





-
城市公园是位于城市范围之内经专门规划建设的绿地,是自然生态的环境空间,具有向公众开放、以游憩为主的功能,兼具提高城市生态质量、美化城市环境、城市防灾减灾等作用[1-3]。城市公园的建设对提高城市生活环境质量与人们环境幸福感有着重要意义,城市公园的服务能力是衡量城市整体环境水平和居民生活品质的一项重要指标[4-5]。目前一些学者正在从各个不同的角度对城市公园服务能力进行评价分析。梁颢严等[6]、李莹等[7]采用缓冲区分析方法,黄曦涛等[8]、翟雪等[9]采用网络分析的方法对公园的服务能力进行了评价。陈永生等[10]以遥感(RS)和地理信息系统(GIS)技术为支撑,采用层次分析法和德尔菲法对合肥市中心城区公园绿地的功能进行了综合评价。BANCROF等[11]对美国的公园绿地空间分布与可达性进行了研究,从可达性角度对公园绿地公平性及效率进行评价,可达性能够较好地研究评价城市公园的空间分布,是评价城市公园空间分布合理性和服务公平性的主要手段[12-13]。李维宇等[14]采用多中心模型,利用中间性和邻近度指标分析了武汉市中心城区道路网的中心性,分析公园绿地的可达性,以可达性指数测度城市公园服务能力。当前相关研究表明:服务能力是满足人们对城市公园需求的重要体现。福州市是自然灾害活动较为活跃的区域之一[15],受地形因素影响,发展空间有限,在一定程度上限制了城市公园的发展。从城市公园现状服务能力与潜在城市公园需求出发,探讨福州市城市公园的布局现状与优化策略,可为提高福州市城市公园的服务能力提供科学依据。
-
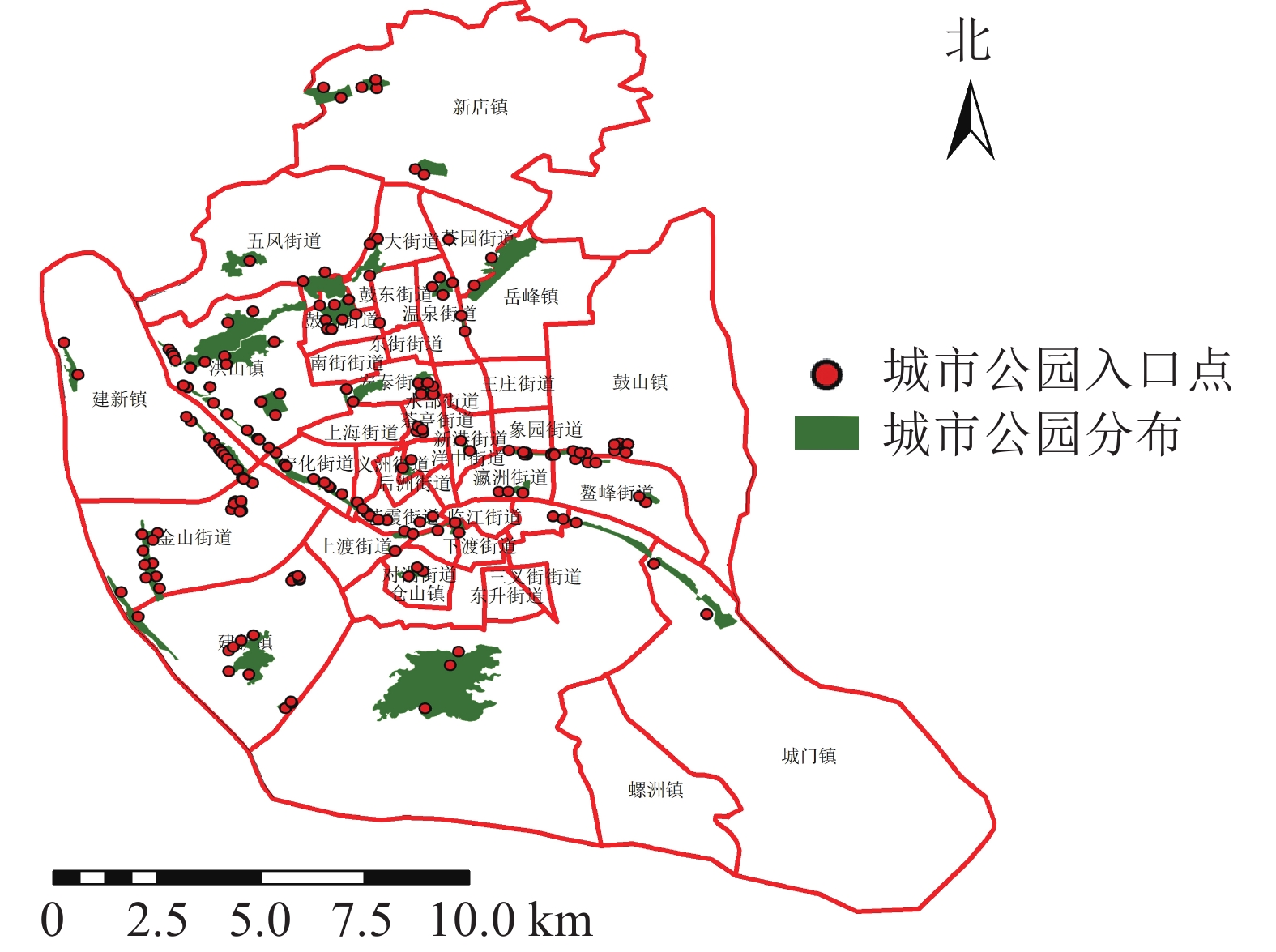
福州市是福建省的省会,是中国东南沿海城市之一。福州市地貌形态较为复杂,四周被群山环绕,位于河口盆地中心。本研究以福州市主城区为研究对象,包含鼓楼区、仓山区、台江区、晋安区(建成区部分) 4个区,面积约248.3 km2,共有30个街道9个镇,其中,鼓楼区9个街道1个镇,台江区10个街道,仓山区8个街道5个镇,晋安区3个街道3个镇。利用谷歌地球(Google Earth)及遥感影像目视解译共提取47个城市公园(图1),其中鼓楼区17个,台江区11个,仓山区13个,晋安区6个,面积共11.11 km2。
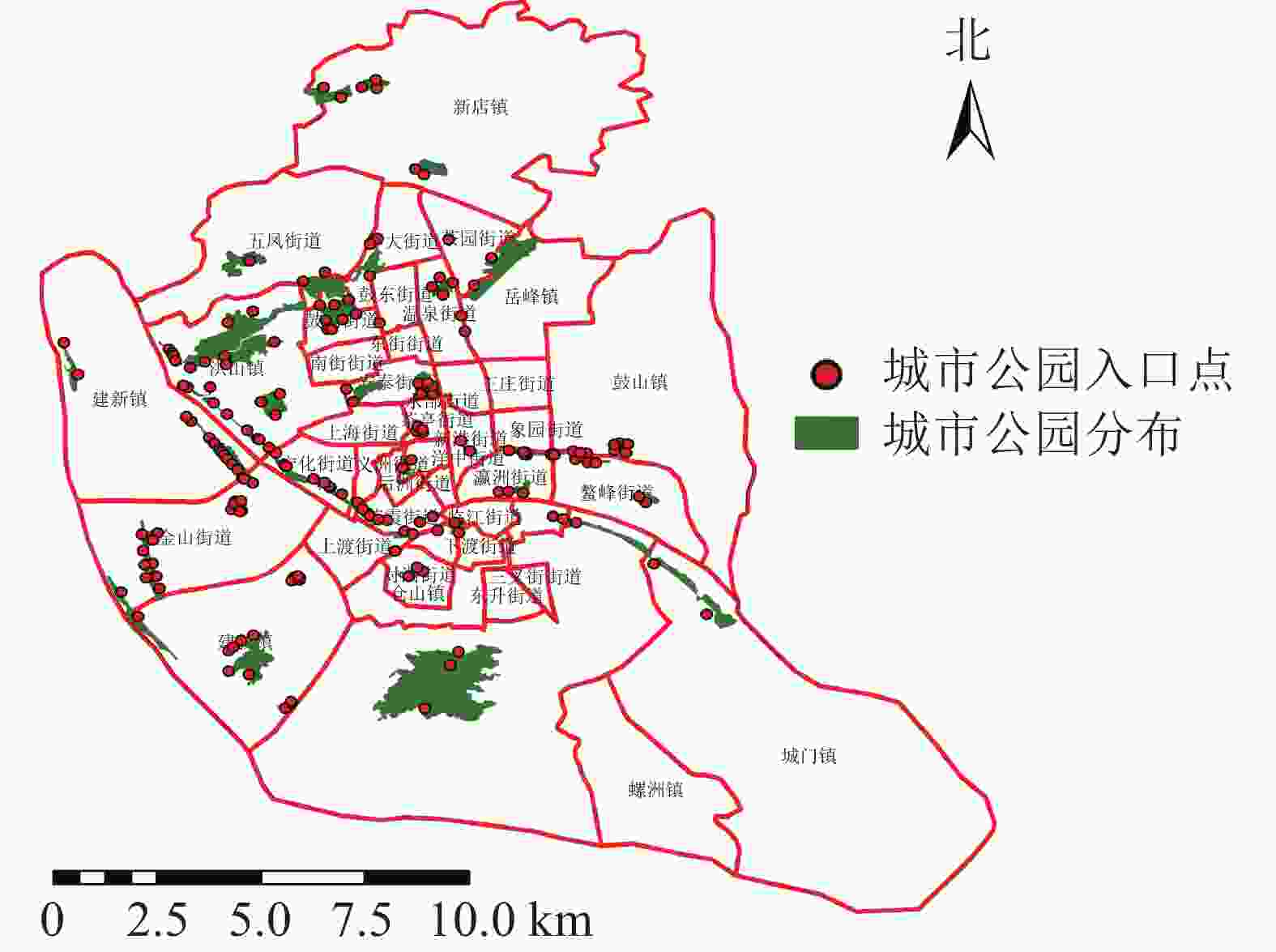
图 1 研究区城市公园及其入口点分布示意图
Figure 1. Distribution of urban parks and their entry points in the study area
-
以福州市主城区为研究区域,绿地、人口等相关数据来源于福州市园林局网(http://ylj.fuzhou.gov.cn/)、《福州年鉴》,遥感影像为地理空间数据云(http://www.gscloud.cn/) 2016年7月27日的Landsat 8卫星遥感影像,影像平均云量4.77,研究区基本无云,有利于地物判读。福州市行政区划分、街道分区图来源于福州市城乡规划局网,以及百度地图与上海脉策数据科技有限公司采集与发布的福州市兴趣点(point of interest,POI)等数据。
-
根据城市公园的入口分析服务域的覆盖率,以研究区的街道作为基本空间单元,根据不同类型城市公园避灾服务到达时间与速度,得到城市公园避灾的服务域面积,服务域覆盖率计算公式如下:Fi=(S1−S2)/(3S)。其中:Fi为某一避灾区i所有城市公园避灾服务域覆盖率,S1为所有避灾类型城市公园避灾服务域的面积(km2),S2为街道边界外避灾服务域面积(km2),S为街道的面积(km2)。
-
利用GIS对服务压力进行可视化及空间化处理,将现有城市公园转换为泰森多边形,生成各城市公园的服务范围。利用GIS对城市公园服务压力进行可视化提取,计算服务压力[16]:U=N/A。其中:U为服务压力(个·km−2),A为城市公园面积(km2),N为城市公园服务范围内POI的数量(个)。
-
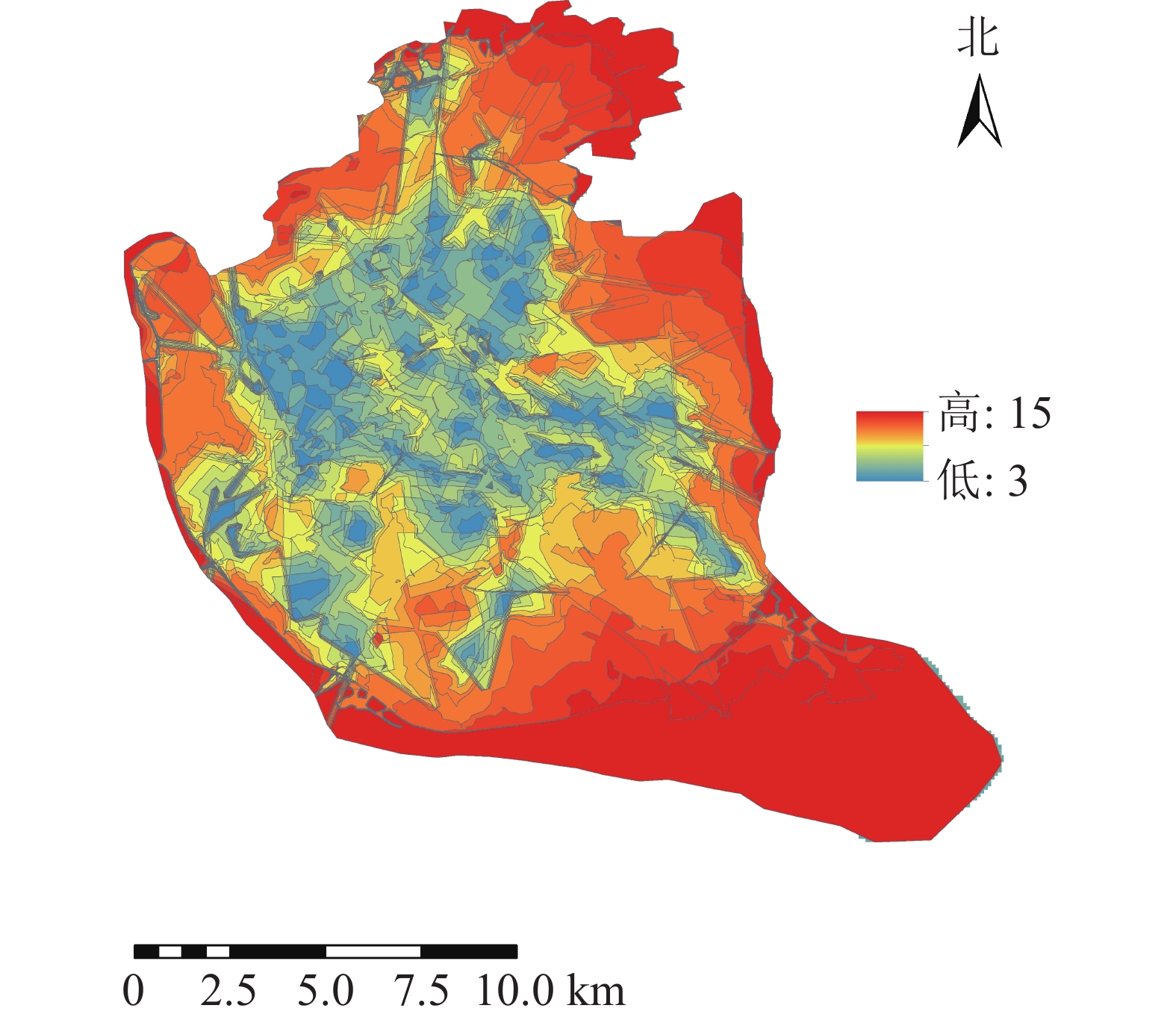
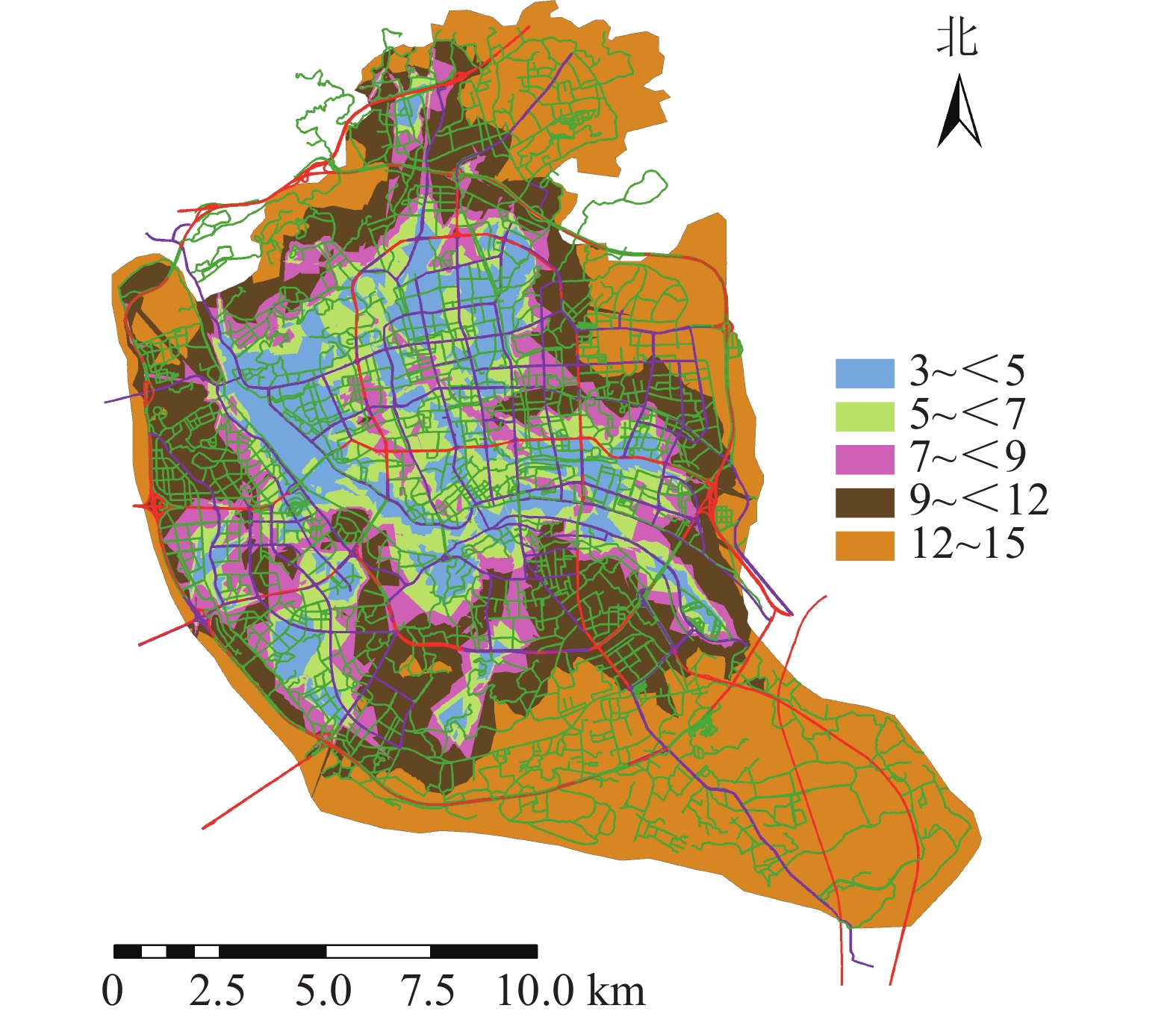
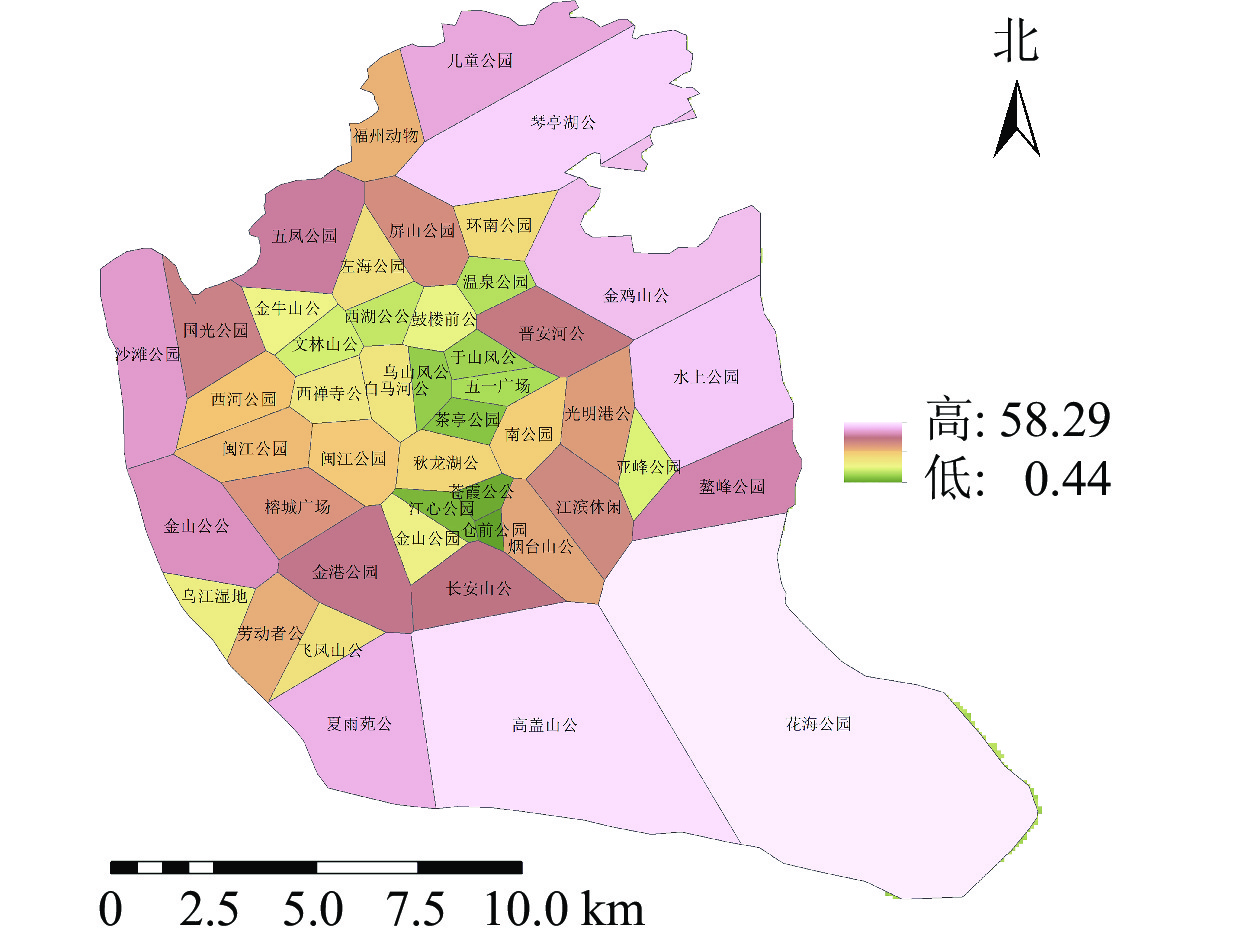
将道路数据库数据整合为Geodatabase网络数据集,利用网络分析模块(network analyst),对不同交通方式的不同时间段的可达面积进行赋值。相关研究表明人们更愿意选择步行到达5~10 min路程,车行10~20 min路程的公园[17]。因此选择以步行5 min,机动车10 min为间隔对不同交通方式下的可达性进行赋值,其中步行0~5 min能到达的区域为1分,大于20 min才能到达的区域为5分;机动车与公交车0~10 min可以到达的地区为1分,大于40 min能到达的为5分。利用GIS的矢量叠加功能对3种交通方式叠加分析并得到综合可达性结果(图2)。为直观分析游憩可达性,对综合可达性进行分级,通过咨询相关专家,结合福州市实际情况,对游憩综合可达性根据不同交通方式下可达性综合得分情况利用自然间断点分级法(Natural Breaks)进行等级划分。综合可达性情况分为5个等级:一级,12~15;二级,9~<12;三级,7~<9;四级,5~<7;五级,3~<5。对应差、较差、一般、较好、好5种情况(图3)。分值越低,等级越高,游憩可达性越好。
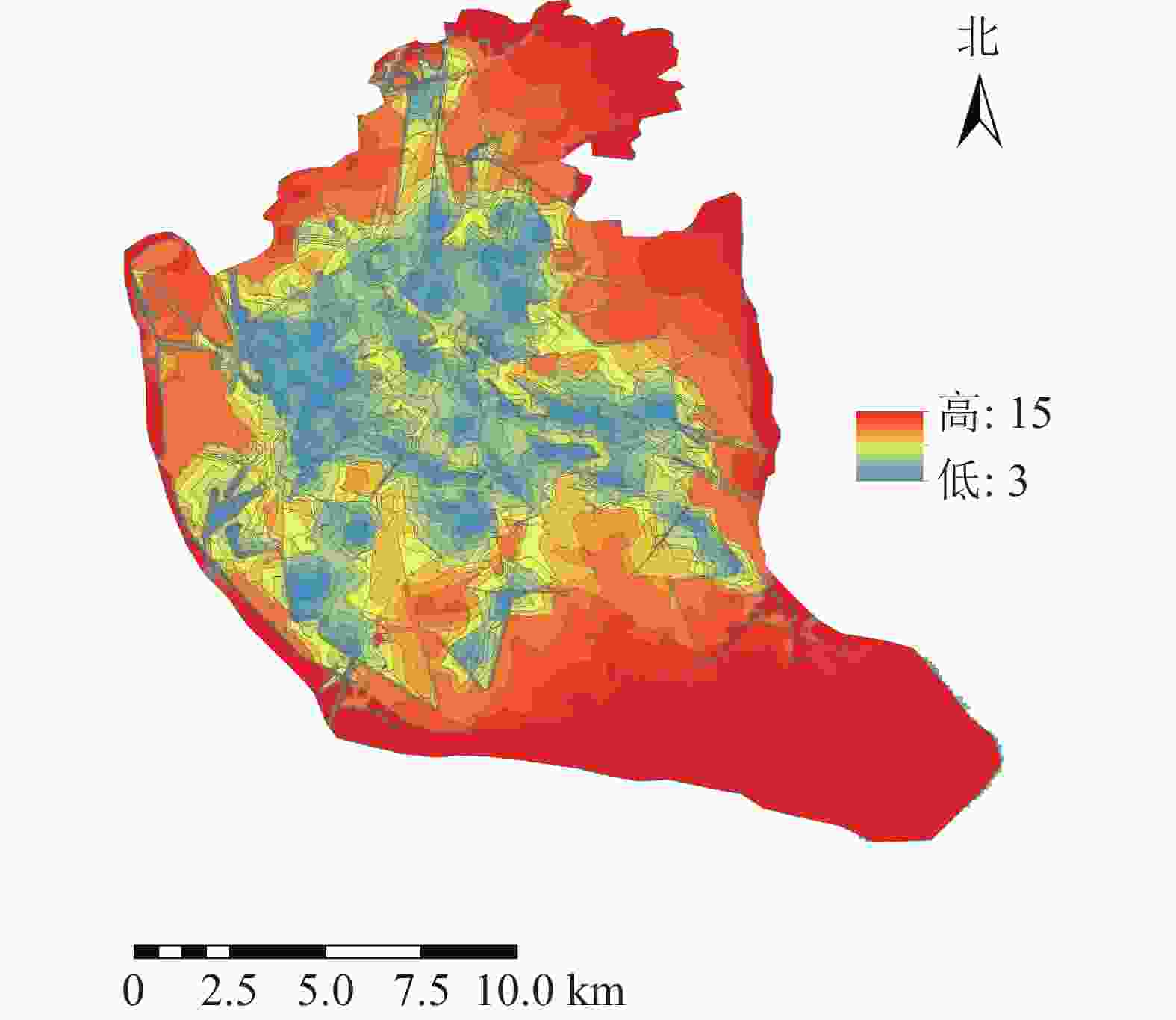
图 2 研究区综合可达性示意图
Figure 2. Comprehensive accessibility of the study area
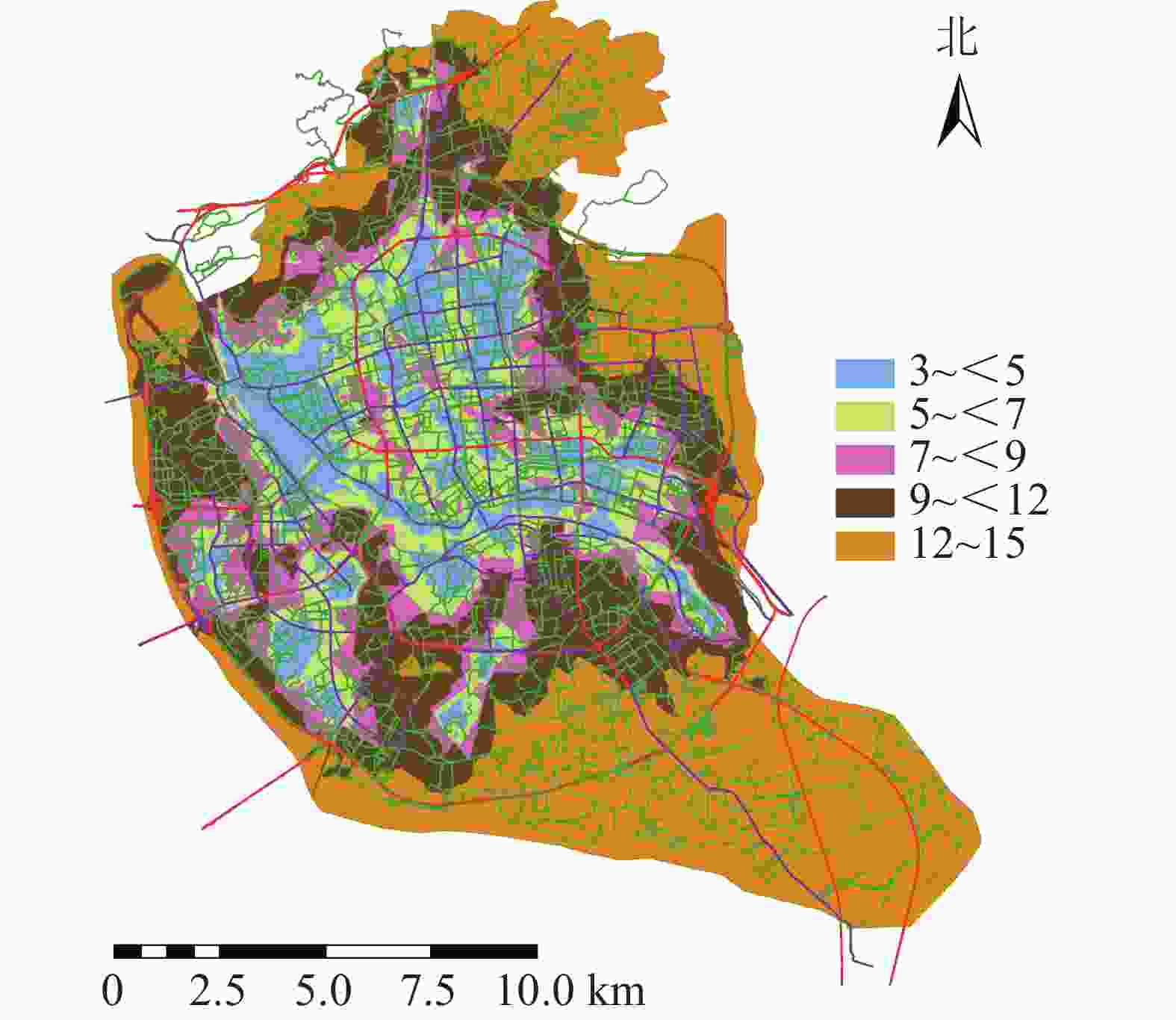
图 3 综合可达性等级分布示意图
Figure 3. Grade of the accessibility overall evaluation
-
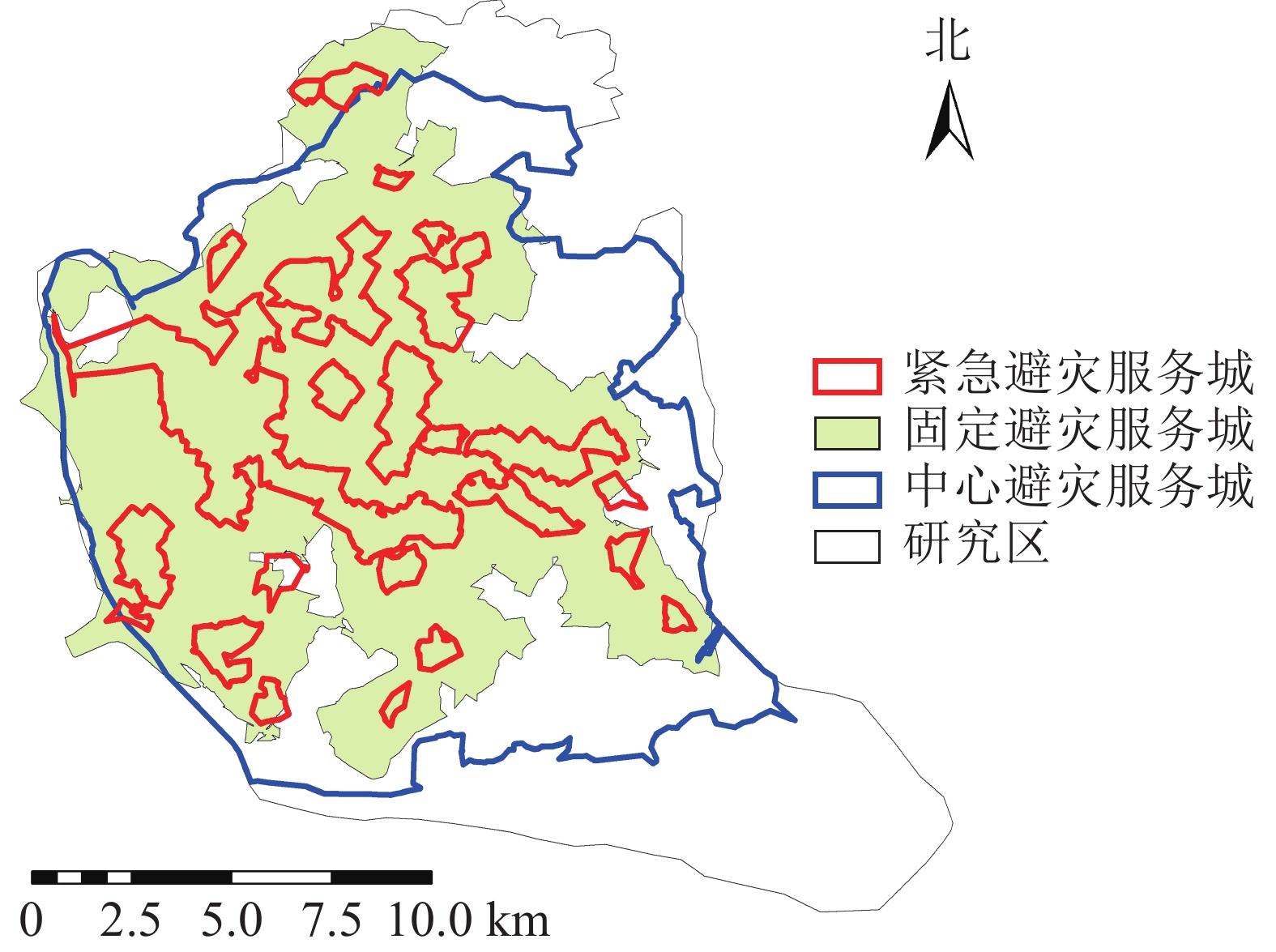
参照国内外学者对避灾绿地的划分标准[18-19],将福州市城市公园避灾类型划分为3类:中心避灾(>0.5 km2)5个、固定避灾(0.1~0.5 km2)19个、紧急避灾(<0.1 km2)23个。参照日本地震经验及国内相关避灾经验[20-21],确定避灾城市公园服务范围(图4)。通过计算可得城市公园的避灾覆盖率分布情况(图5)。相关研究表明城市公园在防灾避险、紧急避灾、疏散居民方面具有重要作用[22],城市避灾服务受重视程度越来越高。为区分福州主城区公园避灾服务分布情况,研究根据等间距法将避灾覆盖率划分为5个等级:一级,<0.2;二级,0.2~<0.4;三级,0.4~<0.6;四级,0.6~<0.8;五级,≥0.8。等级越高避灾覆盖率越大。
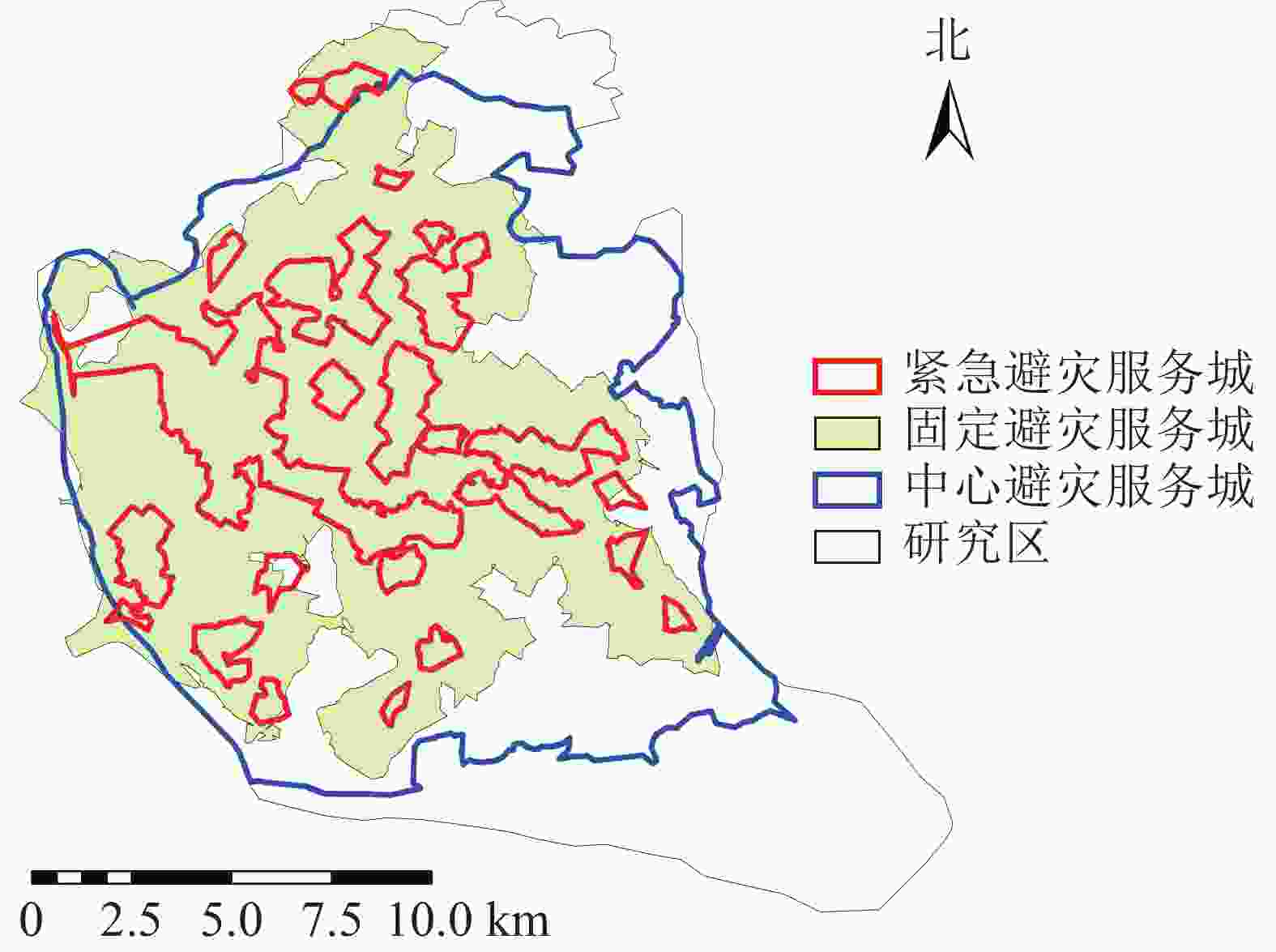
图 4 城市公园不同避灾类型服务域示意图
Figure 4. Service areas of different types of disaster prevention in city parks
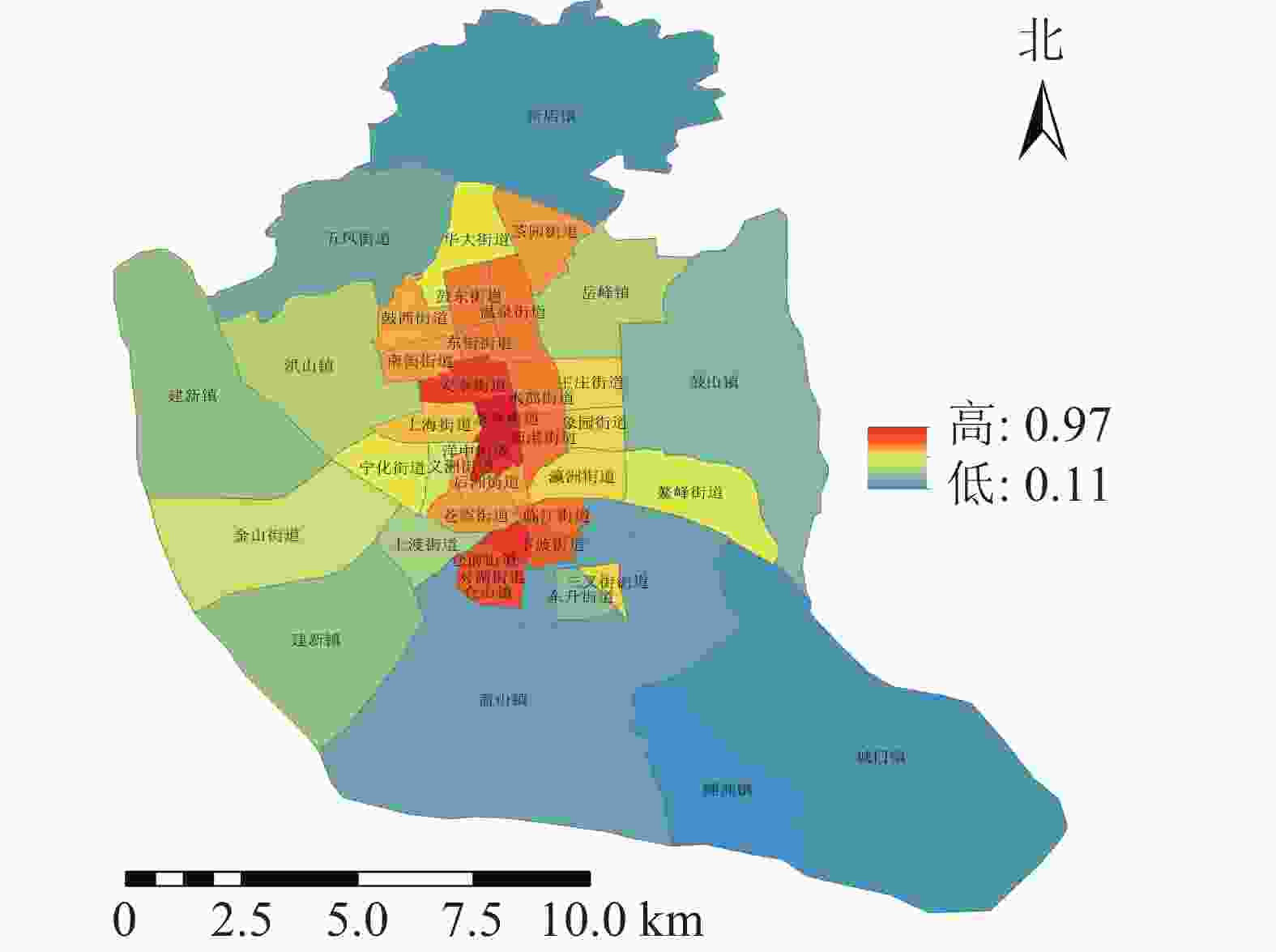
图 5 不同街道城市公园避灾服务域覆盖率示意图
Figure 5. Coverage rate of disaster prevention service areas in urban parks in different streets
-
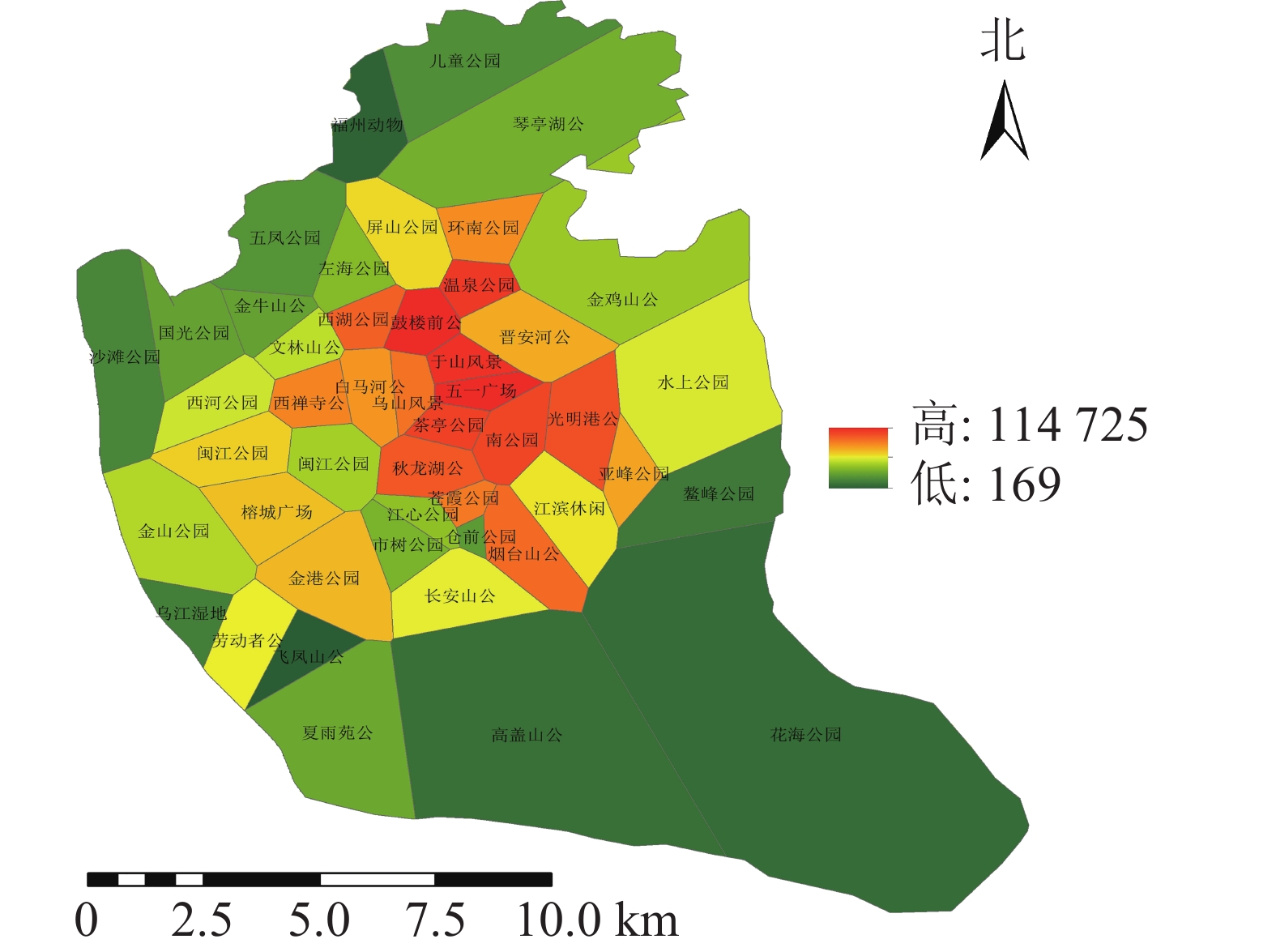
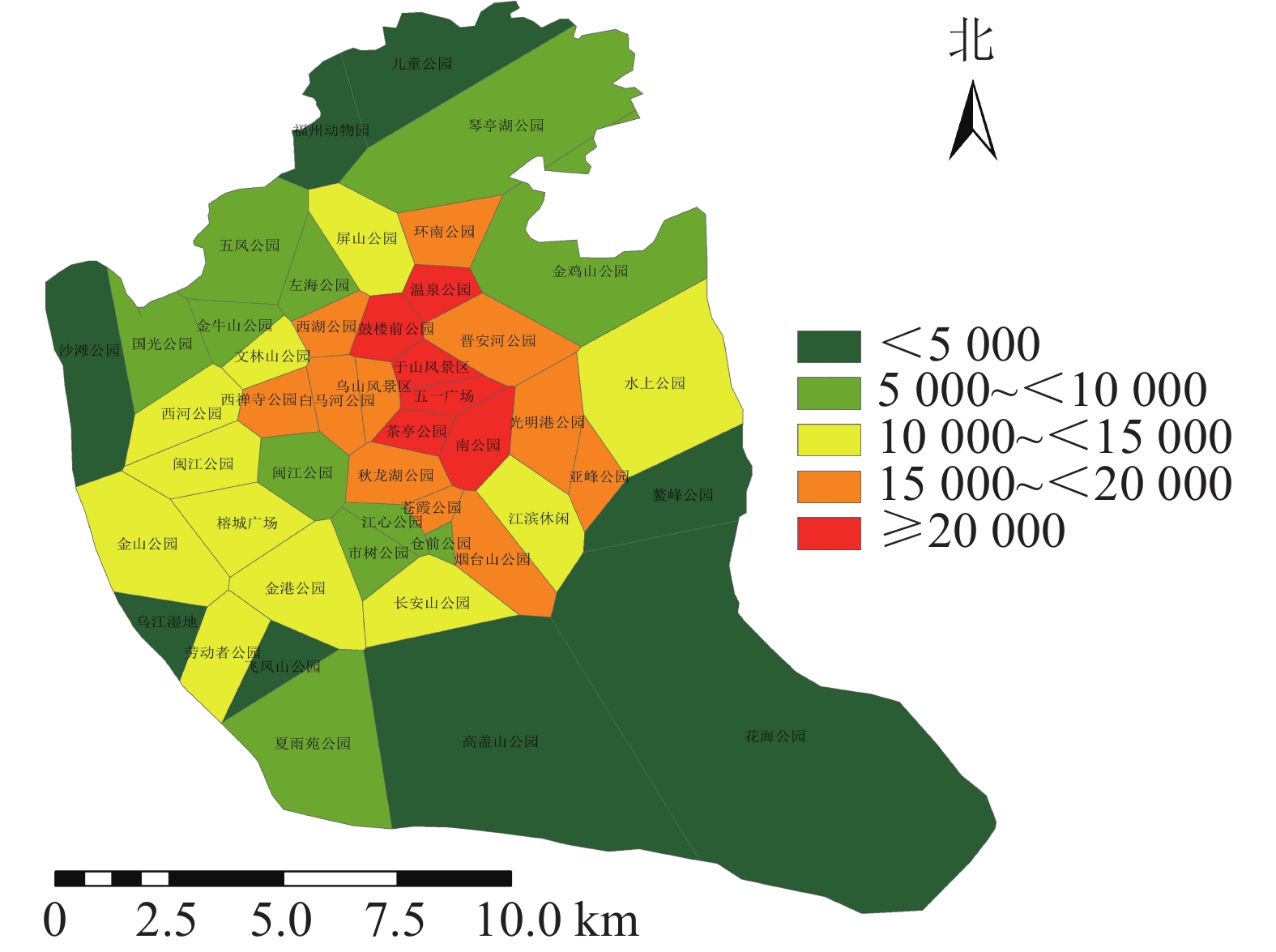
利用百度地图等开源开发者平台可得到完整准确的城市POI数据。选取与人们生活及活动相关的POI数据,根据衣食住行选取8种类型数据,利用GIS的统计提取功能得到POI数量:居住区6 395个,购物区680个,餐饮区13 121个,公司企业16 216个,交通设施6 724个,生活服务7 088个,体育休闲服务区2 379个,科教文化服务区6 192个,共计58 795个。根据POI数据的分布,确定研究区不同街道的POI数据。基于研究区域内所有POI点覆盖情况,利用泰森多边形法建立城市公园的服务范围(图6)。根据计算公式得出服务压力(图7),根据相关文献对服务压力分级研究,并对服务压力进行分级处理(图8)。服务压力以人们在城市公园服务范围内的人群活动强度进行量化分析。研究表明人类活动强度大的区域,居民人口密度不一定高,但对城市公园需求程度较高[16]。结合福州市人口密度情况,咨询相关专家,将福州市主城区城市公园服务压力依次划分为5个等级:一级,<5 000;二级,5 000~<10 000;三级,10 000~<15 000;四级,15 000~<20 000;五级,≥20 000。等级越高服务压力越大。
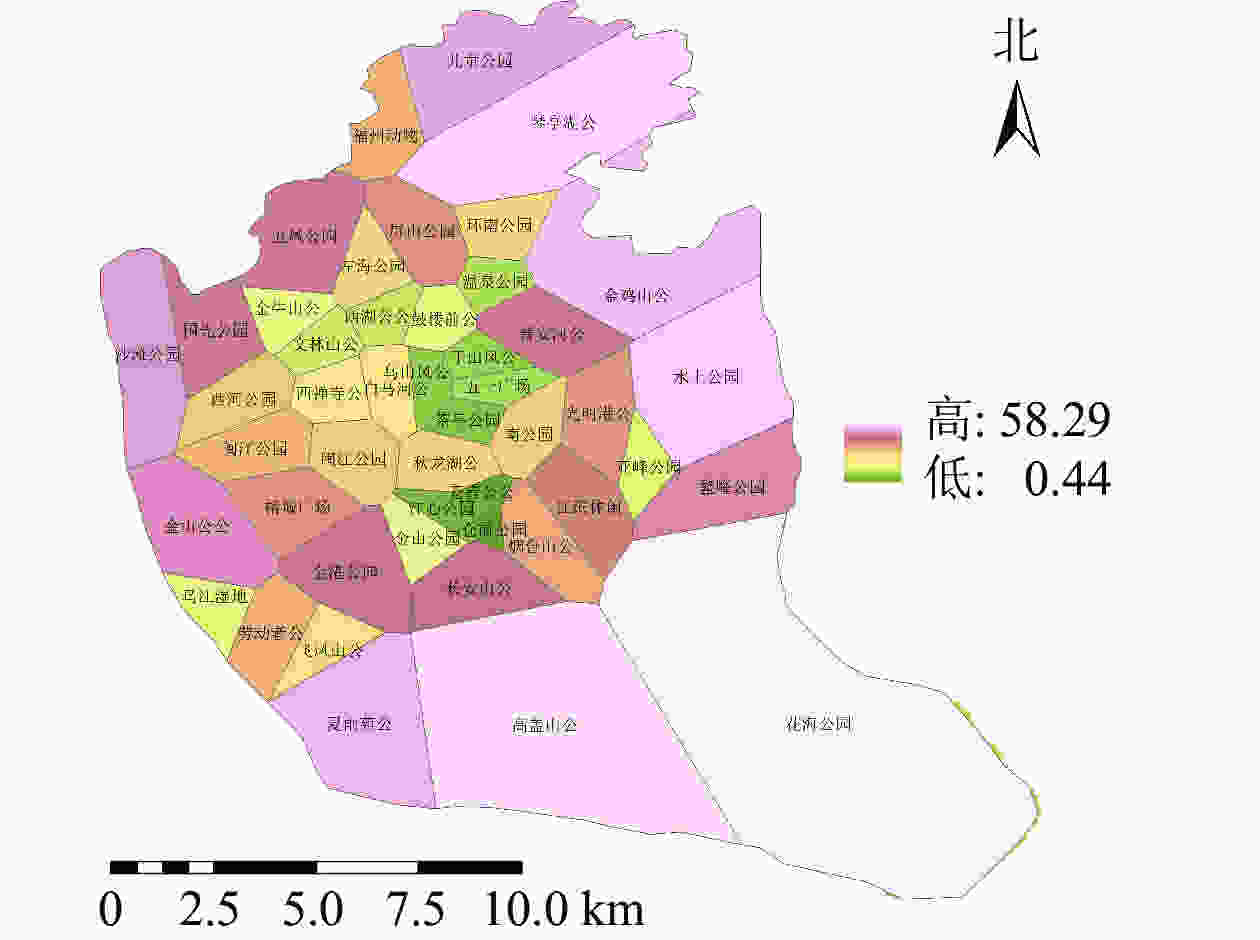
图 6 研究区城市公园服务范围示意图
Figure 6. Scope of service of urban parks in the study area
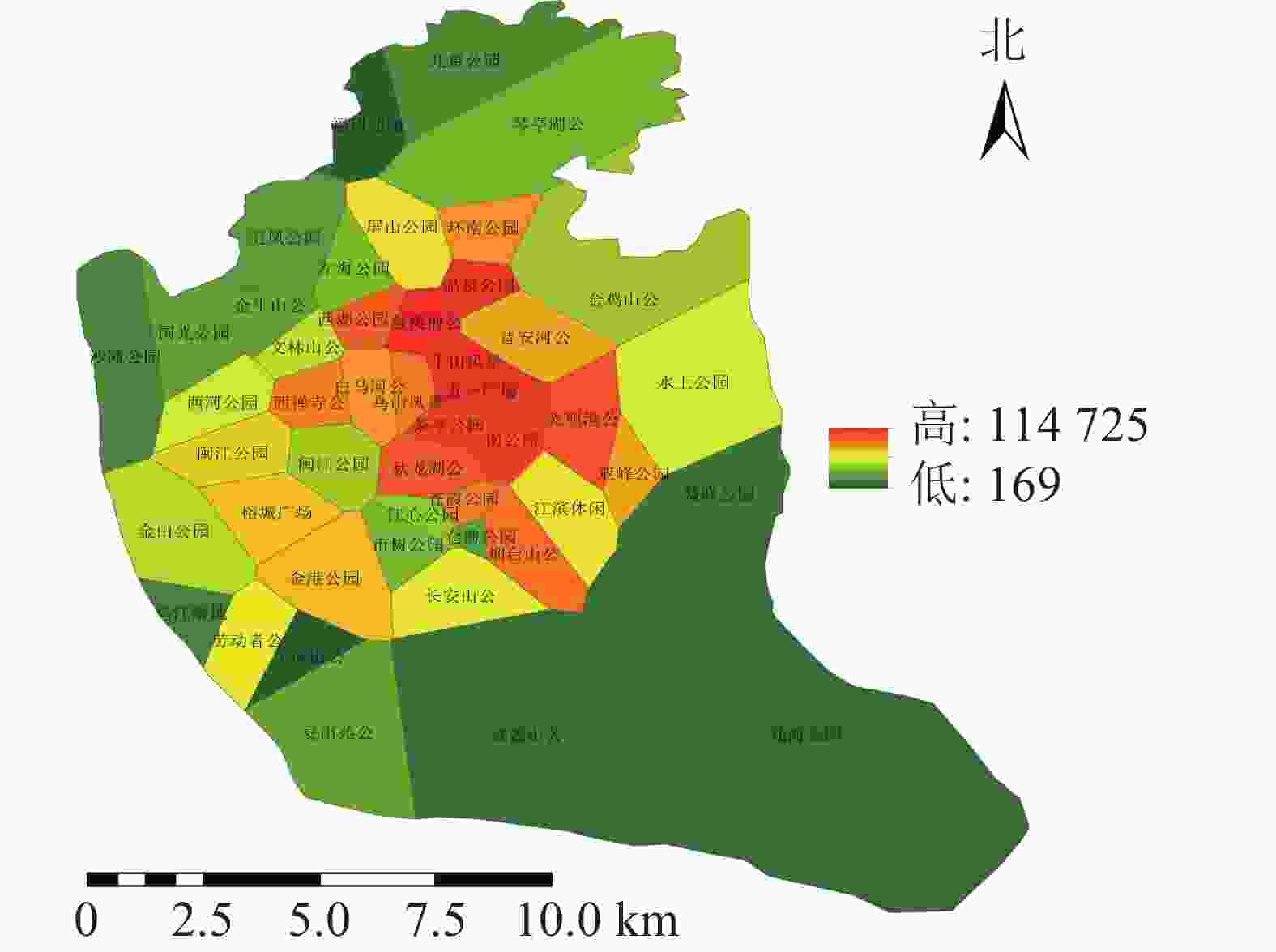
图 7 研究区城市公园服务压力分布示意图
Figure 7. Service pressure distribution of urban parks in the study area
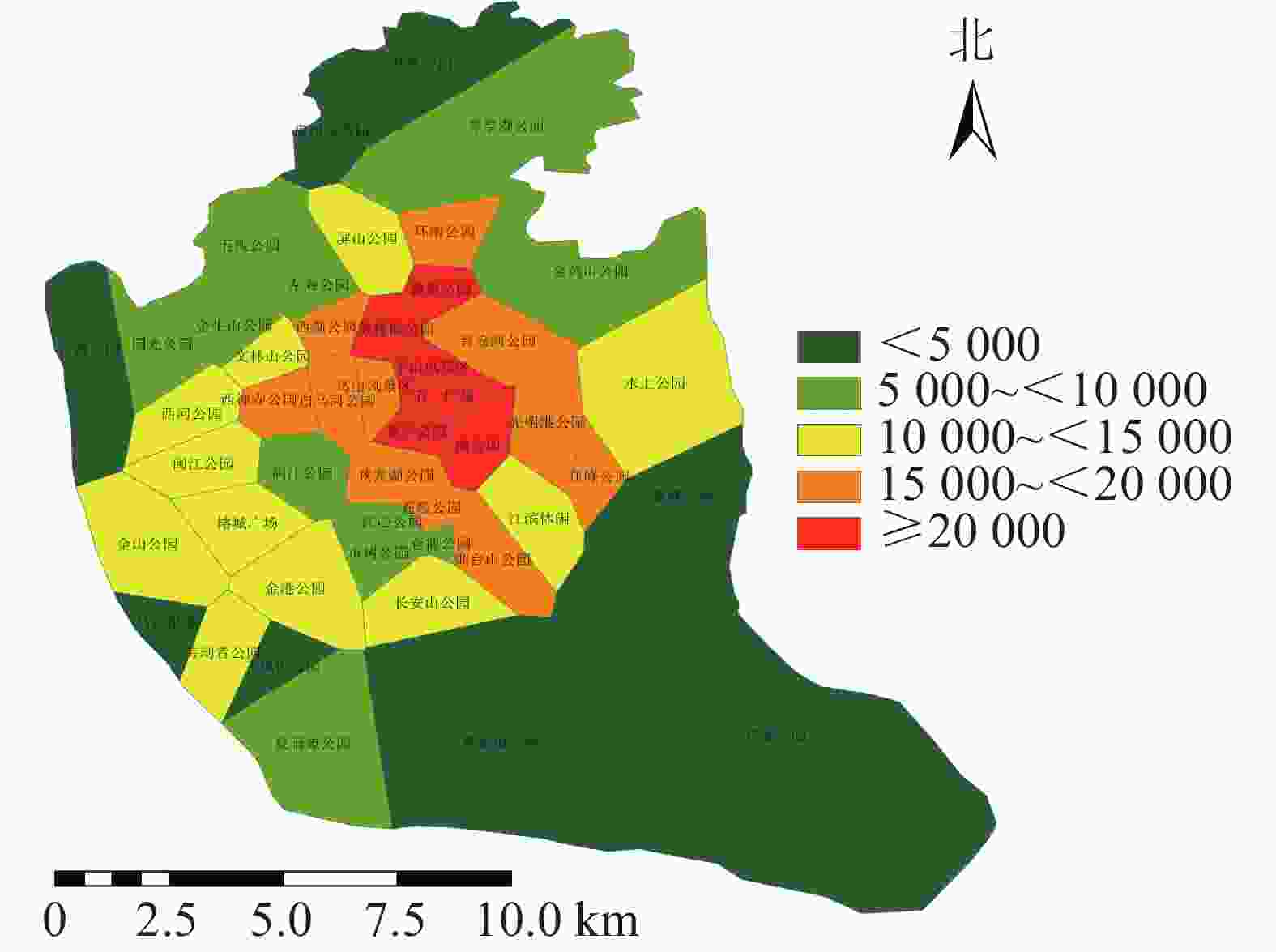
图 8 研究区城市公园服务压力等级分布示意图
Figure 8. Distribution of service stress levels in urban parks
-
福州市城市公园游憩可达性呈中心向周围扩散的现象,中心城区的综合可达性较好,并逐渐向四周扩散。由图3可知:福州市主城区城市公园可达性情况分布不均。由表1可知:城市公园可达性差的面积占福州市主城区面积的38.43%;可达性较差的面积占福州市主城区面积的23.98%;城市公园可达性一般的面积占福州市主城区面积的11.86%;城市公园可达性较好的面积占福州市主城区面积的12.81%;城市公园可达性好的面积占州市主城区面积的12.92%;位于中心城区的鼓楼区与台江区的城市公园可达性较好,因为这些区域城市公园较多,且大多为商业区及生活区,路网密度高,但其对日常维护的要求也相对较高。其他区域可达性较差,这些区域地处边缘,城市公园较少,路网密度小。福州市主城区城市公园游憩可达性很差以及差的面积比达到62.41%,游憩可达性总体较差。
表 1 不同等级游憩可达性面积
Table 1. Area of different grades of recreational accessibility
等级 面积/ km2 比例/% 一级 95.42 38.43 二级 59.54 23.98 三级 29.45 11.86 四级 31.81 12.81 五级 32.08 12.92 -
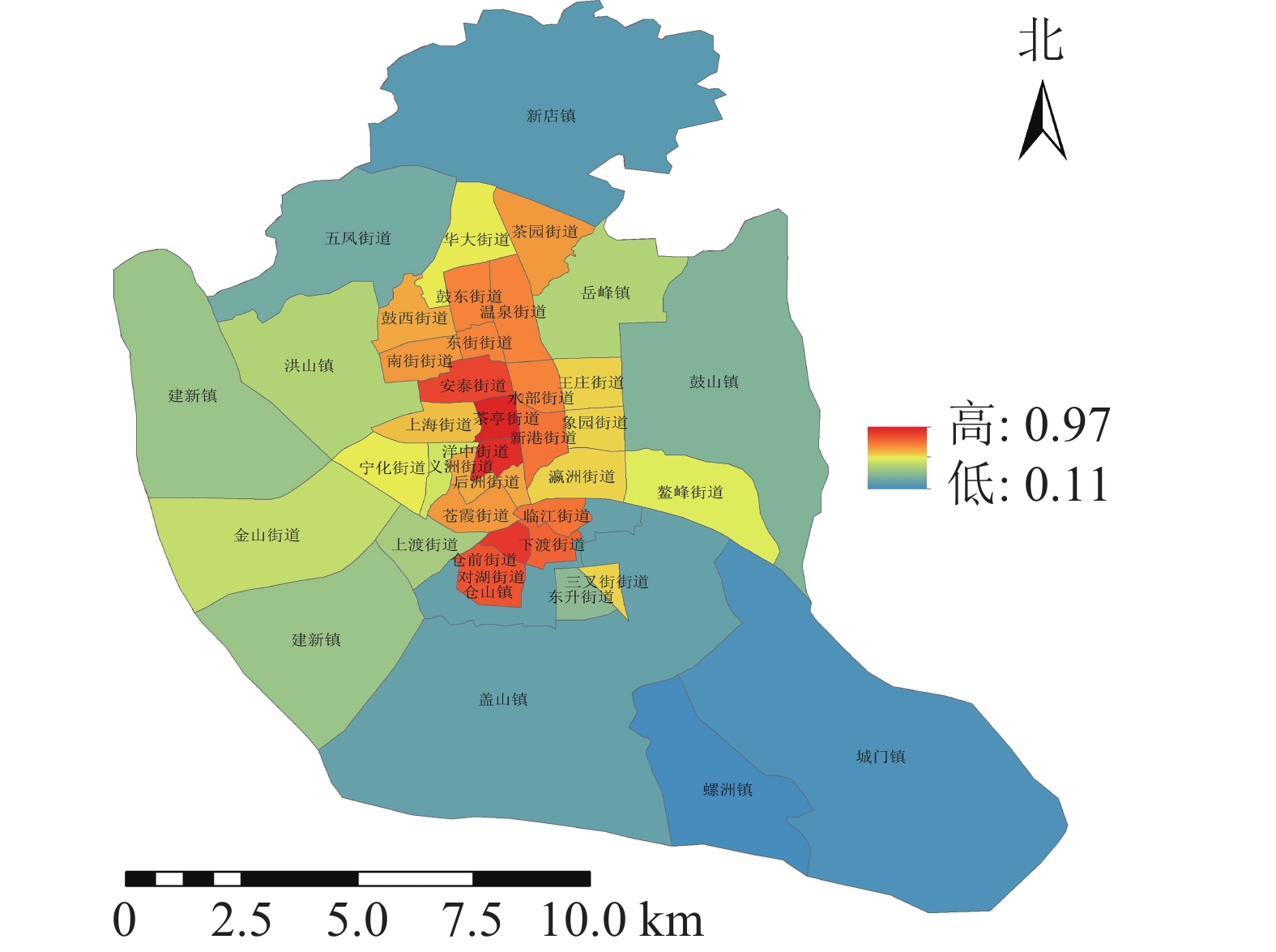
不同类型的避灾服务域结果分析(图4):研究区中心避灾服务情况最好,服务域面积为187.11 km2,服务域覆盖率为75.89%;固定避灾服务域面积为141.01 km2,服务域覆盖率为56.79%,紧急避灾城市公园服务域为40.1 km2,服务域覆盖率为16.15%。根据不同街道城市公园避灾服务情况,利用避灾服务域覆盖率计算公式可得不同街道避灾服务域覆盖率。由表2可知:城市公园避灾覆盖率的分布规律为由中心向四周扩散,城市公园避灾覆盖率最高达97%,最低的为11%。避灾覆盖率大的区域主要分布于台江区的茶亭街道与洋中街道,仓山区的城门镇与螺洲镇的城市公园避灾覆盖率较小。
表 2 不同等级避灾覆盖率街道分布
Table 2. Street distribution of different levels of disaster avoidance coverage
等级 数量/个 街道 一级 3 新店镇、城门镇、螺洲镇 二级 7 鼓山镇、五凤街道、建新镇、上渡街道、仓山镇、盖山镇、东升街道 三级 11 洪山镇、华大街道、岳峰镇、王庄街道、象园街道、瀛洲街道、义洲街道、宁化街道、金山街道、鳌峰街道、三叉街道 四级 16 茶园街道、温泉街道、水部街道、新港街道、鼓东街道、东街街道、鼓西街道、南街街道、安泰街、上海街道、后洲街道、苍霞街道、临江街道、仓前街道、下渡街道、对湖街道 五级 2 茶亭街道、洋中街道 -
对游憩可达性与避灾覆盖率综合分析可知:福州市主城区城市公园综合服务能力为由中心城区向四周扩散;中心城区的路网较丰富,避灾覆盖率大,综合服务能力较好,位于福州市主城区的台江区与鼓楼区服务能力较好,仓山区与晋安区的服务能力较差。由于城市发展差异性与土地利用不同,福州市主城区城市公园的布局存在因服务能力的差异较大产生的布局不合理等问题。
-
利用城市公园服务范围与POI计算得出城市公园的服务压力(图7),服务压力最小的是仓山区的飞凤山公园,最大的是鼓楼区的鼓楼前公园。各个公园的服务压力高低差距较大,且服务压力大的公园较为集中。城市公园服务压力等级分布(图8)可知:服务压力等级较大的主要分布于鼓楼区、台江区及晋安区的交界处。主要原因是这些区域属于老城区范围,人们活动强度比较大。由表3可知:服务压力五级的城市公园有5个,服务压力三级及服务压力四级的城市公园各有11个,服务压力二级的城市公园有12个,服务压力一级的城市公园有8个。
表 3 不同等级服务压力的公园分布
Table 3. Park distribution of different grades of service pressure
等级 数量/个 街道 一级 8 飞凤山公园、福州动物园、花海公园、高盖山公园、鳌峰公园、乌江湿地公园、沙滩公园、儿童公园 二级 12 五凤公园、仓前公园、国光公园、金牛山公园、夏雨苑公园、琴亭湖公园、市树公园、左海公园、江心公园、金鸡山公园、闽江公园(北园)、金山公园 三级 11 金山公园、文林山公园、西河公园、水上公园、长安山公园、劳动者公园、江滨休闲广场、闽江公园(南园)、榕成广场、金港公园、屏山公园 四级 11 晋安河公园、亚峰公园、白马河公园、环南公园、西禅寺公园、苍霞公园、乌山风景区、烟台山公园、西湖公园、秋龙湖公园、光明港公园 五级 5 南公园、鼓楼前公园、茶亭公园、温泉公园、于山风景区 根据服务压力的分布可知:中心城区的服务压力较大。分析可知,这些区域内的人口活动密度强,对城市公园的需求也越大。通过对服务压力的分析发现:台江区对城市公园的需求最大,仓山区的城市公园的需求最小。与福州市主城区现有城市公园服务能力相比较,区域服务能力较好的区域服务压力也相应增加,这些现象表明福州主城区城市公园的布局需要进一步优化。
-
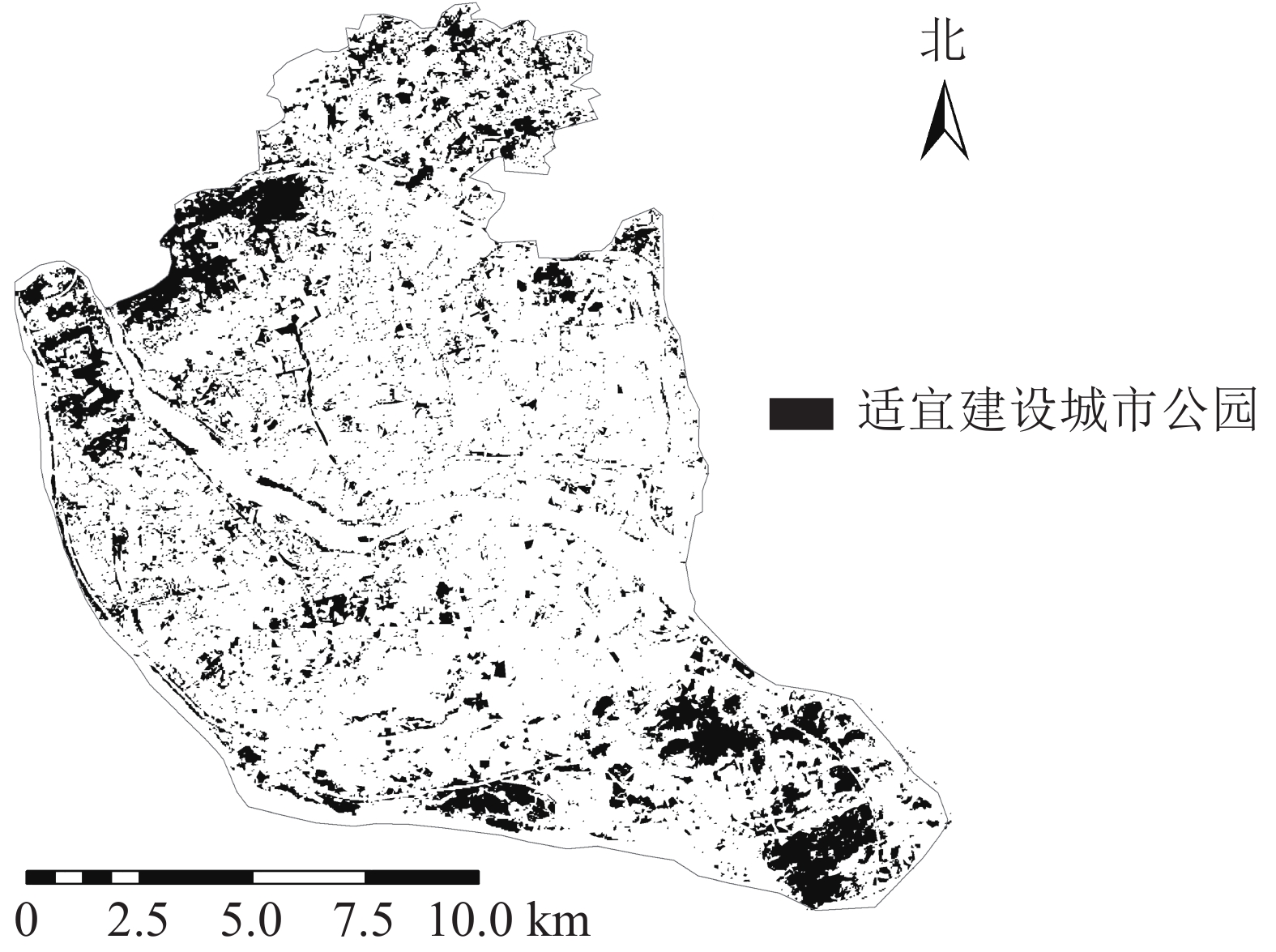
由于公园绿地布局合理性很差的区域呈聚集态分布,因此以福州市主城区公园绿地布局合理性等级为很差的区域为主要研究对象。根据服务能力的分布情况,仓山区与晋安区服务能力差,城市公园数量与需求不符合。根据土地利用分类结果对其进行灰度化处理,将图像划分成256个等级的灰度图像,在全黑0与全白255之间插有254个灰度等级颜色,再对灰度图像进行自适应阈值的二值化处理,使其像素值仅为0或255,灰度级为2。其中适宜建设公园绿地的区域草地、林地为255(剔除现有公园部分),不适宜建设公园绿地的区域为0,得到城市公园潜在建设范围(图9),利用GIS的按位置选择功能,根据服务压力很大的区域选择适合建设公园绿地的区域,利用GIS面转点功能最终得到公园绿地潜在建设点(图10)。共获得潜在城市公园71个,其中,仓山区可建45个城市公园,鼓楼区可建7个城市公园,晋安区可建19个城市公园,台江区并无可建城市公园,主要原因是台江区属于老城区,适宜建设城市公园区域很少,其公园数量较多,可通过优化部分公园改善其城市公园服务情况,以减少服务压力。

图 9 研究区土地分类图像二值化结果示意图
Figure 9. Land classification image binarization result
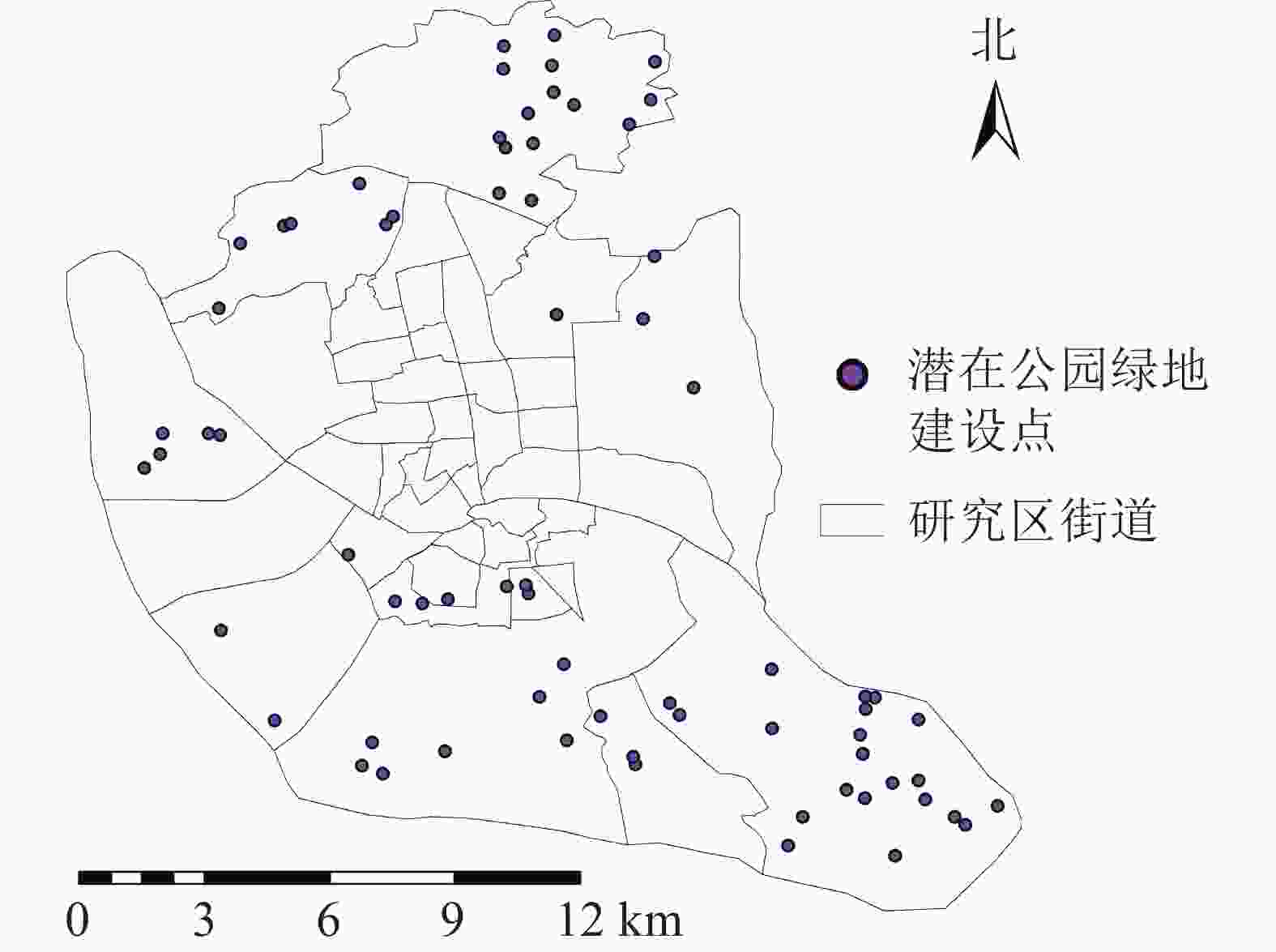
图 10 研究区城市公园潜在建设点示意图
Figure 10. Potential construction sites of urban parks in the study area
-
POI数据以及GIS的应用为研究城市公园如何以人的需求为准则合理选址、确定规模提供更有效方法。通过对大数据的合理应用,对不同城市空间中人们的活动强度分析和对公园进行评价,综合分析服务压力[23]对城市公园布局的影响。人均公园绿地面积、人均绿地面积、绿地率和绿化覆盖率[24]等指标没有反映公园绿地的空间布局,公园绿地与其他用地的关系也难以有效地评价公园使用的效率和公平性。城市公园服务能力的高低是衡量城市居民生活品质的一项重要指标[25],对城市公园服务能力与城市公园布局的分析,有利于提高城市公园以及社会的生态效益。本研究表明:福州主城区城市公园现存服务能力与潜在城市公园布局情况不吻合,反映城市公园建设与潜在需求不吻合,通过对比分析可为后期的城市公园建设提供参考意见。
-
通过比较现状服务能力与潜在服务能力需求对福州主城区城市公园的布局进行分析,现状服务能力由游憩可达性与避灾覆盖率2项指标体现。研究表明:福州主城区城市公园中心城区服务能力好,综合服务设施较丰富,服务能力由中心城区向四周降低。服务能力的分布情况主要与城市道路交通网络的分布相关,游憩可达性越高,城市公园自身避灾覆盖率越大,城市公园的服务能力越好。服务压力的分布与人群活动更为密切,服务压力较大的区域主要在鼓楼区、台江区及晋安区的交界处,主要原因是这些区域属于老城区,人们活动强度比较大。城市公园的布局合理性较好的区域呈聚集性分布,主要分布在服务能力较好,潜在需求较低的区域,包括鼓楼区北部与台江区南部。布局合理性很差的区域主要分布于仓山区的南部及东南部地区,这些区域公园绿地少,避灾覆盖率及游憩可达性较差;晋安区东部及北部地区各项指标也较差,主要原因是其东部存在大量工厂,公园绿地较少,避灾覆盖率较差,服务压力较大,其北部区域避灾覆盖率较差,游憩可达性较差,因此对公园绿地的需求程度较高。
-
对现状服务能力与潜在服务需求分析可知:台江区服务压力最大,服务能力好,在对城市公园布局分析时要考虑其服务设施以及出入口位置的优化;对服务压力小且服务能力差的区域,需结合土地利用情况等对城市公园数量进行优化,即通过对城市土地利用情况的分析显示城市潜在优化城市公园。服务能力与服务压力的分析显示潜在优化城市公园共12个,即福州市主城区共需优化城市公园12个,其中仓山区城市公园需优化3个,鼓楼区需优化城市公园3个,台江区需优化城市公园4个,晋安区需优化城市公园2个。根据实地调查可知:部分城市公园主要缺陷为城市公园内部植物群落单薄,竖向植被结构还需加强,城市公园内部吸引力不足;需提升其城市公园内部属性,通过城市公园内部合理规划来改善游憩环境,包括福州动物园、鼓楼前公园、南公园、劳动者公园、闽江公园(南园)、江滨休闲广场、国光公园、市树公园。部分城市公园主要缺陷为公园入口点过少、公园内部绿地破碎化严重,包括琴亭湖公园、秋龙湖公园、长安山公园、五凤公园;应增加城市公园入口点,减少硬质铺装,提高城市公园的植被覆盖率,以提高其服务能力与游憩环境。
服务压力大,城市公园数量与人口密度不吻合的情况,可通过潜在城市公园建设优化,增加城市公园数量。研究分析城门镇可建21个城市公园,新店镇可建15个城市公园,盖山镇、建新镇、五凤街道各可建7个城市公园,鼓山镇、螺洲镇、仓山镇各可建3个城市公园,东升街道可建2个城市公园,岳峰镇、上渡街道、对湖街道各可建1个城市公园。
Urban park layout in the main urban area of Fuzhou in the light of service ability
-
摘要:
目的 城市公园布局合理性是城市人居环境建设重要体现,分析福州市主城区城市公园的服务能力对提高布局合理性具有重要意义。 方法 利用地理信息系统(GIS)空间统计法,提出游憩可达性与避灾覆盖率2项指标对福州主城区城市公园现状布局的影响;通过城市兴趣点(POI)数据计算城市服务压力对城市公园潜在服务的需求。 结果 台江区、鼓楼区现状服务能力较好,避灾覆盖率大且路网丰富,可达性较好,服务能力最好的是鼓楼前公园;晋安区与仓山区现状服务能力较差,避灾覆盖率小且路网较差,较偏僻地区可达性较差,服务能力最差的是飞凤山公园。城市公园潜在需求分析表明鼓楼区服务压力最大,仓山区服务压力最小,城市公园集中在鼓楼区与仓山区。 结论 城市公园的布局较好,呈聚集性分布;布局合理性很好的区域主要分布在在鼓楼区北部与台江区南部,布局合理性很差的区域主要分布于仓山区的南部及东南部地区、晋安区东部及北部地区,原因是这些区域本来公园绿地就少,避灾覆盖率及游憩可达性较差。从城市空间布局上对城市公园进行优化;对城市公园现状优化有12个,其中,台江区4个,鼓楼区3个,晋安区2个,仓山区3个;根据土地利用情况,建设潜在城市公园:仓山区可建45个城市公园,鼓楼区可建7个城市公园,晋安区可建19个城市公园。图10表3参25 Abstract:Objective With an analysis conducted of the service capacity of urban parks in the main urban area of Fuzhou, this study is aimed to optimize the layout of urban parks, which is reflective of the construction of urban human settlements. Method Firstly, using the GIS spatial statistical method, the current status of urban parks was analyzed with the two indicators of recreational accessibility and disaster avoidance coverage employed. Then the city service pressure was calculated before an analysis was made of the potential service demand of city parks by means of city POI (point of interest) data. Result (1) The current service capacity of Taijiang District and Gulou District is good, as is shown in the large disaster avoidance coverage a well-built transport network and the easy access with Gulouqian Park demonstrating the best service ability; (2) The current service ability of Jinan District and Cangshan District is poor, shown in the small disaster avoidance coverage, the inadequate transport network and the poor accessibility with Feifengshan Park demonstrating the worst service capacity; (3) In terms of potential demand for urban parks, with the urban parks mainly distributed in Gulou District and Cangshan District, Gulou District has the highest service pressure while Cangshan District has the least service pressure. Conclusion In conclusion, the layout of city parks is generally favorable with a distribution featured with spacial aggregation: the city parks with a good layout are mainly distributed in the north of Gulou District and the south of Taijiang District while the ones with poor layout rationality are mainly distributed in the southern and southeastern regions of Cangshan District as well as the eastern and northern regions of Jin’an District. The lack of parkland, small avoidance coverage and poor access in such areas are the main reasons behind the low rationality. As a result, it is of vital importance to optimize the urban spatial layout of the 12 optimized urban parks: 4 are located in Taijiang District, 3 are located in Gulou District, 2 are located in Jin’an District with the rest 3 in Cangshan District. On the other hand, with the current land use circumstances taken into consideration, potential urban parks should be built: 45 in Cangshan District, 7 in Gulou District, and 19 in Jin’an District. [Ch, 10 fig. 3 tab. 25 ref.] -
Key words:
- city park /
- service capacity /
- layout analysis /
- service pressure /
- main area of Fuzhou
-
图 1 研究区城市公园及其入口点分布示意图
Figure 1 Distribution of urban parks and their entry points in the study area
图 4 城市公园不同避灾类型服务域示意图
Figure 4 Service areas of different types of disaster prevention in city parks
图 5 不同街道城市公园避灾服务域覆盖率示意图
Figure 5 Coverage rate of disaster prevention service areas in urban parks in different streets
图 7 研究区城市公园服务压力分布示意图
Figure 7 Service pressure distribution of urban parks in the study area
图 10 研究区城市公园潜在建设点示意图
Figure 10 Potential construction sites of urban parks in the study area
表 1 不同等级游憩可达性面积
Table 1. Area of different grades of recreational accessibility
等级 面积/ km2 比例/% 一级 95.42 38.43 二级 59.54 23.98 三级 29.45 11.86 四级 31.81 12.81 五级 32.08 12.92  下载: 导出CSV
下载: 导出CSV
表 2 不同等级避灾覆盖率街道分布
Table 2. Street distribution of different levels of disaster avoidance coverage
等级 数量/个 街道 一级 3 新店镇、城门镇、螺洲镇 二级 7 鼓山镇、五凤街道、建新镇、上渡街道、仓山镇、盖山镇、东升街道 三级 11 洪山镇、华大街道、岳峰镇、王庄街道、象园街道、瀛洲街道、义洲街道、宁化街道、金山街道、鳌峰街道、三叉街道 四级 16 茶园街道、温泉街道、水部街道、新港街道、鼓东街道、东街街道、鼓西街道、南街街道、安泰街、上海街道、后洲街道、苍霞街道、临江街道、仓前街道、下渡街道、对湖街道 五级 2 茶亭街道、洋中街道
下载: 导出CSV
表 3 不同等级服务压力的公园分布
Table 3. Park distribution of different grades of service pressure
等级 数量/个 街道 一级 8 飞凤山公园、福州动物园、花海公园、高盖山公园、鳌峰公园、乌江湿地公园、沙滩公园、儿童公园 二级 12 五凤公园、仓前公园、国光公园、金牛山公园、夏雨苑公园、琴亭湖公园、市树公园、左海公园、江心公园、金鸡山公园、闽江公园(北园)、金山公园 三级 11 金山公园、文林山公园、西河公园、水上公园、长安山公园、劳动者公园、江滨休闲广场、闽江公园(南园)、榕成广场、金港公园、屏山公园 四级 11 晋安河公园、亚峰公园、白马河公园、环南公园、西禅寺公园、苍霞公园、乌山风景区、烟台山公园、西湖公园、秋龙湖公园、光明港公园 五级 5 南公园、鼓楼前公园、茶亭公园、温泉公园、于山风景区
下载: 导出CSV
-
[1] 陶晓丽, 陈明星, 张文忠, 等. 城市公园的类型划分及其与功能的关系分析:以北京市城市公园为例[J]. 地理研究, 2013, 32(10): 1964 − 1976. TAO Xiaoli, CHEN Mingxing, ZHANG Wenzhong, et al. Classification and its relationship with the functional analysis of urban parks: taking Beijing as an example [J]. Geogr Res, 2013, 32(10): 1964 − 1976. [2] 周媛. 多元目标导向下的成都中心城区绿地生态网络构建[J]. 浙江农林大学学报, 2019, 36(2): 359 − 365. ZHOU Yuan. Developing urban greenspace ecological network in Chengdu City center based on multiple objectives [J]. J Zhejiang A&F Univ, 2019, 36(2): 359 − 365. [3] HUANG Boxun, CHIOU Shangchia, LI Wenying. Accessibility and street network characteristics of urban public facility spaces: equity research on parks in Fuzhou City based on GIS and Space Syntax Model [J]. Sustainability, 2020, 12(9): 3618. [4] 连丽花. 常州市公园绿地布局研究[D]. 南京: 南京林业大学, 2010. LIAN Lihua. Study on Urban Parkland Layout of Changzhou[D]. Nanjing: Nanjing Forestry University, 2010. [5] GUO Shihui, YANG Gege, PEI Tao, et al. Analysis of factors affecting urban park service area in Beijing: perspectives from multi-source geographic data [J]. Landscape Urban Plann, 2019, 181: 103 − 117. [6] 梁颢严, 肖荣波, 廖远涛. 基于服务能力的公园绿地空间分布合理性评价[J]. 中国园林, 2010, 26(9): 15 − 19. LIANG Haoyan, XIAO Rongbo, LIAO Yuantao. Evaluation of reasonable city parks distribution based on service ability [J]. Chin Landscape Archit, 2010, 26(9): 15 − 19. [7] 李文, 张林, 李莹. 哈尔滨城市公园可达性和服务效率分析[J]. 中国园林, 2010, 26(8): 59 − 62. LI Wen, ZHANG Lin, LI Ying. Accessibility and service efficiency of Harbin’s urban parks [J]. Chin Landscape Archit, 2010, 26(8): 59 − 62. [8] 黄曦涛, 李怀恩, 张瑜, 等. 基于人口离散化和交通网络分析的西安市城市公园绿地可达性研究与评价[J]. 遥感信息, 2014, 29(4): 98 − 102. HUANG Xitao, LI Huaien, ZHANG Yu, et al. Research and evaluation of accessibility of Xi’an City Park green space based on population discretization and transportation network analysis [J]. Remote Sensing Inf, 2014, 29(4): 98 − 102. [9] 翟雪, 张涛, 王泽. 基于网络分析法的山地城市公园绿地可达性研究:以承德市中心城区为例[J]. 林业与生态科学, 2018, 33(3): 336 − 342. ZHAI Xue, ZHANG Tao, WANG Ze. Rasearch of the mountainous urban park accessibility based on network analysis: a case study of downtown in Chengde City [J]. For Ecol Sci, 2018, 33(3): 336 − 342. [10] 陈永生, 李莹莹, 张前进. 基于GIS的合肥市中心城区公园绿地功能综合评价[J]. 中国农业大学学报, 2019, 24(3): 137 − 145. CHEN Yongsheng, LI Yingying, ZHANG Qianjin. Comprehensive evaluation of park green space function in the urban area of Hefei City centre: based on GIS [J]. J China Agric Univ, 2019, 24(3): 137 − 145. [11] BANCROFT C, JOSHI S, RUNDLE A, et al. Association of proximity and density of parks and objectively measured physical activity in the United States: a systematic review [J]. Soc Sci Med, 2015, 138: 22 − 30. [12] 刘常富, 李小马, 韩东. 城市公园可达性研究: 方法与关键问题[J]. 生态学报, 2010, 30(19): 5381 − 5390. LIU Changfu, LI Xiaoma, HAN Dong. Acessibility analysis of urban parks: methods and key issues [J]. Acta Ecol Sin, 2010, 30(19): 5381 − 5390. [13] EKKEL E D, de VRIES S. Nearby green space and human health: evaluating accessibility metrics [J]. Landscape Urban Plann, 2017, 157: 214 − 220. [14] 李维宇, 乔庆华, 王中辉, 等. 中心城区广场公园绿地可达性研究[J]. 测绘与空间地理信息, 2018, 41(4): 94 − 97. LI Weining, QIAO Qinghua, WANG Zhonghui, et al. Study on accessibility of square and park with green belt in center city [J]. Geomatics Spat Inf Technol, 2018, 41(4): 94 − 97. [15] 郭亚淑. 福州市土地利用/覆被变化对生态环境脆弱性的影响评价[D]. 福州: 福建农林大学, 2016. GUO Yashu. The Impact Assessment of on Eco-environmental Vulnerability Based on Land Use/Cover Change in Fuzhou[D]. Fuzhou: Fujian Agriculture and Forestry University, 2016. [16] 戚荣昊, 杨航, 王思玲, 等. 基于百度POI数据的公园绿地评估与规划研究[J]. 中国园林, 2018, 34(3): 32 − 37. QI Ronghao, YANG Hang, WANG Siling, et al. Study on evaluation and planning of urban parks based on Baidu POI data [J]. Chin Landscape Archit, 2018, 34(3): 32 − 37. [17] 贺建雄. 西安城市居民日常生活空间供需耦合研究[D]. 西安: 西北大学, 2018. HE Jianxiong. Study on the Supply and Demand Coupling of Xi’an Urban Residents’ Daily Living Space[D]. Xi’an: Northwest University, 2018. [18] 李翔. 基于GIS的地震应急避难场所布局分析与优化研究[D]. 西安: 西安建筑科技大学, 2016. LI Xiang. Research on Layout Analysis and Optimization of Earthquake Emergency Shelters Based on GIS[D]. Xi’an: Xi’an University of Architecture and Technology, 2016. [19] 唐少军. 基于GIS的公共服务设施空间布局选址研究[D]. 长沙: 中南大学, 2008. TANG Shaojun. The Research of Spatial Layout and Location of Public Service Facility Based on GIS[D]. Changsha: Central South University, 2008. [20] OH K, JEONG S. Assessing the spatial distribution of urban parks using GIS [J]. Landscape Urban Plann, 2007, 82(1/2): 25 − 32. [21] COMBER A, BRUNSDON C, GREEN E. Using a GIS-based network analysis to determine urban green space accessibility for different ethnic and religious groups [J]. Landscape Urban Plann, 2008, 86(1): 103 − 114. [22] 和树明. 城市公园绿地防灾避险功能评价与规划研究[D]. 济南: 山东建筑大学, 2017. HE Shuming. Research on Functional Assesment and Planning of Park Greenspace with Disaster Prevention and Danger Avoidance in Zaozhuang City[D]. Ji’nan: Shandong Jianzhu University, 2017. [23] 蒋理, 殷振轩, 刘晓. 基于可达性分析的城市公园绿地供给研究: 以广西玉林市为例[J]. 风景园林, 2019, 26(8): 83 − 88. JIANG Li, YIN Zhenxuan, LIU Xiao. Study on urban park green space supply based on accessibility analysis: a case study of Yulin City, Guangxi [J]. Landscape Archit, 2019, 26(8): 83 − 88. [24] 福州统计局. 福州统计年鉴[Z]. 北京: 中国统计出版社, 2018. [25] 屠星月, 黄甘霖, 邬建国. 城市绿地可达性和居民福祉关系研究综述[J]. 生态学报, 2019, 39(2): 421 − 431. TU Xingyue, HUANG Ganlin, WU Jianguo. Geview of the relationship between urban greenspace accessibility and human well-being [J]. Acta Ecol Sin, 2019, 39(2): 421 − 431. -

-
链接本文:
https://zlxb.zafu.edu.cn/article/doi/10.11833/j.issn.20950756.20200314
 点击查看大图
点击查看大图
计量
- 文章访问数: 2373
- HTML全文浏览量: 658
- PDF下载量: 51
- 被引次数: 0
