








-
谷胱甘肽转移酶(GSTs)是一个多功能的二聚体酶超家族,具有解毒、清除细胞内活性氧等功能[1]。在植物中,GSTs根据其功能和序列等特征被分为8个亚家族:alpha,mu,pi,sigma,theta,kappa,zeta和线粒体(microsomal)GSTs[1-3]。目前,很多植物中的GST基因被陆续克隆[4-5],并进行功能研究,发现来自不同植物的GST同源基因或相同植株的不同GST基因,其功能具有一定差异。大多数GSTs基因具有响应逆境胁迫的功能。如前期研究发现,柽柳Tamarix hispida的zeta家族基因ThGSTZ1能被氯化钠,聚乙二醇(PEG 6000),脱落酸(ABA)和甲基紫精(MV)等胁迫调控,过量表达情况下能提高转基因株系抵抗盐、旱、MV及ABA等胁迫的能力[6-7]。核桃Juglans regia的JrGSTTau1基因也能响应不同逆境刺激,过量表达能改善植株应对低温胁迫的能力[8]。在羽衣甘蓝Brassica oleracea中分离获得65个GST基因,其中BoGSTU19,BoGSTU24,BoGSTF10能被冷胁迫强诱导,推测它们与冷胁迫响应具有重要关系[9]。水稻Oryza sativa OsGSTl2转基因拟南芥Arabidopsis thaliana表现出较高的重金属耐受力[10]。这些研究表明:GST基因在植物响应逆境应答及调节中具有多方面的作用,因此具有重要的研究价值。GST家族成员众多,目前对GST基因的研究主要集中在草本植物,对木本植物GST基因的研究较少,特别是经济干果核桃鲜见报道。核桃属多年生落叶乔木,是中国主要经济树种之一。近年来全球环境的变化,环境因子特别是西北地区越冬入春出现的“倒春寒”及夏秋核桃成熟期严重的高温干旱等气候现象,严重制约了核桃产业的发展。因此,筛选核桃逆境响应重要基因,研究其逆境响应功能机制,将对了解核桃的逆境适应机制具有指导作用。本研究从核桃中鉴定获得1条GST基因JrGSTU23,通过生物信息及定量表达分析其生物学功能,以期为核桃抗逆响应研究提供候选基因。
HTML
-
选取培养于相同条件下的2年生‘香玲’‘Xiangling’核桃嫁接苗用作研究材料。处理包括非生物胁迫[100 g·kg-1聚乙二醇(PEG 6000),0.3 mol·L-1氯化钠、低温6 ℃]和激素处理[0.1 mmol·L-1脱落酸(ABA)],100.0 mg·L-1茉莉酸(MeJA)及2.0 mg·L-1水杨酸(SA)。分别在0,3,6,12,24,48 h取样,以0 h正常浇水作为对照,重复3次·处理-1。分别收集各处理后的根和叶,用液氮速冻后保存于-80 ℃冰箱备用。
-
以“glutathione transferase”为关键词在‘香玲’核桃转录组数据中查找GST基因,经BLAST比对选取其中1条GST基因(命名为JrGSTU23)进行分析。用ORF finder()确定JrGSTU23基因开放读码框(ORF),再根据ORF两端序列设计引物JrGSTU23-F和JrGSTU23-R(表 1),进行聚合酶链式反应(PCR)扩增。产物经回收纯化后与pMD-18-T载体连接并转化大肠埃希菌Escherichia coli DH5ɑ感受态细胞。挑取阳性克隆扩大培养进行菌液PCR验证,对获得目的片段的克隆测序。