






-
种群是物种存在、发展进化以及自然演替的基本单元。种群的结构及动态是种群生态学重要研究内容[1],既反映种群内个体的配置状况、数量动态及种群的未来发展与演变趋势[2−3],又反映种群与环境的关系[4]。研究种群结构及动态的基本方法是种群统计,年龄结构、静态生命表、存活曲线、生存分析、动态指数和时间序列分析等是种群统计学的核心内容。年龄结构是反映种群结构现状的基本指标,也是植物对立地条件适应能力的具体体现。种群目前的存活状态及植物与环境之间的适合度可通过种群静态生命表与其构建的存活曲线和生存分析来反映[5−6]。种群经历的生态学过程及预测种群发展趋势可通过种群动态量化分析和时间序列预测来阐明[7]。开展种群结构及动态特征研究有助于揭示种群的生存现状和发展方向,对于揭示群落演替规律和指导森林生态系统保护修复工作具有重要意义。火山喷发为研究植被演替尤其是原生演替提供了难得的条件[8]。五大连池火山处于小兴安岭和松嫩平原的交错地带,由距今时间跨度300 a至200万 a的2座新期火山和12座老期火山组成[9]。五大连池火山区生态条件独特复杂,由熔岩裸地到演化中的不同生境内可见低等植物和高等植物(演替中的不同植被生态系列)[10]。五大连池火山群具有保存完好的内陆单成因火山地貌,原生而完整的植被生态演替过程,是研究火山干扰、种群的生态学过程和植被演替等诸多科学问题难得的科研基地[11]。近年来对火山生境生态学的研究集中在物种多样性、土壤养分、生态价值等方面,但对火山生境种群的结构和动态的研究仅见于长白山区。植被动态是植被生态学研究的主要内容之一,而植被演替是植被动态研究的核心内容[12]。群落在各阶段通过优势种群的数量增长和消亡来表达群落的演变特征,种群动态则是揭示演替过程的关键[13]。蒙古栎Quercus mongolica具有相对耐贫瘠、耐寒、耐干旱、萌生能力强等特性,广泛分布在东北地区阳坡的低海拔地段,形成典型的蒙古栎林,对中国北方森林群落的演替与发展具有重要意义[14]。蒙古栎是五大连池老期火山南坡乔木层中的优势种群[15]。本研究设置4座老期火山(东焦得布山、小孤山、尾山和南格拉球山)南坡为研究样地,以蒙古栎种群为研究对象,采用种群统计法分析种群的结构与动态特征,揭示种群的生存状态及发展方向,为五大连池火山森林植被的演替、恢复与可持续发展提供科学依据。
-
样点设在黑龙江黑河(五大连池)国家森林生态系统定位观测研究站(48°30′~48°50′N,126°00′~126°45′E),是中国第1个以火山自然环境及生态系统为保护对象的自然保护区[16]。研究区属温带大陆性季风气候,夏季温暖短促,冬季严寒漫长,温度与降水四季变化明显,年均气温为−0.50 ℃,无霜期为121 d,年均降水量为476.33 mm,土壤主要为火山石质土、火山灰土。主要森林植被类型为温带落叶阔叶混交林,乔木层优势植物有蒙古栎、黑桦Betula davurica、紫椴Tilia amurensis、山杨Populus davidiana、白桦Betula platyphylla等,灌木层有珍珠梅Sorbaria sorbifolia、山刺玫Rosa davurica、榛Corylus heterophylla,草本层有万年蒿Artemisia vestita、乌苏里苔草Carex ussuriensis等[17]。
-
本研究区的4座老期火山景观已经17~80 万 a的演化,过去曾是针阔混交林,由于人为对针叶树的间伐,形成现阶段主要分布在火山锥山头附近半原生状态的郁闭度较大的阔叶林[18]。于2020年10—11月,在4座山南坡受人为干扰少的山顶附近蒙古栎种群,从高海拔向低海拔方向设置20 m × 20 m相邻样方各3个,样地信息见表1和参考文献[14, 19]。利用生长锥对样方内胸径≥2.5 cm的植株,在胸高1.3 m处锥取1~2个年轮条,保留过髓心的样品,并记录胸径、树高和幼苗更新情况。对于10 a以下的小龄树从最末端枝条往主干溯源确定树龄(温带的树木通常1 a内只形成1个生长轮,且每年生长结束与来年生长之间会形成1个明显的节痕,将1节枝条划分为1龄,2节枝条划分为2龄,以此类推)[20]。将野外采取的年轮样芯,带回实验室固定在木槽内自然风干,待木芯完全风干后,用乳白胶固定在木槽上。固定后的芯样用砂粒由粗到细的砂纸打磨抛光,直到年轮清晰可见。用LINTAB年轮分析仪测年[21]。
研究区 地理位置 喷发时间/万a 海拔/m 坡向 坡度/(°) 胸径/ cm 郁闭度 乔木树种 东焦德布山 48°39′13ʺN, 126°16ʹ30ʺE 17~19 531.9 南 35~40 14.05±3.23 0.65 247蒙+6色+3黑 小孤山 48°40′45ʺN, 126°22ʹ06ʺE 28~34 453.5 南 37~42 13.61±3.21 0.70 309蒙+6色+4黑+1白 尾山 48°47′23ʺN, 126°15ʹ26ʺE 40~50 516.6 南 30~35 14.95±5.12 0.60 187蒙+31色+7黑+5黄 南格拉球山 48°44′13ʺN, 126°00ʹ46ʺE 70~80 596.9 南 45~50 11.74±3.11 0.70 324蒙+20色+2黑+2黄 说明:胸径数值为平均值±标准差;蒙指蒙古栎;色指色木槭Acer pictum;黑指黑桦;白指白桦;黄指黄檗Phellodendron amurense;乔木树种中的数值表示株数 Table 1. Basic information of plots
-
以10 a为1个龄级[22],1~10 a为龄级Ⅰ,11~20 a为龄级Ⅱ,以此类推龄级Ⅲ、Ⅵ、Ⅴ、Ⅳ、Ⅶ、
$ \cdots $ ,以龄级为纵坐标,对应个体数为横坐标,绘制蒙古栎种群的年龄结构图。 -
种群统计核心是编制静态生命表。静态生命表不是对某一种群全部生活史的追踪,而是反映多个世代重叠的年龄动态历程中的一个特定时间[23],由于本研究所调查的蒙古栎是自然种群,利用不同龄级结构的个体数来研究特定时间段的种群动态,难免会因系统抽样误差与数学假设不符,出现死亡率为负的情况,对此有学者采用匀滑技术对统计数据进行处理[24]。本研究采用方程拟合法进行匀滑,以龄级为自变量,存活株数为因变量拟合方程,并编制静态生命表,计算公式参考文献[23-24]。
-
存活曲线是借助于存活个体数量描述特定年龄死亡率,反映种群个体在各年龄段存活状态的曲线。本研究经过SPSS 20.0拟合存活数和龄级关系的指数函数和幂函数的模型,依据决定系数(R2)、F检验值来判定模型拟合效果,依据HETT和LOUCKS的数学模型[25],如果指数方程Nx=N0e–bx拟合效果好,存活曲线为Deevey Ⅱ型,若幂函数Nx=N0x–b拟合效果好,存活曲线为Deevey Ⅲ型。其中:Nx为lnlx数值,x为树龄,N0、b为常数。以龄级为横坐标,标准化存活数对数lnlx为纵坐标,绘制蒙古栎种群存活曲线。
-
基于静态生命表和生存分析理论,应用生存分析中的4个函数,即种群生存率函数S(i)、累积死亡率函数F(i)、死亡密度函数f(ti)和危险率函数λ(ti)来分析蒙古栎种群生存状况,计算公式参考文献[26],其中:t为时间,i为时间段龄级。
-
种群数量化分析是根据种群内两相邻龄级间个体数量变化动态指数(Vn)、忽略外部干扰时种群的数量变化动态指数(Vpi)及考虑外部干扰时种群的数量变化动态指数(V'pi)客观精确地定量比较和评价种群结构动态,计算公式参考文献[24, 27],其中:n为时间段龄级、pi为种群的所有龄级时间段,Pmax为风险概率极大值。
-
采用时间序列分析的一次移动平均法对蒙古栎的种群龄级结构进行模拟和预测,计算公式参考文献[28],分别对种群经历2、4和6个龄级的时间后,种群未来发展趋势进行预测。
-
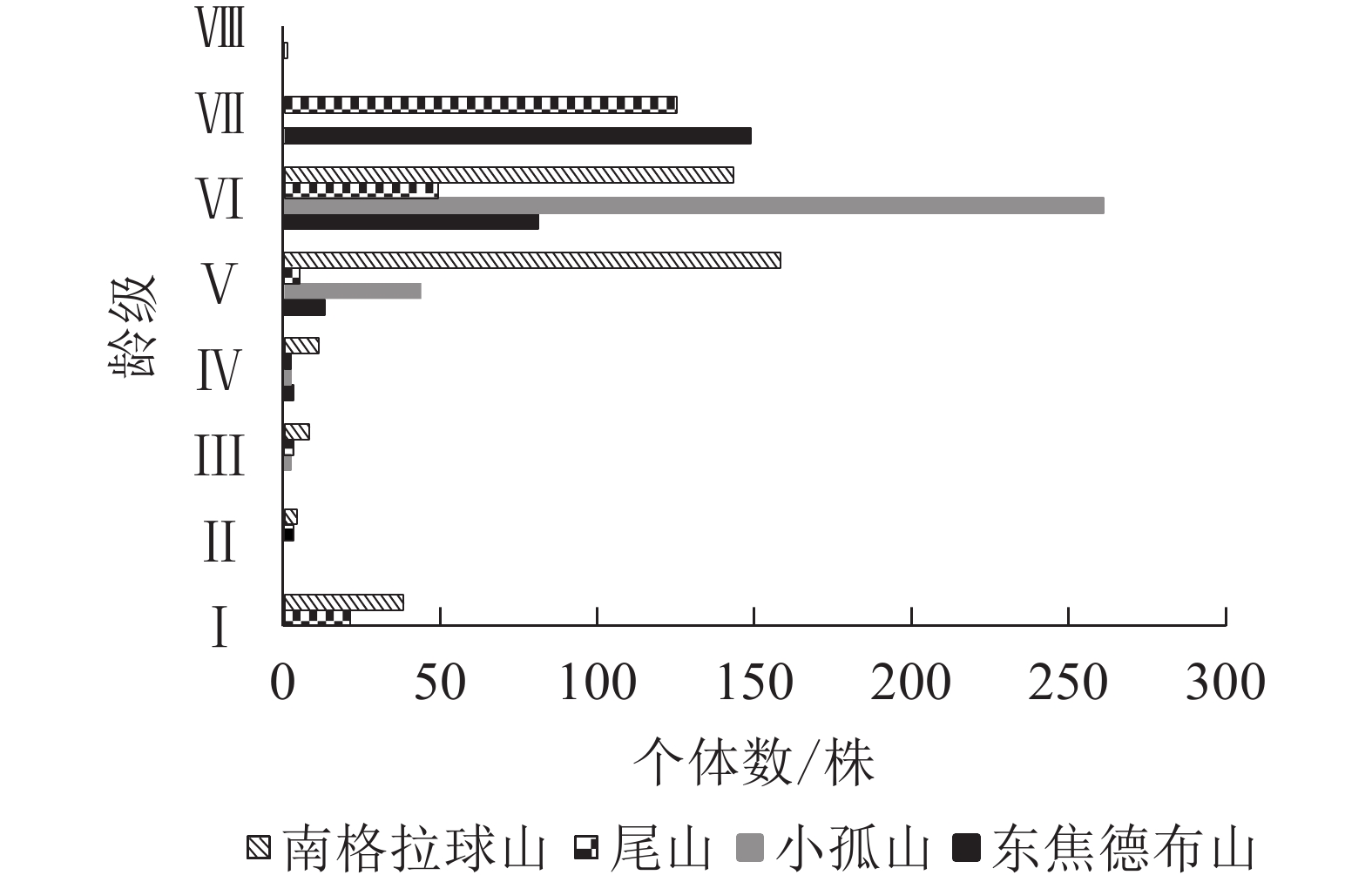
本研究调查蒙古栎共计1 195株(图1),最大胸径为26.3 cm,最大树龄为76 a。东焦德布山共8个龄级275株;尾山共7个龄级208株;小孤山和南格拉球山各有6个龄级,分别为350和362株。4座火山蒙古栎种群年龄结构龄级Ⅰ的个体数占10.10%~11.71%;龄级Ⅱ~Ⅳ的个体数占1.09%~6.35%;龄级Ⅴ~Ⅵ或龄级Ⅵ~Ⅶ的个体数高达83.15%以上,其中,小孤山和南格拉球山的龄级Ⅴ~Ⅵ的个体数分别占87.14%和83.15%,东焦德布山和尾山的龄级Ⅵ~Ⅶ的个体数分别占83.64%和83.65%;均处于成熟树阶段,种群的稳定性都较好,表现为稳定型种群。
Figure 1. Age structure of Q. mongolica populations in Wudalianchi Volcanos
-
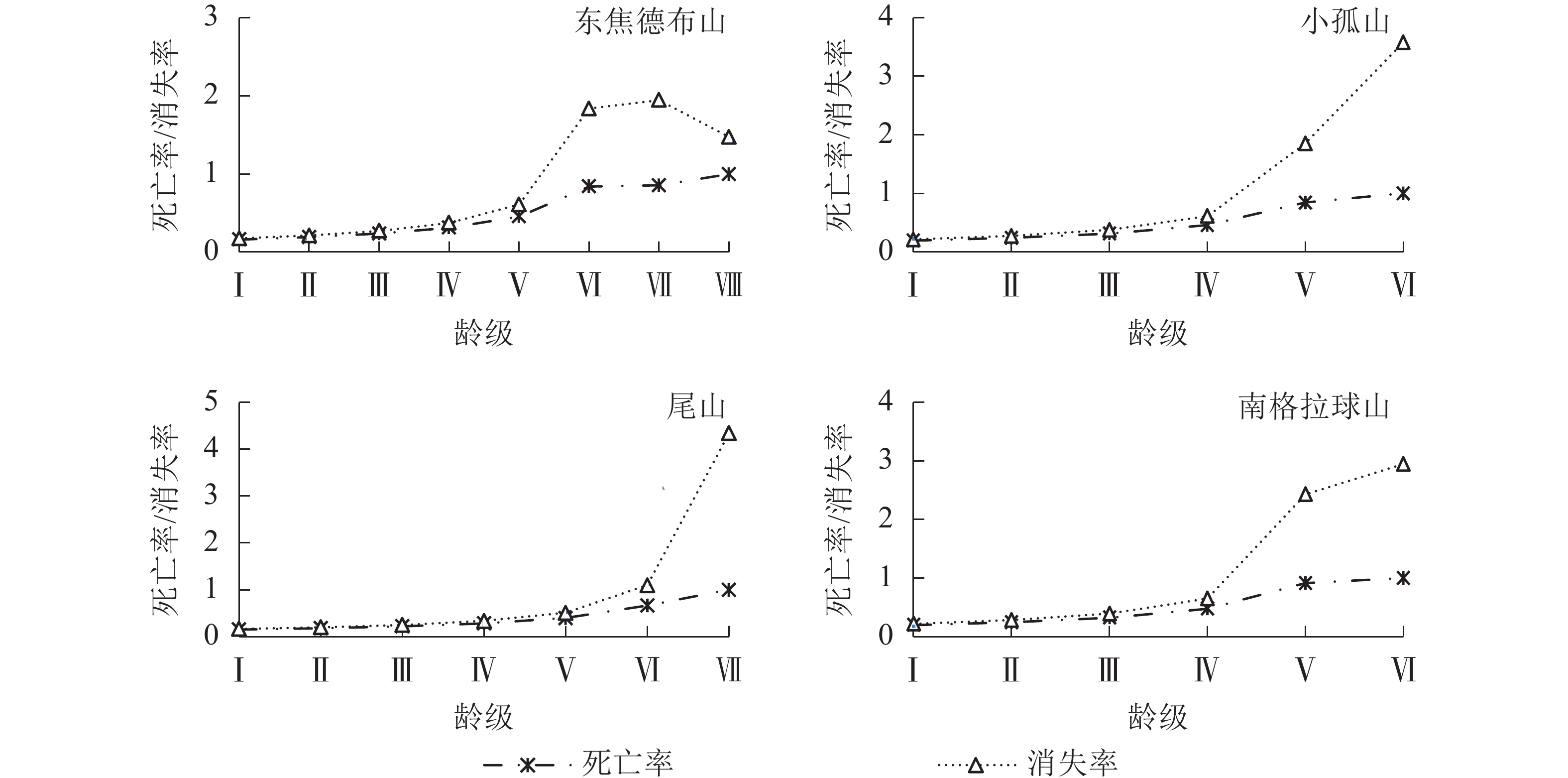
静态生命表分析结果显示(表2):4座火山蒙古栎种群的标准化存活数(lx)、个体的期望寿命(ex)和存活率(Sx)均随年龄增加而逐渐减小,这与其生物学特性一致;死亡率(qx)和消失率(Kx)均随龄级增加而上升,且龄级Ⅵ和Ⅶ显著高于龄级Ⅰ~Ⅴ。
研究区 龄级 Ax ax lx lnlx dx qx Lx Tx ex Kx Sx 东焦德布山 Ⅰ 28 229 1000 6.908 162 0.162 919 3111 3.111 0.176 0.838 Ⅱ 0 192 838 6.732 162 0.193 758 2192 2.614 0.214 0.807 Ⅲ 0 155 677 6.517 162 0.239 596 1434 2.119 0.273 0.761 Ⅳ 3 118 515 6.245 162 0.314 434 838 1.626 0.376 0.686 Ⅴ 13 81 354 5.868 162 0.457 273 404 1.142 0.610 0.543 Ⅵ 81 44 192 5.258 162 0.841 111 131 0.682 1.838 0.159 Ⅶ 149 7 31 3.420 26 0.857 17 19 0.622 1.946 0.143 Ⅷ 1 1 4 1.474 4 1.000 2 2 0.458 1.474 0.000 小孤山 Ⅰ 41 446 1000 6.908 193 0.193 904 2608 2.608 0.214 0.807 Ⅱ 0 360 807 6.694 193 0.239 711 1704 2.111 0.273 0.761 Ⅲ 2 274 614 6.421 193 0.314 518 993 1.616 0.377 0.686 Ⅳ 2 188 422 6.044 193 0.457 325 475 1.127 0.611 0.543 Ⅴ 44 102 229 5.432 193 0.843 132 132 0.577 1.852 0.157 Ⅵ 261 16 36 3.580 36 1.000 18 18 0.502 3.580 0.000 尾山 Ⅰ 21 260 1000 6.908 154 0.154 923 3269 3.269 0.167 0.846 Ⅱ 3 220 846 6.741 154 0.182 769 2346 2.773 0.201 0.818 Ⅲ 3 180 692 6.540 154 0.222 615 1577 2.278 0.251 0.778 Ⅳ 2 140 538 6.289 154 0.286 462 962 1.787 0.336 0.714 Ⅴ 5 100 385 5.952 154 0.400 308 500 1.300 0.511 0.600 Ⅵ 49 60 231 5.441 154 0.667 154 192 0.832 1.099 0.333 Ⅶ 125 20 77 4.343 77 1.000 38 38 0.494 4.343 0.000 南格拉球山 Ⅰ 38 158 1000 6.908 196 0.196 902 2557 2.557 0.218 0.804 Ⅱ 4 127 804 6.689 196 0.244 706 1655 2.059 0.280 0.756 Ⅲ 8 96 608 6.410 196 0.323 509 949 1.562 0.390 0.677 Ⅳ 11 65 411 6.020 196 0.477 313 440 1.070 0.648 0.523 Ⅴ 158 34 215 5.372 196 0.912 117 127 0.590 2.428 0.088 Ⅵ 143 3 19 2.944 19 1.000 9 9 0.474 2.944 0.000 说明:Ax为x龄内个体数;ax为Ax匀滑后的个体数;lx为x龄标准化存活数;lnlx为lx的对数;dx为从x龄到x+1龄间的标准化死亡 数;qx为从x龄到x+1龄间的死亡率;Lx为x龄到x+1龄间的平均存活数;Tx为超过x 龄的个体总数;ex为x龄个体的生 命期望值;Kx为消失率;Sx为x龄开始时个体存活率 Table 2. Static life table of Q. mongolica populations in Wudalianchi Volcanos
-
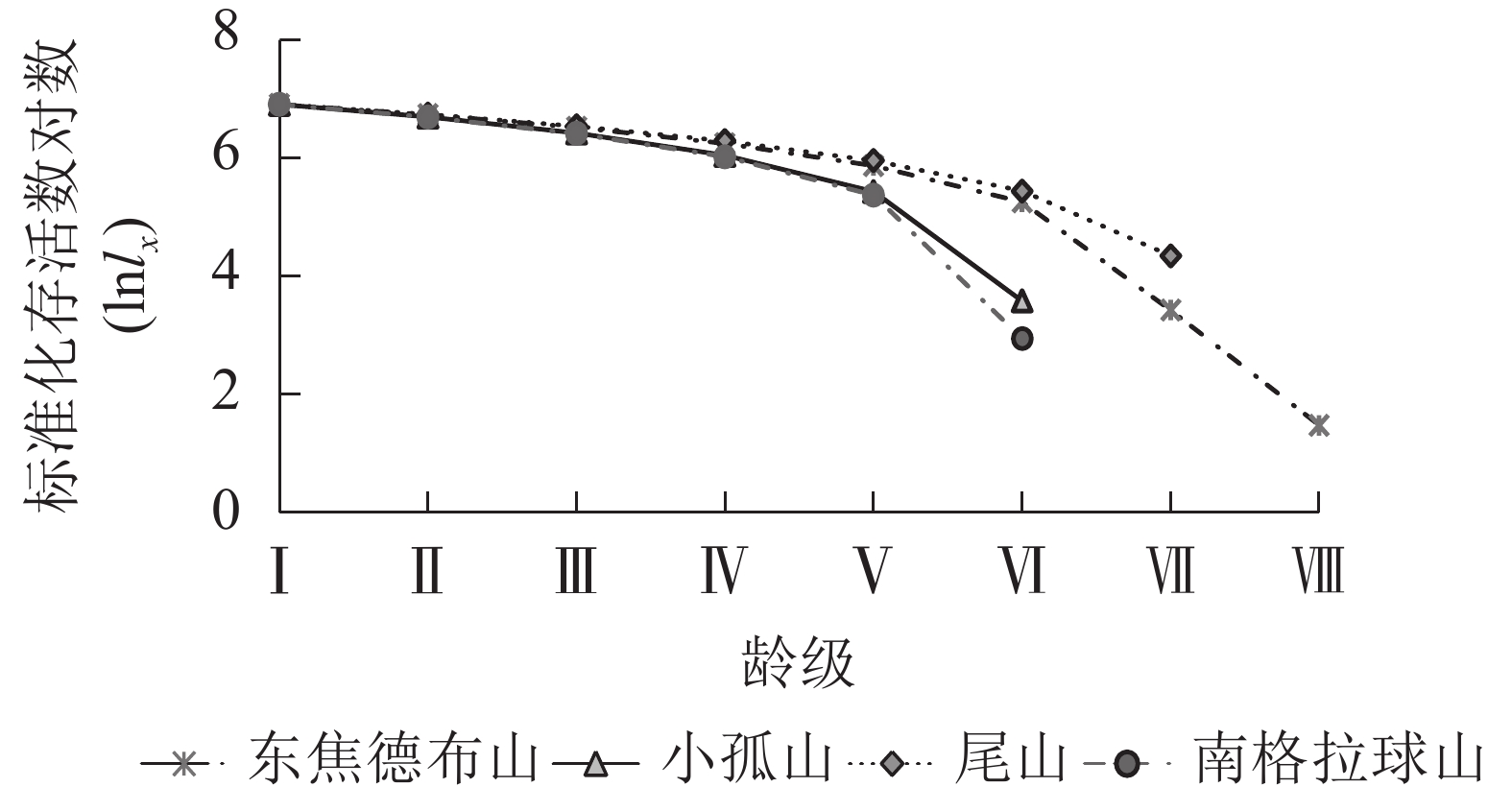
由图2可知:4座火山的蒙古栎种群进入龄级Ⅴ后下降趋势开始加剧,这与死亡率和消失率进入龄级Ⅴ后的快速上升趋势相一致(图3),且个体期望寿命(ex)在龄级Ⅴ后下降趋势也更显著,说明蒙古栎在龄级Ⅳ前的生存能力较好,从龄级Ⅴ起个体生存能力开始快速下降。由表3可知:4座火山指数模型的R2和F均大于幂函数模型,P值更小,且均达到了显著水平(P<0.05),说明指数模型的拟合效果更好;4座火山蒙古栎种群的存活曲线均属于Deevey Ⅱ型,表明蒙古栎种群随着年龄的增长,存活个体数逐渐减少,这与静态生命表的分析结果基本相同。
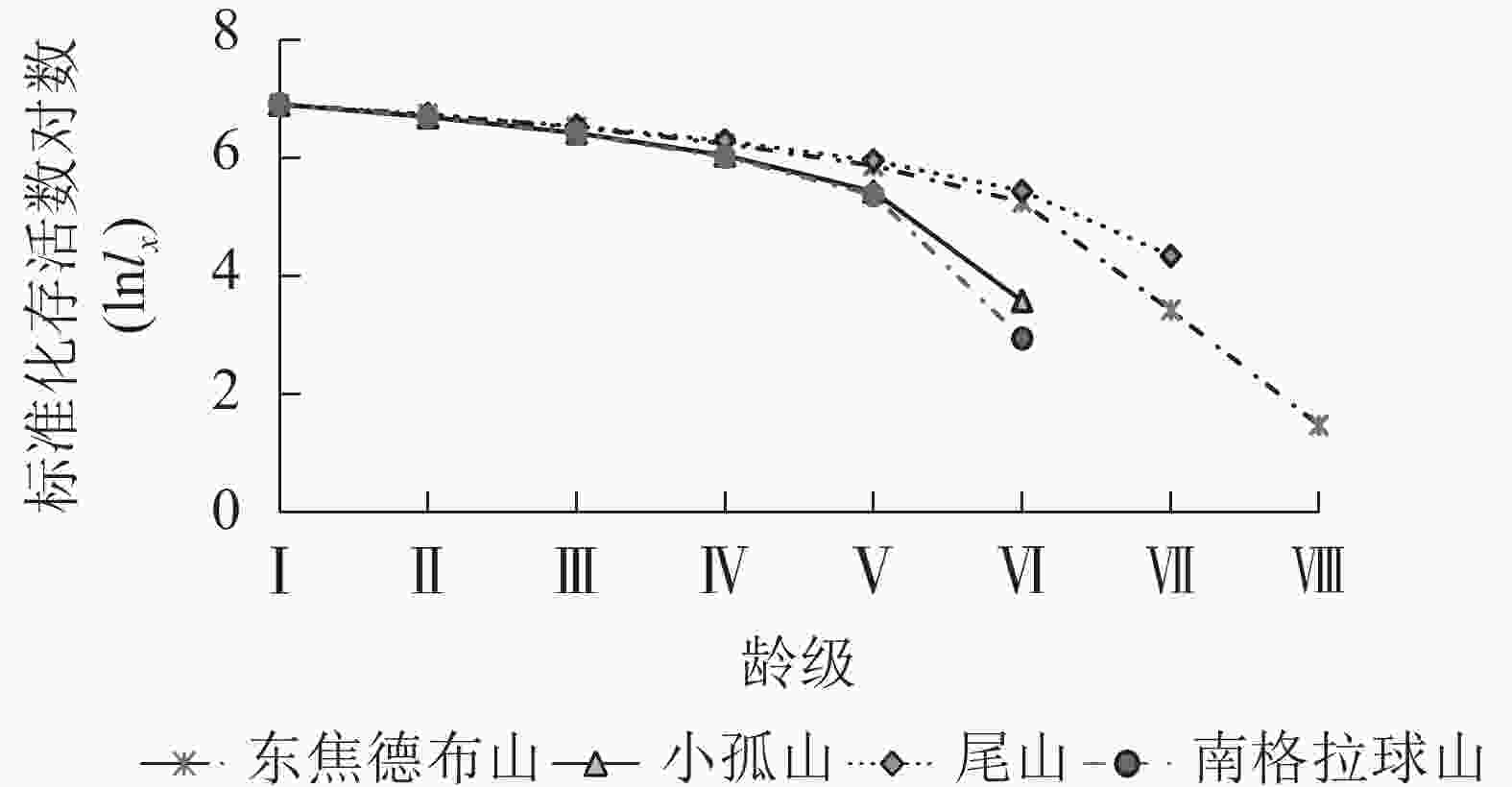
Figure 2. Survival curve of Q. mongolica populations in Wudalianchi Volcanos

Figure 3. Mortality and vanish rate of Q. mongolica populations in Wudalianchi Volcanos
研究区 方程 R2 F P 类型 东焦德布山 Nx=10.725e−0.177x 0.672 12.311 0.013 Deevey Ⅱ Nx=9.433x−0.506 0.450 4.908 0.069 小孤山 Nx=8.505e−0.114x 0.775 12.350 0.025 Deevey Ⅱ Nx=7.739x−0.276 0.561 5.119 0.086 尾山 Nx=7.845e−0.068x 0.835 25.235 0.004 Deevey Ⅱ Nx=7.505x−0.188 0.635 8.709 0.032 南格拉球山 Nx=9.090e−0.143x 0.692 8.997 0.040 Deevey Ⅱ Nx=8.024x−0.342 0.498 3.972 0.117 Table 3. Test models of survival curves of Q. mongolica populations in Wudalianchi Volcanos
-
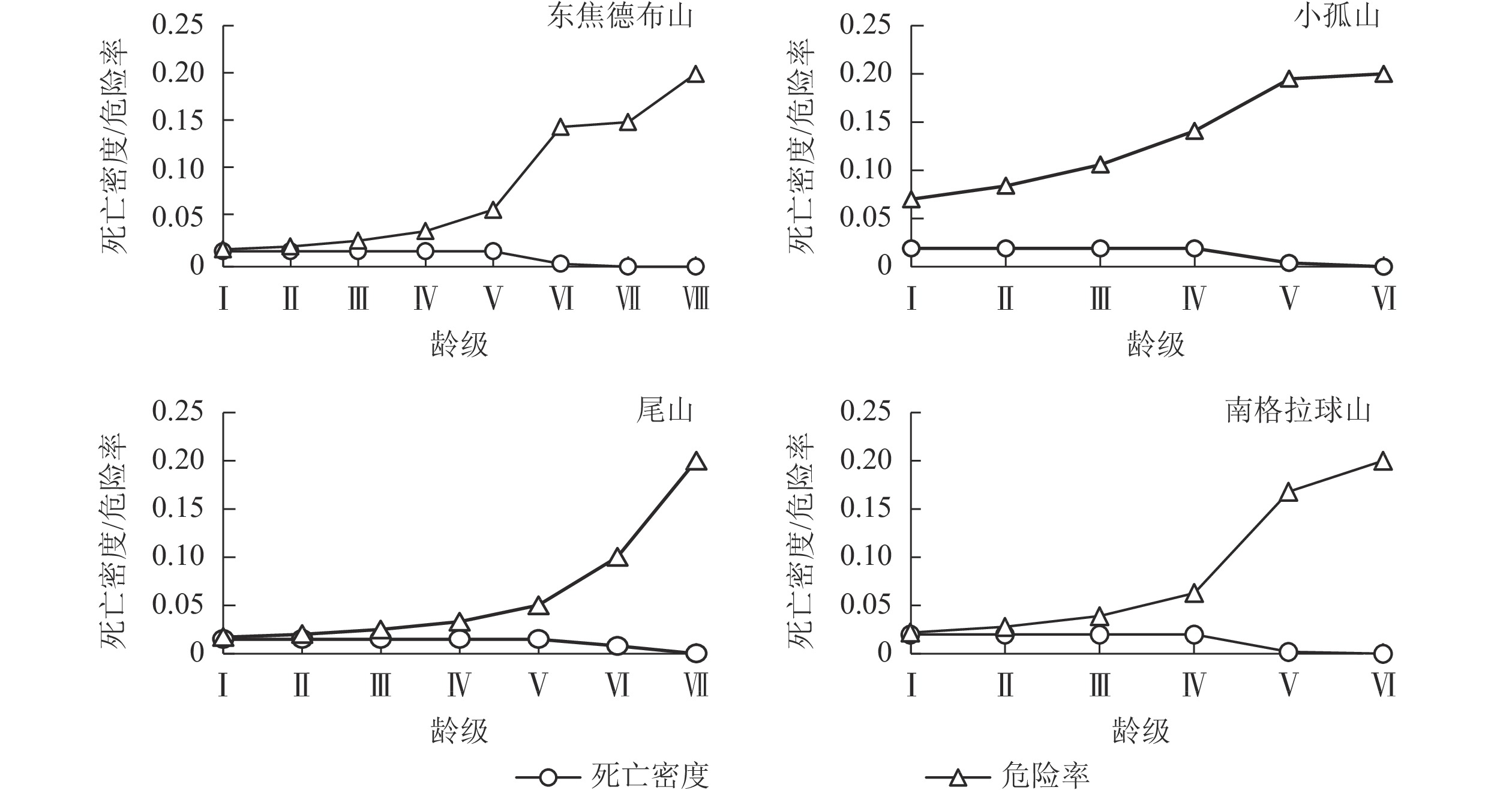
图4和图5显示:生存率和累计死亡率在龄级Ⅰ~Ⅴ分别呈单调的下降和上升趋势,到龄级Ⅵ时,种群的生存率小于1%,而累计死亡率大于99%,种群在此龄级之后开始进入生理衰退期;死亡密度在龄级Ⅰ~Ⅴ都平缓保持在0.02以下,而危险率整体逐渐上升,到龄级Ⅵ死亡密度降为0,危险率加剧上升,表明随着个体逐渐进入生理衰退期,其受环境影响更明显。综上,4座火山蒙古栎种群具有前期逐渐减少、中期趋于稳定的特点,这与种群的存活曲线、死亡率曲线和消失率曲线结果一致。

Figure 4. Survival rate and cumulative mortality rate of Q. mongolica populations in Wudalianchi Volcano
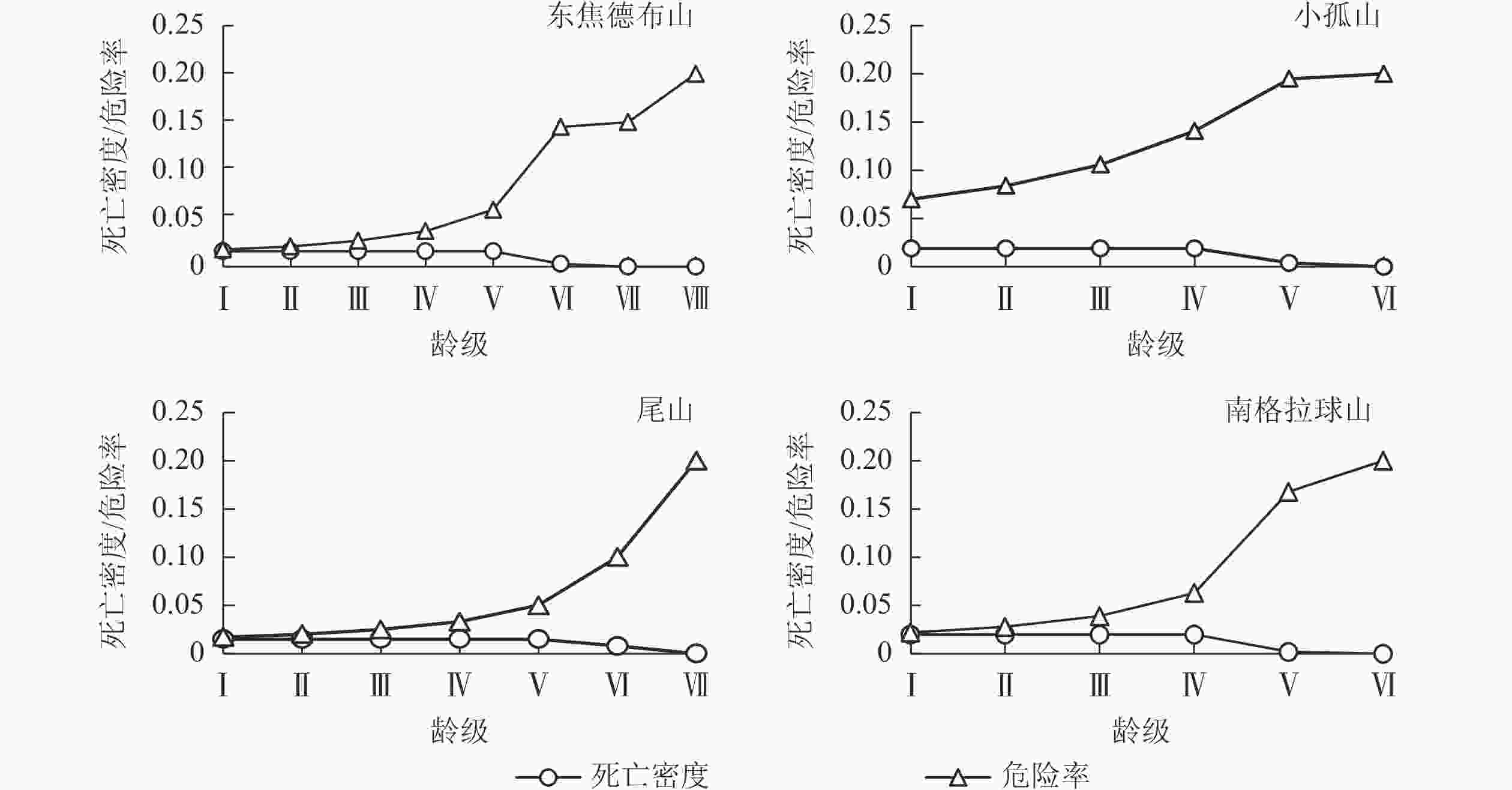
Figure 5. Mortality density and hazard rate of Q. mongolica populations in Wudalianchi Volcano
-
由表4可见:东焦德布山和南格拉球山种群的数量变化动态指数分别呈明显的波动,东焦德布山种群的数量变化动态指数V1和V7大于0、V3~V6小于0,南格拉球山种群动态指数V1和V5大于0、V2~V4小于0,表明这2座山蒙古栎种群整体上表现为“增长—衰退—增长”的结构动态关系。小孤山和尾山均呈现种群的数量变化动态指数V1~V3呈明显的波动,V4~V7都小于0,表明小狐山和尾山蒙古栎种群开始表现为衰退的结构动态关系。不考虑外部干扰的情况下东焦德布山、小孤山和南格拉球山种群总体数量变化动态指数(Vpi)均为正值,表明忽略外部干扰时这3座山蒙古栎种群表现为增长型种群;而考虑随机干扰的情况下种群总体数量变化动态指数(V'pi)分别为0、0和0.66,等于0或趋近于0,随机干扰所承担的风险概率极大值(Pmax)分别为0、0.07和0.04,说明受外界环境干扰时,这3座火山蒙古栎种群表现为稳定型,且对外界随机干扰均具有较高的敏感性。尾山Vpi和V'pi均为负值,Pmax为0,表明在有或无随机干扰时尾山蒙古栎种群都表现为衰退型,且对外界随机干扰敏感度较高。
研究区 种群动态
指数级Vn动态指
数值/%研究区 种群动态
指数级Vn动态指
数值/%研究区 种群动态
指数级Vn动态指
数值/%研究区 种群动态
指数级Vn动态指
数值/%东焦德布山 V1 100.00 尾山 V1 85.71 小孤山 V1 100.00 南格拉球山 V1 89.47 V2 0.00 V2 0.00 V2 −100.00 V2 −50.00 V3 −100.00 V3 33.33 V3 0.00 V3 −27.27 V4 −76.92 V4 −60.00 V4 −95.45 V4 −93.04 V5 −83.95 V5 −89.80 V5 −83.14 V5 9.49 V6 −45.64 V6 −60.80 Vpi 2.82 Vpi 15.79 V7 99.33 Vpi −19.86 V'pi 0.00 V'pi 0.66 Vpi 45.92 V'pi −1.42 Pmax 0.07 Pmax 0.04 V'pi 0.00 Pmax 0.00 Pmax 0.00 说明:Vn是n到n+1龄级之间个体数量变化动态指数;Vpi是忽略外部干扰时种群的数量变化动态指数;V'pi是考虑外部干扰时种群的 数量变化动态指数 Table 4. Dynamic index of Q. mongolica populations in Wudalianchi Volcanos
-
时间序列预测表明(表5):在未来经过2、4、6个龄级后,4座火山蒙古栎种群个体的数量整体上均呈减少趋势。在未来经过2个龄级后,4座火山蒙古栎种群仅有Ⅱ龄级的个体数增加,其他龄级的个体数均减少;在未来经过4个龄级后,4座火山蒙古栎种群仅有Ⅳ龄级的个体数增加,其他龄级的个体数均减少;在未来经过6个龄级后,4座火山蒙古栎种群各龄级的个体数量均减少。由此可知,种群低龄个体的数量较少,中、成年个体数会随着林木生长而得不到补充,种群结构趋于不合理。
研究区 龄级 实际存活数 M2 M4 M6 研究区 龄级 实际存活数 M2 M4 M6 东焦德布山 Ⅰ 28 尾山 Ⅰ 21 Ⅱ 0 14 Ⅱ 3 12 Ⅲ 0 0 Ⅲ 3 3 Ⅳ 3 2 8 Ⅳ 2 3 7 Ⅴ 13 8 4 Ⅴ 5 4 3 Ⅵ 81 47 24 21 Ⅵ 49 27 15 14 Ⅶ 149 115 62 41 Ⅶ 125 87 45 31 Ⅷ 1 75 61 41 南格拉球山 Ⅰ 38 小孤山 Ⅰ 41 Ⅱ 4 21 Ⅱ 0 21 Ⅲ 8 6 Ⅲ 2 1 Ⅳ 11 10 15 Ⅳ 2 2 11 Ⅴ 158 85 45 Ⅴ 44 23 12 Ⅵ 143 151 80 60 Ⅵ 261 153 77 58 说明:M2表示未来经过2个龄级时各龄级的种群个体存活数;M4表示未来经过4个龄级时各龄级的种群个体存活数;M6表示未来经 过6个龄级时各龄级的种群个体存活数 Table 5. Time sequence prediction of number dynamics of Q. mongolica populations in Wudalianchi Volcanos
-
植物种群的生物学特征与环境因素共同作用的结果通过种群龄级结构体现[29]。本研究发现:4座火山蒙古栎种群在龄级Ⅰ~Ⅳ时,种群个体数量随着龄级的增加而减少,80%以上个体都处于龄级Ⅴ~Ⅵ的成熟树阶段,种群结构总体呈稳定型,但种群在小树阶段个体数量显著减少,导致老龄树个体数量的发展潜力较小,种群有向衰退转化的趋势。蒙古栎喜光不耐荫的习性导致随着幼树的生长,其个体数量大幅度减少,这种高死亡率现象主要是由幼树阶段有限的光照资源所致[30]。本研究区火山锥地势高,坡度大,多碎屑岩,风化过程和成土过程都较缓慢,土壤上层厚30~50 cm,疏松且夹有1~2 cm大的砾石块,下层砾石块增大增多,pH 6.6。发育着不完善的粗骨土的干燥阳坡,土层浅薄且石粒较多,腐植质含量甚微,显著影响了蒙古栎幼树的正常生长。本研究区蒙古栎林的郁闭度高于0.60,在此生境中,空间和光照不足进一步加剧了小龄级树木死亡,导致种群幼龄树很难向中龄树和成龄树阶段转化,这在一定程度上制约了蒙古栎种群的自然更新。本研究的结果与解婷婷等[30]、陈国鹏等[24]、赵阳等[31]的研究结果一致,即幼树阶段的高死亡率是种群更新和发展的瓶颈。
-
种群个体在各龄级的存活状况和种群的发展趋势可以通过种群静态生命表及存活曲线来判断[32],死亡率和消失率反映了种群数量随龄级增加的动态变化[33]。本研究4座火山蒙古栎种群的存活曲线都为Deevey Ⅱ型,表明当前各种群总体处于稳定状态。种群个体数量的变化直接受种群更新情况和个体存活情况的影响,蒙古栎种群幼龄个体相对不足,随龄级增大种群死亡率和消失率都呈上升趋势,且在进入龄级Ⅴ后快速上升,这可能因为火山区的独特生态条件和老龄个体的生理生长极限导致老龄个体对环境资源的利用能力下降,致使老龄个体的发展遇到了瓶颈,目前依靠数量较为充足的中龄个体维持种群的相对稳定,但是长期来看种群存在偏离稳定状态的风险。在野外调查时发现:较多蒙古栎存在萌发生长,更新个体主要源于母树根部,萌生蒙古栎的寿命期远低于实生个体,个体数量的扩繁和种群范围的扩大均受到较大限制[34]。种群结构与动态更进一步直观地反映可以通过4个生存分析函数来表达。本研究生存分析表明:4座火山蒙古栎种群到龄级Ⅵ,种群的累计死亡率大于99%,而生存率小于1%,说明种群在此龄级之后开始进入生理衰退期,同时死亡密度降为0,危险率加剧上升,也表明蒙古栎种群目前相对稳定,种群后期有衰退的趋势。种群数量变化动态反映种群个体生存能力与环境的相互作用[35],研究种群未来年龄结构和发展趋势的有力手段是时间序列分析[36]。4座火山蒙古栎种群动态量化分析结果显示:东焦德布山、小孤山和南格拉球山在忽略外部干扰时种群表现为增长型,在受外界环境干扰时为稳定型,且对外界随机干扰均具有较高的敏感性;尾山蒙古栎种群在2种情况下都表现为衰退型,且对外界随机干扰敏感度较高,这可能是因为只有尾山的南坡山麓地带大面积土石被移走,储藏的资源更少有关。时间序列分析结果显示4座火山蒙古栎种群未来的发展趋于衰退,主要原因可能是缺乏幼龄个体的补充。
-
植物种群的发展是复杂且漫长的过程,不仅有物种内和物种间的竞争,同时还伴有物种和环境的适应过程[37]。蒙古栎种群的年龄结构与动态特征与其繁殖策略和有限的环境资源密切相关。本研究调查发现:4座火山蒙古栎种群样地内幼龄个体几乎都为根蘖苗,因此蒙古栎种群的更新很难通过有性生殖的方式完成。根蘖繁殖所导致的聚集分布模式使蒙古栎种群大多数幼龄个体因上层林冠的庇萌而死亡,难以成长为低龄和中龄个体。蒙古栎由低龄级向中高龄级生长的过程中,虽然过渡到低龄个体在抗性方面有所增强,但自身对环境资源的需求增大,有限的环境资源成为限制蒙古栎种群增长的重要因素。本研究中,4座火山南坡有限的环境资源条件使蒙古栎种群在龄级Ⅴ就开始处于高危险率和低生存率中。种群动态是揭示演替过程的关键所在[13]。不耐荫的蒙古栎生物学特性决定其在演替过程中将被耐荫的树种取代,加之森林的自我恢复能力,最终蒙古栎林可向原生林方向演替[38]。五大连池火山蒙古栎林是针阔混交林针叶树被间伐,逐步形成半原生状态的郁密度较大的阔叶林。本研究区1974年就成立五大连池自然保护区管理机构,通过相关政策和保护措施,保护和恢复了该区的天然林资源。多年的样地调查发现:随着立地条件的改善,更新层中已出现较多的色木槭,且在五大连池火山群中的新期火山老黑山北坡已出现面积较大的山杨-白桦-兴安落叶松Larix gmelini针阔混交林群落,老黑山熔岩台地上也零散分布有红松Pinus koraiensis幼树。在老期火山块状熔岩包围的“孤岛”上分布兴安落叶松纯针叶林群落,孤岛面积小,分布离散[39]。在黑龙江省各地的蒙古栎林观察发现:蒙古栎林在不被人类活动所影响下,会逐渐演替为阔叶混交林,直至演替为红松针阔叶混交林[40]。以红松为优势种的针阔混交林是小兴安岭等地的地带性植被,目前伊春市丰林自然保护区蒙古栎林下红松幼苗和幼树发育良好[40]。本研究区域蒙古栎种群地处火山锥山头附近,生境十分特殊,难以排除地形顶极群落的可能,在没有外界因素干扰的情况下,蒙古栎林演替发展是否与已有研究结果一致,向地带性植被红松针阔混交林方向演替还有待进一步研究。
-
五大连池4座老期火山蒙古栎种群当前均为稳定性型,但目前幼苗数量较少,自然更新较差,容易受外界环境的干扰。种群自然更新过程中低龄个体死亡率高,制约了种群的更新和发展,种群的增长潜力较小。时间序列预测结果表明:未来随着中、老龄个体的不断衰老、死亡,种群将趋于衰退。但对于火山生态条件下的蒙古栎种群演替动态规律还有待进一步研究。
Population structure and dynamic characteristics of Quercus mongolica in Wudalianchi Volcanoes, China
doi: 10.11833/j.issn.2095-0756.20210785
- Received Date: 2021-12-03
- Accepted Date: 2022-04-11
- Rev Recd Date: 2022-04-06
- Available Online: 2022-09-22
- Publish Date: 2022-10-20
-
Key words:
- Wudalianchi Volcano /
- Quercus mongolica /
- age structure /
- static life table /
- survival curve /
- suvival analysis /
- population dynamic index
Abstract:
| Citation: | XIE Lihong, HUANG Qingyang, CAO Hongjie, et al. Population structure and dynamic characteristics of Quercus mongolica in Wudalianchi Volcanoes, China[J]. Journal of Zhejiang A&F University, 2022, 39(5): 960-970. DOI: 10.11833/j.issn.2095-0756.20210785

|


 DownLoad:
DownLoad: