




-
随着全球工农业活动强度的持续增加,重金属污染已在多种土壤生态系统中广泛存在,成为重要的土壤污染问题。砷(As)、镉(Cd)在稻田土壤中的长期累积,不仅威胁农田生态系统的结构与功能安全,还可能通过水稻Oryza sativa吸收并在食物链中富集,对粮食安全和人类健康构成潜在风险[1]。研究表明,土壤中过量的砷、镉会破坏土壤理化性质,抑制有益微生物的生长与代谢活动,从而干扰土壤养分循环过程和生态功能的正常发挥[2]。
稻田土壤具有典型的水分动态特征,淹水与排水过程会引起土壤氧化还原电位(Eh)和pH的显著变化,从而深刻影响砷、镉的迁移转化行为。稻田淹水条件下,铁锰(氢)氧化物的还原性溶解以及As(Ⅴ)向As(Ⅲ)的还原过程,通常会增强砷的迁移性和生物有效性[3]。排水条件下,氧化还原电位升高、pH降低,有利于As(Ⅲ)氧化为迁移性较弱的As(Ⅴ)[4],但同时可能导致结合镉的硫化物和铁氧化物发生不稳定转化,进而增加镉的迁移风险[5]。因此,仅通过水分管理难以实现稻田土壤中砷镉的同步钝化,亟需一种绿色、廉价的技术或材料实现对砷-镉复合污染稻田土壤的高效修复。
生物炭能够通过离子交换、化学络合及共沉淀作用降低土壤重金属的生物有效性[6],还能通过调节土壤理化性质及重塑微生物群落结构,间接影响土壤中砷、镉的生物有效性[7]。由于原始生物炭表面官能团的物理化学特性的限制,带负电荷且对阴离子砷的钝化效果较差,对砷-镉复合污染土壤的修复能力有限,难以满足实际农田修复需求,因此通过物理或化学改性制备高效功能型生物炭逐渐成为研究热点[8]。JIAO等[9]研究表明:铁镁改性生物炭可显著降低稻田土壤中高生物有效性砷的形态比例,其表面的铁镁复合氧化物能与砷形成内球络合物,从而促进可迁移态砷向稳定形态转化。
土壤微生物是维持土壤生态功能和调控重金属生物地球化学循环的关键,其群落结构受水分管理以及有机肥和化肥施用等因素的影响[10]。微生物群落结构和功能的变化可通过调节氧化还原反应、元素循环及沉淀作用进一步影响重金属的生物有效性[11]。在XIAO等[12]的研究中,负载氧化镁(MgO)的生物炭可通过固定土壤重金属,降低甲基杆菌属Methylobacillus、贪铜菌属Cupriavidus等重金属耐受类群丰度,优化优势类群组成并增强土壤生态功能;ISLAM等[13]研究发现:铁镁改性的玉米Zea mays秸秆生物炭和蛋壳炭能够提高污染稻田土壤中厌氧黏细菌属Anaeromyxobacter等参与重金属解毒或抗性细菌属的丰度,同时降低砷镉的生物有效性。目前,关于铁镁改性生物炭对稻田土壤中砷镉的固定效果以及微生物调控机制的研究多集中于单一水分条件下,在不同氧化还原(水分管理)条件下的修复效果及作用机制不明,且相关研究相对较少。因此,探索在不同水分条件下,添加铁镁改性生物炭后对污染土壤的修复效应及土壤微生物的作用与响应机制,具有重要的理论意义和实际应用价值。
本研究以海口市琼山区典型砷-镉复合污染稻田土壤为研究对象,以海南省热带作物衍生的椰壳生物炭为原料,采用铁镁双金属负载改性方法制备新型功能型钝化剂,通过室内土壤培养试验,系统比较其在淹水(厌氧)与湿润(好氧) 2种水分管理条件下对土壤砷镉生物有效性、理化性质以及微生物群落结构与功能的影响特征,旨在阐明水分调控与铁镁改性生物炭协同作用下砷、镉迁移转化及微生物调控机制,为热带稻田重金属污染土壤的精准修复与资源化治理提供科学依据。
-
供试土壤于2024年8月采自海南省海口市琼山区某收获后的典型稻田。去除土壤中可见的动植物残体后充分混匀,并过2 mm筛备用。土壤中砷和镉的背景质量分数分别为197.1和0.86 mg·kg−1,pH为6.1,属于酸性土壤。土壤中砷和镉质量分数超过GB
15618 —2018《土壤环境质量 农用地土壤污染风险管控标准(试行)》规定的风险筛选值。本研究所用生物炭为购自泰禹生物工程有限公司的椰壳生物炭(CBC, 650 ℃热解)。铁镁改性椰壳生物炭(FCBC)的制备过程如下:将20 g 椰壳生物炭均匀分散于300 mL含0.15 mol·L−1氯化镁(MgCl2)和0.15 mol·L−1硝酸铁[Fe(NO3)3]的混合溶液中,磁力搅拌下混合1 h。随后通入氢氧化钠(NaOH)溶液,将体系pH调至10.5,并在95 ℃条件下持续搅拌反应1 h,之后转移至90 ℃水浴中恒温反应6 h。反应结束后,用去离子水反复洗涤并用滤纸过滤,过滤后的样品置于60 ℃烘箱中干燥至恒量。
-
2025年8—9月,将生物炭与供试土壤按2%质量比充分混合后置于敞口烧杯中进行培养试验。试验设置淹水和湿润2种水分管理方式:淹水处理通过加入去离子水,使水层高度保持在土壤表面以上1~2 cm;湿润处理通过定期补水,使土壤含水量维持在田间持水量的60%~70%。试验共设置6种处理:①YS-ck,不施生物炭的淹水处理;②SR-ck,不施生物炭的湿润处理;③YS-CBC,施入2% CBC的淹水处理;④SR-CBC,施入2% CBC的湿润处理;⑤YS-FCBC,施入2% FCBC的淹水处理;⑥SR-FCBC,施入2% FCBC的湿润处理。每种处理设置4个重复,所有培养试验均在恒温(24±1) ℃条件下进行,培养周期为30 d。
-
通过比表面积分析仪(BET)、傅里叶红外光谱仪(FTIR)、X射线衍射分析(XRD)和扫描电子显微镜(SEM)对生物炭表面官能团、比表面积及形貌特征进行分析。稻田土壤的砷、镉背景值采用盐酸-硝酸-氢氟酸消解法测定。采用电位法对土壤的氧化还原电位(Eh)进行原位测定。按土液质量比1∶10将鲜土与去离子水混合,所得土壤浸提液用于测定溶解性有机碳(DOC)和水溶态硫酸盐($\mathrm{SO}_4^{2-} $)质量分数。采用0.5 mol·L−1盐酸提取-邻菲啰啉比色法测定土壤无定形Fe(Ⅱ)。