利用Expasy ProtParam(http://web.expasy.org/protparam/)对确认的JrGSTU23基因序列特征进行分析。利用BLASTP()进行序列同源性搜索;利用Clustal 3.0软件对不同物种的GST蛋白进行多序列比对和进化分析。使用PlantCARE(http://bioinformatics.psb.ugent.be/webtools/plantcare/html/)分析该基因启动子中含有的顺式作用元件。利用Expasy中Swiss Model程序同源建模,推测该蛋白的三维结构模型。
引物名称 正向引物(5′→3′) 反向引物(5′→3′) 18SrRNA GGTCAATCTTCTCGTTCCCTT TCGCATTTCGCTACGTTCTT JrGSTU23-DL-F/JrGSTU23-DL-R GTGAAGCTGATTGCCACT GTCCTTCCATGTCTCCTC JrGSTU23-F/JrGSTU23-R ATGGGGGATAAGGTGAAG TCATGGTGTATTGGCTGC Table 1. The used primers
-
各样品总核糖核酸(RNA)采用十六烷基三甲基溴化铵(CTAB)方法提取[8],RNA经DNA消化酶处理后采用PrimeScriptTM RT Reagent Kit(CWBIO,康为世纪,中国)反转录为cDNA,稀释10倍后用作实时荧光定量PCR(qRT-PCR)的模板。qRT-PCR参照SYBR Green Real time PCR Master mix(CWBIO)进行,内参基因为核桃18S rRNA(HE574850)基因[10]。JrGSTU23定量引物为DL-F和DL-R(表 1)。定量反应仪器为Applied Biosystems生产的Step OneTM Real-Time PCR System。反应程序为:94 ℃预变性30 s;94 ℃变性12 s,60 ℃退火45 s,72 ℃延伸45 s,45个循环;81 ℃读板1 s,重复3次·样品-1。采用2-ΔΔCt法对定量结果进行相对分析[11],所有表达值均做了以2为底的对数转化。
1.1. 材料及处理
1.2. JrGSTU23基因的克隆与分析
1.3. JrGSTU23的表达分析
-
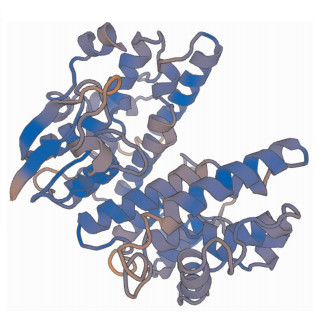
通过查找核桃转录组数据获得1条GST基因,根据获得的cDNA序列设计引物JrGSTU23-F/R进行PCR验证,经分析发现该基因ORF长684 bp,拟推导的蛋白分子量为25.89 kDa,含有氨基酸数为227,理论等电点为5.20。BLAST分析发现:该基因与核桃转录组中的GST23基因相同。保守结构域分析发现:该蛋白具有GST-Tau保守域(图 1),表明该蛋白属于GST氧硫还蛋白亚家族(thioredoxin-line superfamily,Thi),因此命名为JrGSTU23(GeneBank登录号:MG356784)。经美国生物技术信息中心(NCBI)同源搜索获得相似蛋白,并进行进化分析,发现JrGSTU23蛋白与香蕉Musa accuminata,毛果杨Populus trichocarpa等的进化关系较近(图 2)。通过Swiss Model程序同源建模,推测该蛋白的三维结构如图 3所示。]

Figure 1. Amino acid sequence alignment between JrGSTU23 and its homologous proteins from other species

Figure 2. Phylogenetic tree of JrGSTU23 protein and its homologs from other species

Figure 3. Predicted 3D structure model of protein JrGSTU23
-
以JrGSTU23在NCBI数据库中进行同源搜索,发现其与‘强特勒’核桃的GST序列(XM_018974105.1)一致。因此,分析XM_018974105.1序列起始密码子上游2 000 bp的DNA序列作为基因启动子,并对其顺式作用元件进行分析,为预测JrGSTU23基因的功能及可能调控机制提供参考依据。PlantCARE预测结果显示,JrGSTU23启动子包含多个与逆境响应及激素调控相关的顺式作用元件,如热胁迫响应元件(HSE),干旱响应元件(MBS),茉莉酸响应元件(CGTCA-motif),水杨酸响应元件(TCA-element)等(表 2)。
顺式作用元件 起始位点 方向 序列 特性 AT-rich element 1 436 + ATAGAAATCAA ATBP-1结合位点 ATC-motif 1 125 + AGTAATCT 光响应 Box 4 298/353/302/368 +/+/+/+ ATTAAT Box Ⅰ 344/453 -/+ TTTCAAA GAG-motif 121/157 -/- AGAGATG GATA-motif 1 235 + GATAGGG 1-box 1 235/1 249 +/- GATAGGG/GATATGG Sp1 1 147/1 354 +/+ CC (G/A) CCC chs-CMA2a 580 - GCAATTCC CCGTCC-box 1 157 + CCGTCC 分生组织激活 CGTCA-motif 197 + CGTCA 茉莉酸响应 TGACG-motif 197 - TGACG HSE 126 - AGAAAATTCG 热胁迫响应 MBS 669 - TAACTG 干旱胁迫响应 02-site 1 249 - GATGATATGG 玉米素代谢调控 RY-element 645 + CATGCATG 种子调控 Skn-1 motif 379 - GTCAT 胚乳表达 TC-rich repeats 393 + ATTTTCTTCA 防卫和胁迫响应 TCA-element 316 + CCATCTTTTT 水杨酸响应 TCA-element 192 + AACGAC 激素响应元件 WUN-motif 736 TCATTACGAA 创伤响应元件 Table 2. Cis-acting regulatory elements in JrGSTU23 promoter predicted by PlantCARE
-
对试材分别进行ABA,MeJA,SA等激素处理。qRT-PCR实验发现:JrGSTU23能被这些激素明显诱导,但在根和叶的表达趋势不同(图 4)。在叶中,ABA处理3~6 h被抑制,24 h达最大表达水平(2.85);MeJA胁迫下与ABA相反,随着胁迫时间延长,JrGSTU23的表达水平逐渐下降,在24 h被抑制(-0.98);SA胁迫3~6 h也被抑制,在24 h达最大值4.13,但其最低值出现在3 h,为-0.01。在根中,JrGSTU23在ABA胁迫下的最大和最小转录水平分别出现在12和24 h,分别为4.35和2.74;MeJA处理下,该基因的最大和最小表达量分别为7.24(12 h)和2.73(3 h);而在SA胁迫下,JrGSTU23的表达随胁迫时间延长而增强,最大值为4.57倍(24 h)。表明JrGSTU23基因能不同程度响应ABA,MeJA,SA的胁迫,并表现出组织特异性,但其具体的响应机制可能不同。

Figure 4. Expression level of JrGSTU23 gene under different stresses
-
qRT-PCR结果显示:JrGSTU23能被氯化钠,聚乙二醇(PEG 6000),6 ℃等不同胁迫明显诱导,且在大多数时间点下的表达差异显著(图 4)。氯化钠胁迫下,JrGSTU23在根和叶中的表达趋势相似,均随着胁迫时间的延长表达增高,在胁迫24 h达最大表达水平,分别为2.49和4.11。但除3 h外,其他处理点该基因在根中的表达水平高于在叶中的表达。PEG 6000模拟干旱胁迫3~12 h,JrGSTU23在叶中的表达被抑制,24 h被诱导为1.76;在根中的表达趋势与氯化钠胁迫相似,随着胁迫时间延长而增大,且在24 h达最高水平,但表达量低于氯化钠胁迫。表明JrGSTU23应对氯化钠和干旱胁迫的响应机制可能相似,但JrGSTU23对盐胁迫可能更为敏感。低温胁迫下,JrGSTU23在叶中的表达在6 h(1.22)和24 h(2.30)出现2个高峰;在根中,其表达趋势与氯化钠和干旱胁迫相似,随着胁迫时间延长而升高,在24 h达最大值(3.06)。
2.1. JrGSTU23全长cDNA序列分析

2.2. JrGSTU23启动子分析
2.3. 外源激素胁迫对JrGSTU23表达的影响

2.4. 不同非生物胁迫对JrGSTU23基因表达的影响
-
GSTs是植物响应逆境的重要基因,在植物解毒等方面具有重要作用。其中,含有Tau保守结构域的GST亚家族基因,参与植物众多的逆境响应。核桃作为中国西北地区扶贫攻坚项目的重要经济树种,在推动区域经济发展上具有重要作用。核桃产业的健康快速发展与核桃产量和质量息息相关。但气候等环境因子严重制约了中国核桃产业的发展,因此,选育抗逆优良核桃品种,掌握核桃抗逆适应机制,对深入了解核桃的适应性具有重要指导作用。本研究从‘香玲’核桃中克隆获得1条Tau家族的GST基因(JrGSTU23),经进化分析发现该基因与来自水稻、香蕉、毛果杨、大豆Glycine max等物种的Tau家族基因具有较近的亲缘关系,推测其可能与这些蛋白具有相似或相近的功能。如,水稻Osgstu4和Osgstu3能迅速被抗氧化剂和过氧化氢诱导,表明Osgstu4和Osgstu3的应答反应涉及氧化还原反应[12]。大豆GmGSTU2-2是严格的渗透胁迫型响应基因,参与植物胁迫反应中的催化和调节功能网络[13]。毛果杨的GSTU16和GSTU45能被三硝基甲苯(2, 4, 6-trinitrotoluene)诱导[14]。因此,推测JrGSTU23与逆境应答具有重要关系。
顺式作用元件一般由5~20个碱基对组成,是同一DNA分子中具有转录调节功能的特异DNA序列[15]。海蓬子Salicornia brachiata的一个Tau类GST基因的上游1 023 bp启动子包含有非生物胁迫响应相关的ABA响应元件(ABRE),干旱响应基因rd22识别位点(MYB),结节特意表达元件(NOD),光响应表达元件(GATA),光调控表达元件(GT1)及激素、病害和损伤等相关的顺式作用元件,参与了该基因响应氯化钠和渗透胁迫的表达调控[16]。玉米Zea mays的ZmCIPK10和ZmZIP71基因启动子序列中含有ABA,SA,赤霉素(GA),低温等相关的顺式作用元件,在氯化钠、干旱、低温胁迫下,ZmCIPK10和ZmZIP71的表达量上升,表明其参与了玉米的逆境响应[17-18]。本研究发现:JrGSTU23基因启动子含有丰富的顺式作用元件,如玉米素代谢、种子调控、分生组织激活、胚乳表达以及干旱胁迫、热胁迫、防卫、MeJA和SA等响应相关的元件(表 2)。由此可推测,JrGSTU23可能参与植物生长发育及逆境响应过程,具有深入研究的价值。
GSTs基因响应逆境具有组织表达特异性。本研究发现的JrGSTU23基因在不同激素(SA,MeJA,ABA)及不同逆境(氯化钠、干旱、低温)下在根和叶中能被不同程度地诱导表达,体现了一定的组织表达特异性和逆境响应特异性。这与其他物种的Tau家族GST基因的逆境响应表达具有一定的相似性。如从香蕉克隆获得的5个GST基因(MaGSTU1,MaGSTU2,MaGSTU3,MaGSTF1,MaGSTL1)的表达具有组织特异性,在盐、干旱、冷等胁迫下Tau亚家族的MaGSTU1,MaGSTU2,MaGSTU3的表达受盐、干旱、冷诱导更为明显,而MaGSTF1和MaGSTL1更受信号分子影响[19],预测这些GST基因对不同逆境的响应功能具有差异。盐胁迫下,番茄Solanum lycopersicum的SlGSTU23和SlGSTU26基因在叶中被上调表达[20],推测这些GST基因在不同逆境下的具体功能可能不同。JrGSTTau1在低温胁迫下也表现出根、叶表达差异,过表达提高了植株的抗寒能力[8]。可见,通过分析基因响应不同逆境的转录水平,可以推测其在逆境响应中可能的生物学功能。JrGSTU23能不同程度地响应激素及非生物胁迫,表明其参与了核桃的逆境响应调控。后续研究将通过在植株中过量表达全面分析JrGSTU23基因的抗逆响应功能。
 DownLoad:
DownLoad: