








-
在气候变暖的背景下,高温和干旱等灾害天气出现更为频繁。温度和水分作为全球气候变化的两大重要表征,是影响植物生长和分布的最重要的非生物因素[1]。两者独立或交互作用,都会对植物生长发育产生极大影响[2]。活性氧(ROS)如过氧化氢(H2O2),超氧阴离子(O2·-),羟自由基(·OH)和单线态氧(1O2)等,是植物中氧化还原级联的有氧代谢产物,如果不能被及时清除,将产生很强的氧化性,导致细胞脂质过氧化,对细胞的蛋白质、核酸和脂质等生物大分子产生永久性伤害,进而引起细胞死亡[3]。此外,随着ROS的积累,生物膜不饱和脂肪酸分解产生的丙二醛(MDA)[4],一方面加剧细胞质膜过氧化程度,导致膜系统损伤,使细胞受到伤害[5];另一方面植物细胞产量显著减少,最终引起机体衰老[6]。植物处于逆境下并不是被动承受伤害,而是主动调节适应。植物在进化过程中形成了相应的酶促保护系统[如超氧化物歧化酶(SOD),过氧化物酶(POD)和过氧化氢酶(CAT)等],能有效清除ROS,提高植物耐受能力[7]。RAMÍREZ等[8]研究发现,拟南芥Arabidopsis thaliana通过维持体内非酶系统[抗坏血酸-谷胱甘肽(AsA-GSH)循环]快速有效运转,减轻逆境胁迫下植物的活性氧伤害,使植物更好地适应环境。学者对单一逆境胁迫条件下植物抗氧化防御系统的研究已有报道,并被认为是一个能够反映植物抗性大小的普遍标记[2, 9],但是高温干旱复合胁迫对抗氧化防御系统的研究较为少见。毛竹Phyllostachys edulis在中国分布极广,具有重要的社会、经济和生态价值[10]。目前,针对毛竹抗逆性的研究主要集中在光合作用[11]、生长形态[12]和单一逆境下的抗氧化酶活性[13]等方面。本研究以毛竹为研究对象,通过设置高温与干旱胁迫不同水平的处理,分析高温和干旱叠加条件下毛竹体内ROS的产生情况,探讨抗氧化酶系统和非酶系统抵御氧化胁迫的变化规律,揭示高温和干旱双重胁迫下毛竹生理生化响应机制及其抗高温和干旱的能力,为毛竹林培育提供理论依据。
HTML
-
试验材料为3年生毛竹实生苗,苗高为1.0 m左右。于2015年8月下旬,选取生长良好形态相近的毛竹幼苗移栽于塑料花盆中(花盆高为50.0 cm,内径为60.0 cm),取浙江临安当地的黄壤土作为栽培土壤,pH 6.9~7.0,1株·盆-1,在缓苗期间,用自来水浇灌。选取生长良好,无病虫害,长势相近的毛竹25盆,其中对照组10盆,其余随机分成3组(处理),5盆·组-1(重复),在常温条件下(25 ℃/20 ℃,昼/夜)栽培。将对照组中的10盆随机分为2组,5盆·组-1,其中1组作为干旱处理的对照组,正常管理,另一组作为高温处理组,正常管理。
干旱处理:在缓苗30 d后进行。设计4个梯度的处理:对照70.0%~80.0%田间持水量(FC),轻度干旱(60.0%~70.0% FC),中度干旱(40.0%~50.0% FC)和重度干旱(20.0%~30.0% FC)。采用称量法控制土壤水分含量,每天17:00-18:00称量并补充消耗的水分,干旱处理10 d。选取毛竹植株中上部完整的功能叶片取样,样品用液氮迅速冷冻,置于-80 ℃低温冰箱内保存。温度处理:在8:00时将其中1组对照组和干旱处理过的毛竹盆栽苗放入40 ℃/30 ℃(昼/夜)人工气候箱,设定昼夜相隔时间12 h,光照强度设定为400 μmol·m-2·s-1,高温处理48 h后取毛竹植株中上部完整的功能叶片进行相关指标测定。
-
O2·-测定参照SHAH等[14]的方法并略作改进:取0.2 g冷冻样品,加5.0 mL磷酸缓冲液(100.0 mmol·L-1,pH 7.2,含1.0 mmol·L-1二乙基二硫代氨基甲酸钠)研磨,15 000 g离心30 min(4 ℃)。3.0 mL反应体系包含:2.0 mL样品提取液和1.0 mL氮蓝四唑(NBT,0.25 mmol·L–1),在540 nm处测定3 min内吸光度的变化。结果表示为ΔD(540)·min-1·g-1。过氧化氢测定参照RAI等[15]的方法并略作改进:取0.2 g冷冻样品,加5.0 mL磷酸缓冲液(50.0 mmol·L-1,pH 6.5)磨碎,3.0 mL样品提取液与体积分数为20%的硫酸(含质量分数为0.1%硫酸钛)1.0 mL混合,16 000 g离心15 min(4 ℃),取上清液测定410 nm处的吸光度。过氧化氢的质量摩尔浓度用吸光系数0.28 μmol-1·cm-1来计算,结果表示为μmol·g-1。MDA测定参照HODGES等[16]的方法。
-
取0.2 g冷冻样品液氮研磨,加5.0 mL磷酸缓冲溶液(50.0 mmol·L-1,pH 7.8)匀浆,10 000 g离心10 min(4 ℃)。上清液用于SOD,POD和CAT活性的测定。SOD活性测定参照GIANNOPOLITIS等[17]的方法。POD活性测定参照KUMARI等[18]的愈疮木酚法。CAT活性测定参照KUMARI等[18]的方法。
-
取0.2 g冷冻样品液氮研磨,加50.0 g·L-1偏磷酸5.0 mL匀浆,15 000 g离心15 min(4 ℃),上清液用于AsA和GSH的测定。AsA和GSH测定参照RAI等[19]的方法。
-
取0.2 g样品用5.0 mL磷酸缓冲溶液(pH 7.4,含1.0 mmol·L-1乙二胺四乙酸EDTA)冰浴研磨成匀浆,混匀后在16 000 g离心20 min(4 ℃)。上清液为毛竹AsA-GSH循环的相关酶活性测定的酶液。抗坏血酸过氧化物酶(APX)活性测定参照NAKANO等[20]的方法。谷胱甘肽还原酶(GR)活性测定参照SCHAEDLE等[21]的方法。单脱氢抗坏血酸还原酶(MDHAR)活性测定参照HOSSAIN等[22]的方法。脱氢抗坏血酸还原酶(DHAR)活性测定参照DOULIS等[23]的方法。
-
数据分为2组进行统计分析:一组为干旱胁迫处理,另一组为高温干旱胁迫处理,高温对照组即高温处理组,所有的数据均取5次重复的平均值±标准误。各组的数据利用Origin 9软件(美国Origin Lab公司)进行统计分析和作图。统计方法采用单因素方差分析(one-way ANOVA)进行检验,并进行Turkey多重比较。采用双因素方差分析(two-way ANOVA)评估高温胁迫×干旱胁迫的协同作用。
1.1. 材料处理
1.2. 测试方法
1.2.1. O2·-,过氧化氢和MDA测定
1.2.2. 抗氧化酶活性测定
1.2.3. AsA和GSH测定
1.2.4. AsA-GSH循环相关酶活性测定
1.3. 数据处理
-
随干旱胁迫程度的增加毛竹叶片中的O2·-,过氧化氢和MDA质量摩尔浓度逐渐上升(表 1),其中在轻度干旱胁迫条件下,O2·-和MDA质量摩尔浓度分别比对照高39.9%和68.8%(P<0.01);中度干旱胁迫时,O2·-,MDA和过氧化氢质量摩尔浓度分别比对照高74.4%,94.5%和38.8%(P<0.01)。在高温胁迫条件下,O2·-,过氧化氢和MDA质量摩尔浓度分别比对照高44.2%,21.9%和140.9%(P<0.01)。在协同胁迫下,三者质量摩尔浓度进一步增加,在轻度干旱和高温协同胁迫下,O2·-,过氧化氢和MDA质量摩尔浓度分别比对照高67.6%,28.8%和170.5%(P<0.01)。
T/℃ 项目 处理 超氧阴离子O2·-/[D(540)·min-1·g-1] 过氧化氢/(μmol·g-1) MDA/(μmol·g-1) 25 对照 7.5 ± 0.3 D 36.5 ± 2.8 B 9.6 ± 2.6 C 轻度干旱 10.5 ± 0.5 C 42.4 ± 5.5 B 16.1 ± 1.6 B 中度干旱 13.1 ± 0.3 B 50.7 ± 5.2 A 18.6 ± 1.1 AB 重度干旱 14.7 ± 0.1 A 50.7 ± 5.2 A 20.6 ± 0.4 A 平方和 组间(df1=3) 118.0 861.1 276.1 组内 1.2 463.0 31.2 40 对照 10.8 ± 0.1 d 44.5 ± 1.6 c 23.0 ±1.3 bc 轻度干旱 12.6 ± 0.4 c 47.0 ± 3.5 c 25.8 ±3.0 b 中度干旱 14.6 ± 0.3 b 53.1 ± 2.9 b 29.0 ±1.0 ab 重度干旱 15.4 ± 0.1 a 57.6 ± 2.7 a 31.8 ± 0.5 a 平方和 组间(df1=3) 51.2 626.8 174.5 组内 0.8 150.9 36.6 P(Fm) * ** ns P(Fs) ** * ** P(Fs×Fm) ns ns ns 说明:同列不同大写字母表示25 ℃时不同处理类型间差异显著, 同列不同小写字母表示40 ℃时不同处理类型间差异显著。Fs表示不同温度间的差异;Fm表示不同处理类型的影响;Fs×Fm表示植物组织应对高温干旱胁迫的不同响应;*表示P<0.05;**表示P<0.01;ns表示差异不显著 Table 1. Effects of high temperature and drought stress on ROS and MDA contents in Phyllostachys edulis
-
随干旱胁迫程度的增加,毛竹叶片的SOD,POD和CAT活性均逐渐增加(图 1)。中度干旱胁迫下,SOD和POD活性分别比对照高1.7倍和1.7倍(P<0.01);轻度干旱胁迫下,CAT活性比对照高1.9倍(P<0.05)。在高温胁迫条件下,SOD,POD和CAT活性分别比对照高0.8倍、0.6倍和1.6倍,三者活性与其对照的差异均极显著(P<0.01)。协同胁迫使SOD和POD活性呈先增加后降低,且均在中度干旱水平的协同胁迫下达到峰值,分别比对照高2.9倍和3.2倍(P<0.01);CAT活性呈逐渐增加趋势,在中度干旱和高温协同胁迫下与对照的差异极显著(P<0.01)。

Figure 1. Changes of antioxidant enzyme activities in Phyllostachys edulis under different treatments
-
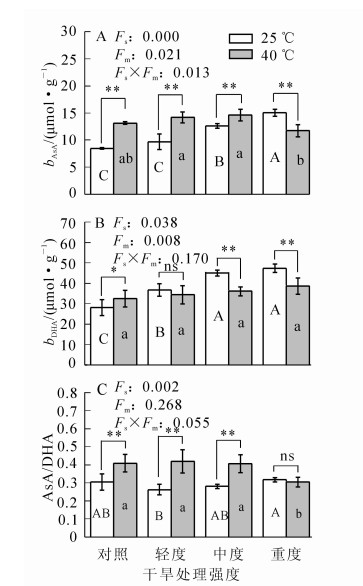
随干旱胁迫程度的增加,毛竹叶片的AsA和二十二碳六烯酸(DHA)质量摩尔浓度均呈逐渐上升趋势(图 2),在中度干旱时AsA质量摩尔浓度比对照高49.5%(P<0.01);在轻度干旱时DHA质量摩尔浓度比对照高30.6%(P<0.01);ρAsA/ρDHA呈先下降后上升,在轻度干旱时达到最小值,比对照降低13.9%(P>0.05)。在高温条件下,AsA质量摩尔浓度和ρAsA/ρDHA分别比对照高55.6%和34.1%(P<0.01);DHA质量摩尔浓度比对照高15.6%(P<0.05)。协同胁迫下,三者变化均不大:其中AsA质量摩尔浓度呈先增加后降低,在中度干旱和高温的协同胁迫时达最大值,比对照高72.9%(P<0.01),随后降低;DHA质量摩尔浓度总体变化不大,轻度干旱和高温协同胁迫时,比对照高22.3%(P<0.05),其他变化不显著;ρAsA/ρDHA变化不大,在轻度干旱和高温协同胁迫时达最大值,比对照高37.3%(P<0.01)。

Figure 2. Changes in the content of ascorbic acid in Phyllostachys edulis under different treatments
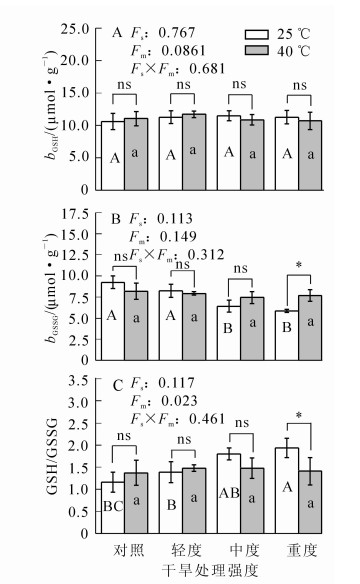
在干旱胁迫条件下,毛竹叶片的还原型谷胱甘肽(GSH)质量摩尔浓度变化与对照差异不显著(P>0.05)(图 3);氧化型谷胱甘肽(GSSG)质量摩尔浓度呈下降趋势,中度干旱胁迫时其质量摩尔浓度比对照降低30.6%(P<0.01);ρGSH/ρGSSG呈上升趋势,中度干旱比对照高55.3%(P<0.01)。在高温胁迫及协同胁迫下,三者的变化与对照比较均不显著(P>0.05)。

Figure 3. Changes in the content of glutathione in Ph. edulis under different treatments
-
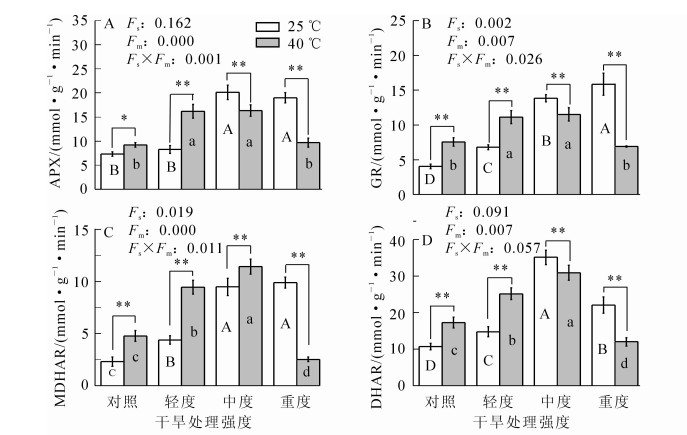
随干旱胁迫程度的增加,毛竹叶片的APX酶活性先逐渐增加后变化缓慢(图 4),在中度干旱胁迫下达到最大值,比对照高1.8倍(P<0.01);GR和MDHAR活性均呈上升趋势,轻度干旱胁迫时其活性分别比对照高0.7倍和0.9倍(P<0.01);DHAR活性先增加后降低,中度干旱胁迫时达到最大值,比对照高2.3倍(P<0.01)。高温胁迫时,APX活性比对照高0.3倍(P<0.05);而GR,MDHAR和DHAR活性分别比对照高0.9倍、1.1倍和0.6倍(P<0.01)。在高温的基础上,随着干旱胁迫程度的加深,四者活性均呈先增加后降低,APX和GR活性增加缓慢,MDHAR和DHAR活性增加迅速,中度干旱和高温协同胁迫时均达到最大值,分别比对照高1.2倍、1.9倍、3.9倍和1.9倍(P<0.01),随后降低。

Figure 4. Changes of enzyme activities in AsA-GSH cycle system in Phyllostachys edulis under different treatments
2.1. O2·-,过氧化氢和MDA变化
2.2. SOD,POD与CAT活性变化
2.3. 抗氧化物的变化
2.4. AsA-GSH循环系统相关酶活性变化
-
细胞膜对维持细胞正常生命活动具有至关重要的作用,而O2·-和过氧化氢是植物代谢过程中产生的2种活性氧,对细胞膜脂质具有较强的氧化伤害作用;MDA作为膜脂过氧化作用的终极产物,其变化可解释逆境胁迫对植物细胞膜的破坏程度及植物对逆境的响应[24]。本试验结果表明:干旱胁迫下,毛竹叶片中的O2·-和MDA产生速率明显增加,而高温胁迫下,过氧化氢、O2·-和MDA产生速率增加均极显著,且高温时MDA质量摩尔浓度较中度干旱胁迫显著增加。这与吴永波等[2]的研究结果不一致,表明高温比干旱对毛竹的影响更大,可能是在不同温度和水分条件下,不同植物体内活性氧对植物产生的伤害程度不同导致的。协同胁迫下,随着干旱程度的增加,三者质量摩尔浓度逐渐增加,且在同等干旱程度下,高温条件下的O2·-和MDA质量摩尔浓度均比正常温度下的质量摩尔浓度显著升高,表明协同胁迫较单一胁迫对毛竹伤害更大,可能协同胁迫已经伤害到抗氧化物质产生的机构,削弱了对氧化物质的清除作用,导致氧化物质不断累积。
毛竹处于逆境中,不仅体内过氧化氢、O2·-和MDA的产量大量增加,还刺激抗氧化酶活性增强。SOD,POD和CAT是植物抗氧化系统重要组成部分[25]。其中SOD是抗氧化系统的第一道防线,可以使植物体内过量的O2·-发生歧化反应,生成过氧化氢(H2O2)和氧气(O2);POD和CAT与SOD具有协同作用,将其氧化产生的H2O2和具有潜在危害的O2·-转化为水和氧气,从而阻止O2·-和过氧化氢在植物体内的累积,减少对植物体的伤害。本研究发现:随干旱高温胁迫程度的增加,毛竹叶片产生过量的ROS,且抗氧化酶活性升高。说明O2·-质量摩尔浓度增加,它的积累刺激叶片中SOD应激反应使活性增强,催化O2·-歧化生产基态分子氧和过氧化氢;毛竹叶片中的过氧化氢和MDA逐渐增加,由于POD和CAT的作用,其质量摩尔浓度增幅缓慢;而在重度干旱和高温协同胁迫下,SOD和POD活性较中度干旱和高温协同胁迫显著降低,CAT活性降低不明显,可能是SOD活性表达受到抑制,且过度胁迫导致一部分已经表达的SOD失活[26]造成的,而POD和CAT酶活性分别在高温干旱前期和后期对过氧化氢清除起主导作用[27],表明在持续时间较长的高温干旱条件下,CAT效率更高,耐性更稳定。总之,在单一胁迫条件时,毛竹在抗氧化酶防御系统作用下,能有效消除因高温干旱胁迫积累的ROS,使过氧化氢、O2·-和MDA质量摩尔浓度增幅减缓,表明一方面胁迫条件下毛竹体内产生的ROS能够对毛竹产生伤害,另一方面毛竹在遇到胁迫会刺激抗氧化酶活性增加,有效清除ROS,维持毛竹体内活性氧平衡,防止氧化伤害,适应环境变化。而在协同胁迫条件时,过氧化氢、O2·-和MDA质量摩尔浓度显著增加,SOD,POD和CAT活性有降低趋势,表明协同胁迫比单一胁迫对植物造成的伤害更大。
AsA-GSH循环是植物体内清除自由基的重要途径。AsA不仅可以直接清除活性氧,在AsA-GSH循环清除过氧化氢过程中也发挥着重要作用[27]。APX是以AsA为电子供体的一种过氧化物酶,能够快速清除细胞中产生的过量的过氧化氢,是植物体内尤其是叶绿体中清除过氧化氢的关键酶[28];植物通过维持较高的ρAsA/ρDHA和ρGSH/ρGSSG比值,使植物体内抗坏血酸库和谷胱甘肽库保持较高的还原态[29-30]。本研究结果表明:在单一胁迫下,毛竹叶片产生过量的H2O2,AsA和DHA质量摩尔浓度随之增加,一方面直接消除过氧化氢降低伤害;另一方面通过AsA-GSH循环清除过氧化氢。但轻度干旱胁迫下,ρAsA/ρDHA比值最低,可能是由于干旱胁迫初期,毛竹叶片产生过量的过氧化氢,AsA清除过氧化氢有所消耗,而AsA-GSH循环不能及时补充AsA,使得ρAsA/ρDHA比值降低;随胁迫程度增强,毛竹适应此逆境后,AsA-GSH循环达到一个新的稳态,及时补充AsA的损失,维持较高的ρAsA/ρDHA比值,抵抗氧化胁迫;同时,GSH质量摩尔浓度变化较对照不明显,GR活性增加,与GSSG质量摩尔浓度变化相反,说明GR在还原型烟酰胺嘌呤二核苷酸磷酸(NADPH)的作用下,加速催化GSSG还原成GSH,使毛竹叶片中的ρGSH/ρGSSG比值增加,维持植物体内较高的还原态,减缓毛竹氧化伤害。这与杨颖丽等[31]研究结果一致。在干旱和高温协同胁迫下,AsA,DHA,GSH和GSSH质量摩尔浓度变化不大,而APX,MDHAR,DHAR和GR活性先增加后降低,均在中度干旱和高温协同胁迫时达到峰值,这是因为在AsA-GSH循环中,毛竹受到较强程度协同胁迫的刺激,使叶片中清除过氧化氢的APX和加快AsA循环再生的MDHAR和DHAR活性的增加,加快AsA和DHA大量快速生成,及时补充AsA清除过氧化氢的消耗;同时,毛竹叶片中GSSG在关键酶GR作用下催化产生GSH,DHAR利用GSH提供的电子供体将DHA还原成AsA,使AsA质量摩尔浓度保持稳定,进而促进AsA-GSH循环,使ρAsA/ρDHA比值维持在较强的还原性范围内,以利于抵御氧化胁迫;此时ρGSH/ρGSSG比值较为稳定,说明GSH循环比AsA循环对过氧化氢的还原作用更为稳定,清除过氧化氢的作用更为持久有效,因此GR作为AsA-GSH循环中最后一步关键酶,在毛竹抵抗高温干旱胁迫中发挥重要作用。但在重度干旱和高温协同胁迫下,APX,MDHAR,DHAR和GR活性降低,AsA质量摩尔浓度随之降低,导致ρAsA/ρDHA比值也略有降低,说明毛竹叶片在高强度的协同胁迫下,AsA-GSH循环中的酶受温度和水分的影响,活性降低,此循环不能及时催化产生AsA,补充AsA清除过氧化氢产生的消耗,导致AsA-GSH循环效率降低,抵御逆境的能力降低。而DHA质量摩尔浓度变化不大,这与MOSTOFA等[32]研究结果不一致,这可能是过度胁迫条件下,由于MDHA极不稳定,既可还原成AsA,也可进一步歧化为DHA,又因为过多的DHA积累对细胞有害,所以MDHAR能限制MDHA产生以及进一步转化为DHA;此外,由于DHAR活性降低,无法利用GSH提供的电子供体把DHA还原成AsA[33],使得毛竹叶片中的AsA循环被打破,形成新的动态循环,也说明这些酶对胁迫的反应因物种而异。这也表明毛竹幼苗的AsA循环对逆境反应较GSH循环更敏感,而GSH循环在抵御逆境的能力更稳定。
-
随高温干旱单一处理强度的增加,毛竹叶片中的过氧化氢、O2·-与MDA质量摩尔浓度增加,且协同胁迫下,三者质量摩尔浓度进一步增加,表明高温干旱协同胁迫下较单一胁迫对毛竹伤害更大。干旱和高温协同胁迫下,毛竹叶片内抗氧化防御酶活性逐渐增强,且CAT清除过氧化氢较POD活性表现更为稳定,说明在抗氧化酶系统中CAT清除过氧化氢效果更为持久,在协同胁迫后期对防御氧化伤害起到主要作用;AsA-GSH循环相关酶活性随处理的增强而增加,通过AsA和GSH的再生,AsA-GSH循环动态平衡得以维持,有效消除过氧化氢,保护毛竹细胞不受损害,且在AsA-GSH循环中,ρGSH/ρGSSG比值较ρAsA/ρDHA比值更为稳定,说明GSH循环清除过氧化氢比AsA循环的效果更为持久,对持续协同胁迫起到重要防御作用。但是在重度干旱和高温协同胁迫下,毛竹叶片抗氧化防御酶活性和AsA-GSH循环相关酶活性均表现下降趋势,说明毛竹的酶系统和非酶系统对高温干旱协同胁迫的抵御能力是有限的,在一定范围内可有效清除活性氧,超出一定范围则降低。
 DownLoad:
DownLoad: