






-
铁线莲属Clematis植物为毛茛科Ranunculaceae多年生草本或木本植物,花色艳丽、花型多样,被广泛用于阳台、庭院等家庭园艺场景,被誉为“藤本皇后”[1]。全世界铁线莲属植物约300种,中国有100多种,各地均有分布[2]。单叶铁线莲Clematis henryi为该属木质藤本植物,广泛分布在中国安徽、江苏、浙江等地,适生于林下荫蔽的环境,花白色或淡黄绿色,花量大且具芳香,花期12月至翌年2月,是铁线莲属中为数不多的冬季开花种类,具有极大的开发利用潜力[3]。
光是植物生长过程中重要的环境因素之一,光强不足或过剩都会对植物的光合作用产生不利的影响[4]。现有研究表明:植物的生长对遮光响应十分敏感。遮光处理改变了植物生长的微环境,从而对植物叶片光合生理和解剖结构产生影响[5]。当植物所处的光照条件发生改变时,叶绿素荧光参数能够直观反映植物光合系统的运转状态[6],因此在植物光合能力和栽培适应性评价中被广泛应用[7−8]。近年来,在园艺市场常见的铁线莲品种多为春(秋)季开花的喜阳类型,单叶铁线莲作为冬季开花且具有良好耐荫性的种类,可以满足林缘、北侧墙体或阳台、室内窗台等多种园林与家庭园艺场景的栽培需求,引种栽培后将有效扩展观赏铁线莲的园林应用范围。目前,有关铁线莲属植物光合特性的研究主要集中在光强对植株生长发育[9−10]以及水分等其他环境因子对光合系统的影响[11]等方面,尚未涉及铁线莲属植物引种栽培和园林应用中对光照环境的选择与调控。本课题组前期对单叶铁线莲耐荫生理变化已有报道[12]。本研究在春末(6月)、盛夏(8月)和仲秋(10月)对单叶铁线莲在遮光环境下的枝叶表型、光合特性与叶绿素荧光参数变化进行了系统分析,更直观和全面地解析了单叶铁线莲光合系统对低光环境的适应机制,可为单叶铁线莲在园林中的栽培应用提供依据。
-
试验于2022年3—10月在铁线莲种质资源圃(30°13′19.2″N,119°22′33.6″E)进行。该地属亚热带湿润季风气候,雨量充足且四季分明,年平均气温为13.0~21.0 ℃,年降水量为1 427.0 mm,年平均日照时数为1 939.0 h,无霜期为234.0 d。2022年3月2日,选取60株长势良好、均一的3年生单叶铁线莲盆栽扦插苗(栽植盘上口径为16.0 cm、下口径为12.0 cm、高为13.5 cm)。栽培基质采用珍珠岩∶泥炭土为1∶1 (体积比),pH 6.5,有机质质量分数为605.0 g·kg−1,置于日光温室内(遮光度为63%),常规水肥管理备用。
-
2022年5月20日,从上述移栽的植株中选取生长健康、长势一致的40株用于遮光试验。共设定5个遮光度:82% (T1)、61% (T2)、53% (T3)、43% (T4)和0% (对照),每个处理8株,植株间距为25 cm,防止相互遮挡。试验期间常规水肥管理。分别于试验初期(6月16—17日)、中期(8月15—16日)和末期(10月16—17日)进行叶片、植株形态及光合与荧光参数测定。
-
以基部叶片着生位置为起点,测定不同处理下各植株的节间距;取植株中上部,高度相近的健康叶片,利用网格法[13]测定叶面积、叶宽、叶长和叶周长。徒手切片参照邱彦芬等[14]的方法,用双面刀片在叶片主脉中段横切若干0.5 mm左右薄片,于显微镜(Olympus Microscope BX43)下观察拍照,测定叶片厚度(TL)、上表皮细胞厚度(TU)、下表皮细胞厚度(TL)、栅栏组织厚度(TP)、海绵组织厚度(TS),计算栅海比(TP/TS)、组织结构紧密度(CTR=TP/TL)及组织结构疏松度(SR=TS/TL)。将新鲜叶片置于烘箱中105 ℃杀青30 min,65 ℃烘干至恒量,称取叶干质量,计算叶片含水量[叶片含水量=(叶片鲜质量-叶片干质量)/叶片鲜质量]和比叶面积(比叶面积=叶面积/叶片干质量)。3次重复。
-
光合参数测定。采用Li-6400XT便携式光合仪进行光响应曲线和光合日变化测定。在测定日7:00—18:00,各处理随机选3株长势健康、一致的植株中上部叶片,在自然光下,隔2 h测定叶片的净光合速率(Pn)、胞间二氧化碳摩尔分数(Ci)、蒸腾速率(Tr)、气孔导度(Gs),3次重复。设置光强梯度为2 000、1 800、1 500、1 200、800、600、400、200、150、100、100、50、20、0 μmol·m−2·s−1,测定各光强下的净光合速率,重复3次,用双曲线修正模型拟合并计算表观量子效率(AQY)、最大净光合速率(Amax)、光饱和点(PLS)、光补偿点(PLC)以及暗呼吸速率(Rd)。叶绿素荧光参数的测定。测定日8:00—11:00,选取与光合参数测定相同的3株植株,在暗反应状态下测定叶片的初始荧光(Fo)和最大荧光(Fm)。测定前用锡纸完全包裹1 h。光下活化30 min后于1 000 μmol·m−2·s−1光强下测定最大荧光($F'_{\mathrm{m}} $)、最小荧光($F'_{\mathrm{o}} $)、光化学猝灭系数(qP)、电子传递效率(RET)。计算出最大光化学效率(Fv/Fm)、 PSⅡ实际光量子产量($F'_{\mathrm{v}}/F'_{\mathrm{m}} $)、PS Ⅱ潜在活性(Fv/Fo)、非光化学猝灭系数(QNP),其中Fv和$F'_{\mathrm{v}} $分别指可变荧光和光适应可变荧光。
-
采用Excel 2021和Origin 2022进行数据整理并作图,利用SPSS 26.0进行方差分析和相关性分析等。
-
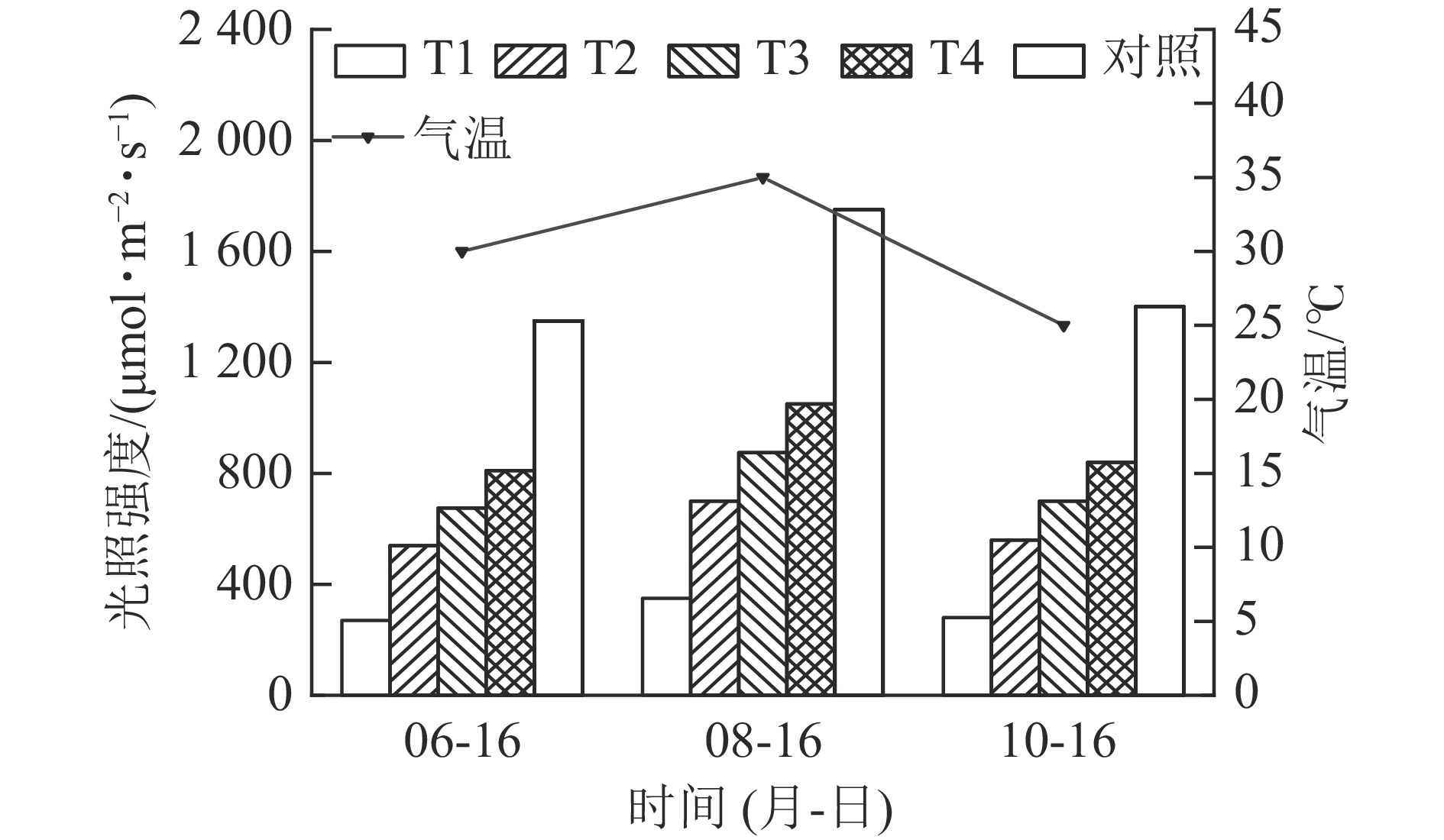
试验期间,外界最高气温和最大光照强度均呈“先升后降”的变化趋势(图1)。8月最高气温达35.0 ℃,最大光照强度达1 750 μmol·m−2·s−1,较6月分别提高23.0%和29.6%;10月的最高气温回落至26.0 ℃,最大光照强度降至1 400 μmol·m−2·s−1。这期间气温和光照强度的变化,为试验材料提供了“适宜—胁迫—恢复”的外界环境条件。
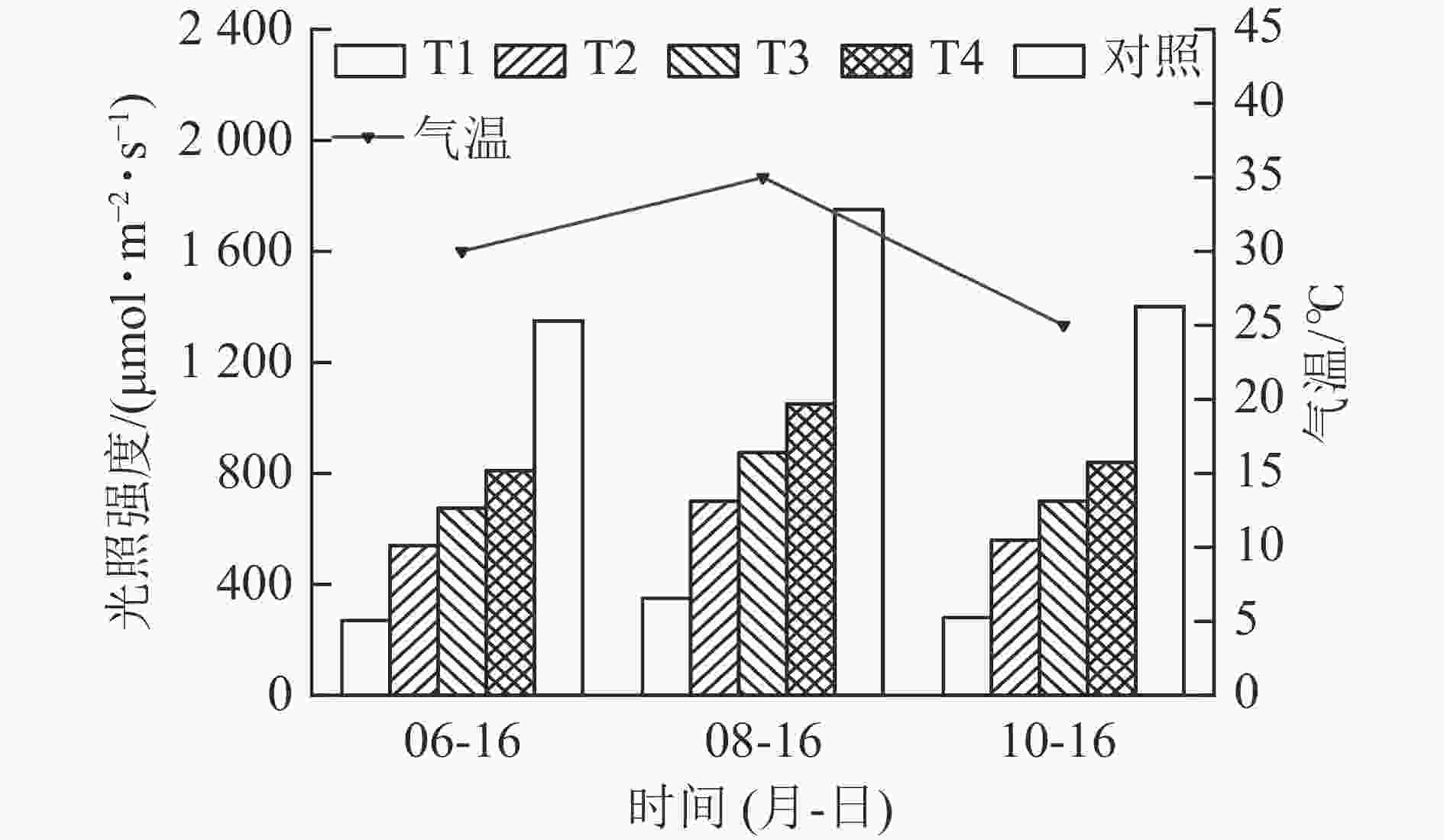
Figure 1. Changes of light intensity and temperature during the test
-
试验初期(6月),各处理植株长势差异不明显,处理组较对照组叶色更浓绿。试验中期(8月),对照组叶片变黄、枯萎,T4处理叶片发黄,部分枯萎;T3处理叶片绿色减淡,少数叶片边缘枯黄;T1和T2处理植株长势和叶色没有明显变化。试验末期(10月),对照组植株全部枯萎死亡,T4处理植株有1/2的叶片枯黄,T3处理植株叶片转绿,T1和T2处理叶色仍保持浓绿,新叶大量萌生。可见,在夏季遮光度递增对植株的保护作用也更好。
表1显示:遮光促进了单叶铁线莲叶片生长。在试验的3个阶段,T1和T2处理的植株叶长度分别达到11.49、12.58、12.65和11.35、12.83、12.33 cm,显著高于其他处理(P<0.05),T1、T2和T3处理植株的叶宽也均大于T4和对照(P<0.05)。在试验的3个阶段,T1处理植株的叶面积、比叶面积、节间距和叶干质量大都显著高于其他处理(P<0.05),随遮光度降低,上述指标也不断减少。方差分析结果显示:遮光度对叶片形态产生了显著影响,叶长、叶面积和叶干质量在T1处理中的均值最高,分别是对照组的1.90、2.41和1.53倍,可见遮光度增加促进了叶片生长和干物质积累。此外,各处理的叶片含水量仅在8月呈显著差异(P<0.05),表明遮光处理在高温强光条件下对叶片保护更有效。
月份 处理 形态特征参数 叶长/cm 叶宽/cm 叶面积/cm2 节间距/cm 干质量/g 含水量/% 比叶面积/(cm2·g−1) 6月 T1 11.49±0.15 a 5.10±0.72 a 37.13±0.13 a 18.27±0.87 a 0.37±0.32 a 79.13±2.11 a 370.14±11.12 a T2 11.35±0.10 a 4.90±0.10 a 30.33±0.38 c 15.83±1.04 ab 0.30±0.16 a 73.31±4.21 a 133.46±12.34 b T3 11.58±0.05 a 5.14±0.25 a 29.85±0.13 c 16.50±1.14 a 0.10±0.02 b 72.23±9.15 a 119.62±10.21 b T4 10.85±0.10 b 4.53±0.05 c 33.96±0.31 b 13.30±1.21 bc 0.26±0.03 ab 72.32±1.15 a 131.75±10.04 b 对照 10.75±0.70 b 4.16±0.07 b 29.25±0.35 d 12.57±2.35 c 0.14±0.01 b 71.33±4.41 a 209.87±24.70 b 8月 T1 12.58±0.45 a 4.50±0.20 a 42.88±3.41 a 18.50±0.44 a 0.35±0.05 a 84.36±0.49 a 417.73±27.92 a T2 12.83±0.06 a 4.40±0.20 ab 33.92±3.76 b 17.57±0.40 a 0.31±0.03 a 80.17±4.41 a 130.20±5.64 b T3 10.10±0.10 b 4.70±0.46 a 32.43±3.79 b 18.90±0.20 a 0.26±0.02 b 65.33±2.21 b 92.92±3.25 c T4 9.90±0.17 b 3.53±0.81 bc 22.25±2.41 c 13.70±0.42 b 0.10±0.02 c 64.36±2.17 b 85.74±6.14 c 对照 8.83±0.21 c 3.10±0.26 c 17.72±0.26 c 12.65±2.35 b 0.05±0.01 c 54.43±2.26 c 54.72±5.48 d 10月 T1 12.65±0.07 a 4.53±0.46 ab 34.52±5.99 a 17.00±1.41 a 0.23±0.07 a 74.27±4.33 a 210.64±7.83 a T2 12.33±0.46 a 4.87±0.31 a 34.25±2.17 ab 13.73±1.17 ab 0.22±0.10 a 71.42±2.17 a 187.32±9.53 b T3 9.77±0.55 b 5.03±0.67 a 33.67±3.54 ab 11.17±2.84 bc 0.19±0.01 ab 74.31±2.33 a 183.27±7.07 b T4 8.77±0.67 c 4.30±0.14 b 25.17±4.91 b 8.13±1.18 c 0.12±0.03 b 70.43±7.15 a 160.87±6.66 c 说明:数据为平均值±标准差。不同小写字母表示同一月份不同处理间差异显著(P<0.05)。10月对照植株死亡。 Table 1. Effect of shade on plant and leaf morphology of C. henryi
由表2可以看出:叶片上表皮厚度、下表皮厚度在试验中期和末期都表现出随遮光度增加而减少的趋势,叶片厚度在试验中期也呈现同样的变化趋势,表明遮光阻碍了叶片表皮和总厚度的增长。叶片栅栏组织厚度在试验的3个时期均表现出随遮光度增加而减少的趋势,以8和10月数据为例,T1处理较T4处理显著降低(P<0.05),海绵组织厚度变化趋势则截然相反,表明遮光在促进叶片海绵组织扩展的同时,抑制了栅栏组织的生长。这在叶片海栅比数据中也得到了证实。
月份 处理 叶片解剖结构参数 叶片厚度/μm 上表皮厚度/μm 下表皮厚度/μm 栅栏组织厚度/μm 海绵组织厚度/μm 栅海比 6月 T1 440.37±5.03 a 33.14±4.26 a 20.79±5.54 b 90.38±8.52 b 292.21±4.26 a 0.31±0.03 a T2 420.24±0.43 a 40.07±8.95 a 24.70±0.85 ab 87.36±4.26 b 284.68±14.91 a 0.31±0.09 a T3 434.10±0.43 a 36.45±0.43 a 21.33±0.34 ab 103.03±0.85 a 264.50±0.85 b 0.39±0.07 a T4 435.01±7.67 a 42.18±0.00 a 24.40±0.43 ab 96.40±8.52 ab 268.11±12.78 b 0.36±0.08 a 对照 443.44±24.71 a 41.57±0.85 a 28.02±1.28 a 105.74±3.83 a 268.31±21.30 b 0.39±0.03 a 8月 T1 399.04±8.51 c 24.13±9.78 c 22.27±5.57 b 77.02±5.80 b 309.23±11.59 a 0.25±0.06 c T2 374.91±17.01 c 26.19±10.04 c 23.20±1.61 b 79.63±11.59 b 263.55±11.59 b 0.30±0.06 b T3 439.50±5.02 b 24.33±3.21 c 27.84±0.00 ab 92.80±3.21 ab 262.25±8.03 b 0.35±0.04 b T4 458.43±11.59 b 30.83±3.21 b 27.84±5.57 ab 98.37±7.01 a 252.64±6.43 b 0.39±0.06 ab 对照 515.97±12.86 a 51.04±4.25 a 35.26±6.43 a 103.01±7.37 a 224.58±17.90 c 0.46±0.06 a 10月 T1 399.04±13.73 a 29.70±3.21 b 24.13±11.59 b 79.81±11.59b 263.55±21.08 a 0.30±0.03 b T2 402.75±14.01 a 28.98±9.64 b 29.70±4.25 b 85.38±3.21 a 236.64±22.10 ab 0.36±0.06 b T3 396.05±6.43 a 39.18±5.80 a 40.83±12.55 ab 82.59±16.31ab 211.58±5.57 b 0.39±0.02 ab T4 409.25±15.50 a 41.97±3.21 a 50.11±5.57 a 93.94±11.59a 217.15±5.57 b 0.43±0.03 a 说明:数据为平均值±标准差。不同小写字母表示同一月份不同处理间差异显著(P<0.05)。10月对照植株死亡。 Table 2. Effects of shade on leaf anatomy of C. henryi
-
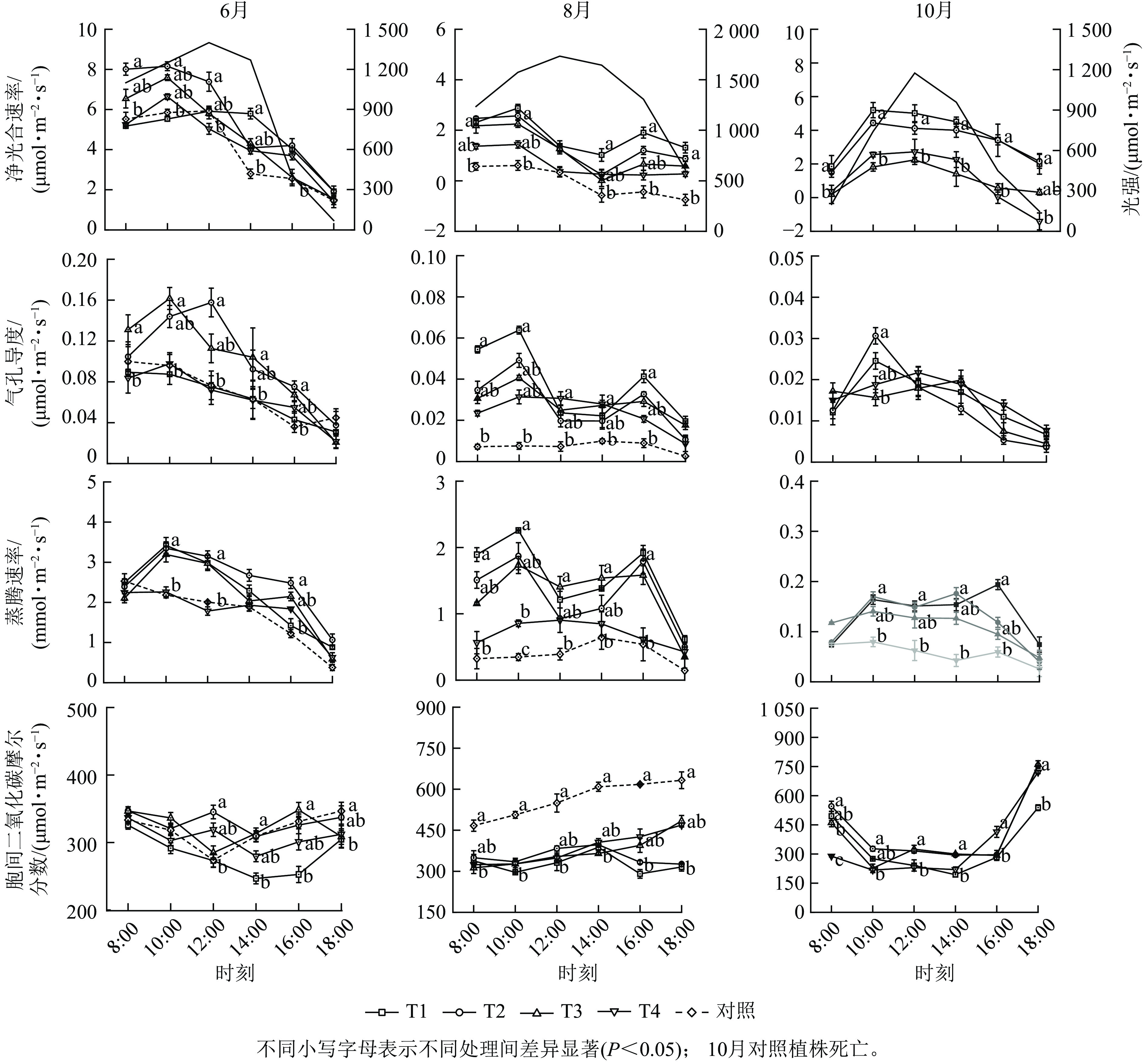
从时间变化来看,不同遮光处理植株的净光合速率、气孔导度和蒸腾速率的最大值均出现在6月,且三者的日变化与外界光强变化趋势相似,呈先升后降的“单峰”曲线(图2),胞间二氧化碳摩尔分数的日变化趋势则相反,细胞间的二氧化碳被充分利用,表明各处理植株的光合系统运转正常。T1、T2和T3处理的净光合速率、气孔导度和蒸腾速率在8月的日变化呈“双峰”曲线(图2),在14:00最低,进入“光合午休”状态,T4和T5处理植株的3个光合参数变化未表现出明显的规律,但对照植株的净光合速率、气孔导度和蒸腾速率远低于遮光处理组(P<0.05),同时胞间二氧化碳摩尔分数显著增高(P<0.05),表明此时的高温和强光胁迫抑制了各处理植株的光合效能,对照植株光合系统受损更严重,这与形态观测结果相符。进入10月后,各遮光处理的植株光合能力逐步恢复,从净光合速率变化来看,T1和T2处理的植株平均光合速率高于T3和T4 (P<0.05),表明遮光度较高(≥61%)的处理对单叶铁线莲植株光合系统的保护更有效。
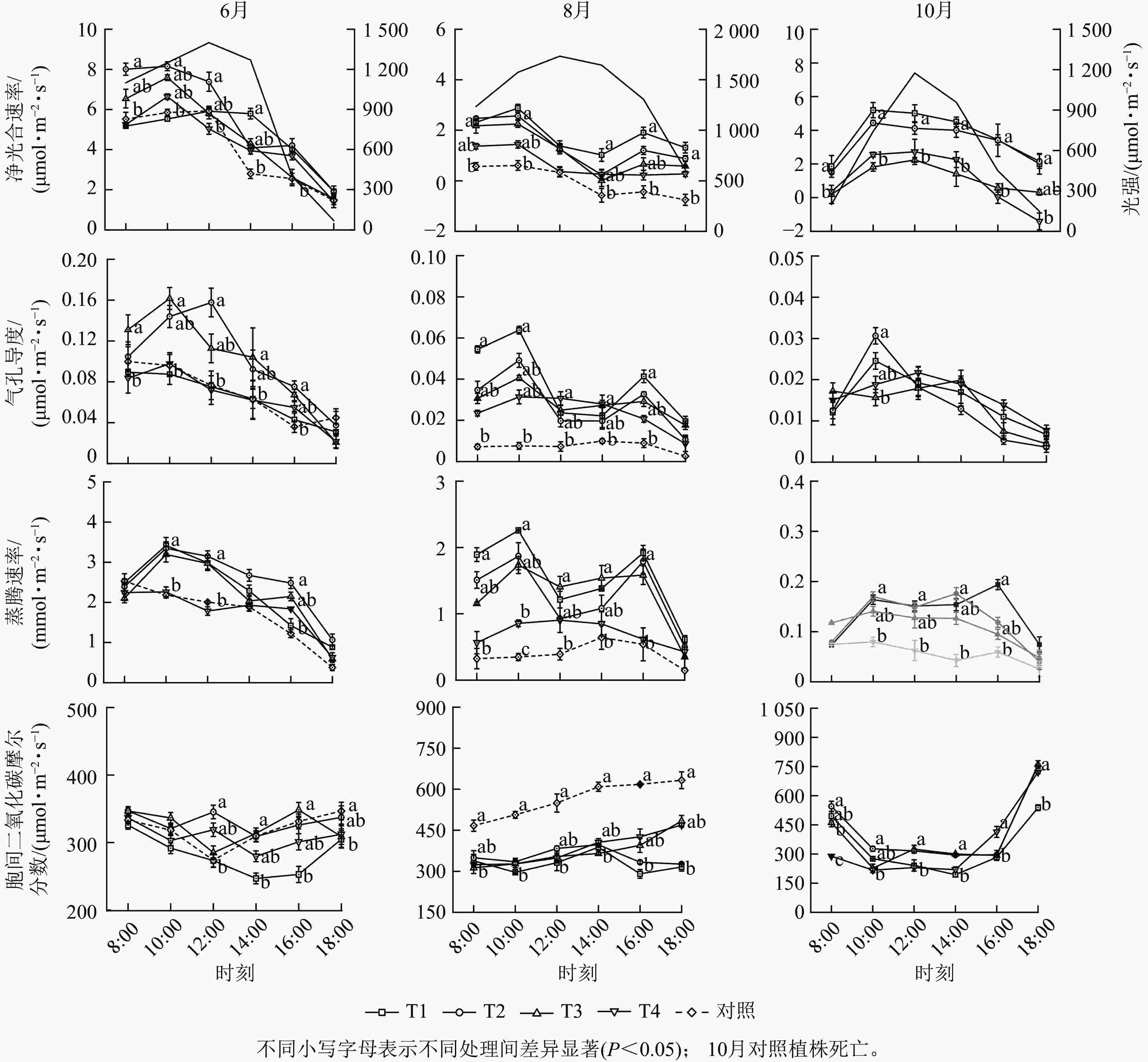
Figure 2. Monthly variations in the daily courses of Pn, Gs, Tr, and Ci of C. henryi under different treatments
-
方差分析和多重比较结果显示:遮光度和时间对表观量子效率、光补偿点、光饱和点、暗呼吸速率、最大净光合速率均产生显著影响(P<0.05)。在同一试验阶段,表观量子效率和最大净光合速率随遮光度增加均呈升高趋势(表3),以6月为例,两者在T1、T2、T3、T4处理中分别是对照组的1.30和1.35,1.37和1.23,1.12和1.12,1.03和1.16倍,其中T1、T2与T4和对照组的差异显著(P<0.05),表明单叶铁线莲有较强的耐荫能力。8月这2个指标分别降至各处理的最低值,相较6月数据,T1、T2、T3、T4处理分别降至原有的0.28和0.45,0.30和0.51,0.15和0.48,0.18和0.42倍,意味着夏季的高温、强光抑制了单叶铁线莲光合系统的运行。对比试验初期和末期的数据,T1、T2和T3植株光补偿点和暗呼吸速率均值光照度增加呈下降趋势,但降幅随遮光度增加而减小,表明在这3种遮光处理下,单叶铁线莲可以通过主动降低光补偿点提高光能利用率,同时减少呼吸作用的营养消耗来适应低光环境,但光饱和点并未表现出规律性的变化。
月份 处理 表观量子效率 /
(μmol·μmol−1)光补偿点 /
(μmol·m−2·s−1)光饱和点 /
(μmol·m−2·s−1)暗呼吸速率 /
(μmol·m−2·s−1)最大净光合速率 /
(μmol·m−2·s−1)6月 T1 0.117±0.01 a 29.35±1.00 b 680.32±9.17 b 0.59±0.03 d 7.83±0.03 a T2 0.123±0.02 a 30.21±2.00 ab 428.25±7.55 c 1.33±0.08 b 7.14±0.16 ab T3 0.101±0.01 ab 32.33±1.73 a 448.36±8.19 c 1.75±0.02 a 6.51±0.01 b T4 0.093±0.02 b 28.65±2.65 b 720.77±13.23 a 0.65±0.04 d 6.74±0.10 b 对照 0.090±0.03 b 34.65±2.65 a 356.52±13.45 d 1.77±0.03 a 5.80±0.26 c 8月 T1 0.033±0.00 a 26.34±2.65 ab 524.24±2.65 a 0.56±0.02 d 3.50±0.04 a T2 0.037±0.00 a 28.58±1.10 a 513.34±4.58 a 1.24±0.02 b 3.63±0.06 a T3 0.015±0.00 b 22.54±2.65 b 320.14±6.50 b 1.47±0.03 a 3.12±0.01 ab T4 0.017±0.00 b 27.55±2.09 ab 240.33±11.36 c 0.64±0.07 c 2.84±0.02 b 对照 0.005±0.00 c 31.45±2.65 a 306.74±9.00 b 1.61±0.03 c 1.84±0.02 c 10月 T1 0.055±0.00 a 26.57±1.13 ab 596.25±8.72 a 0.27±0.02 a 3.82±0.04 a T2 0.051±0.00 a 28.55±2.65 a 504.24±16.37 b 0.11±0.02 b 3.68±0.02 a T3 0.029±0.00 b 22.58±1.73 b 324.11±14.93 c 0.03±0.03 c 3.26±0.07 ab T4 0.025±0.00 b 29.45±1.73 a 248.14±8.00 d 0.13±0.05 b 2.99±0.22 b 说明:数据为平均值±标准差。不同小写字母表示同一月份不同处理间差异显著(P<0.05)。10月对照植株死亡。 Table 3. Photophotoresponse characteristics of C. henryi under different shading treatments in different months
-
方差分析和多重比较结果显示:遮光度和时间对7个叶绿素荧光参数均产生显著(P<0.05)影响(表4)。试验初期至中期,对照和T4处理植株叶绿素荧光参数相较于另外3个处理初始荧光显著升高(P<0.05),电子传递速率显著下降(P<0.05)。试验末期对照植株死亡,T4处理初始荧光和电子传递速率均降至较低水平。T1、T2、T3处理植株初始荧光和电子传递速率在试验的3个时期均保持平稳,表明强光限制了T4和对照叶片内天线色素对光能的捕获能力,对PSⅡ反应中心造成了损伤,而≥53%的遮光度有效保护了叶片光合系统,维持着PSⅡ的光化学效率。在试验中期,各处理植株的最大光化学效率、PSⅡ最大潜在活性、PSⅡ实际光量子产量和光化学猝灭系数均降至最低值(表4),在试验后期又有不同程度回升,且T1、T2和T3处理均显著高于T4处理 (P<0.05),表明强光造成PSⅡ的潜在光化学活性下降,PSⅡ反应中心趋于关闭。随着试验后期胁迫缓解,≥53%的遮光处理下植株叶片光合系统实际量子产能得以有效恢复。此外,T1、T2和T3处理植株的非光化学猝灭系数在试验的3个时期均相对稳定,表明反应中心非光化学猝灭损耗较少,光合系统运转正常。
月份 处理 初始荧光 最大光化学效率 PSⅡ潜在活性 PSII 实际光量子产量 光化学猝灭系数 电子传递速率 非光化学猝灭系数 6月 T1 230.56±14.81 c 0.79±0.00 a 3.71±0.07 a 0.41±0.01 a 0.25±0.04 a 38.58±3.15 a 0.98±0.15 b T2 212.68±20.67 c 0.76±0.04 ab 2.62±0.15 b 0.43±0.03 a 0.22±0.02 a 35.51±0.77 a 1.08±0.08 b T3 231.91±16.79 c 0.75±0.02 ab 2.95±0.34 a 0.40±0.02 a 0.21±0.01 a 33.99±0.62 a 1.28±0.64 b T4 281.90±5.93 b 0.77±0.03 a 2.42±0.07 b 0.37±0.01 b 0.21±0.01 a 26.39±0.75 ab 1.56±0.03 ab 对照 436.67±2.36 a 0.72±0.01 b 2.22±0.16 b 0.33±0.02 b 0.13±0.01 b 16.15±6.40 b 2.93±0.13 a 8月 T1 228.10±5.86 bc 0.73±0.03 a 2.68±0.02 a 0.40±0.01 a 0.20±0.04 a 31.22±1.76 a 1.33±0.03 b T2 204.41±45.48 c 0.53±0.05 b 2.15±0.25 a 0.33±0.01 a 0.15±0.01 b 31.90±0.86 a 1.21±0.12 b T3 205.11±69.04 c 0.59±0.05 b 2.44±0.29 a 0.29±0.01 b 0.15±0.01 b 30.59±1.28 a 1.66±0.15 b T4 321.59±19.21 b 0.39±0.02 c 1.20±0.18 b 0.22±0.02 b 0.11±0.04 b 19.20±4.61 b 2.52±0.25 a 对照 442.17±5.40 a 0.09±0.01 d 0.01±0.00 c 0.02±0.00 c 0.06±0.01 c 2.52±0.13 c 0.46±0.01 c 10月 T1 207.07±7.89 b 0.75±0.01 a 2.97±0.09 a 0.40±0.01 a 0.21±0.01 a 37.08±0.73 a 1.34±0.04 b T2 211.45±16.49 b 0.76±0.02 a 3.02±0.28 a 0.35±0.01 ab 0.20±0.01 a 36.24±1.75 a 1.13±0.19 b T3 236.24±21.45 a 0.74±0.01 a 2.92±0.22 a 0.29±0.02 b 0.16±0.01 ab 29.49±1.18 b 2.24±0.14 a T4 104.50±6.89 c 0.53±0.01 b 1.83±0.08 b 0.10±0.02 c 0.11±0.01 c 19.58±2.47 c 2.62±0.15 a 说明:不同小写字母表示不同处理间差异显著(P<0.05)。10月对照植株死亡。 Table 4. Chlorophyll fluorescence parameters of C. henryi in different months in different treatments
-
叶片是植物对光照环境变化最敏感的器官,其形态结构特征最能体现植物对光环境的适应能力[15]。本研究的3个阶段中,遮光处理下植株叶面积、节间长度、叶片干质量等6个指标的平均值都高于对照,说明单叶铁线莲通过增加叶片长和宽获得更大的叶面积,提高节间长度扩展生长空间,以此获得更多的光照资源,促进光合有机产物积累。在低光照条件下,玉簪Hosta plantaginea通过调整自身形态来增加对光照的截获概率,表现出叶面积增大、茎秆伸长等特点[16];绿萝Epipremnum aureum在荫蔽环境中也会通过增加叶面积和节间长度来应对低光胁迫,保障自身的有机物质积累[17]。这些结果都与单叶铁线莲在遮光处理下的生长调节策略相似。
随着遮光度增加,单叶铁线莲通过减少叶片总厚度,抑制栅栏组织生长和扩展海绵组织空间,增加细胞间隙,提高光能利用率。这种主动适应荫蔽环境的叶片生长调节机制在金莲花Trollius chinensis[18]和黑茶藨子Ribes nigrum[19]中也有报道。有研究表明:玉簪在遮光环境下叶片干质量和含水量有显著变化,尤其是在夏季,遮光能更好地维持叶片的水分状况和干物质积累速率[16]。鸟巢蕨Asplenium nidus在遮光度增加时,叶片干质量和含水量也随之增加,在夏季这种变化更显著[20]。与玉簪和鸟巢蕨2种植物类似,单叶铁线莲叶片干质量和含水量随着遮光度提高而增加,且远高于对照组。这种差异在8月更为明显。这一结果说明单叶铁线莲具有很强的耐荫性,在荫蔽环境中能保持光合系统高效运转,为植株生长提供保障。
-
植物叶片的光合参数能反映生理代谢和物质积累的能力,也被用于分析环境因子对植物代谢和生长的影响[21]。有报道显示:“光合午休”现象主要是气孔和非气孔限制因素导致[22−23]。当胞间二氧化碳摩尔分数与气孔导度同时下降时,净光合速率下降主要是由气孔限制引起,反之则是非气孔因素[24]。本研究中,各处理植株8月的净光合速率日变化曲线均呈“双峰”,是典型的“光合午休”表现。10:00后外界光强和气温快速升高,各处理组净光合速率持续下降,气孔导度也均随之降低,此时,T1、T2处理的胞间二氧化碳摩尔分数下降,但T3、T4和对照组则上升,表明T1、T2处理植株的 “光合午休”是气孔关闭所致。T3、T4和对照组植株的净光合速率下降则是非气孔因素造成的,可能是过剩的光能促进了活性氧积累,导致脂质过氧化,植物膜系统受损,光合电子传递系统遭到破坏所致[25]。现有研究也表明:植物在光照轻度胁迫下,光合速率的降低是由于气孔限制值降低引起的;在中度或严重胁迫下,主要是非气孔因素造成的。2种因素对光合作用影响的大小取决于光照胁迫的程度[26]。由此可见:T1和T2处理有效缓解了夏季高温、强光对单叶铁线莲光合生理系统的伤害。有研究表明:光补偿点和光饱和点的变化不仅与植物的种类、环境因素有关,还与植物自身的生理调节机制密切相关[27−28]。6和8月的数据显示,遮光处理的单叶铁线莲植株光补偿点均低于对照,T1处理光补偿点平均值为24.50 μmol·m−2·s−1,表明单叶铁线莲可以通过主动降低光补偿点来适应弱光环境。这与白及Bletilla striata耐荫性研究结果[29]相似。
-
初始荧光表示叶片PSⅡ反应中心处于完全开放时的荧光水平。本研究初期(6月)对照的初始荧光较处理组显著上升,说明全光照下单叶铁线莲PSⅡ反应中心天线的热耗散程度较大;随后8月高温强光使叶片PSⅡ反应中心出现不可逆的失活,开放程度不断降低,初始荧光也大幅降低,可能是叶片类囊体膜受到损伤所致[30]。电子传递速率反映实际光强条件下的电子传递效率,光化学猝灭参数代表了光合能量用于暗反应固定能量的部分[31]。本研究初期和中期,遮光提高了单叶铁线莲电子传递速率,电子传递速与光化学猝灭参数随遮光度增加而增加,说明单叶铁线莲通过增加PSⅡ反应中心的开放比例,促进电荷分离,光合暗反应得以顺利进行。这与番茄Solanum lycopersicum幼苗应对高光胁迫的机制类似[32]。
-
单叶铁线莲具有很强耐荫性,遮光处理促进了叶面积、节间距、叶片干质量、叶片含水量的增加,植株通过增加叶片海栅比,降低光饱和点和减少呼吸作用的营养消耗来适应低光环境。夏季高温、强光对单叶铁线莲生长产生显著负面影响,61%~82%遮光处理可保护单叶铁线莲叶片光合系统免受不可逆的损伤。单叶铁线莲可以通过提高光能转换效率适应弱光环境,维持PSⅡ反应中心正常运转,有效保障植株的正常生长。上述研究结果为单叶铁线莲在园林中的引种、栽培提供了依据。此外,单叶铁线莲的耐荫特性将有效扩展铁线莲属植物的园林应用范围,为营造林下、北侧阳台和室内空间植物景观提供了更多选择。
Effect of shading on the growth and photosynthetic characteristics ofClematis henryi
doi: 10.11833/j.issn.2095-0756.20240625
- Received Date: 2024-11-05
- Accepted Date: 2025-03-10
- Rev Recd Date: 2025-03-02
- Available Online: 2025-11-26
- Publish Date: 2025-12-20
-
Key words:
- Clematis /
- Clematis henryi /
- shading /
- photosynthetic characteristics /
- chlorophyll fluorescence
Abstract:
| Citation: | ZHAO Changgao, CHEN Zihang, ZHANG Chengchai, et al. Effect of shading on the growth and photosynthetic characteristics ofClematis henryi[J]. Journal of Zhejiang A&F University, 2025, 42(6): 1174−1183 doi: 10.11833/j.issn.2095-0756.20240625

|
 DownLoad:
DownLoad: