






-
云锦杜鹃Rhododendron fortunei为杜鹃花科Ericaceae常绿小乔木或灌木,主要分布于中国长江流域,生长在海拔600 m以上的沟谷阔叶林或山顶灌草丛中,为中国丘陵山地较高海拔地区常见的观赏价值较高的木本花卉[1],以“苍干如松柏,花姿若牡丹”著称。
近年来,中国关于云锦杜鹃的研究较多。不同地区的云锦杜鹃群落物种组成差异较大。陈艳华等[2]对湖南阳明山云锦杜鹃纯林和混交林的比较发现:无论是纯林还是混交林,物种组成均不丰富,群落垂直结构单一。邓贤兰等[3]对江西井冈山、袁丛军等[4]对贵州雷公山调查发现:不同海拔的云锦杜鹃样地物种多样性较高。在种群结构方面,赵丽娟等[5]对湖南平江幕阜山、黄川腾等[6]对广东天井山、付剑等[7]对湖北大别山的云锦杜鹃种群调查均发现:云锦杜鹃种群结构稳定,但幼苗较少,从而导致其种群更新不良。管康林等[8]分析了浙江华顶山云锦杜鹃林衰退的原因,认为可能系上层树种黄山松Pinus taiwanensis和柳杉Cryptomeria fortunei不断扩大和低灌层箬竹Indocalamus tessellatus的入侵蔓延所致。
边才苗等[9]研究发现:云锦杜鹃的种子不适合在过于湿润的土壤环境中萌发,但其种子对干旱胁迫比较敏感。杨华等[10]测定了2种光照条件下低海拔引种的云锦杜鹃的光合作用日变化,发现栽培的云锦杜鹃喜光特性并不会因为气温高而改变。此外,研究还涉及云锦杜鹃的扦插繁育[11]、花期的挥发物质成分分析[12]、转录组分析[13]及分子标记开发[14]等。
尽管云锦杜鹃群落特征、种群结构、种子萌发特性、光合生理等已有研究报道,但针对其种群及群落动态变化的研究仍显不足。此外,当前针对云锦杜鹃的研究方法通常是选取有限的小型样地进行静态分析,这种方法虽然有助于了解特定时间点的物种丰富度和群落组成,但未能充分反映种群动态变化的复杂性,而样地面积大小影响群落物种多样性及种群结构的分析结果。本研究基于浙江大盘山国家级自然保护区1 hm2固定监测样地的长期监测和调查资料,分析该区云锦杜鹃群落的基本特征、种群结构的动态变化,为资源保护、管理和利用等提供理论依据,同时也为大盘山国家级自然保护区较高海拔的地带性植被恢复与林相改造提出参考建议。
-
浙江大盘山国家级自然保护区位于浙江省金华市磐安县内,是浙江省钱塘江、灵江、瓯江三大水系主要支流的发源地,28°57′05″~29°01′58″N,120°28′05″~120°33′40″E。该区地貌属于中山山地,为典型的亚热带季风气候区,年均降水量为1 427.8 mm,年均气温为15.0 ℃,年均日照时数为1 827.6 h。地形复杂,气候优越,植物种类丰富。目前构成大盘山植被的主要类型为人工杉木Cunninghamia lanceolata林、竹林、马尾松Pinus massoniana林和常绿阔叶林,另有针阔混交林植被等[15]。
-
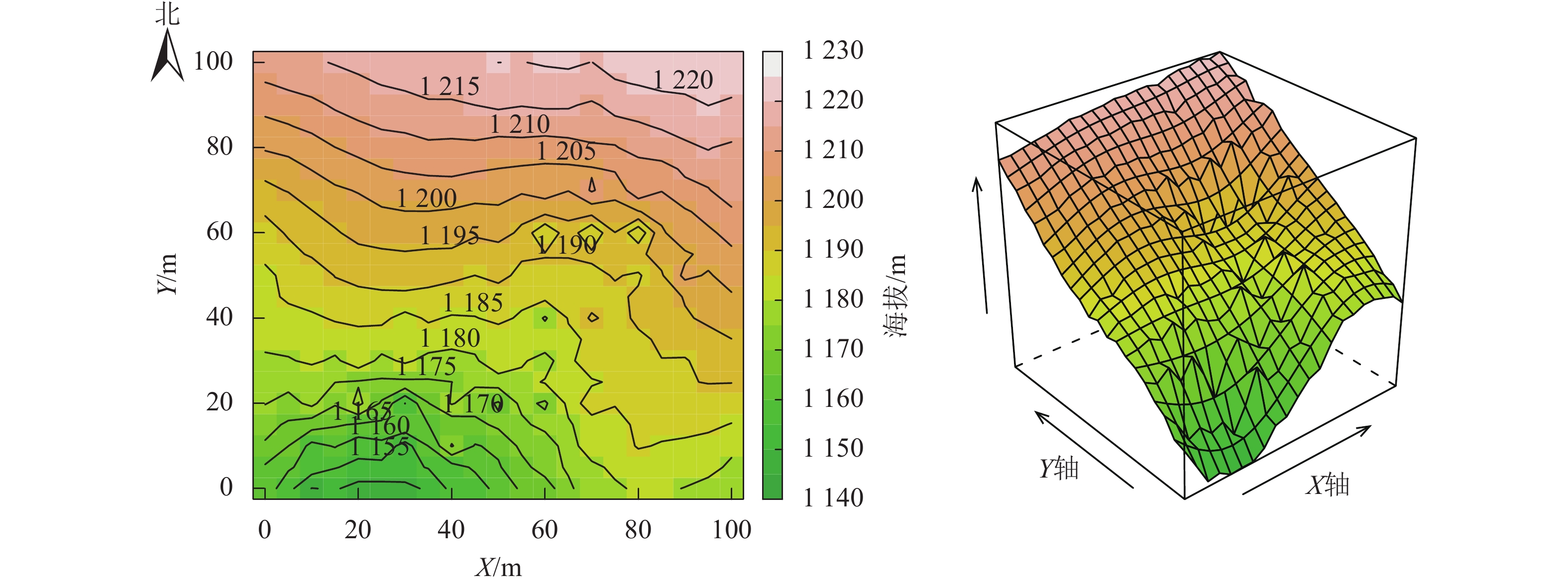
参照美国史密森热带研究所热带森林研究中心(Center for Tropical Forest Science,CTFS)的技术规范[16],2018年在大盘山国家级自然保护区内选择云锦杜鹃集中分布且生长状态较好的林地,建立1 hm2 (100 m×100 m)的固定监测样地,地理位置为28°58′33.24″N,120°31′36.12″E。样地最高点海拔为1 224 m,最低点海拔为1 148 m。样地地形较复杂,南北方向有1条纵沟,北部平均海拔高(图1)。用全站仪将整个样地划分为25个20 m×20 m的样方,每个样方再划分为16个5 m×5 m的小样方。对样地内胸径(DBH)≥1.0 cm的木本植株主干及其分支挂牌编号,以便永久监测,记录其胸径、冠幅、树高、小样方内坐标、生长状况等。2023年对该样地内所有挂牌木本植株进行复查,对新增树木(DBH≥1.0 cm)挂牌编号,记录死亡个体。
Figure 1. Topography of 1 hm2 forest dynamics plot in Mount Dapanshan of Zhejiang Province
-
参照《浙江植物志(新编)》 [1]鉴定样地植物。参照仲磊等[17]、丁炳扬等[18]的方法,按生长型划分样地木本植物的径级。乔木:1.0 cm≤DBH<5.0 cm (小树)、5.0 cm≤DBH <10.0 cm (中树)、10.0 cm≤DBH<25.0 cm (大树)、DBH≥25.0 cm (老树)。小乔木:1.0 cm≤DBH<3.0 cm (小树)、3.0 cm≤DBH<5.0 cm (中树)、5.0 cm≤DBH<15.0 cm (大树)、DBH≥15.0 cm (老树)。灌木:1.0 cm≤DBH <2.0 cm (小树)、2.0 cm≤DBH<3.0 cm (中树)、3.0 cm≤DBH<10.0 cm (大树)、DBH≥10.0 cm (老树)。稀有种:1 hm2仅为1株的物种。偶见种:样地内个体数大于1株且小于10株的物种。常见种:样地内个体数大于等于10株的物种[19]。
物种重要值=(相对多度+相对频度+相对显著度)/3,其中相对多度和相对频度仅计算独立个体,相对显著度计算包括了分枝的胸高截面积[20]。死亡率$ M=\dfrac{(\mathrm{l}\mathrm{n}{N}_{0}-\mathrm{l}\mathrm{n}{S}_{t})}{{T}} $;补员率$ R=\dfrac{(\mathrm{l}\mathrm{n}{N}_{t}-\mathrm{l}\mathrm{n}{S}_{t})}{{T}} $。其中:N0和Nt分别为第1、2次调查的个体数,St是该物种在第1、2次调查时都存在的活个体数,T为2次调查时间间隔。种群大小变化率(λ)用补员率和死亡率的差值表示。
参照文献[18],采用径级结构代替年龄结构的方法,将该种群径级结构划分为7个等级:Ⅰ级 (1.0 cm≤DBH<2.5 cm),Ⅱ级 (2.5 cm≤DBH<5.0 cm),Ⅲ级 (5.0 cm≤DBH<8.0 cm),Ⅳ级 (8.0 cm≤DBH<11.0 cm),Ⅴ级 (11.0 cm≤DBH<15.0 cm),Ⅵ级 (15.0 cm≤DBH<20.0 cm),Ⅶ级(DBH≥20.0 cm)。根据7个径级的个体数编制云锦杜鹃静态生命表[21]。生命表最关键的原始数据为ax,lx=ax/a0×
1000 ;dx=lx−lx+1;qx=dx/lx;Lx=(lx+lx+1)/2;$ {{T}}_{{x}}{=}\displaystyle \sum_{{x}}^{{\infty }}{{L}}_{{x}} $;ex=Tx/lx;Kx=lnlx−lnlx+1。其中:x为龄级;ax为匀滑后x龄级存活数;a0为最大个体数;lx为x龄级初始状态标准化存活个体数;dx为x到x+1龄级区间内标准化死亡数;qx为x到x+1龄级间隔期死亡率;Lx为从x到x+1龄级间隔期还存活的个体数;Tx为从x龄级到超过x龄级的个体总数;ex为进入x龄级的平均寿命;Kx为各龄级致死力(消失率)。根据静态生命表数据,绘制存活曲线(lnlx)、死亡率曲线(qx)。数据分析作图使用SPSS 27.0.1、R 4.3.2和Origin 2022等。
-
云锦杜鹃群落1 hm2固定监测样地于2018年调查并记录4 773株DBH≥1.0 cm的木本植物,分属于35科55属90种。其中,常绿树种只有16种,但个体数占调查总数的67%。样地中,云锦杜鹃的重要值最高,其次为黄山松(表1),因此该样地为常绿针阔混交林。从乔木层科的组成来看,含属数最多的为壳斗科Fagaceae (4属)、蝶形花科Fabaceae (4属)等。从属的组成来看,含有种数最多的为山胡椒属Lindera (4种)、槭属Acer (4种)和李属Prunus (4种)等。样地内常见种33种,其中个体数≥1 000株的仅有云锦杜鹃1种,个体数≥100株的常见种有黄山松、水马桑Weigela japonica var. sinica、红果钓樟Lindera erythrocarpa等6种;偶见种共35种,包括茅栗Castanea seguinii、盐肤木Rhus chinensis等;稀有种22种。2023年复查记录了5 018株DBH≥1.0 cm的木本植物,共计35科53属87种。间隔5 a后,有3种稀有种植物在样地消失,分别是接骨木Sambucus williamsii、圆叶鼠李Rhamnus globosa和隔药柃Eurya muricata。此外,样地中DBH≥1.0 cm的个体年死亡率为1.38%,年补员率为2.38%,种群大小变化率为1.00%。
树种 个体数/株 重要值/% 生活型 2018年 2023年 2018年 2023年 云锦杜鹃Rhododendron fortunei 2 341 2 380 36.21 36.43 常绿小乔木 黄山松Pinus taiwanensis 359 336 15.61 13.71 常绿乔木 红果钓樟Lindera erythrocarpa 228 254 5.30 5.53 落叶灌/小乔木 水马桑Weigela japonica var. sinica 243 197 4.72 3.86 落叶灌木 朝鲜白檀Symplocos coreana 208 289 4.59 5.41 落叶乔木 杉木Cunninghamia lanceolata 112 101 4.11 3.87 常绿乔木 尖连蕊茶Camellia cuspidata 219 312 3.44 4.56 常绿灌木 灯台树Bothrocaryum controversum 98 97 2.77 2.63 落叶乔木 短柄榛Corylus heterophylla var. brevipes 90 103 1.88 1.95 落叶灌/小乔木 合轴荚蒾Viburnum sympodiale 93 134 1.84 2.51 落叶灌木 Table 1. Top ten species with the highest importance values in the 1 hm2 forest dynamics plot in Mount Dapanshan
样地内群落的垂直结构分层明显。根据高度和生长型划分群落的垂直结构,树高(H)≥15.0 m的林冠层包括黄山松、杉木、灯台树等;H≥5.0 m的亚乔木层主要有云锦杜鹃、红果钓樟等;H<5.0 m的灌木层主要有尖连蕊茶、合轴荚蒾等。样地中优势度明显,重要值前10位的物种个体数占总个体数的80.45%,这10个物种的相对多度总和为83.62%,相对显著度总和为89.09%。2023年复查结果显示:样地内重要值前10位的物种未发生变化,但黄山松的重要值降幅最大,尖连蕊茶的重要值增幅最大。
-
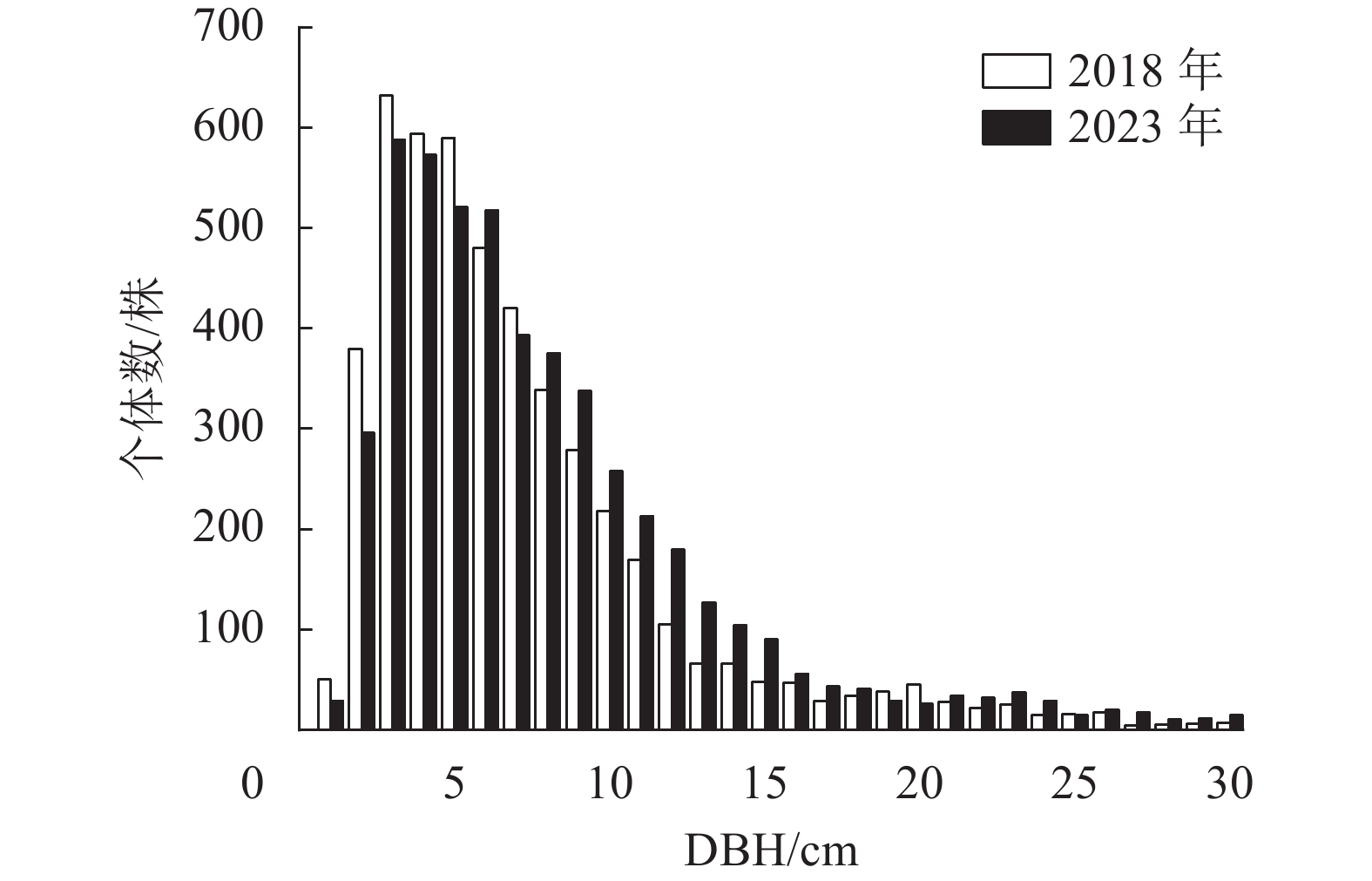
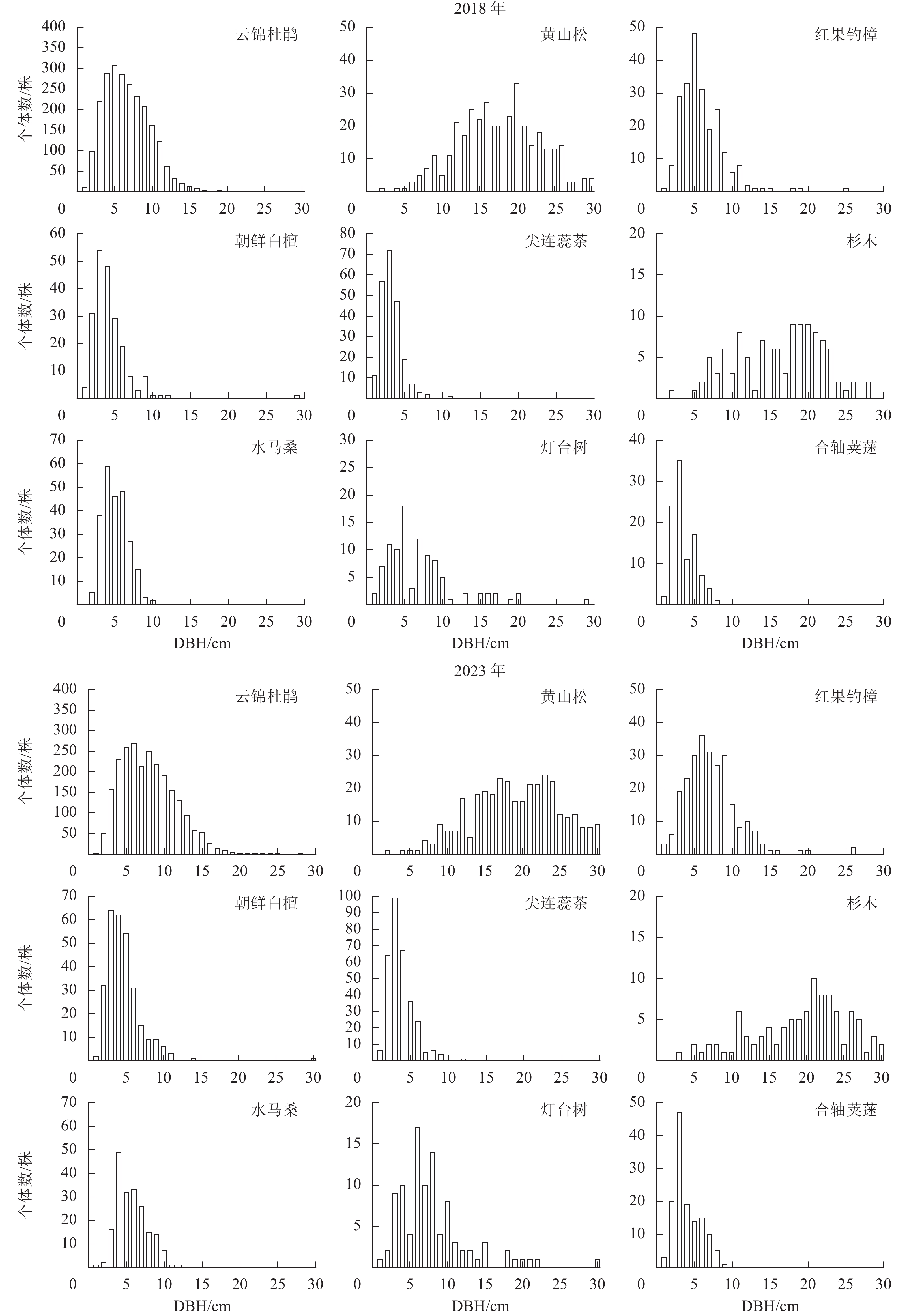
样地内所有木本植物(DBH≥1.0 cm)的径级分布呈现倒“J”型(图2)。2018年所有木本植物的平均径级为7.5 cm,其中1.0 cm≤DBH <5.0 cm的个体有1 655株,占总个体数的34.67%;5.0 cm≤DBH <20.0 cm的个体有2 928株,占总个体数的61.36%;DBH≥20.0 cm的个体有190株,占总个体数的3.98%。2023年所有木本植物的平均径级为8.3 cm,5 a间该样地平均径级增长了0.8 cm,径级分布依然为倒“J”型,其中1.0 cm≤DBH <5.0 cm的个体减少了169株;5.0 cm≤DBH <20.0 cm的个体增加了358株,DBH≥20.0 cm的个体增加了56株。样地重要值排名前9位的径级结构如图3。云锦杜鹃的径级分布表现为金字塔型,2018年小树和老树个体数较少,中树个体数占总个体数的21.66%,大树个体数占总个体数的72.32%;2023年大树的比例有所增加。黄山松小树和中树的储备较少,大树和老树的数量较多,2023年老树的比例上升。红果钓樟、朝鲜白檀、尖连蕊茶的径级分布总体呈现“L”型。杉木集中在大径级个体,并且径级分布不连续,有多个不同的峰;水马桑、灯台树和合轴荚蒾在样地内的种群数量少,且集中在小径级范围内。
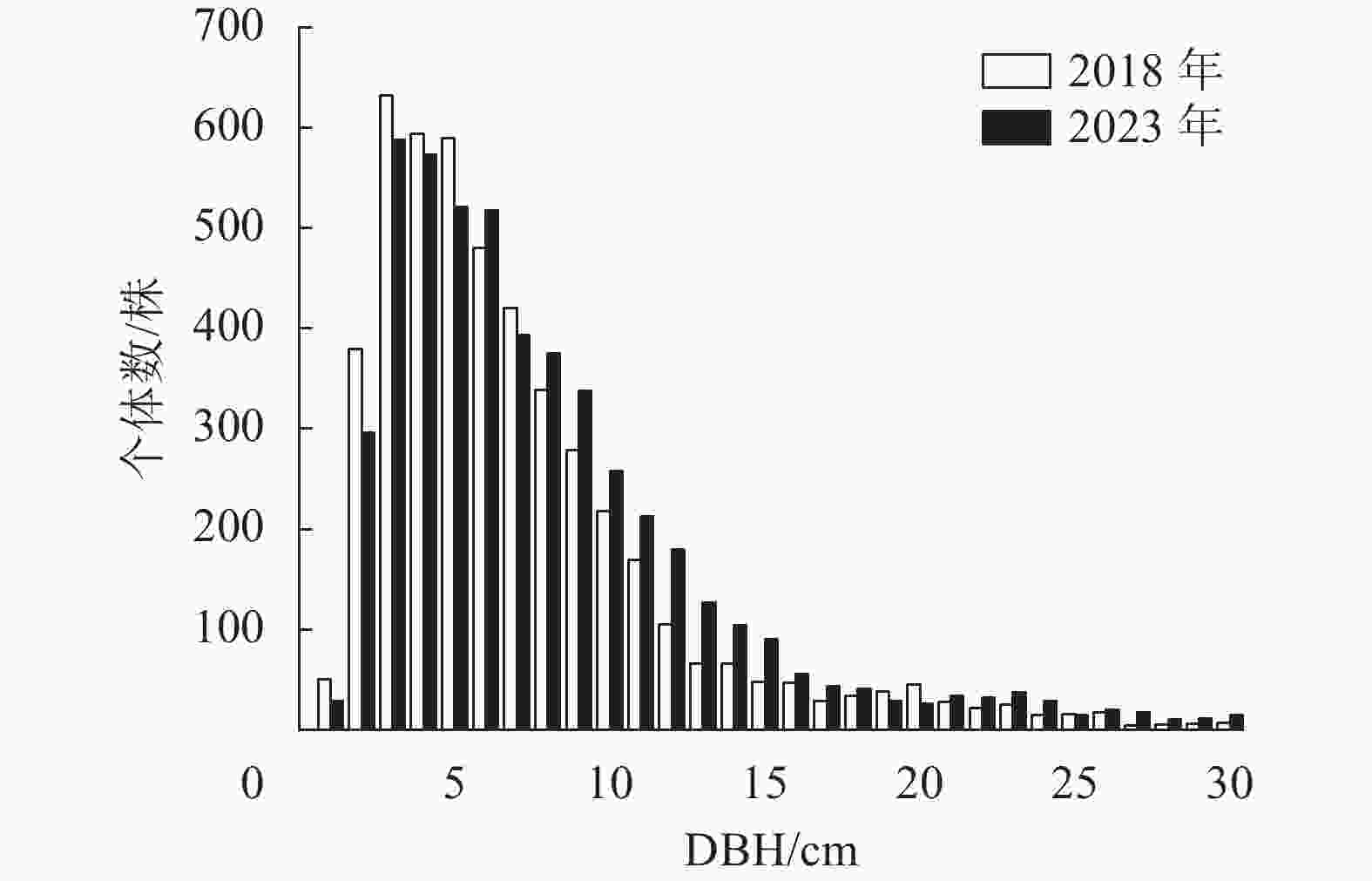
Figure 2. DBH class distribution of 1 hm2 forest dynamics plot of Mount Dapanshan

Figure 3. DBH class distribution of the top 9 tree species in the 1 hm2 forest dynamics plot in Mount Dapanshan
-
样地沿南北方向存在1条纵向沟谷,且北部的平均海拔高于南部,整体坡度为10°~30°。优势种云锦杜鹃(图4A)在样地内的分布与地形表现出明显的相关性,集中分布在山沟的两侧,山沟处有极少数分布。黄山松(图4B)在样地内分布较分散,且老树和大树的比例偏高。红果钓樟(图4C)和朝鲜白檀(图4D)在整个样地内均有分布。灌木树种尖连蕊茶(图4E)主要聚集在样地南部海拔相对较低的位置,大树占比为73%。杉木(图4F)聚集于样地西北角,位于海拔较高的地方,其中大树73株,占比为70%,老树19株,占比为20%,这意味着杉木以大树为主。灌木树种水马桑(图4G)多见于样地北部,南部稀疏分布。灯台树(图4H)在样地内共有97株,表现为零星分布。灌木树种合轴荚蒾(图4I)聚集分布在样地南部海拔较低的地方,北部零星分布,且以大树为主,占比为80%。
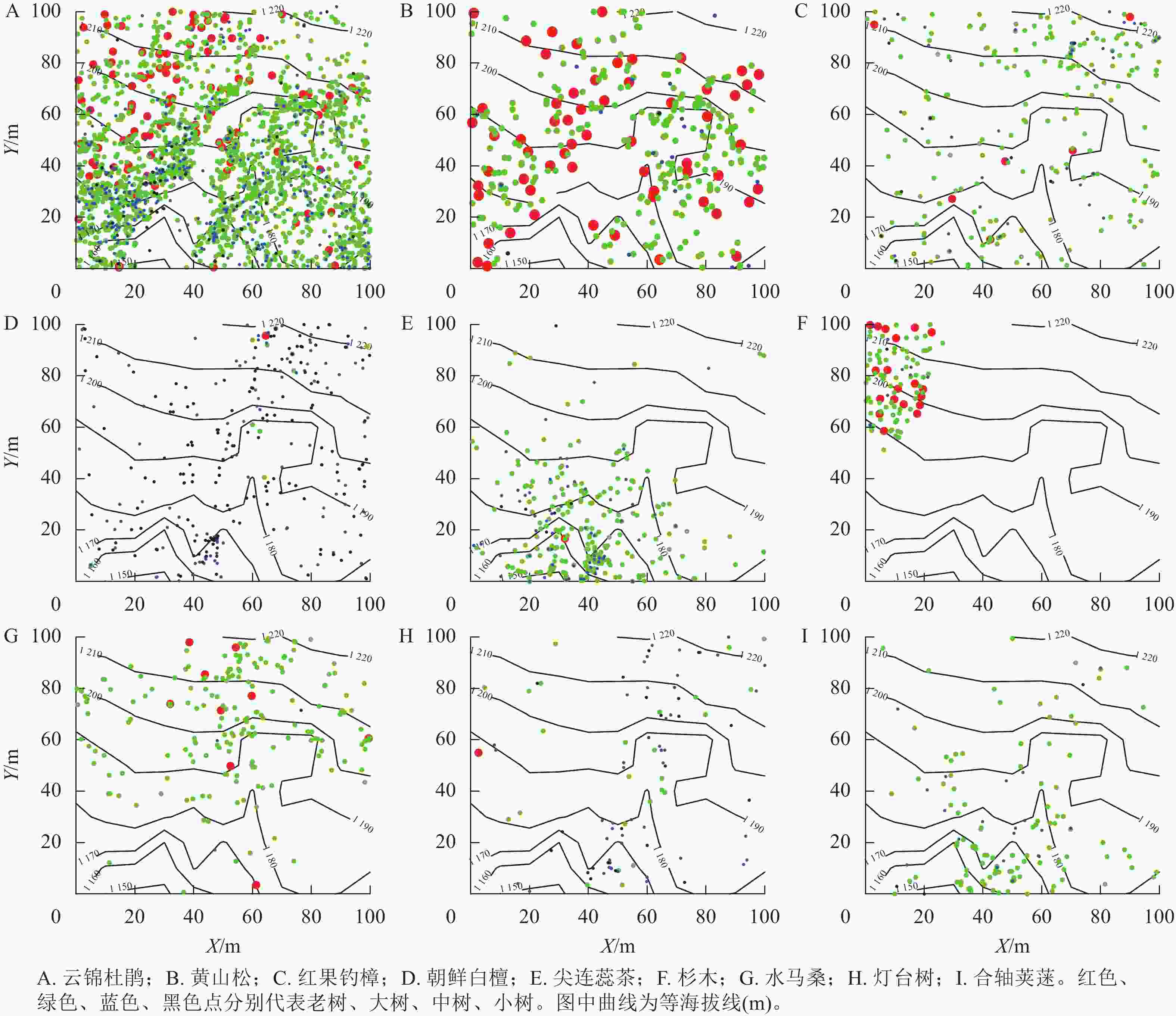
Figure 4. Spatial distribution pattern of the top 9 dominant species with important values in Mount Dapanhsan
-
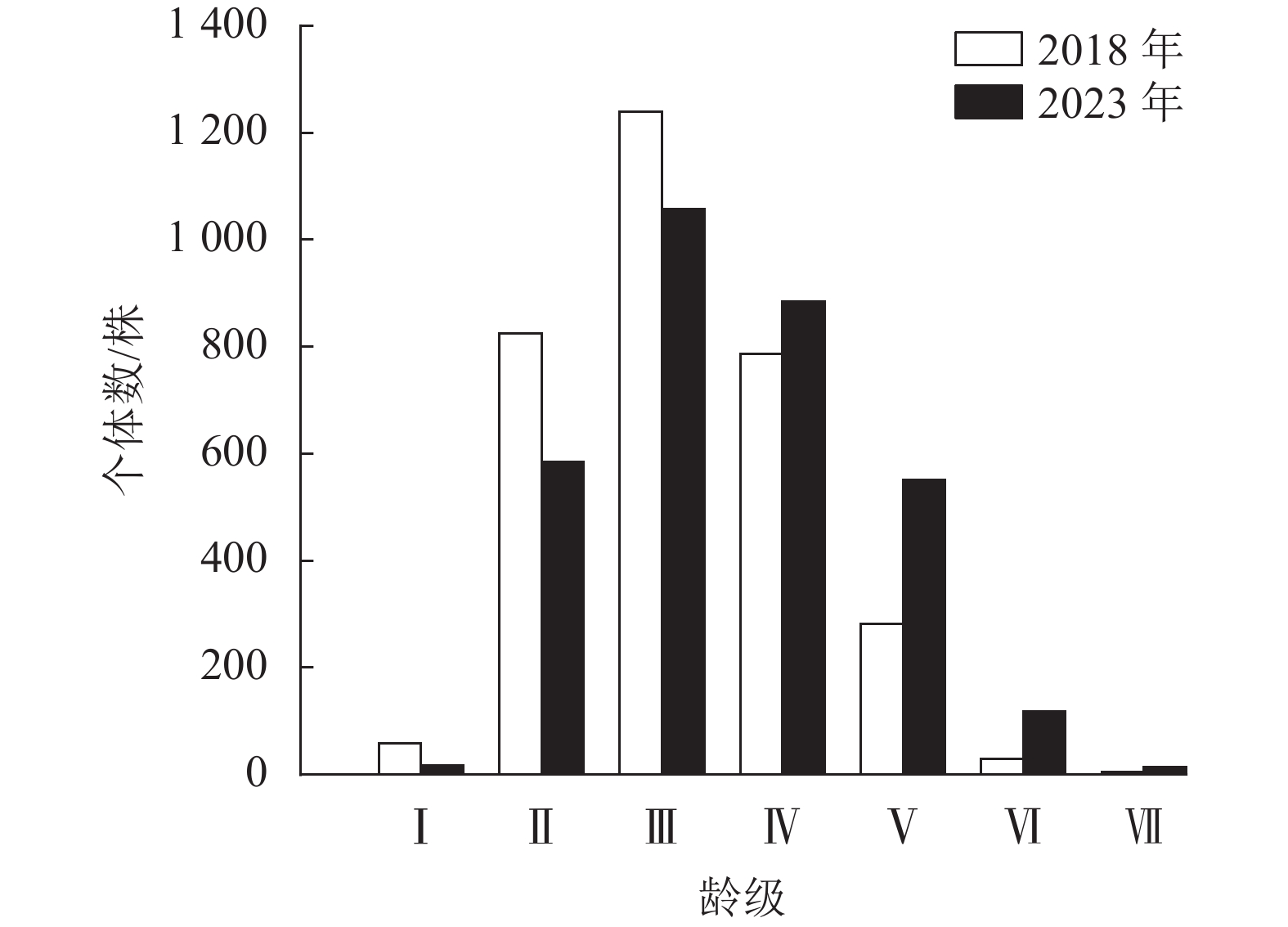
2018年云锦杜鹃个体数为2 341株,占样地内总个体数的49.00%;2023年为2 380株,占比为47.43%。云锦杜鹃的年龄结构总体表现为金字塔结构(图5),即集中在Ⅱ、Ⅲ、Ⅳ龄级,Ⅰ、Ⅵ、Ⅶ龄级的个体数很少。2023年Ⅳ、Ⅴ、Ⅵ龄级的个体数相比2018年明显增多,说明其5 a间对自然环境变化适应较好,龄级生长发育良好。根据云锦杜鹃种群存活曲线判断,2018、2023年均为Ⅰ型(凸型)(图6),表现为绝大多数个体都能活到生理年龄,早期死亡率极低,但是达到一定生理年龄时,短期内几乎全部死亡。云锦杜鹃死亡率在2018、2023年都表现出随着龄级逐渐增加,存活数逐渐减少,说明幼龄个体数较多(表2)。2018年调查时显示:Ⅰ~Ⅳ龄级个体死亡率较低,Ⅴ、Ⅵ龄级个体死亡率较高,分别为0.9和0.8;2023年死亡率随着龄级的增加而逐渐增大,Ⅵ龄级的个体死亡率最高,为0.9 (表2)。2018和2023年的个体平均寿命随着龄级的增加而降低,在Ⅰ龄级最高,说明该阶段种群生存能力强。
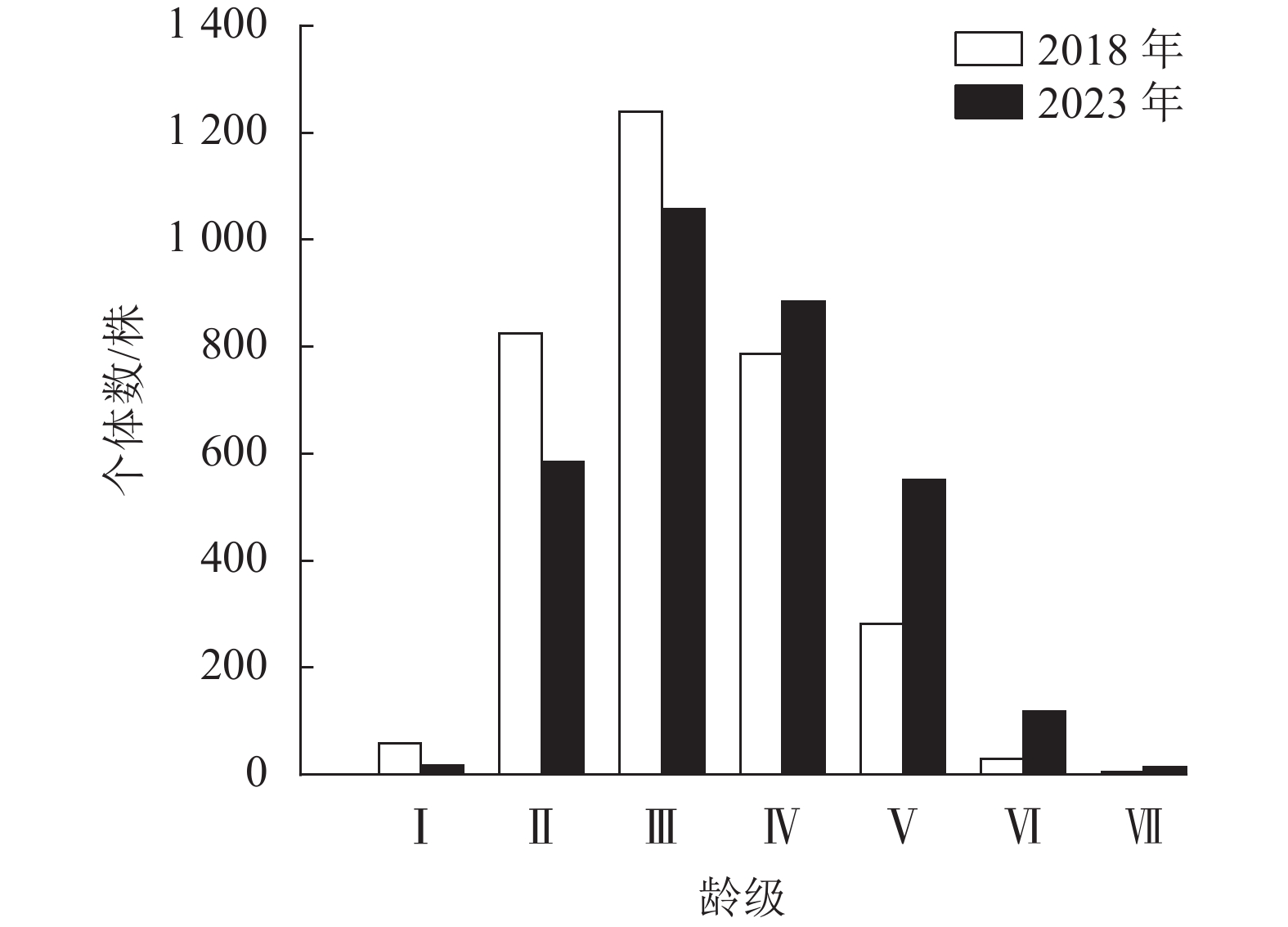
Figure 5. Age structure of R. fortunei population
调查年份 龄级 Ax ax lx lnlx dx qx Lx Tx ex Kx 2018 Ⅰ 43 888 1 000.0 6.9 208.3 0.2 895.8 2 554.1 2.6 0.2 Ⅱ 572 703 791.7 6.7 209.5 0.3 686.9 1 658.2 2.1 0.3 Ⅲ 854 517 582.2 6.4 208.3 0.4 478.0 968.5 1.7 0.4 Ⅳ 600 332 373.9 5.9 104.7 0.3 321.5 493.2 1.3 0.3 Ⅴ 239 239 269.1 5.6 237.6 0.9 150.3 171.7 0.6 2.1 Ⅵ 28 28 31.5 3.5 25.9 0.8 18.6 21.4 0.7 1.7 Ⅶ 5 5 5.6 1.7 - - 2.8 2.8 0.5 - 2023 Ⅰ 14 665 1 000.0 6.9 159.4 0.2 920.3 3 079.0 3.1 0.2 Ⅱ 422 559 840.6 6.7 157.9 0.2 761.7 2 158.7 2.6 0.2 Ⅲ 739 454 682.7 6.5 159.4 0.2 603.0 1 397.0 2.1 0.3 Ⅳ 658 348 523.3 6.3 157.9 0.3 444.4 787.2 1.5 0.4 Ⅴ 436 243 365.4 5.9 212.0 0.6 259.4 349.6 1.0 0.9 Ⅵ 102 102 153.4 5.0 139.9 0.9 83.5 90.2 0.6 2.4 Ⅶ 9 9 13.5 2.6 - - 6.8 6.8 0.5 - 说明:Ax表示龄级内现有存活数。由于缺少下一龄级数据,dx、qx和Kx无法计算,用-表示。 Table 2. Static life table of R. fortunei population
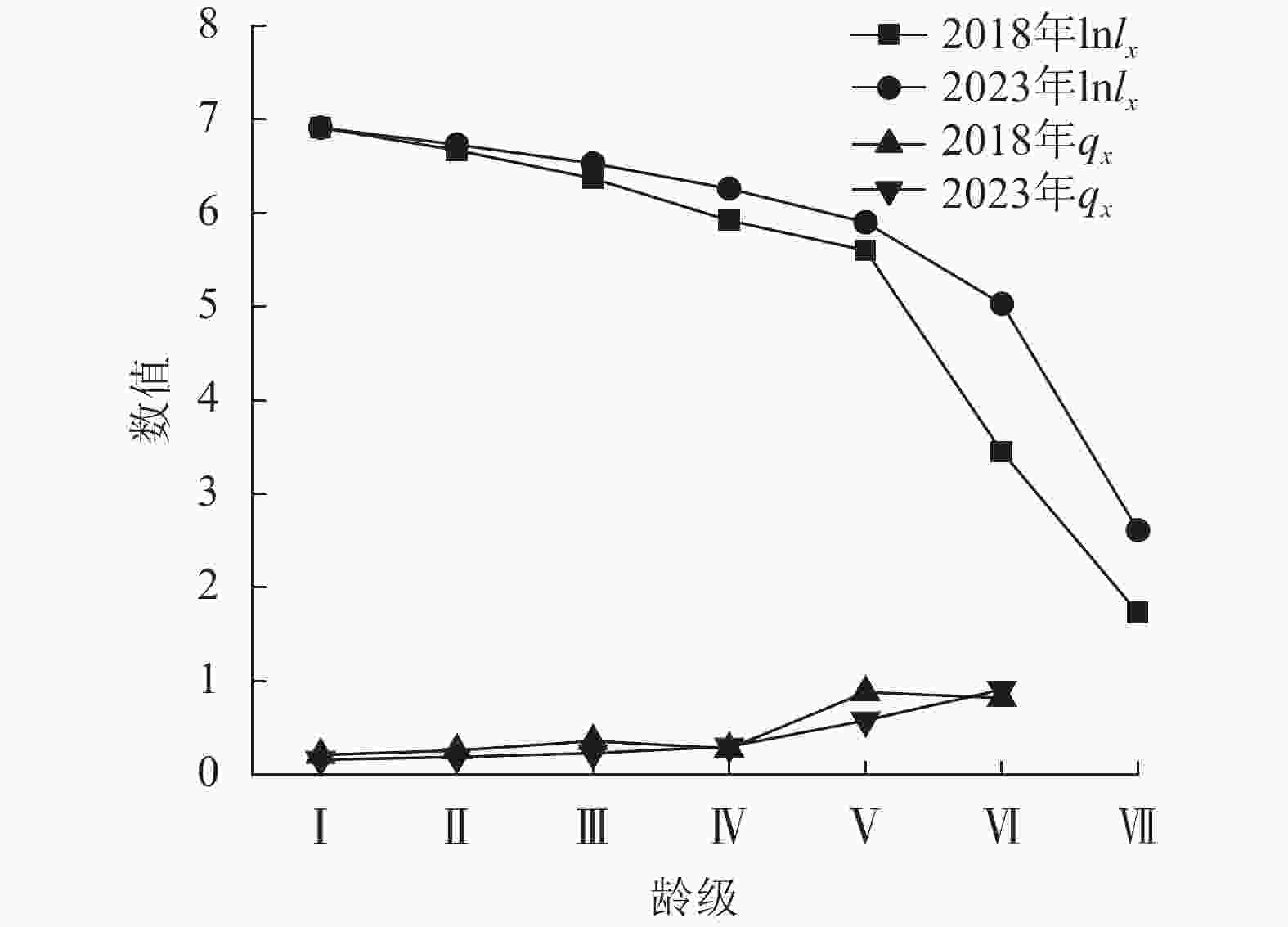
Figure 6. Survival curve (lnlx) and mortality rate curve (qx) of R. fortunei population
-
物种组成是决定群落性质的重要因素,也是了解生态系统维持机制的重要途径[22]。大盘山的云锦杜鹃群落物种组成丰富,5 a间样地排前10位的优势物种组成无变化,说明样地内的优势种总体上自我更新较好[23],与同样以云锦杜鹃为优势种的湖南阳明山群落10 a间乔木层的物种组成研究结果一致[2],即样地内优势种稳中有变,更新状态良好。样地常绿树种个体占比高达67%,整体上可反映亚热带较高海拔的森林群落特征。5 a间样地林冠层常绿树种云锦杜鹃重要值增长0.22%,尖连蕊茶增幅最大,相比5 a前增加了1.12%,而针叶树种黄山松和杉木的重要值则分别降低了1.90%和0.24%。该结果与地理位置和植被类型较为接近的浙江东白山1 hm2次生针阔混交林样地[24]的研究结果相似,即喜光的先锋树种黄山松等针叶树种的优势地位在未来可能会丧失。该样地随着时间推移将可能逐渐演替为以云锦杜鹃等为优势种的常绿阔叶林。大盘山群落的物种多样性较高,而同样以云锦杜鹃为优势种的江西井冈山样地[3]的群落多样性也同样丰富。这2个样地均位于中国东部较高海拔地区,且都处于亚热带温暖湿润气候区。这种气候特征为植物种群多样化生长提供了良好的环境。此外,较高海拔的地形和微气候的多样性进一步丰富了物种的生态位,推动了物种间的相互作用和竞争,有助于维持和增强物种多样性[25]。样地内偶见种和稀有种数较多,复查时死亡的3个物种全部为稀有种,可能由于其适合度较低,在样地中竞争力较弱,因而更容易被竞争排除[26]。
-
径级结构可以表示在该群落内各物种的不同胸径大小的个体数量分布情况,而径级结构的动态则反映种群内不同年龄个体的生长和发展趋势[27]。2018—2023年,大盘山云锦杜鹃群落1 hm2样地内径级结构呈现倒“J”型分布,说明群落总体上幼龄植株数量多,更新良好。5 a间样地内所有物种的径级结构无显著变化,小径级个体数下降,中径级个体增加,说明外界条件适宜,样地内物种稳定增长。优势种云锦杜鹃呈现金字塔型,以4.0 cm≤DBH<6.0 cm的个体为主,更新良好,但1.0 cm≤DBH<2.0 cm的幼树个体储备不足,应确保幼苗库的补给,否则长时间演替可能出现“青黄不接”的情况。群落内黄山松和杉木主要为10.0 cm≤DBH<25.0 cm的大树。这表明个体从小径级发育为大径级的过程中,为了获取更多的水分、光照、营养和空间,同种个体的自疏作用导致相邻个体的死亡,综合竞争力较强的植株生长较为稳定,最终得以长成大树[28−29]。红果钓樟、朝鲜白檀、尖连蕊茶的径级分布均呈现“L”型,表明其更新状况良好,种群数量会保持稳定增长。此外,一些数量较少的树种,如水马桑等多以小径级个体为主,种群更新较快,有利于群落结构的稳定[30−31]。
-
种群在空间上的分布反映了外界环境因子对个体生存的影响,是分析种内和种间竞争、种子扩散和干扰等生态过程的基础[32]。该样地的优势物种空间分布具有生境偏好,且不同物种之间的生境偏好存在差异。对浙江古田山样地[33]的研究表明:物种在同一生态系统中能够通过时间、空间或资源使用上的差异,避免直接竞争,从而实现更有效的资源利用,即生态位分化机制在样地森林群落的生物多样性维持中有重要的作用。优势种云锦杜鹃种群在样地内均有分布,但山沟底部分布稀少,可能是由于其环境较为潮湿。有研究表明:云锦杜鹃的种子不适宜在过于湿润的土壤环境中萌发[9]。乔木亚层树种小径级较多,如云锦杜鹃、红果钓樟和尖连蕊茶等出现聚集分布,有利于改善群落微环境,增强种群抵御不良环境的能力[34]。LIU等[35]研究发现:竞争是构建群落和驱动树木空间分布模式的主要机制,在小范围聚集,且聚集程度随着距离增加而降低与种子扩散能力有关。乔木亚层植株高度有限,种子落在母株附近的概率更大,新生个体更容易聚集在母株周围。树种从小径级到大径级的空间分布特征一般由聚集分布到随机分布,最终为规则分布[36]。在样地中,黄山松、杉木等高大乔木由于个体生长发育对空间资源的竞争加强,逐渐过渡为随机分布。
-
云锦杜鹃种群的年龄结构呈金字塔,为增长型,种群数量随着龄级的增加呈先升后降的趋势,但5 a间Ⅰ龄级个体更新少。在复查过程中发现:灌木层云锦杜鹃幼苗的出现概率很低,推测可能是由于种子发育为幼苗的过程存在障碍。广东天井山、湖北大别山的云锦杜鹃种群均报道有幼苗不足的情况。也有研究表明:杜鹃花属Rhododendron植物种子自然更新对生境条件要求严格[37]。边才苗等[11]研究发现:云锦杜鹃结实率和结籽率很低。另外,样地内黄山松大树的冠幅较大,郁闭度高也会使幼苗生长光照不够,还可能与区域地形复杂有关,如坡度较大、林下的凋落物层较厚,飘落的种子难以直接在枯枝落叶层上萌发。种群内Ⅵ、Ⅶ龄级数量稀少,说明云锦杜鹃老树对环境的适应能力减弱。本研究中云锦杜鹃种群的个体平均寿命总体呈现下降的趋势,在Ⅰ龄级最大,说明此时种群生存能力较强,对环境具有较强的适应力。存活曲线属于Deevey-I型,说明种群稳定增长,自然环境适宜大多数个体存活到生理寿命。结合年龄结构和存活曲线,5 a间种群更新较好,但Ⅰ龄级数量少将难以保障其种群向中龄级植株和大龄级植株生长的需求,在演替的过程中遭受环境波动和气候变化等自然灾害易造成种群年龄结构的变动[38]。为实现云锦杜鹃种群结构的稳定和种群数量的增长,建议保护现有幼龄植株,并通过人为辅助措施提高幼苗的存活率,加大就地保护力度。
-
浙江大盘山国家级自然保护区王盼、金京华,浙江农林大学段雨豪、王冠舜、解江涛、张坤、张秀娟、何淑冉,杭州师范大学蔡鑫、耿新、何芳参加了野外样地调查;鲁益飞在数据分析过程中提供了帮助;温州大学刘金亮对论文初稿提出宝贵修改意见。谨致谢意!
Community structure and dominant population dynamics of Rhododendron fortunei in Mount Dapanshan, Zhejiang Province
doi: 10.11833/j.issn.2095-0756.20240476
- Received Date: 2024-08-03
- Accepted Date: 2024-11-16
- Rev Recd Date: 2024-11-14
- Available Online: 2024-12-12
- Publish Date: 2025-02-20
-
Key words:
- Rhododendron fortunei /
- community structure /
- spatial distribution pattern /
- population structure /
- species diversity /
- Mount Dapanshan
Abstract:
| Citation: | LI Qige, YU Yefei, CHEN Weijie, et al. Community structure and dominant population dynamics of Rhododendron fortunei in Mount Dapanshan, Zhejiang Province[J]. Journal of Zhejiang A&F University, 2025, 42(1): 1−11 doi: 10.11833/j.issn.2095-0756.20240476

|


 DownLoad:
DownLoad: