inhibition rate of Z3 strain (Dichotomopilus funicola) against Colletotrichum gloeosporioides (the pathogen of orchid anthracnose) was 68.56%. The in vivo inoculation test of Cymbidium goeringii ‘Green Cloud’ showed that the rate of diseased leaves and disease index of the experimental group treated with Z3 strain were significantly lower than those of the control group (P<0.01). Conclusion Z3 is selected as the dominant antagonistic biocontrol strains against orchid diseases. [Ch, 4 fig. 2 tab. 35 ref.] -
Key words:
- Cymbidium faberi ‘Hongxiangfei’ /
- endophytic fungi /
- Chaetomium spp. /
- Fusarium spp. /
- bacteriostatic
-
图 1 蕙兰‘红香妃’根部内生真菌菌株Z1~Z6的菌落形态特征
Figure 1 Colony morphological characteristics of endophytic fungi strains Z1-Z6 from the roots of C. faberi ‘Hongxiangfei’
图 2 基于菌株及其相似真菌的rDNA ITS序列构建的系统发育树
Figure 2 Phylogenetic tree based on rDNA ITS sequences of mycorrhizal fungi and similar fungi
图 3 菌株Z1~Z6对5种主要病原真菌的拮抗抑制率
Figure 3 Antagonistic inhibition of Z1-Z6 against five pathogen fungi
图 4 菌株Z1~Z6对5种主要病原真菌的拮抗效果
Figure 4 Antagonistic effects of Z1-Z6 against five pathogen fungi
表 1 病害程度分级标准
Table 1. Classification standard of disease
病情等级 代表值 病叶数量/片 0 0 0 1 1 1~3 2 3 4~5 3 5 6~7 4 7 8~9 5 9 ≥10  下载: 导出CSV
下载: 导出CSV
表 2 Z3菌株对春兰‘绿云’炭疽病的防治效果
Table 2. Control effect of Z3 strain on C. gloeosporioides of C. goeringii ‘Green Cloud’
处理 病叶率/% 病情指数 防治效果/% 对照组 26.43 17.53 ± 2.61 a 处理组 14.81 10.27 ± 2.17 b 41.41 说明:病情指数中“±”后为标准差;同列不同小写字母表示差异极显著(P<0.01)。
下载: 导出CSV
-
[1] 金效华, 李剑武, 叶德平. 中国野生兰科植物原色图鉴: 上卷[M]. 郑州: 河南科学技术出版社, 2019: 10 − 20. JIN Xiaohua, LI Jianwu, YE Deping. Atlas of Native Orchids in China: Vol. 1 [M]. Zhengzhou: Henan Science and Technology Press, 2019: 10 − 20. [2] 吕昂, 王成, 汤亚芳, 等. 兰花茎腐病拮抗菌HJ-5的筛选和生防潜力评估[J]. 河南农业科学, 2021, 50(8): 85 − 93. LÜ Ang, WANG Cheng, TANG Yafang, et al. Isolation and evaluation of the antagonistic bacterium HJ-5 against orchid stem rot disease [J]. Journal of Henan Agricultural Sciences, 2021, 50(8): 85 − 93. [3] 李颖, 鞠昊伦, 宋子诺, 等. 作物疫病的绿色防控措施及其未来发展[J]. 江苏农业科学, 2022, 50(7): 18 − 27. LI Ying, JU Haolun, SONG Zinuo, et al. Environmental friendly prevention and control measures of crop Phytophthora blight and its development trend [J]. Jiangsu Agricultural Sciences, 2022, 50(7): 18 − 27. [4] 李梅蓉, 陈兴全, 郑亚强, 等. 铁皮石斛圆斑病病原菌鉴定及其生防菌筛选[J]. 植物医生, 2020, 33(2): 28 − 36. LI Meirong, CHEN Xingquan, ZHENG Yaqiang, et al. Identification of thepathogen for round apot on Dendrobium officinale and screening of the biocontrol fungus against it [J]. Plant Doctor, 2020, 33(2): 28 − 36. [5] 任爱梅, 张丽珂, 孟宪刚. 植物内生真菌研究进展与存在问题[J]. 广东农业科学, 2010, 37(2): 103 − 106. REN Aimei, ZHANG Like, MENG Xiangang. Research progress and existing problems of endophytic [J]. Guangdong Agricultural Sciences, 2010, 37(2): 103 − 106. [6] 王美娜, 胡玥, 李鹤娟, 等. 兰科植物菌根真菌研究新见解[J]. 广西植物, 2021, 41(4): 487 − 502. WANG Meina, HU Yue, LI Hejuan, et al. New insights into orchid mycorrhizal fungi research [J]. Guihaia, 2021, 41(4): 487 − 502. [7] 谢海伟, 冯嘉琪, 付晓晴, 等. 药用植物内生真菌的研究进展[J]. 江苏农业科学, 2020, 48(14): 1 − 6. XIE Haiwei, FENG Jiaqi, FU Xiaoqing, et al. Research progress of endophytic fungi in medicial plants [J]. Jiangsu Agricultural Sciences, 2020, 48(14): 1 − 6. [8] 张晋彦. 春兰‘宋梅’和杂交兰组培苗的菌根化研究[D]. 杭州: 浙江农林大学, 2017. ZHANG Jinyan. Studies on Mycorrhizal of Cymbidium goeringii ‘Songmei’ and Cymbidium hybridum[D]. Hangzhou: Zhejiang A&F University, 2017. [9] 杨前宇, 何聪芬, 梁立雄, 等. 菌根真菌对3种兰花幼苗生长作用研究[J]. 核农学报, 2019, 33(4): 687 − 695. YANG Qianyu, HE Congfen, LIANG Lixiong, et al. Effect of mycorrhizal fungi on plantlets growth of three species of orchid [J]. Journal of Nuclear Agricultural Sciences, 2019, 33(4): 687 − 695. [10] 陈宝玲, 杨开太, 龚建英, 等. 野生兜兰菌根真菌对带叶兜兰生长和生理指标的效应[J]. 热带亚热带植物学报, 2022, 30(1): 88 − 96. CHEN Baoling, YANG Kaitai, GONG Jianying, et al. Effects of wild Paphiopedilum mycorrhizal fungi on growth and physiological indexes of Paphiopedilum hirsutissimum seedlings[J]. Journal of Tropical and Subtropical Botany, 2022, 30(1): 88 − 96. [11] 杨鑫凤, 周雅琴, 谭小明, 等. 青天葵叶斑病病原菌的分离鉴定及其生物防治[J]. 北方园艺, 2021(24): 115 − 121. YANG Xinfeng, ZHOU Yaqin, TANG Xiaoming, et al. Isolation and identification of pathogen causing leaf spot disease in Nervilia fordii (Hance) Schltr. and its biocontrol [J]. Northern Horticulture, 2021(24): 115 − 121. [12] 夏伟, 霍雯雯, 张范琳, 等. 春兰根内生真菌的分离鉴定及生防菌筛选[J]. 生物资源, 2022, 44(2): 189 − 197. XIA Wei, HUO Wenwen, ZHANG Fanlin, et al. Isolation and identification of endophytic fungi from the root of Cymbidium goeringii and screening of biocontrol fungi [J]. Biotic Resources, 2022, 44(2): 189 − 197. [13] 闵长莉, 汪学军, 刘文博. 春兰内生真菌的分离及其抑菌活性的初步研究[J]. 西北植物学报, 2012, 32(3): 596 − 599. MIN Changli, WANG Xuejun, LIU Wenbo. Preliminary study on isolation of endophytic fungi from Cymbidium goeringii and its antimicrobial activity [J]. Acta Botanica Boreali-Occidentalia Sinica, 2012, 32(3): 596 − 599. [14] 莫维弟, 卯婷婷. 辣椒枯萎病土壤拮抗真菌的筛选与鉴定[J]. 贵州农业科学, 2019, 47(8): 66 − 70. MO Weidi, MAO Tingting. Screening and identification of antagonistic fungi in soil against Fusarium oxysporum in pepper [J]. Guizhou Agricultural Sciences, 2019, 47(8): 66 − 70. [15] 孙嘉莹, 刘美琦, 任伟超, 等. 人参曲霉属内生真菌发酵液抑菌效果研究[J]. 现代农业科技, 2021(18): 211 − 213, 216. SUN Jiaying, LIU Meiqi, REN Weichao, et al. Study on antibacterial effect of fermentation broth of Panax ginseng Aspergillus endophytic fungi [J]. Modern Agricultural Science and Technology, 2021(18): 211 − 213, 216. [16] 朱艳秋, 张荣意, 孟锐, 等. 黄花美冠兰根部内生真菌研究[J]. 西南农业学报, 2009, 22(3): 675 − 680. ZHU Yanqiu, ZHANG Rongyi, MENG Rui, et al. Endophytic fungi of roots of Eulophia flava [J]. Southwest China Journal of Agricultural Sciences, 2009, 22(3): 675 − 680. [17] 潘英文, 张凌, 王安石, 等. 濒危兰科植物 DNA 的提取方法[J]. 热带生物学报, 2019, 10(1): 94 − 98. PAN Yingwen, ZHANG Ling, WANG Anshi, et al. Extraction of genome DNA from endangered orchid plants [J]. Journal of Tropical Biology, 2019, 10(1): 94 − 98. [18] 袁洪波, 侯珲, 周增强, 等. 梨树腐烂病拮抗真菌JK2的分离和鉴定[J]. 果树学报, 2021, 38(10): 1760 − 1766. YUAN Hongbo, HOU Jun, ZHOU Zengqiang, et al. Isolation and identification of endophytic fungus JK2 antagonistic against pear Valsa canker caused by Valsa pyri [J]. Journal of Fruit Science, 2021, 38(10): 1760 − 1766. [19] 陈泉, 徐永红, 何锦辉, 等. 柑橘轮斑病抗性鉴定方法的建立[J]. 果树学报, 2022, 39(2): 295 − 301. CHEN Quan, XU Yonghong, HE Jinhui, et al. Establishment of an identification method of citrus resistance to target spot [J]. Journal of Fruit Science, 2022, 39(2): 295 − 301. [20] 魏景超. 真菌鉴定手册[M]. 上海: 上海科学技术出版社, 1979: 196. WEI Jingchao. Fungal Identification Manual[M]. Shanghai: Shanghai Scientific and Technical Publishers, 1979: 196. [21] CAMPBELL C K, JOHNSON E M, WARNOCK D W. 病原真菌鉴定[M]. 邹先彪, 周红, 译. 2版. 上海: 上海科学技术出版社, 2020: 174 − 187. CAMPBELL C K, JOHNSON E M, WARNOCK D W. Identification of Pathogenic Fungi[M]. ZOU Xianbiao, ZHOU Hong, tran. 2nd ed. Shanghai: Shanghai Scientific and Technical Publishers, 2020: 174 − 187. [22] 何洁, 梁霜, 李忠, 等. 过路黄叶斑病病原鉴定及拮抗细菌的筛选[J]. 核农学报, 2022, 36(2): 282 − 290. HE Jie, LIANG Shuang, LI Zhong, et al. Pathogen identification of Lysimachia christinae leaf sopt and screening of antagonistic bacterial strains [J]. Journal of Nuclear Agricultural Sciences, 2022, 36(2): 282 − 290. [23] 唐婕, 陈健鑫, 尼玛此姆, 等. 栽培墨兰根腐病病原鉴定与生防菌筛选[J]. 中国农学通报, 2021, 37(19): 128 − 133. TANG Jie, CHEN Jianxin, NIMA Cimu, et al. Pathogen identification and biocontrol screening of root rot of Cymbidium sinense [J]. Chinese Agricultural Science Bulletin, 2021, 37(19): 128 − 133. [24] 李艳梅, 李小六, 陈超, 等. 大花蕙兰根腐病病原菌的分离与鉴定[J]. 河南农业大学学报, 2007, 56(1): 85 − 89. LI Yanmei, LI Xiaoliu, CHEN Chao, et al. Isolation and identification of the pathogens causing root rot disease of Cymbidium hybrida [J]. Journal of Henan Agricultural University, 2007, 56(1): 85 − 89. [25] SRIVASTAVA S, KADOOKA C, UCHIDA J Y. Fusarium species as pathogen on orchids [J]. Microbiological Research, 2018, 207: 188 − 195. [26] BOOTH C. The Genus Fusarium[R]. Kew: Commonwealth Mycological Institute, 1971. [27] ALABOUVETTE C, LEMANCEAU P, STEINBERY C. Recent advances in the biological control of Fusarium wilts [J]. Pesticide Science, 1993, 37(4): 365 − 373. [28] 陈大为, 卢幸蓉, 张树武, 等. 生防腐皮镰刀菌JK-13对黄瓜抗白粉病的诱导作用[J]. 甘肃农业大学学报, 2018, 53(4): 82 − 86, 91. CHEN Dawei, LU Xingrong, ZHANG Shuwu, et al. Inductive effect of bio-control Fusarium solani JK-13 on cucumber resistance against powdery mildew [J]. Journal of Gansu Agricultural University, 2018, 53(4): 82 − 86, 91. [29] 张静雅, 李欣雨, 张成, 等. 木薯炭疽病拮抗木霉菌筛选与室内防效研究[J]. 中国生物防治学报, 2022, 38(1): 115 − 124. ZHANG Jingya, LI Xinyu, ZHANG Cheng, et al. Screening of antagonistic Trichoderma against cassava anthracnose and investigation on its control effect in laboratory [J]. Chinese Journal of Biological Control, 2022, 38(1): 115 − 124. [30] 徐波, 宋凤鸣, 冯淑杰, 等. 兜兰炭疽病病原鉴定及其室内防治药剂筛选[J]. 南方农业学报, 2018, 49(2): 271 − 279. XU Bo, SONG Fengming, FENG Shujie, et al. Identification for anthracnose pathogen of Paphiopedilum and selection of its indoor chemical control chemicals [J]. Journal of Southern Agriculture, 2018, 49(2): 271 − 279. [31] HUNG P M, WATTANACHAI P, KASEM S, et al. Biological control of Phytophthora palmivora causing root rot of pomelo using Chaetomium spp. [J]. Mycobiology, 2015, 43(1): 63 − 70. [32] ZHAO Shuangshuang, ZHANG Yingying, YAN Wei, et al. Chaetomium globosum CDW7, a potential biological control strain and its antifungal metabolites [J]. FEMS Microbiology Letters, 2017, 364(3): 1 − 6. [33] PHONG N H, PONGNAK W, SOYTONG K. Antifungal activities of Chaetomium spp. against Fusarium wilt of tea [J]. Plant Protection Science, 2016, 52(1): 10 − 17. [34] SONG J, SOYTONG K, KANOKMEDHAKUL S, et al. Antifungal activity of microbial nanoparticles derived from Chaetomium spp. against Magnaporthe oryzae causing rice blast [J]. Plant Protection Science, 2020, 56(3): 180 − 190. [35] 徐姣, 常越, 刘振鹏, 等. 5种人参内生真菌对6种人参病害拮抗作用的研究[J]. 中医药学报, 2016, 44(6): 16 − 18. XU Jiao, CHANG Yue, LIU Zhenpeng, et al. Five kinds of endophytic fungi in Ginseng against six kinds of ginseng disease [J]. Acta Chinese Medicine and Pharmacology, 2016, 44(6): 16 − 18. -

-
链接本文:
https://zlxb.zafu.edu.cn/article/doi/10.11833/j.issn.2095-0756.20220578
 点击查看大图
点击查看大图
计量
- 文章访问数: 2066
- HTML全文浏览量: 355
- PDF下载量: 43
- 被引次数: 0