其余理化指标在自然条件下风干并过2 mm尼龙筛后的土样中测定。采用1 mol·L−1磷酸二氢钾(KH2PO4)和0.01 mol·L−1氯化钙(CaCl2)浸提土壤中有效态砷和有效态镉[14],经电感耦合等离子体质谱仪(ICP-MS)测定其质量分数。土壤有机碳(SOC)采用重铬酸钾-外加热法测定;土壤有效磷(AP)采用碳酸氢钠浸提-钼锑抗比色法测定;土壤pH采用电位法测定;土壤速效钾(AK)采用醋酸铵浸提-火焰光度法测定。
-
培养30 d后采集各处理土壤样品,用于微生物群落结构分析。采用E. Z. N. A. ® soil DNA kit (Omega Bio-tek)按照说明书提取土壤总DNA。利用NanoDrop 2000分光光度计(Thermo)测定DNA的浓度和纯度。经纯化和质量控制合格的DNA样品用于后续扩增与测序分析。
以细菌16S rRNA基因V4~V5高变区为扩增目标,选用引物对338F (5′-ACTCCTACGGGAGGCAGCAG-3′)和806R (5′-GGACTACHVGGGTWTCTAAT-3′)进行聚合酶链式反应(PCR)扩增。纯化后的PCR产物采用NEXTFLEX Rapid DNA-Seq Kit构建测序文库,文库质量合格后在Illumina MiSeq平台(上海美吉生物有限公司)进行高通量测序分析。
-
数据处理与统计分析通过Excel 2010、Origin 2024和SPSS 27完成。采用单因素方差分析(one-way ANOVA)检验不同处理间的差异性,并结合Duncan多重比较法进行显著性分析,显著性水平设定为P<0.05。采用mothur软件对α多样性(Ace指数、Chao1指数、Shannon-Wiener指数和Simpson指数)进行计算分析[15],用于评估土壤微生物群落的丰富度和多样性。采用R统计软件中的“vegan”包进行Mantel分析。基于Spearman相关系数(|r|>0.5,P<0.05)构建土壤细菌共现网络,网络的可视化与拓扑结构分析采用Gephi软件完成。
-
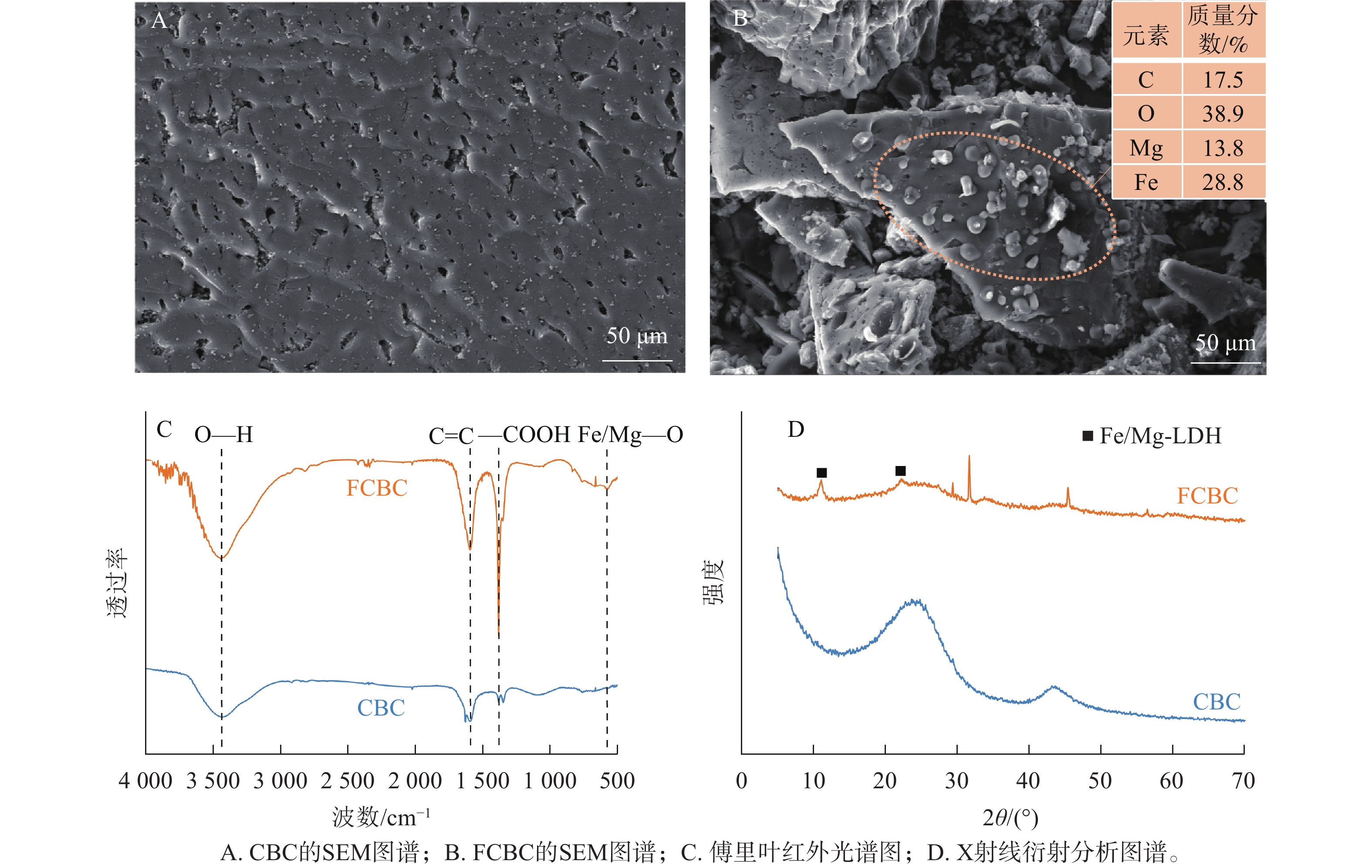
扫描电子显微镜图像(图1A、B)显示:CBC表面相对光滑,孔隙分布不均;而FCBC的形貌更为复杂。铁镁改性显著改变了生物炭表面结构,在炭基体与孔隙表面均形成大量粗糙的负载颗粒,能谱分析证实其成分包含铁和镁。傅里叶红外光谱仪分析结果(图1C)显示:FCBC在576 cm−1处出现吸收峰Fe/Mg—O[16]。X射线衍射分析图谱(图1D)显示:在11.03°和22.15°处的特征峰分别对应典型铁镁层状双金属氢氧化物(Fe/Mg-LDH)的(003)和(006)晶面。表1显示:相较于CBC,FCBC的碳元素质量分数降低、氧元素质量分数升高。这可能与负载大量铁镁羟基化合物,引入了更多含氧官能团有关。同时,FCBC比表面积和孔体积的减少进一步说明铁镁负载占据了生物炭表面与孔隙结构。可见,铁镁羟基化合物有效负载在生物炭表面。
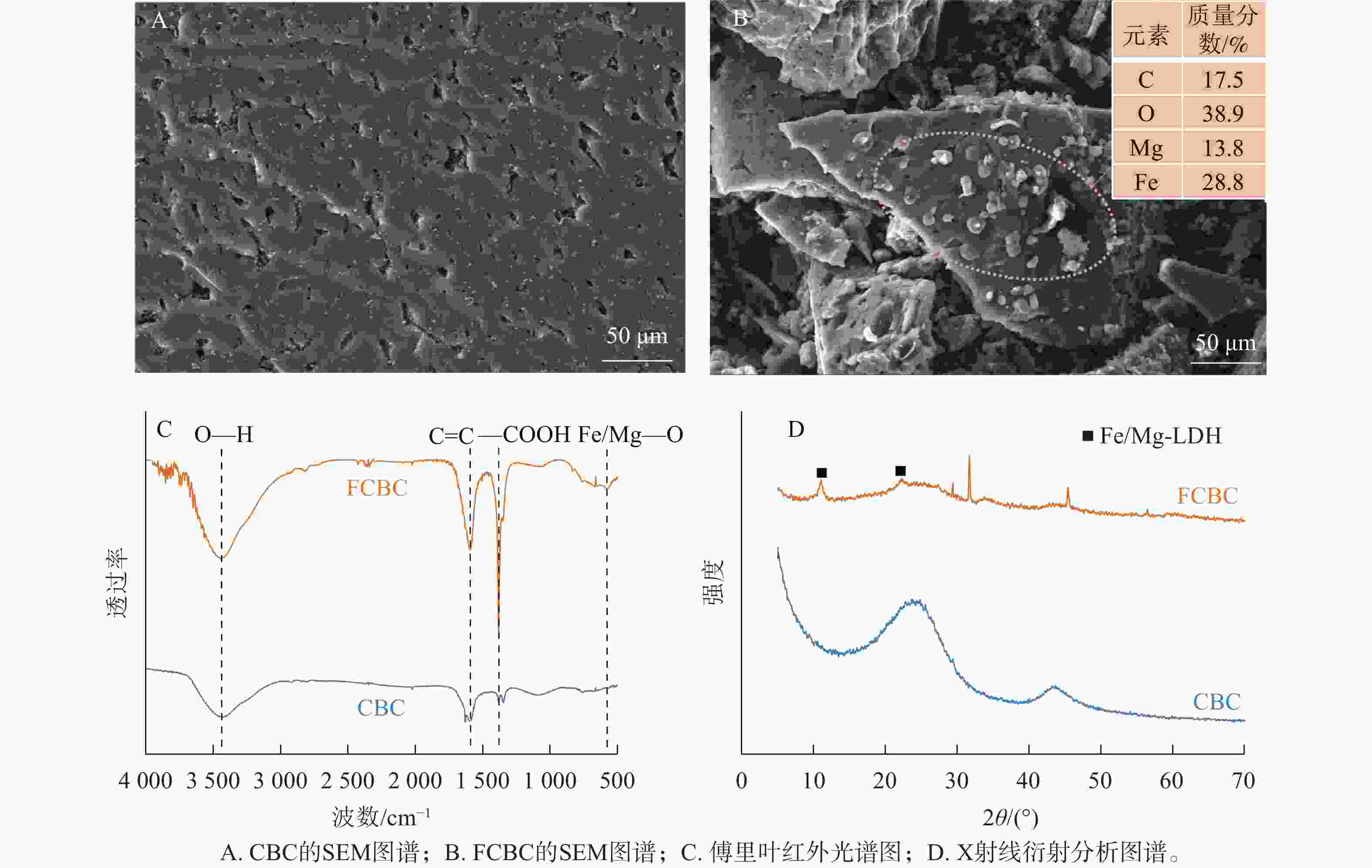
图 1 供试生物炭的形貌和表征
Figure 1. Morphology and characterization of biochar before and after modification
表 1 生物质炭的基本理化性质
Table 1. Selected physicochemical properties of the used biochars
生物炭 碳/
(g·kg−1)氮/
(g·kg−1)氧/
(g·kg−1)氢/
(g·kg−1)比表面积/
(m2·g−1)孔体积/
(cm3·g−1)CBC 744.0 3.0 204.0 34.0 1 017.36 0.46 FCBC 489.0 23.0 272.0 29.0 266.57 0.14 -
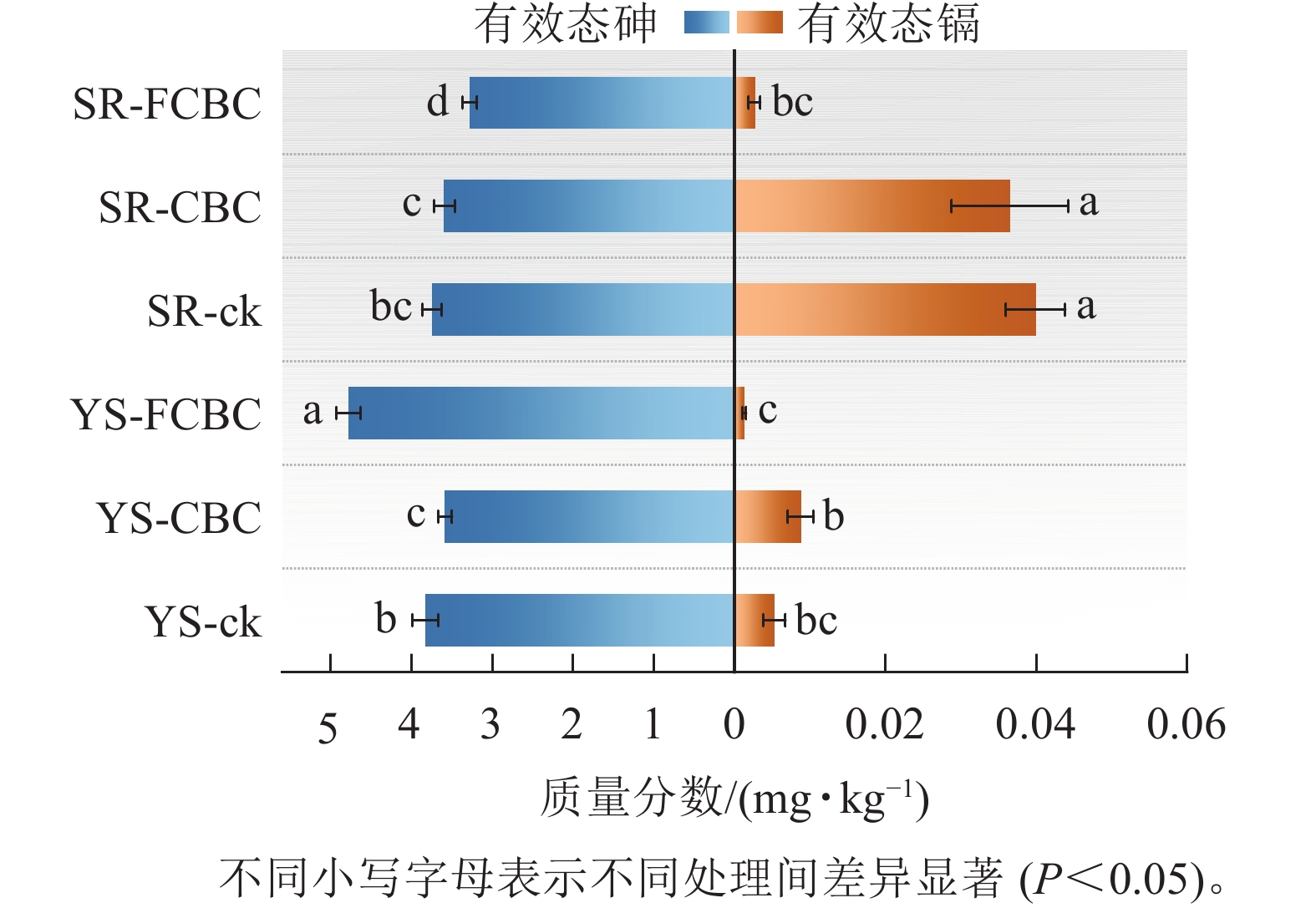
如图2所示:与SR-ck处理相比,SR-CBC处理下有效态砷质量分数差异不显著,而SR-FCBC处理下土壤有效态砷质量分数显著降低12.5%(P<0.05),表明在湿润条件下,FCBC对砷具有显著的钝化效果,而CBC钝化效果不显著。该现象可能归因于:①CBC表面缺少与砷阴离子有较强亲和力的结合位点(如Fe—O),这与戴志楠等[17]的研究结果一致。②FCBC施入土壤后引入了更多Fe3+,在氧化条件下易与砷形成Fe-As共沉淀物[18]。此外,FCBC表面的Fe—OH活性官能团与$\mathrm{AsO}_4^{3-} $等阴离子形成稳定的Fe—O—As络合物,促进有效态砷向铁氧化物结合态等更稳定形态转化[6]。
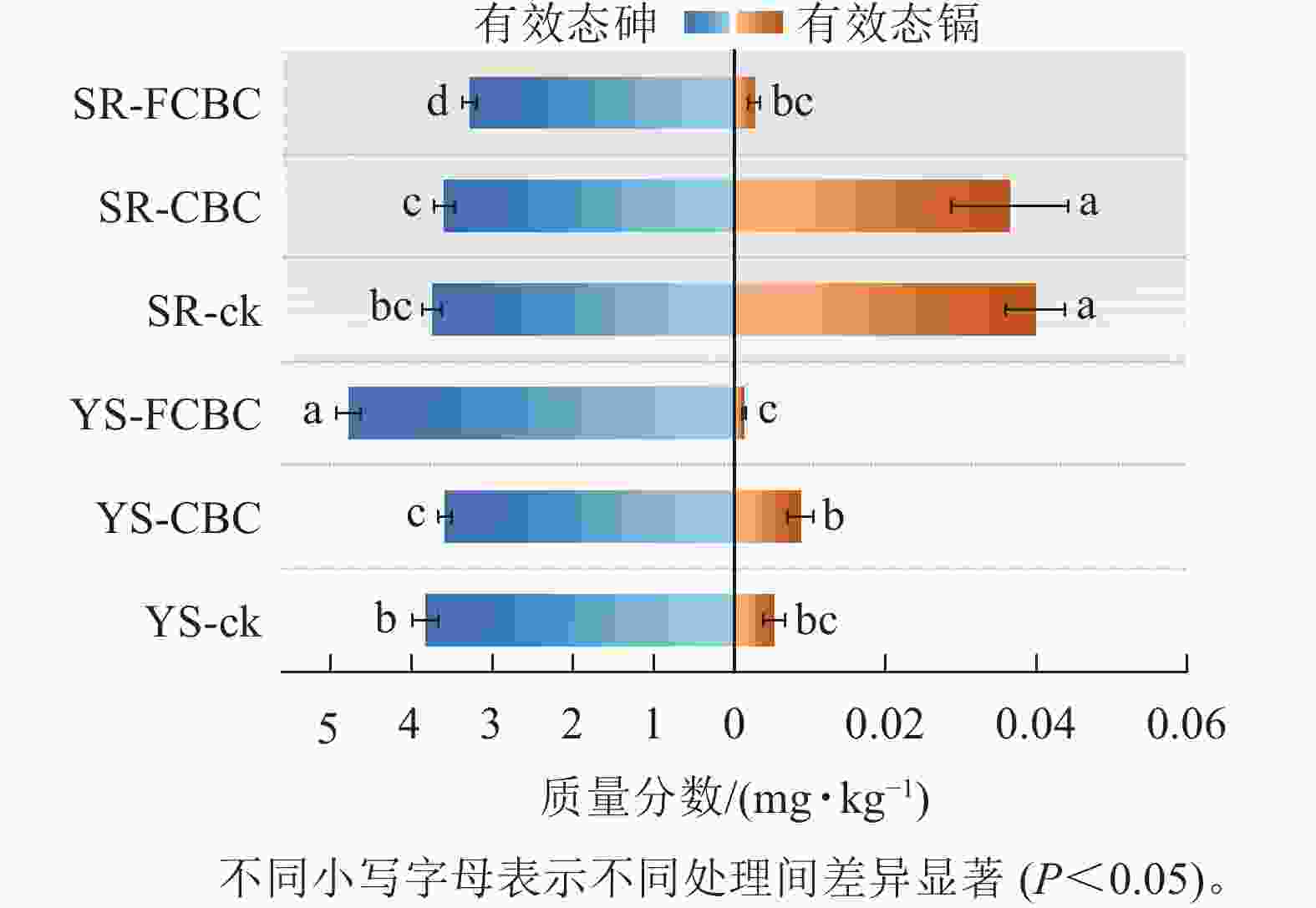
图 2 不同处理下土壤有效态砷镉质量分数
Figure 2. Concentrations of available arsenic and cadmium in soil under different treatments
与YS-ck和YS-CBC处理相比,YS-FCBC处理下土壤有效态砷质量分数显著(P<0.05)升高24.95%~33.45%,表明在厌氧条件下FCBC的施入反而增强了砷的生物有效性和迁移潜势。其原因可能主要包括以下2个方面:①FCBC施入土壤后可提高土壤pH,进而增加土壤颗粒表面的负电荷,通过静电排斥作用促进表面吸附态砷阴离子(如$\mathrm{H}_2 \mathrm{AsO}_4^{-} $)的释放[19];②土壤溶解性有机碳与砷具有较强的亲和力,易形成As-DOC络合物,而FCBC的施入提高了土壤中溶解性有机碳的质量分数,从而促进砷的解吸和迁移[20]。③FCBC施入土壤后可作为电子供体促进电子转移,从而加速淹水水稻土中铁氧化物的还原性溶解,进而导致铁氧化物结合态砷的释放[21]。
YS-FCBC处理下土壤有效态镉质量分数相较YS-CBC显著(P<0.05)降低86.20%,相较YS-ck降低76.92%;同时,与SR-ck和SR-CBC处理相比,SR-FCBC处理下土壤有效态镉质量分数显著(P<0.05)降低92.88%~93.50%。上述结果表明:在不同水分条件下,FCBC均能显著降低土壤中镉的生物有效性,其钝化效果优于CBC。该结果可能归因于以下机制:①FCBC可提高土壤pH,增加铁氧化物表面的负电荷密度,从而为Cd2+提供更多吸附位点;同时,pH的升高有利于碳酸盐结合态镉(CdCO3)的生成,增强镉的固定[22]。②FCBC表面的Mg/Fe-LDH吸附位点与Cd2+发生化学反应(络合、沉淀),促进有效态镉向铁氧化物结合态镉转化[16, 23]。此外,湿润环境中土壤有效态镉质量分数显著(P<0.05)高于淹水环境,归因于厌氧条件下硫酸盐还原菌介导的$\mathrm{SO}_4^{2-} $还原过程,可通过生成难溶性的硫化亚铁(FeS)固定镉,或由HS−/S2−与Cd2+反应形成硫化镉(CdS)沉淀,从而降低镉的生物有效性[5]。同时,厌氧环境中铁的还原与再分配过程有利于形成比表面积更大、表面官能团更丰富的无定形铁氧化物,进一步增强其对镉的吸附和固定能力[24]。
-
表2展示了不同处理下土壤理化性质的变化特征。与2种水分管理方式不施生物炭处理(ck)和CBC处理相比,FCBC的施用显著(P<0.05)提高了土壤pH,归因于铁镁改性过程中在生物炭表面引入了大量羟基等碱性官能团,施入土壤后提升其pH。淹水处理的土壤pH均显著(P<0.05)高于湿润处理,这可能与厌氧条件下铁、锰氧化物及硫酸盐的还原反应持续消耗H+有关[24]。厌氧环境下施入CBC和FCBC均使土壤氧化还原电位显著降低至−473.45~−498.51 mV (P<0.05)。这是由于生物炭作为还原性物质施入土壤后,加速氧化耗氧过程,导致氧化还原电位显著降低[25]。生物炭处理均显著(P<0.05)提高了土壤有机质质量分数,增幅为9.15%~28.47%,归因于生物炭本身富含稳定有机碳组分,施入土壤后直接增加有机碳库。与ck、CBC相比,FCBC处理下土壤有效磷质量分数未出现显著变化,归因于30 d的短期培养难以影响土壤磷组分的变化。与ck、FCBC相比,CBC处理显著(P<0.05)提高了土壤速效钾质量分数,表明原始生物炭能有效提升土壤速效钾。
表 2 不同处理下稻田土的理化性质
Table 2. Physicochemical properties of paddy soil under different treatments
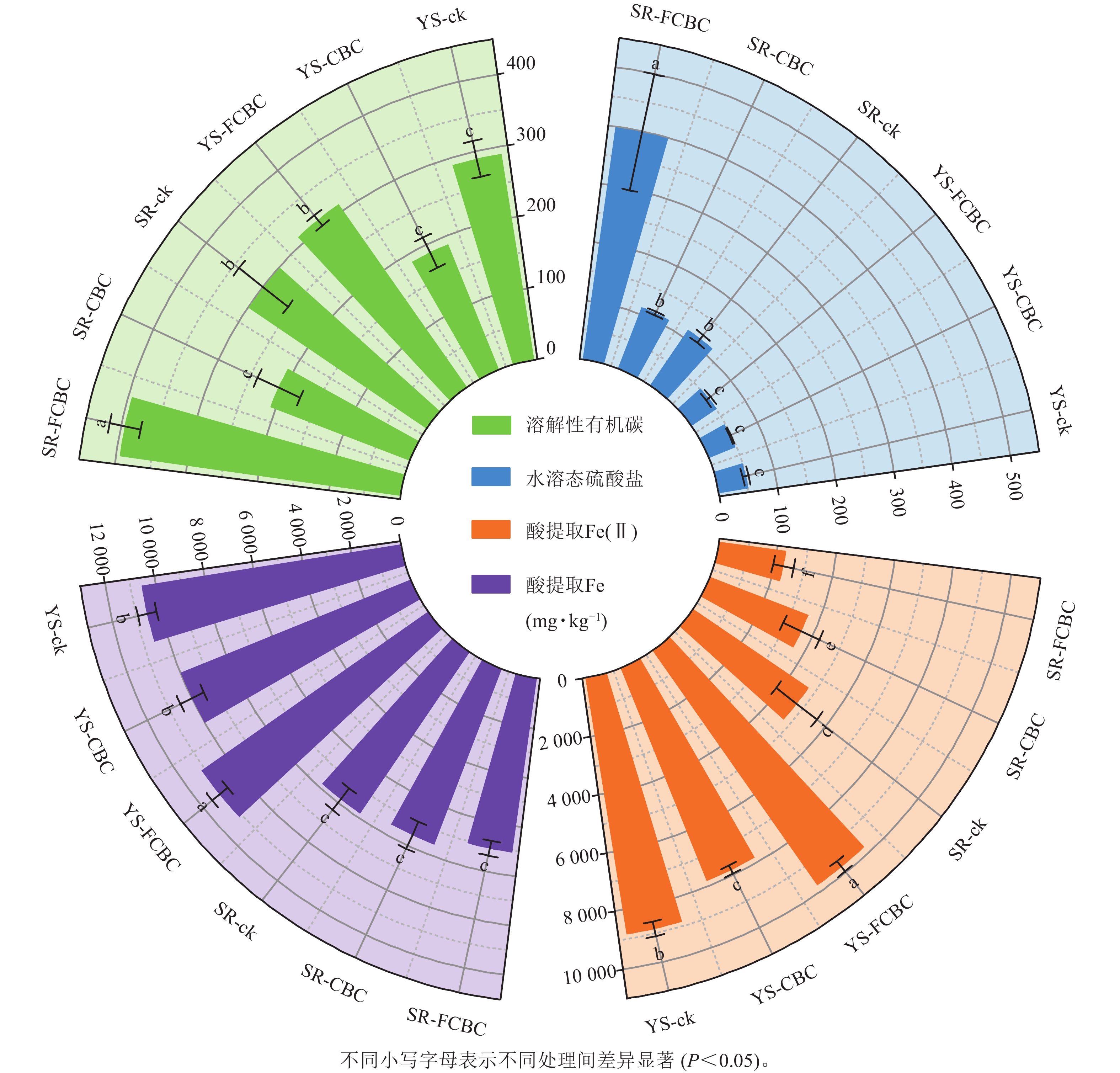
处理 pH 氧化还原电位/mV 有机质/(g·kg−1) 有效磷/(mg·kg−1) 速效钾/(mg·kg−1) YS-ck 6.05±0.02 c −204.13±4.50 b 28.69±1.01 d 10.69±0.82 c 67.31±4.05 c YS-CBC 6.11±0.08 c −473.45±6.68 c 36.86±1.06 a 12.41±0.50 ab 162.47±15.49 a YS-FCBC 7.38±0.05 a −498.51±8.42 c 34.98±1.08 b 12.01±0.75 b 58.39±6.79 cd SR-ck 5.69±0.09 e 275.65±32.66 a 31.80±1.27 c 11.80±0.99 b 46.87±1.53 d SR-CBC 5.87±0.05 d 310.07±14.81 a 36.83±0.49 a 13.37±0.53 a 93.02±16.32 b SR-FCBC 6.99±0.11 b 235.15±22.93 a 34.71±0.51 b 12.25±0.55 b 51.56±7.27 cd 说明:同列不同小写字母表示不同处理间差异显著(P<0.05)。 如图3所示,淹水处理下土壤酸提取Fe质量分数显著(P<0.05)高于湿润处理,表明还原条件下高结晶度铁氧化物发生还原溶解,形成无定形水铁矿或黄铁矿等次生矿物[5]。相较于ck和CBC处理,淹水环境下FCBC显著(P<0.05)提升了酸提取Fe(Ⅱ)的质量分数,增幅最高达25.00%。这是因为FCBC表面的Fe/Mg—O键与Fe/Mg-LDH结构具备良好的电子传递能力更有利于促进土壤电子转移,加速淹水环境下土壤Fe(Ⅲ)的还原。WU等[21]研究发现:生物炭作为电子供体,其施入提高了水稻土中盐酸可提取态Fe(Ⅱ)/Fe(Ⅲ)质量分数。与ck和CBC处理相比,FCBC施入显著(P<0.05)提高了土壤溶解性有机碳质量分数,这可能是由于FCBC促进了铁结合态有机质的释放与分解,从而增加了土壤中溶解性有机碳[26]。淹水环境下土壤水溶态硫酸盐质量分数显著(P<0.05)低于湿润环境,表明在厌氧条件下硫酸盐大量还原为硫化物。此外,与SR-ck和SR-CBC处理相比,SR-FCBC处理下土壤硫酸盐质量分数显著(P<0.05)升高,说明FCBC的施用对土壤硫循环过程具有一定的调节作用。
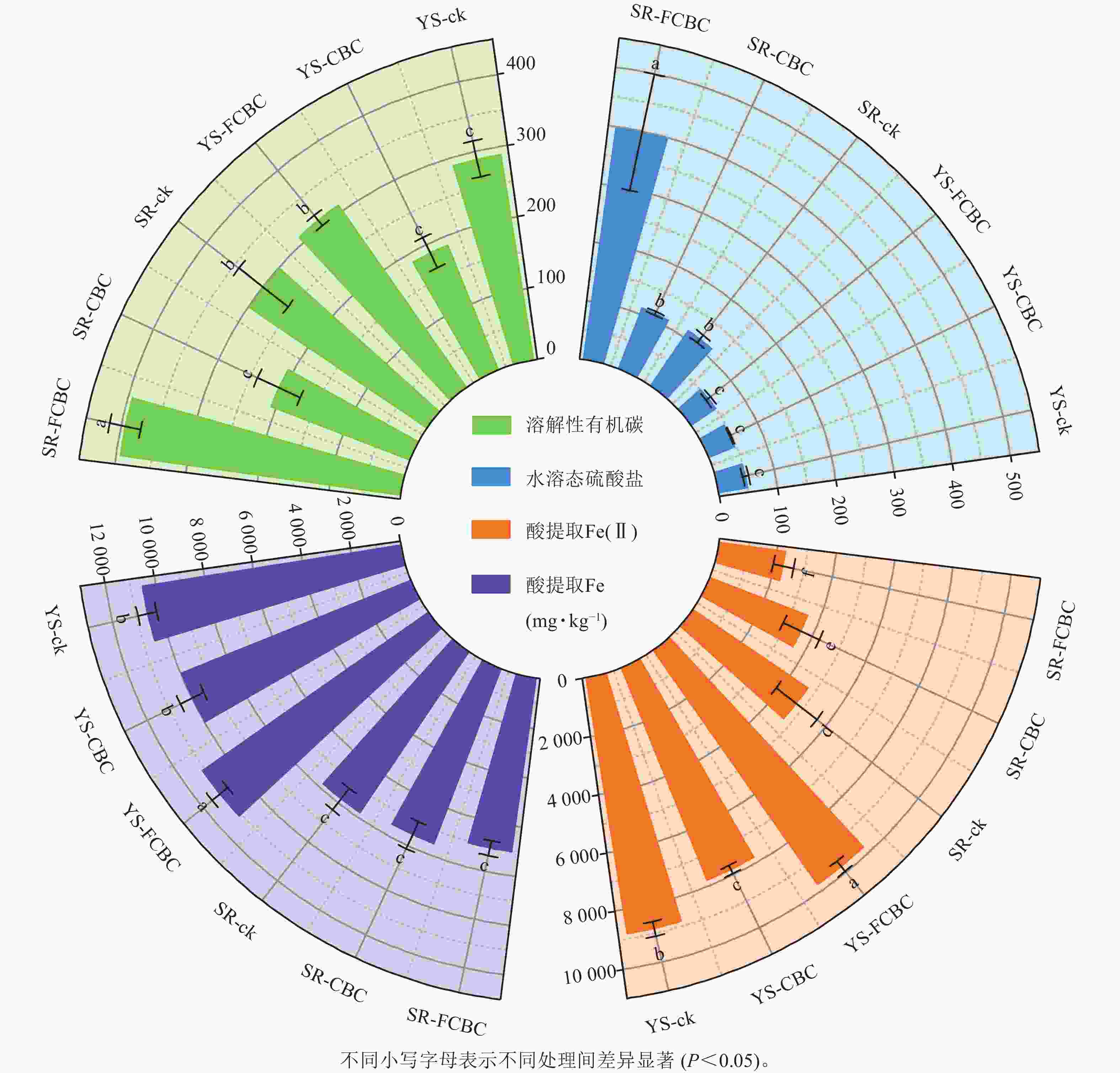
图 3 不同处理下土壤鲜土指标
Figure 3. Physicochemical properties of fresh soil under different treatments
-
土壤微生物多样性及群落结构变化可作为评价重金属污染土壤修复效果的重要生态指标。如表3所示,与ck和CBC处理相比,FCBC处理下土壤细菌群落的Ace指数、Chao1指数和Shannon-Wiener指数均显著降低(P<0.05),而Simpson指数升高,表明FCBC的施用显著降低了细菌群落的物种丰富度和多样性,使群落结构趋于由少数优势类群主导。根据ZHANG等[27]的研究,pH能驱动土壤微生物群落结构与多样性变化。因此FCBC处理引起的土壤pH变化可能是导致细菌群落多样性和丰富度下降的重要原因。
表 3 不同处理下土壤细菌群落多样性指数
Table 3. Diversity indices of soil bacterial communities under different treatments
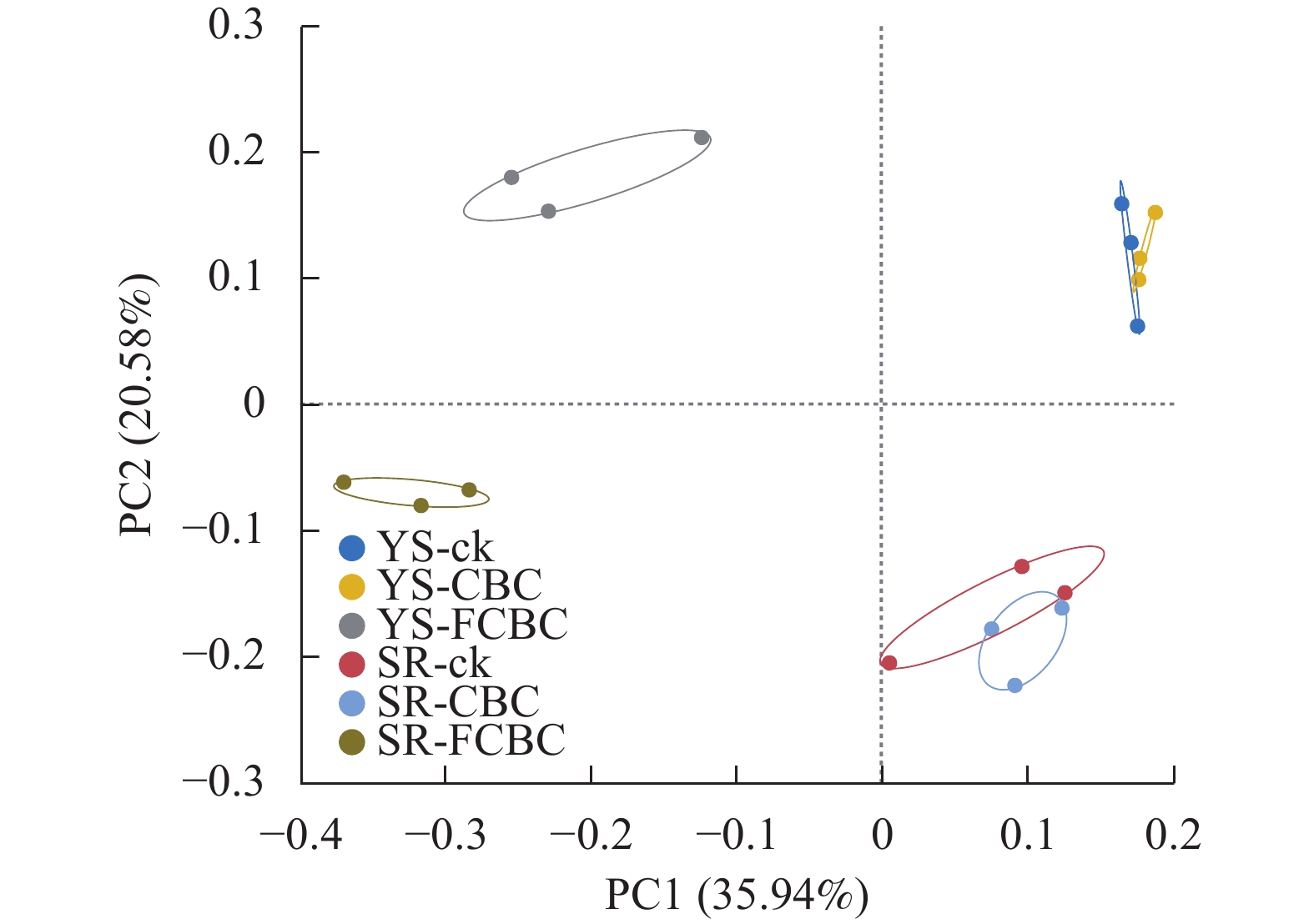
处理 Ace指数 Chao 1指数 Shannon-Wiener指数 Simpson指数 YS-ck 2 410.5±105.8 a 2 375.1±117.0 a 5.86±0.17 a 0.009±0.002 bc YS-CBC 2 500.7±122.4 a 2 496.3±111.5 a 5.90±0.17 a 0.009±0.002 bc YS-FCBC 2 143.5±175.6 b 2 126.4±172.9 b 5.63±0.04 b 0.011±0.002 b SR-ck 2 417.4±61.9 a 2 402.7±58.3 a 5.96±0.10 a 0.007± 0.001 cSR-CBC 2 558.8±33.2 a 2 530.6±60.6 a 6.05±0.04 a 0.007±0.001 c SR-FCBC 2 113.5±131.7 b 2 079.4±193.0 b 5.47±0.10 b 0.015±0.002 a 说明:同列不同小写字母表示不同处理间差异显著(P<0.05)。 主坐标分析(PCoA)结果显示(图4):FCBC处理与ck和CBC处理在PC1轴上呈现出明显分离,而淹水与湿润2种水分管理方式在PC2轴上分离,表明FCBC的施用和水分条件均对稻田土壤细菌群落组成产生了显著影响,且两者在驱动群落差异方面具有不同的贡献方向。
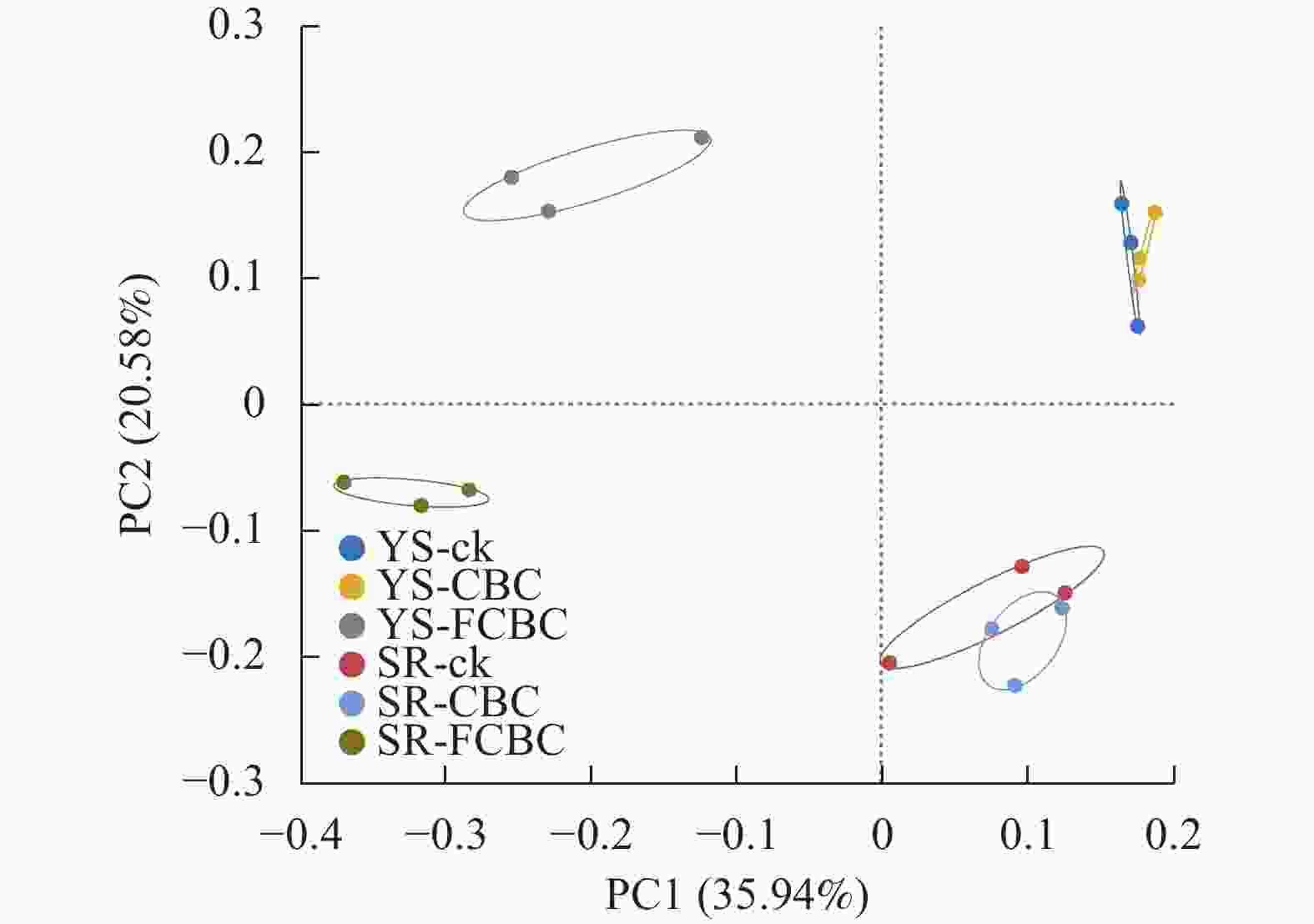
图 4 不同处理下土壤细菌群落的PCoA分析
Figure 4. PCoA analysis of soil bacterial communities of species numbers under different treatments
-
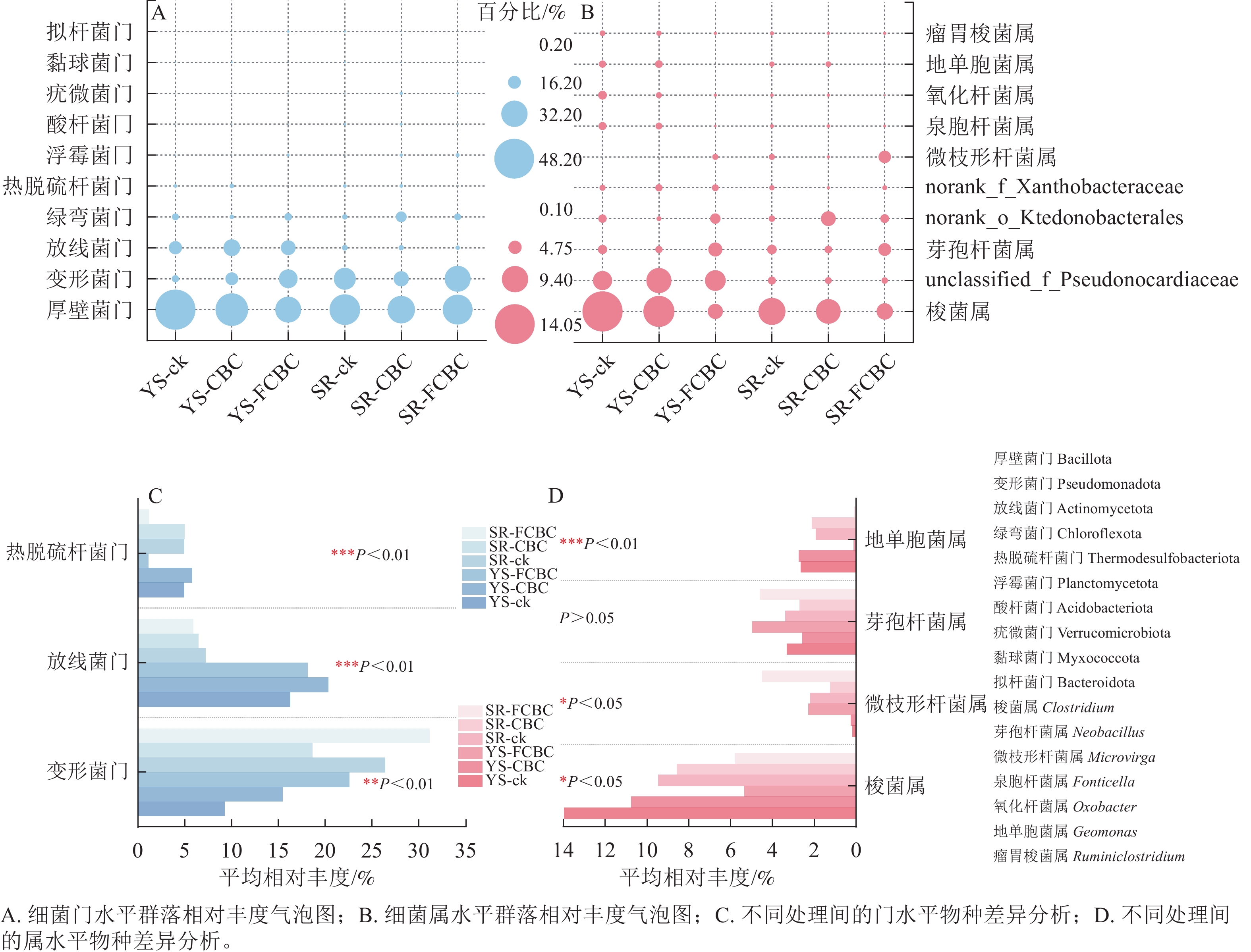
在门水平上,稻田土壤细菌群落主要由厚壁菌门Bacillota、变形菌门Pseudomonadota、放线菌门Actinomycetota、绿弯菌门Chloroflexota、热脱硫杆菌门Thermodesulfobacteriota等组成(图5A)。在属水平上,优势菌属主要包括梭菌属Clostridium、芽孢杆菌属Neobacillus、微枝形杆菌属Microvirga、泉胞菌属Fonticella和地单胞菌属Geomonas等(图5B)。
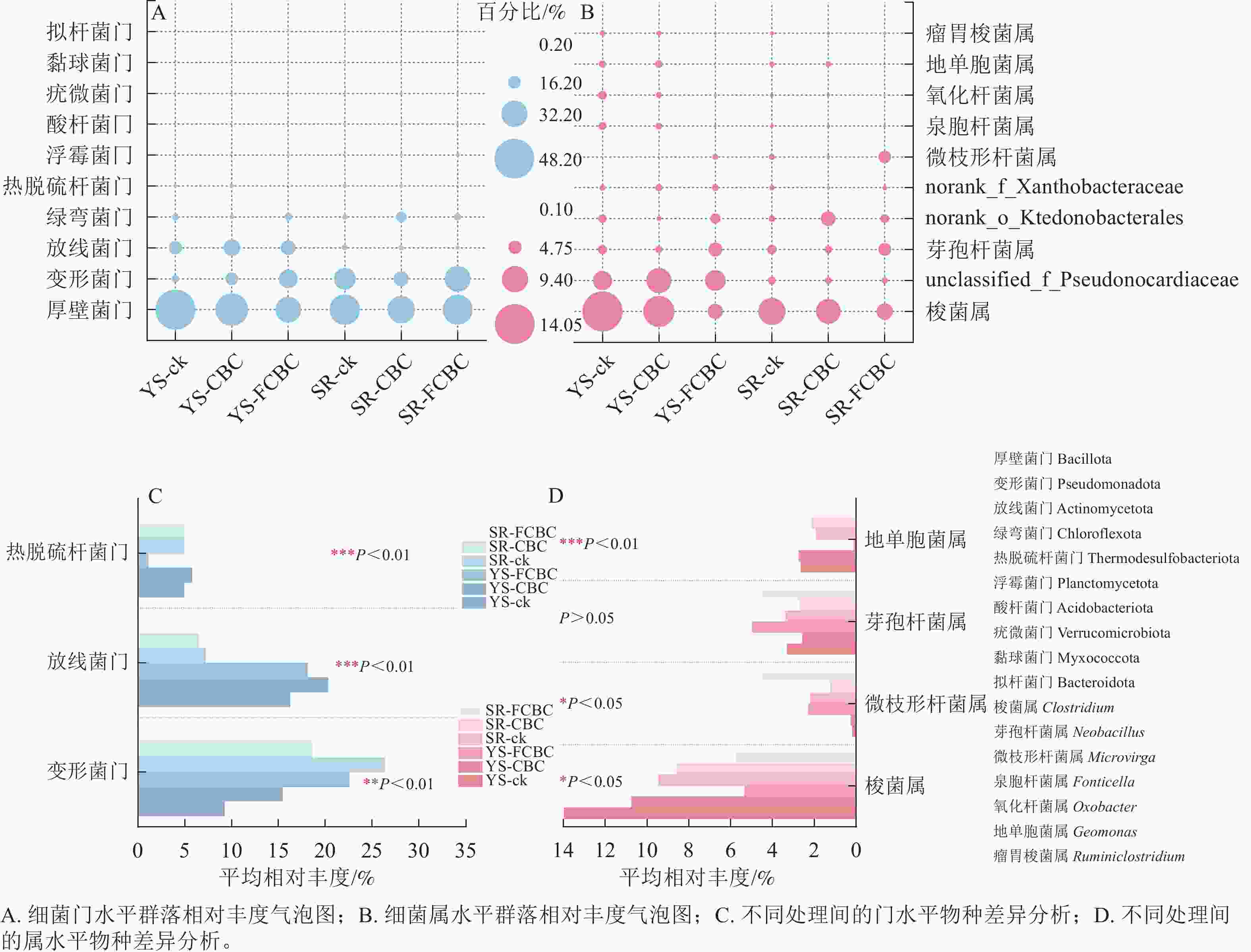
图 5 不同处理下土壤细菌群落丰度分布及物种差异分析
Figure 5. Relative abundance of soil bacterial communities and species difference analysis under different treatments.
如图5C所示,与ck和CBC处理相比,FCBC的施用降低了热脱硫杆菌门的相对丰度,提高了变形菌门的相对丰度。变形菌门具有较强的环境适应能力,可分泌铁载体、有机酸及胞外聚合物,通过络合或吸附作用参与重金属的固定过程[28],因而在好氧环境下能有效钝化砷和镉。与湿润处理相比,淹水处理降低了变形菌门的相对丰度,同时提高了放线菌门的相对丰度。放线菌门具有螯合特性,通过与镉发生螯合进一步增强厌氧环境下镉的固定[16],放线菌门还可参与铁循环过程,通过还原反应产生Fe(Ⅱ)[29],这进一步影响厌氧环境下铁氧化物结合态砷的释放。
如图5D所示,与ck和CBC处理相比,FCBC处理提高了微枝形杆菌属和芽孢杆菌属的相对丰度,降低了梭菌属和地单胞菌属的相对丰度;与湿润处理相比,淹水处理提高了梭菌属的相对丰度,同时降低了微枝形杆菌属的相对丰度。梭菌属作为典型的硫酸盐还原菌,能够在土壤中参与硫循环将$\mathrm{SO}_3^{2-} $还原为S2−,厌氧环境下该菌属相对丰度的提升通过促进硫化镉的形成,更有效固定镉[30]。微枝形杆菌属能够合成铁载体进而增强对重金属的固定能力。芽孢杆菌属在稻田土壤的铁与砷生物地球化学循环中具有重要作用,其在厌氧条件下能驱动异化铁还原过程。FCBC的施入通过提升其相对丰度促进淹水环境下含砷铁矿物中无机砷的释放,增强砷的迁移性。芽孢杆菌属还可将有机砷转化为无机砷,从而影响土壤中砷的形态分布[31]。此外,隶属于芽孢杆菌属的部分磷酸盐活化细菌能够提高土壤磷的溶解度,并通过沉淀反应对金属阳离子镉产生钝化作用[32]。
-
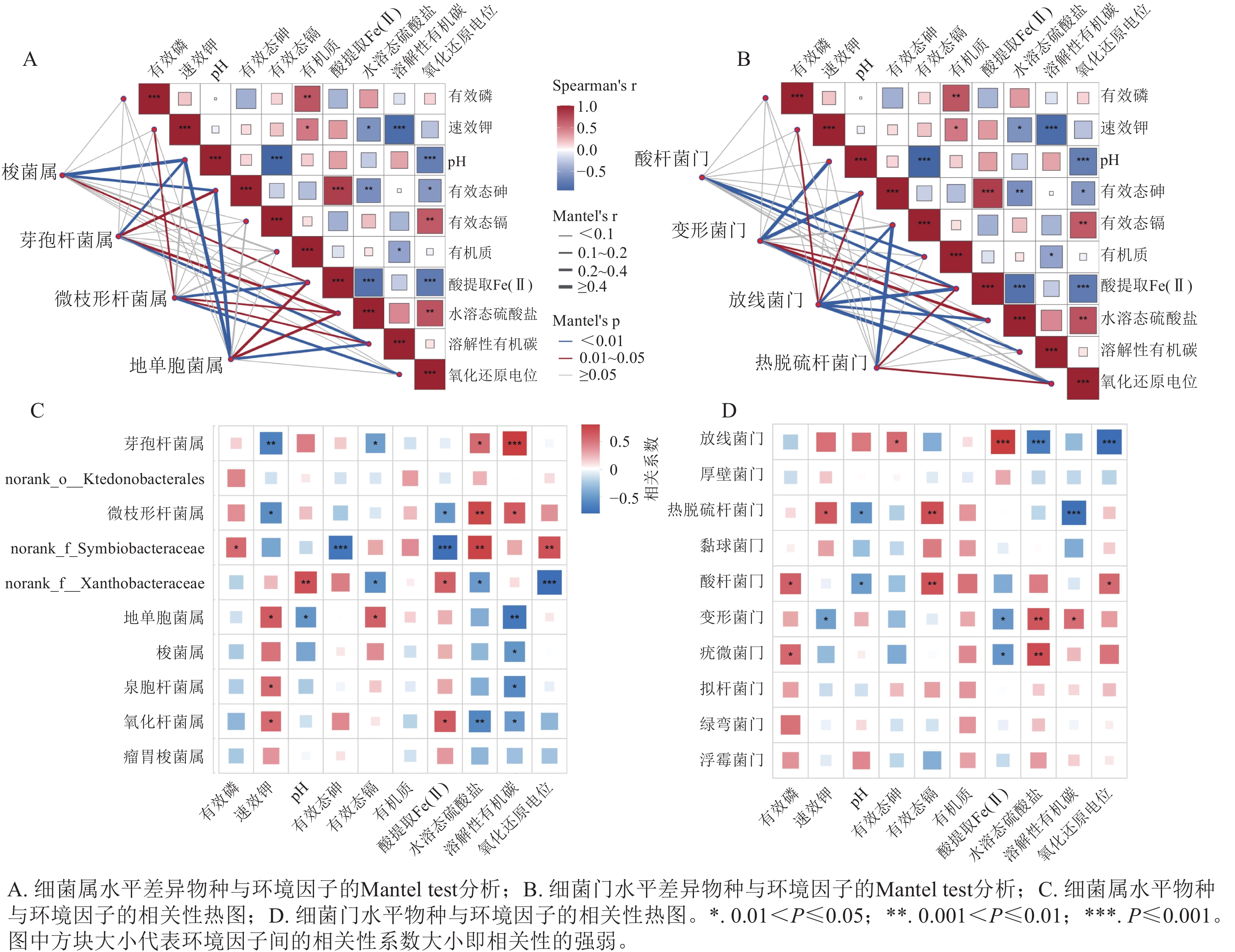
在门和属水平上对差异物种类群与环境因子进行了Mantel分析(图6A、B)。有效态砷与酸提取态Fe(Ⅱ)呈极显著(P<0.001)正相关,而与水溶态硫酸盐和氧化还原电位呈显著(P<0.05)负相关,表明不同生物炭类型及水分管理方式通过驱动Fe(Ⅱ)、水溶态硫酸盐和氧化还原电位的变化,进而调控土壤中砷的生物有效性。有效态镉与氧化还原电位呈显著(P<0.01)正相关,与pH呈极显著(P<0.001)负相关,说明pH和氧化还原电位是影响镉钝化与迁移转化的关键环境因子[22]。Mantel分析结果进一步表明:环境因子对门、属水平上差异物种类群的分布具有显著影响。
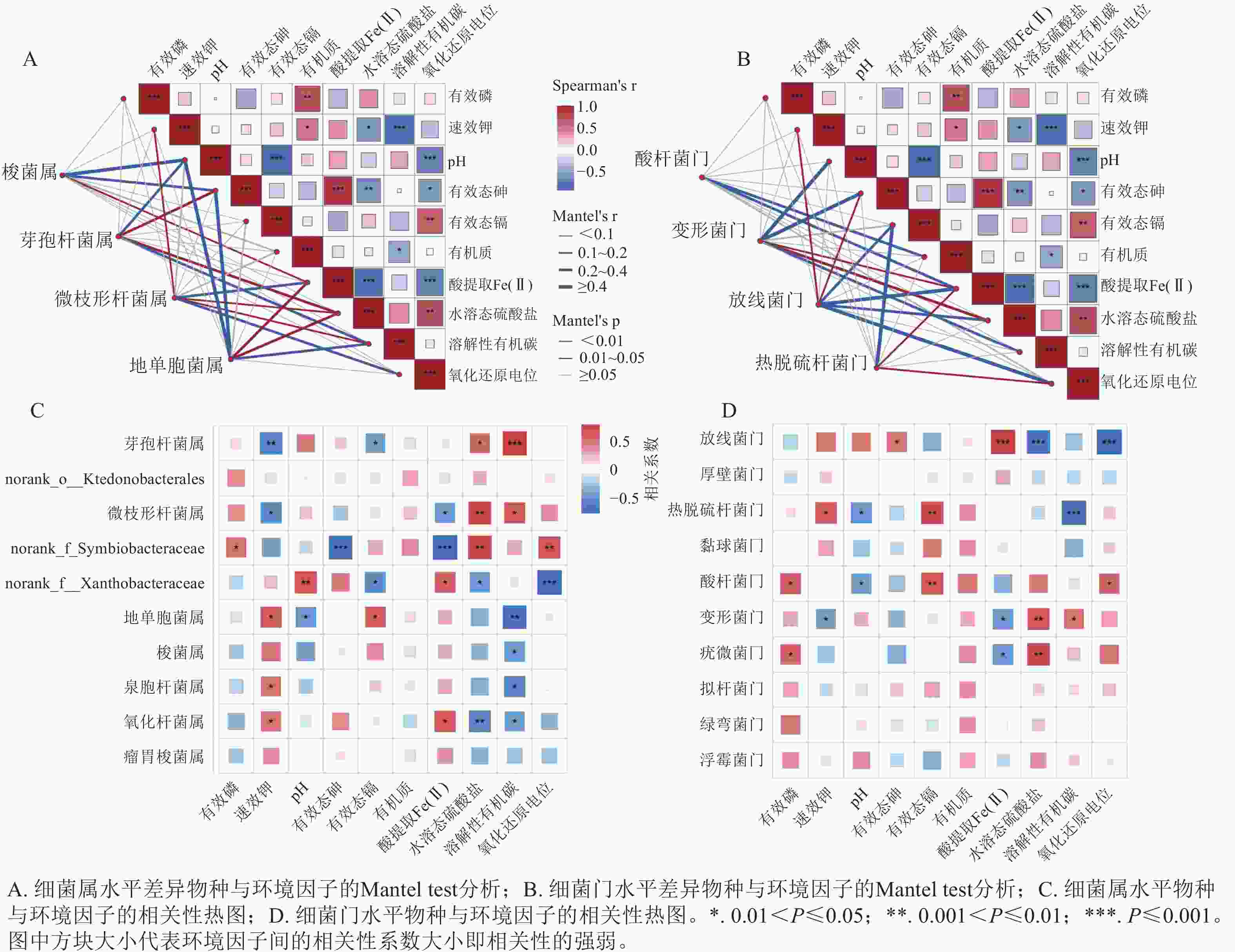
图 6 环境因子及土壤细菌群落与环境因子的关联分析
Figure 6. Correlation analysis among environmental factors and between soil bacterial communities and environmental factors
如图6C、D所示:放线菌门与水溶态硫酸盐和氧化还原电位呈极显著(P<0.001)负相关,而与Fe(Ⅱ)呈极显著(P<0.001)正相关,表明厌氧环境驱动放线菌门相对丰度的提升。该菌门可能通过还原水溶态硫酸盐和Fe(Ⅲ)将氧化环境转变为还原环境,从而影响砷、镉的迁移转化过程[33]。酸杆菌门与有效态镉质量分数呈显著(P<0.01)正相关,表明该菌门对镉胁迫具有较强的适应能力,其相对丰度随生物有效态镉质量分数的降低而降低[34]。在属水平上,溶解性有机碳与梭菌属和地单胞菌属的相对丰度呈显著(P<0.05)负相关,而与微枝形杆菌属和芽孢杆菌属呈显著(P<0.05)正相关,表明FCBC可能通过调控土壤溶解性有机碳及其组成特征,进而重塑优势细菌属的分布格局。
-
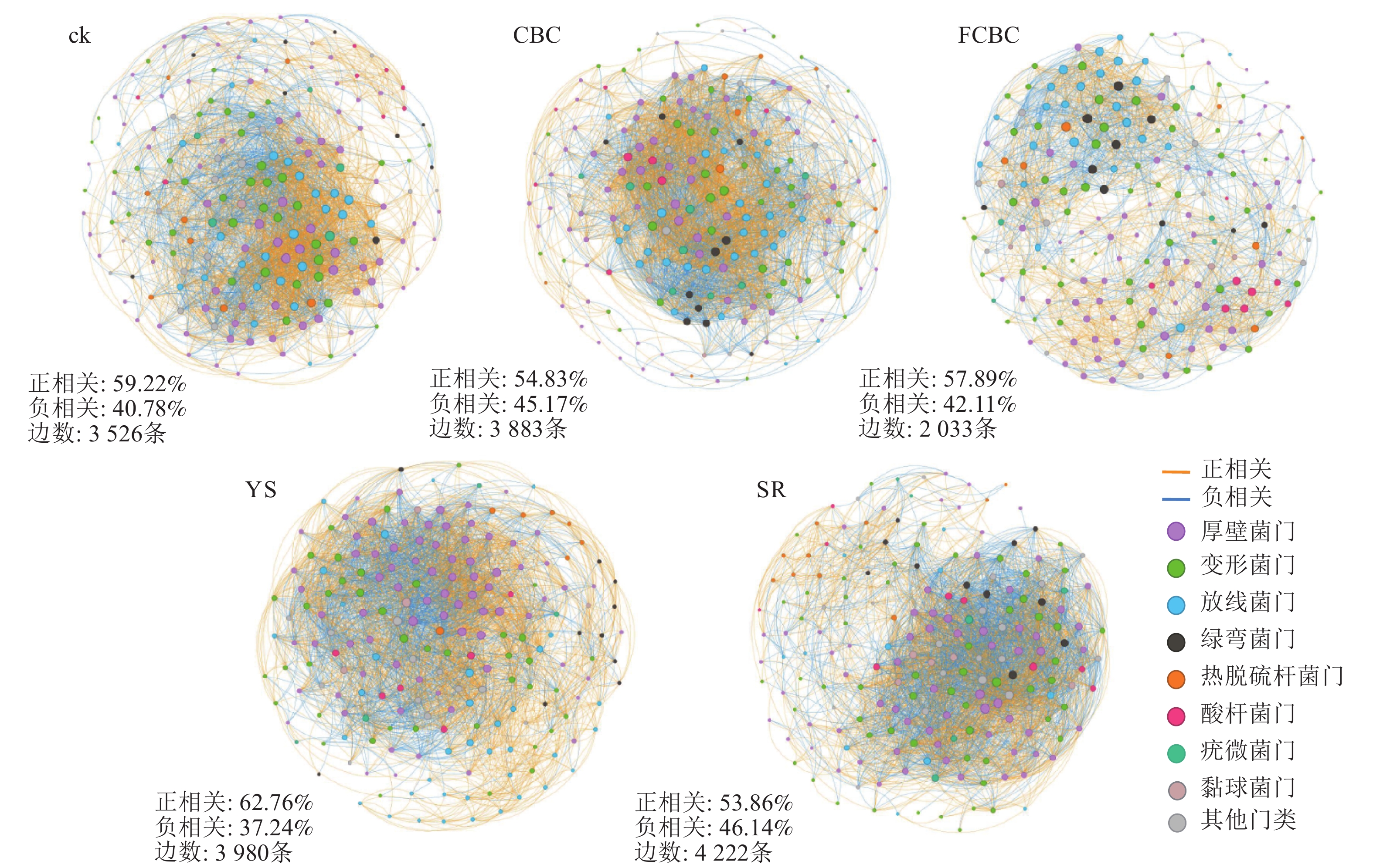
微生物共现网络分析可用于揭示微生物类群之间的潜在关联模式及相互作用特征。如图7所示,分布于模块枢纽(module hubs)和连接体(connectors)类型的关键物种多隶属于厚壁菌门、变形菌门和放线菌门等。FCBC处理下的共现网络连接数量(2 033条)显著低于ck和CBC处理(分别为3 526和3 883条),且网络结构整体更加松散,表明FCBC施用降低了土壤微生物网络的复杂性和稳定性,削弱了细菌物种间的相互作用强度[35]。该现象可能由以下2个原因导致:①FCBC在土壤中释放的铁、镁离子可能对部分微生物产生一定的选择性胁迫,从而削弱其与其他类群之间的共生代谢关系。②TIAN等[35]研究发现:高重金属污染下土壤微生物群落会构建更为复杂的网络结构来增强对环境胁迫的适应能力,而FCBC能更有效地钝化土壤中的重金属,可能使网络的复杂性降低。从相关性类型来看,厌氧环境下网络中正相关连接线比例(62.76%)显著高于好氧环境(53.86%),表明厌氧条件下稻田土壤细菌之间的协同与互利共生关系增强,而竞争关系相对减弱[36]。这可能是由于厌氧环境中可利用的氧气及电子受体受限,微生物需通过代谢互补和协同作用来提高资源利用效率。
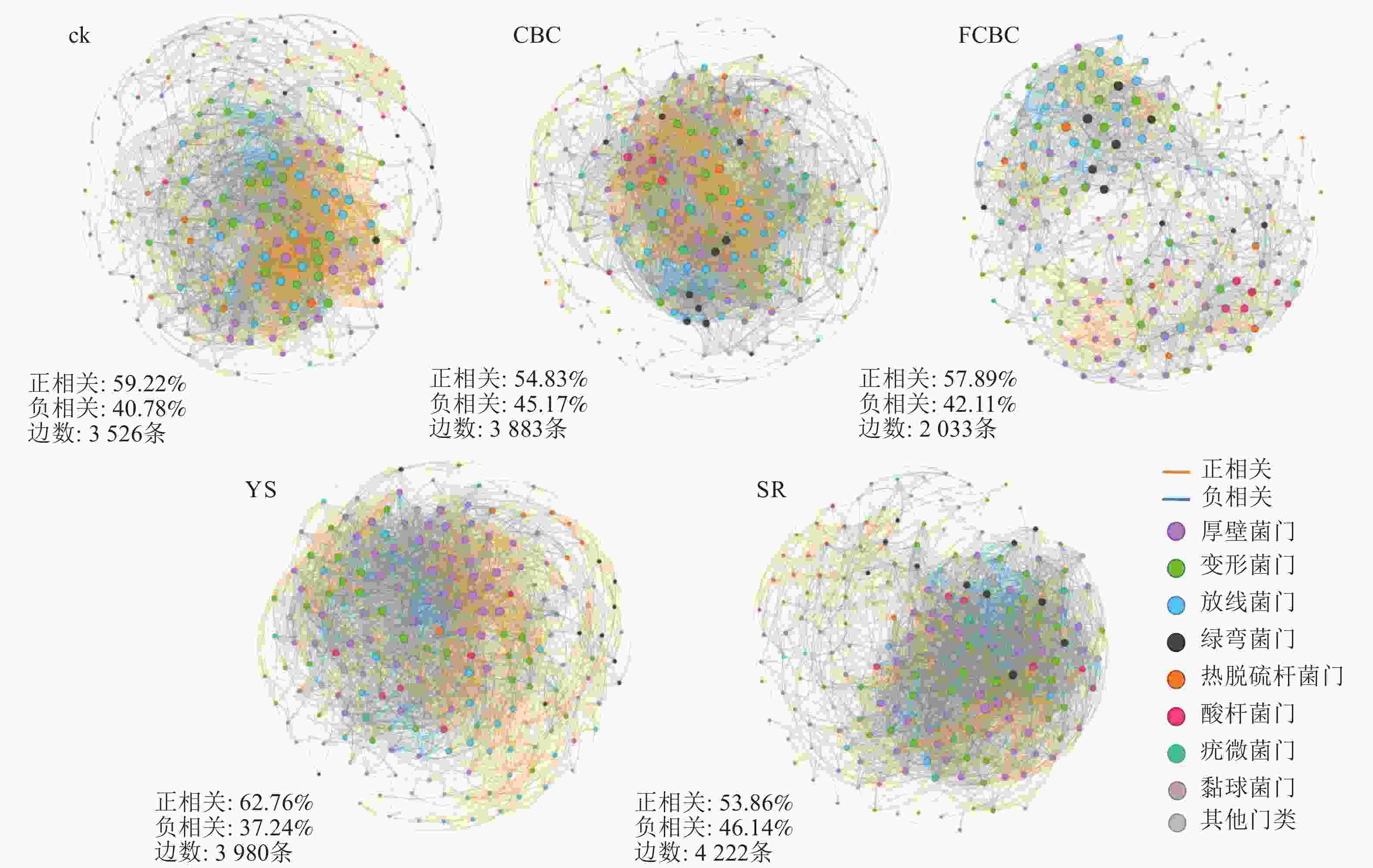
图 7 土壤细菌群落共现网络结构特征
Figure 7. Co-occurrence network structure of soil bacterial communities
-
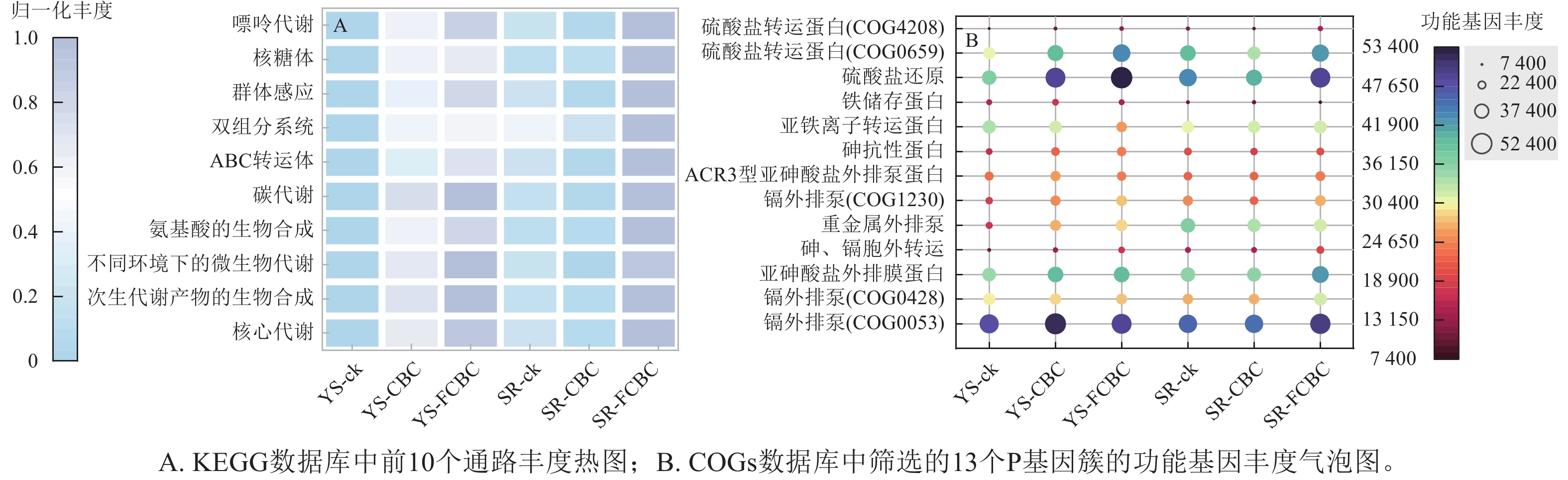
基于PICRUSt2对土壤细菌群落的功能潜力进行了预测分析,并对前10位丰度的京都基因与基因组百科全书(KEGG)三级代谢通路进行比较(图8A)。各处理下土壤细菌群落的核心功能主要集中于碳水化合物代谢、ATP结合盒(ABC)转运系统以及氨基酸合成与代谢等基础生命过程相关通路,这些功能对于维持微生物的生长、代谢和环境适应能力具有重要意义[37]。与ck和CBC处理相比,FCBC的施入显著提高了上述功能通路的相对丰度,表明FCBC有助于增强土壤微生物的代谢潜力和环境适应性。
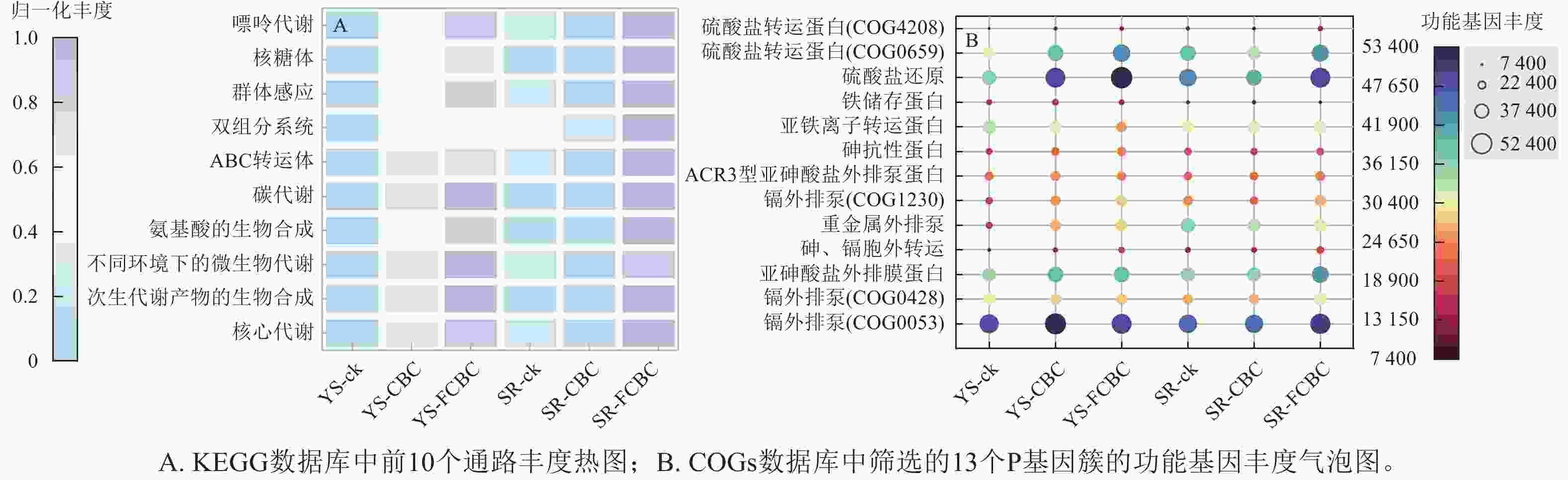
图 8 PICRUSt2预测的土壤细菌群落功能基因丰度
Figure 8. Functional gene abundance of soil bacterial communities predicted by PICRUSt2
本研究还选取同源蛋白簇(COGs)的P类功能注释中,与砷镉抗性、铁转运以及硫循环相关的8个基因进行丰度比较(图8B) [11]。与ck和CBC处理相比,在好氧条件下FCBC的施入显著提高了大多数重金属抗性相关基因的丰度,而在淹水条件下该提升效应不明显,表明FCBC在好氧环境下可通过上调砷镉转运与外排相关蛋白的表达,增强土壤微生物对重金属的耐受性,尤其是强化镉的外排机制,将Cd2+从细胞质或细胞膜转移至胞外,从而降低其细胞毒性[38]。此外,FCBC的施入均提高了与硫转运和硫代谢相关基因的丰度,说明FCBC通过调控土壤硫的转运与转化过程,可能间接促进砷、镉的钝化固定[5]。
-
本研究表明:铁镁改性显著增强了椰壳生物炭在淹水和湿润条件下对土壤镉的钝化能力,而对砷的钝化效果表现出明显的水分依赖性,即在湿润条件下可降低砷的生物有效性,而在淹水条件下促进了砷的迁移转化。此外,铁镁改性生物炭的施用重塑了土壤细菌群落结构且显著降低了细菌α多样性和土壤细菌共现网络复杂性。同时,改性生物炭可选择性富集与土壤矿质元素循环及重金属调控相关的优势类群,从而影响土壤砷镉的迁移转化。另外,不同水分管理方式也可显著影响细菌群落组成。在好氧条件下,铁镁改性生物炭通过增强与重金属转运、抗性蛋白表达及氨基酸和有机质代谢相关的功能通路,提高了微生物介导的重金属迁移转化。
综上所述,水分管理与铁镁改性椰壳生物炭通过调控土壤理化环境和微生物群落结构与功能,显著影响了稻田土壤中砷、镉的生物有效性和形态转化。由于本研究为短期室内培养试验,结果尚未结合作物生长及田间实际环境条件进一步验证,因此,后续可开展盆栽及田间试验,进一步探究铁镁改性椰壳生物炭在实际稻田生态系统中的修复效应。
本文已在中国知网网络首发,可在知网搜索、下载并阅读全文。
铁镁改性生物炭对热带稻田土壤砷镉形态及微生物群落的调控效应
DOI: 10.11833/j.issn.2095-0756.20260146
Regulatory effects of Fe-Mg modified biochar on arsenic and cadmium forms and microbial communities in tropical paddy soil
-
摘要:
目的 探究铁镁双金属改性椰壳生物炭(FCBC)在不同水分条件下对热带稻田土壤中砷、镉的钝化效果及对微生物群落结构的影响特征。 方法 于2025年8—9月开展了为期30 d的室内土壤培养试验,设置淹水与湿润2种水分处理,系统分析FCBC施加后土壤中砷、镉迁移特性,土壤理化性质及微生物群落结构的响应规律。 结果 与不加生物炭(对照)及未改性生物炭处理相比,在湿润条件下FCBC显著(P<0.05)降低了土壤中有效态砷(降幅为8.74%~12.5%)、有效态镉(降幅为92.88%~93.50%);在淹水条件下降低了有效态镉(降幅为76.92%~86.2%),但显著(P<0.05)促进了砷的迁移转化(增幅为24.95%~33.45%),增加其潜在生态风险。Mantel分析结果表明:FCBC对砷的钝化效果主要受水分变化驱动的铁、硫元素循环调控,而对镉的钝化则主要通过调节土壤pH等理化性质实现。FCBC的施加整体降低了土壤微生物α多样性,并改变了不同水分条件下的细菌群落结构。与其他处理相比,FCBC降低了热脱硫杆菌门Thermodesulfobacteriota、梭菌属Clostridium等类群的相对丰度,提高了变形菌门Pseudomonadota、微枝形杆菌属Microvirga和芽孢杆菌属Neobacillus等类群的相对丰度,进而影响砷、镉的迁移转化,且水分条件进一步影响了细菌群落的组成差异。共现网络分析表明:FCBC降低了土壤细菌网络的复杂性和稳定性,而厌氧环境增强了细菌物种间的协同共生关系。PICRUSt2功能预测结果显示:在好氧环境下FCBC的施入增强了与砷、镉抗性相关的蛋白功能,并促进了氨基酸代谢及有机质合成等关键代谢过程。 结论 水分管理与铁镁改性椰壳生物炭的协同作用通过调控微生物群落结构与功能,进而影响砷、镉在稻田土壤中的迁移转化过程。图8表3参38 Abstract:Objective The objective is to investigate the passivation effects of Fe-Mg bimetallic modified coconut shell biochar (FCBC) on arsenic (As) and cadmium (Cd) in tropical paddy soil under different moisture conditions, as well as its impact on microbial community structure. Method A 30-day indoor soil incubation experiment was performed from August to September 2025, with two moisture treatments: flooded and moist. The effects of FCBC on the mobility of As and Cd, soil physicochemical properties, and microbial community structure were systematically analyzed. Result Compared with the control and unmodified biochar, FCBC significantly (P<0.05) reduced the contents of available As (by 8.74%–12.5%) and available Cd (by 92.88%–93.50%) in soil under moist conditions. Under flooded conditions, it decreased the available Cd content (by 76.92%–86.2%), but significantly (P< 0.05) promoted the migration and transformation of As (by 24.95%–33.45%), thereby increasing its potential ecological risk. Mantel analysis revealed that the passivation effect of FCBC on As was mainly regulated by Fe and S cycling driven by moisture changes, while its effect on Cd was primarily achieved by altering soil pH and other physicochemical properties. The application of FCBC overall reduced soil microbial α-diversity and significantly altered the bacterial community structure under different moisture conditions. Compared with other treatments, FCBC significantly decreased the relative abundance of taxa such as Thermodesulfobacteriota, and Clostridium, and increased that of Pseudomonadota, Microvirga and Neobacillus, thereby affecting the migration and transformation of As and Cd. Moisture conditions further affected bacterial community composition. Co-occurrence network analysis showed that FCBC reduced the complexity and stability of the soil bacterial network, while the anaerobic environment enhanced the synergistic relationships among bacterial species. The functional prediction results of PICRUSt2 revealed that the application of FCBC under aerobic conditions enhanced protein functions related to As and Cd resistance, and promoted key metabolic processes such as amino acid metabolism and organic matter synthesis. Conclusion The synergistic effect of moisture management and Fe-Mg modified coconut shell biochar affects the migration and transformation of As and Cd in paddy soil by regulating the structure and function of microbial communities.[Ch, 8 fig. 3 tab. 38 ref.] -
Key words:
- biochar /
- Fe-Mg modification /
- soil microorganisms /
- redox /
- heavy metals
-
图 1 供试生物炭的形貌和表征
Figure 1 Morphology and characterization of biochar before and after modification
图 2 不同处理下土壤有效态砷镉质量分数
Figure 2 Concentrations of available arsenic and cadmium in soil under different treatments
图 3 不同处理下土壤鲜土指标
Figure 3 Physicochemical properties of fresh soil under different treatments
图 4 不同处理下土壤细菌群落的PCoA分析
Figure 4 PCoA analysis of soil bacterial communities of species numbers under different treatments
图 5 不同处理下土壤细菌群落丰度分布及物种差异分析
Figure 5 Relative abundance of soil bacterial communities and species difference analysis under different treatments.
图 6 环境因子及土壤细菌群落与环境因子的关联分析
Figure 6 Correlation analysis among environmental factors and between soil bacterial communities and environmental factors
图 7 土壤细菌群落共现网络结构特征
Figure 7 Co-occurrence network structure of soil bacterial communities
图 8 PICRUSt2预测的土壤细菌群落功能基因丰度
Figure 8 Functional gene abundance of soil bacterial communities predicted by PICRUSt2
表 1 生物质炭的基本理化性质
Table 1. Selected physicochemical properties of the used biochars
生物炭 碳/
(g·kg−1)氮/
(g·kg−1)氧/
(g·kg−1)氢/
(g·kg−1)比表面积/
(m2·g−1)孔体积/
(cm3·g−1)CBC 744.0 3.0 204.0 34.0 1 017.36 0.46 FCBC 489.0 23.0 272.0 29.0 266.57 0.14  下载: 导出CSV
下载: 导出CSV
表 2 不同处理下稻田土的理化性质
Table 2. Physicochemical properties of paddy soil under different treatments
处理 pH 氧化还原电位/mV 有机质/(g·kg−1) 有效磷/(mg·kg−1) 速效钾/(mg·kg−1) YS-ck 6.05±0.02 c −204.13±4.50 b 28.69±1.01 d 10.69±0.82 c 67.31±4.05 c YS-CBC 6.11±0.08 c −473.45±6.68 c 36.86±1.06 a 12.41±0.50 ab 162.47±15.49 a YS-FCBC 7.38±0.05 a −498.51±8.42 c 34.98±1.08 b 12.01±0.75 b 58.39±6.79 cd SR-ck 5.69±0.09 e 275.65±32.66 a 31.80±1.27 c 11.80±0.99 b 46.87±1.53 d SR-CBC 5.87±0.05 d 310.07±14.81 a 36.83±0.49 a 13.37±0.53 a 93.02±16.32 b SR-FCBC 6.99±0.11 b 235.15±22.93 a 34.71±0.51 b 12.25±0.55 b 51.56±7.27 cd 说明:同列不同小写字母表示不同处理间差异显著(P<0.05)。
下载: 导出CSV
表 3 不同处理下土壤细菌群落多样性指数
Table 3. Diversity indices of soil bacterial communities under different treatments
处理 Ace指数 Chao 1指数 Shannon-Wiener指数 Simpson指数 YS-ck 2 410.5±105.8 a 2 375.1±117.0 a 5.86±0.17 a 0.009±0.002 bc YS-CBC 2 500.7±122.4 a 2 496.3±111.5 a 5.90±0.17 a 0.009±0.002 bc YS-FCBC 2 143.5±175.6 b 2 126.4±172.9 b 5.63±0.04 b 0.011±0.002 b SR-ck 2 417.4±61.9 a 2 402.7±58.3 a 5.96±0.10 a 0.007± 0.001 cSR-CBC 2 558.8±33.2 a 2 530.6±60.6 a 6.05±0.04 a 0.007±0.001 c SR-FCBC 2 113.5±131.7 b 2 079.4±193.0 b 5.47±0.10 b 0.015±0.002 a 说明:同列不同小写字母表示不同处理间差异显著(P<0.05)。
下载: 导出CSV
-
[1] ZHANG Siwen, ZHU Qichao, de VRIES W, et al. Effects of soil amendments on soil acidity and crop yields in acidic soils: a world-wide meta-analysis[J]. Journal of Environmental Management, 2023, 345: 118531. DOI: 10.1016/j.jenvman.2023.118531. [2] HU Xuewu, WANG Jianlei, LÜ Ying, et al. Effects of heavy metals/metalloids and soil properties on microbial communities in farmland in the vicinity of a metals smelter[J]. Frontiers in Microbiology, 2021, 12: 707786. DOI: 10.3389/fmicb.2021.707786. [3] ROY M, GIRI A K, DUTTA S, et al. Integrated phytobial remediation for sustainable management of arsenic in soil and water[J]. Environment International, 2015, 75: 180−198. DOI: 10.1016/j.envint.2014.11.010. [4] ARAO T, KAWASAKI A, BABA K, et al. Effects of water management on cadmium and arsenic accumulation and dimethylarsinic acid concentrations in Japanese rice[J]. Environmental Science & Technology, 2009, 43(24): 9361−9367. DOI: 10.1021/es9022738. [5] YIN Yuepeng, WANG Yurong, DING Changfeng, et al. Impact of iron and sulfur cycling on the bioavailability of cadmium and arsenic in co-contaminated paddy soil[J]. Journal of Hazardous Materials, 2024, 465: 133408. DOI: 10.1016/j.jhazmat.2023.133408. [6] 毛浩楠, 杨兴, 陆扣萍, 等. 铁改性生物质炭对农田土壤养分及砷、铅有效性的影响[J]. 浙江农林大学学报, 2024, 41(6): 1222−1232. MAO Haonan, YANG Xing, LU Kouping, et al. Effect of iron-modified biochars on soil nutrients and bioavailability of As and Pb[J]. Journal of Zhejiang A&F University, 2024, 41(6): 1222−1232. DOI: 10.11833/j.issn.2095-0756.20240171. [7] 梅闯, 蔡昆争, 黎紫珊, 等. 稻秆生物炭对稻田土壤Cd形态转化和微生物群落的影响[J]. 生态环境学报, 2022, 31(2): 380−390. MEI Chuang, CAI Kunzheng, LI Zishan, et al. Effects of rice-straw biochar on the transformation of cadmium fractions and microbial community in paddy soils[J]. Ecology and Environment Sciences, 2022, 31(2): 380−390. DOI: 10.16258/j.cnki.1674-5906.2022.02.019. [8] 孙远, 陈敏, 周育智, 等. 改性生物炭对镉砷复合污染土壤的修复研究进展[J]. 江苏农业科学, 2024, 52(2): 1−11. SUN Yuan, CHEN Min, ZHOU Yuzhi, et al. Research progress of modified biochar for remediation of cadmium-arsenic contaminated soils[J]. Jiangsu Agricultural Sciences, 2024, 52(2): 1−11. DOI: 10.15889/j.issn.1002-1302.2024.02.001. [9] JIAO Yonghong, WANG Tianning, HE Mengchang, et al. Simultaneous stabilization of Sb and As co-contaminated soil by Fe-Mg modified biochar[J]. Science of the Total Environment, 2022, 830: 154831. DOI: 10.1016/j.scitotenv.2022.154831. [10] LI Weitao, WU Meng, LIU Ming, et al. Responses of soil enzyme activities and microbial community composition to moisture regimes in paddy soils under long-term fertilization practices[J]. Pedosphere, 2018, 28(2): 323−331. DOI: 10.1016/S1002-0160(18)60010-4. [11] ZHANG Han, CUI Jianglong, XIONG Ying, et al. Microbial response and the crucial function of predominant phyla in Sedum alfredii-mediated remediation of high concentration of multiple heavy metal soils[J]. Environmental Pollution, 2025, 374: 126211. DOI: 10.1016/j.envpol.2025.126211. [12] XIAO Shiqi, DING Sheng, GAO Wei, et al. Hydroxyapatite self-doped biochar with MgO modification immobilizes heavy metal and alters soil microbial community[J]. Journal of Environmental Chemical Engineering, 2024, 12(6): 114283. DOI: 10.1016/j.jece.2024.114283. [13] ISLAM M S, DENG Hui, DONG Youming, et al. Improving arsenic and cadmium contaminated paddy soil health and rice quality with plant-animal-based modified biochar: a mechanistic study[J]. Journal of Cleaner Production, 2024, 448: 141659. DOI: 10.1016/j.jclepro.2024.141659. [14] ZHANG Ting, SUN Yifei, PARIKH S J, et al. Water-fertilizer regulation drives microorganisms to promote iron, nitrogen and manganese cycling: a solution for arsenic and cadmium pollution in paddy soils[J]. Journal of Hazardous Materials, 2024, 477: 135244. DOI: 10.1016/j.jhazmat.2024.135244. [15] SCHLOSS P D, WESTCOTT S L, RYABIN T, et al. Introducing mothur: open-source, platform-independent, community-supported software for describing and comparing microbial communities[J]. Applied and Environmental Microbiology, 2009, 75(23): 7537−7541. DOI: 10.1128/AEM.01541-09. [16] ZHU Xiaohui, LI Zhexuan, GU Tianyu, et al. Functional materials based on kitchen waste fermentation residue for the remediation of cadmium-contaminated soil[J]. Journal of Environmental Chemical Engineering, 2025, 13(3): 116616. DOI: 10.1016/j.jece.2025.116616. [17] 戴志楠, 杨兴, 陈翰博, 等. 原始及铁改性生物质炭对污染土壤中As、Pb生物有效性和微生物群落结构的影响[J]. 环境科学学报, 2022, 42(7): 456−465. DAI Zhinan, YANG Xing, CHEN Hanbo, et al. Effect of raw and iron-modified biochars on the bioavailability of as and Pb and functional diversity of the microbial community in soils[J]. Acta Scientiae Circumstantiae, 2022, 42(7): 456−465. DOI: 10.13671/j.hjkxxb.2021.0527. [18] ZHANG Yuliang, FU Pingfeng, LI Sheng, et al. Remediation of As, Sb, and Pb co-contaminated mining soils by using Fe/C based solid wastes: synergistic effects and field applications[J]. Chemical Engineering Journal, 2024, 498: 155476. DOI: 10.1016/j.cej.2024.155476. [19] MENSAH A K, MARSCHNER B, SHAHEEN S M, et al. Biochar, compost, iron oxide, manure, and inorganic fertilizer affect bioavailability of arsenic and improve soil quality of an abandoned arsenic-contaminated gold mine spoil[J]. Ecotoxicology and Environmental Safety, 2022, 234: 113358. DOI: 10.1016/j.ecoenv.2022.113358. [20] CHEN Hanbo, GAO Yurong, DONG Huiyun, et al. Chitin and crawfish shell biochar composite decreased heavy metal bioavailability and shifted rhizosphere bacterial community in an arsenic/lead co-contaminated soil[J]. Environment International, 2023, 176: 107989. DOI: 10.1016/j.envint.2023.107989. [21] WU Chuan, AN Wenhui, LIU Ziyu, et al. The effects of biochar as the electron shuttle on the ferrihydrite reduction and related arsenic (As) fate[J]. Journal of Hazardous Materials, 2020, 390: 121391. DOI: 10.1016/j.jhazmat.2019.121391. [22] CHEN Xin, SI Tianren, WANG Senhan, et al. Enhancement of soil DOC and electron transport by biochar strengthens Cd immobilization via iron oxides transformations in waterlogged soils[J]. Journal of Environmental Management, 2025, 391: 126532. DOI: 10.1016/j.jenvman.2025.126532. [23] SI Tianren, CHEN Xin, YUAN Rui, et al. Iron-modified biochars and their aging reduce soil cadmium mobility and inhibit rice cadmium uptake by promoting soil iron redox cycling[J]. Journal of Environmental Management, 2024, 370: 122848. DOI: 10.1016/j.jenvman.2024.122848. [24] WANG Lu, CHEN Hanrui, WU Jizi, et al. Effects of magnetic biochar-microbe composite on Cd remediation and microbial responses in paddy soil[J]. Journal of Hazardous Materials, 2021, 414: 125494. DOI: 10.1016/j.jhazmat.2021.125494. [25] REN Jieling, PENG Yuanzhe, ZHU Aibin, et al. Soil water management-induced transformation of iron phases in kaolinite-loaded amorphous zero-valent iron enhances cadmium immobilization[J]. Journal of Environmental Chemical Engineering, 2025, 13(4): 117282. DOI: 10.1016/j.jece.2025.117282. [26] ZHOU Guowei, YANG Xiaoru, MARSHALL C W, et al. Biochar addition increases the rates of dissimilatory iron reduction and methanogenesis in ferrihydrite enrichments[J]. Frontiers in Microbiology, 2017, 8: 589. DOI: 10.3389/fmicb.2017.00589. [27] ZHANG Yuliang, FU Pingfeng, LI Sheng, et al. Dual regulation of as release and soil environment by Fe(Ⅱ) assisted steel slag and coal fly ash: effects and potential mechanisms[J]. Journal of Hazardous Materials, 2025, 482: 136599. DOI: 10.1016/j.jhazmat.2024.136599. [28] GU Xiang, LI Zhaoshuang, XU Xu, et al. Hydrangea-like biomimetic MgO-modified coconut shell biochar for remediation of multi-media heavy metal pollution: morphological innovation and underlying mechanisms[J]. Journal of Hazardous Materials, 2025, 500: 140337. DOI:10.1016/j.jhazmat.2025.140337. [29] ZHANG Limin, ZENG Qiang, LIU Xi, et al. Iron reduction by diverse Actinobacteria under oxic and pH-neutral conditions and the formation of secondary minerals[J]. Chemical Geology, 2019, 525: 390−399. DOI: 10.1016/j.chemgeo.2019.07.038. [30] ZHANG Dan, GONG Yipeng, QIN Rui, et al. Effect and mechanism of rice-Solanum nigrum L. rotation system inoculated with sulfate-reducing bacteria (SRB) and sulfur-oxidizing bacteria (SOB) for the remediation of Cd-contaminated paddy soil[J]. Journal of Hazardous Materials, 2025, 496: 139540. DOI: 10.1016/j.jhazmat.2025.139540. [31] ZHANG Guogang, LIU Xuewei, GAO Minling, et al. Effect of Fe-Mn-Ce modified biochar composite on microbial diversity and properties of arsenic-contaminated paddy soils[J]. Chemosphere, 2020, 250: 126249. DOI: 10.1016/j.chemosphere.2020.126249. [32] TENG Zedong, ZHAO Xin, YUAN Junjun, et al. Phosphate functionalized iron based nanomaterials coupled with phosphate solubilizing bacteria as an efficient remediation system to enhance lead passivation in soil[J]. Journal of Hazardous Materials, 2021, 419: 126433. DOI: 10.1016/j.jhazmat.2021.126433. [33] YANG Peng, WU Bin, ZHENG Shuai, et al. Effective cadmium immobilization in paddy soil by the interaction of sulfate reducing bacteria and manganese fertilizer[J]. Journal of Environmental Management, 2024, 371: 123261. DOI: 10.1016/j.jenvman.2024.123261. [34] LI Linfeng, LI Qi, XIAO Anwen, et al. Regulation of soil properties by amendments and their impact on Cd fractions and bacterial community structure: exploring the mechanism of inhibition on Cd phytoavailability[J]. Ecotoxicology and Environmental Safety, 2025, 294: 118033. DOI: 10.1016/j.ecoenv.2025.118033. [35] TIAN Shengni, DONG Yufei, PANG Shouyang, et al. Driving role of acid mine drainage on microbial community assembly and species coexistence in paddy soil profiles[J]. Journal of Environmental Sciences, 2025, 156: 771−783. DOI: 10.1016/j.jes.2024.12.034. [36] TANG Tao, ZENG Peng, WANG Jiangyao, et al. Remediation of multi-metal contaminated soil using cow dung and its biochar: effects on heavy metal uptake and soil microbiome in Triticum aestivum L.[J]. Environmental Technology & Innovation, 2025, 39: 104243. DOI: 10.1016/j.eti.2025.104243. [37] YU Fangming, HE Ziang, XIN Xiaomin, et al. Evidence that beneficial microbial inoculation enhances heavy metal-contaminated soil remediation: variations in plant endophyte communities[J]. Journal of Hazardous Materials, 2024, 480: 135883. DOI: 10.1016/j.jhazmat.2024.135883. [38] WANG Xu, LI Daping, GAO Ping, et al. Analysis of biosorption and biotransformation mechanism of Pseudomonas chengduensis strain MBR under Cd(Ⅱ) stress from genomic perspective[J]. Ecotoxicology and Environmental Safety, 2020, 198: 110655. DOI: 10.1016/j.ecoenv.2020.110655. -

-
链接本文:
https://zlxb.zafu.edu.cn/article/doi/10.11833/j.issn.2095-0756.20260146
 点击查看大图
点击查看大图
计量
- 文章访问数: 218
- HTML全文浏览量: 68
- PDF下载量: 43
- 被引次数: 0
