






-
植物功能性状是植物生命周期中获取、利用和保留资源等一系列活动的关键指示,是植物对外界环境长期适应和演化的结果,能够在一定程度上反映植物对环境资源的利用情况[1−3]。当外界环境发生变化时,为维持体内环境的相对稳定,植物功能性状会进行适应性调整,以适应复杂多变的环境[4]。叶片作为植物进行光合作用、呼吸作用和蒸腾作用的核心器官,是植物与外界进行水、气、热交换的关键门户,对外界环境反应最为敏感[5]。通过对大量植物叶功能性状进行研究,不仅可以表征植物物种间的差异,还可进一步揭示和推测植物对环境的适应及生长策略[6]。因此,从叶功能性状视角出发,研究植物叶功能性状特征及其权衡关系已经成为植物生态学研究的热点领域之一[7]。
近年来,大量研究通过分析植物叶功能性状的变异特征及其资源权衡分析,深入揭示了植物在资源获取与利用之间的策略选择[8−9]。WRIGHT等[10]对全球范围内多个物种的叶功能性状进行了研究,在全球尺度上首次揭示了叶性状组合之间的相互联系和协同变化规律,并提出了叶经济谱理论。叶经济谱理论认为叶功能性状沿着一条连续变化的资源梯度有序排列,并表现出2种截然不同的叶性状变化策略:资源轴的一端表现为物种具有较长的叶片寿命、较低的叶氮和叶磷、低的光合速率、高的干物质和低的比叶面积,被划分为“缓慢投资-收益”型策略(保守型策略);在资源轴的另一端则表现为短的叶片寿命、较高的叶氮和叶磷、高的光合速率、低的干物质和高的比叶面积,属于“快速投资-收益型”策略(获取型策略)[11−13]。因此,植物通过叶性状间的协作与权衡来实现对环境变化的响应与适应[14]。目前,对植物叶功能性状适应策略的研究多集中于草本植物,有关木本植物的研究相对较少,且多以跨科、跨属的大范围研究为主[15−17],对小尺度范围特别是属内木本植物在相似演化背景下,如何通过精细的叶功能性状权衡适应小尺度生境的认识有限,也难以完善从大尺度到小尺度的性状权衡理论体系。因此,深入研究植物叶功能性状关系有助于揭示属内植物的资源分配方式。
杜鹃花属Rhododendron隶属于杜鹃花科Ericaceae,是北半球最大的木本植物属,有1 000多个物种,广泛分布于亚洲、欧洲和北美洲,其中以亚洲分布的种类最多[18−19]。中国杜鹃花属植物已超600种,以云南、西藏、四川所在的横断山脉分布最多[20−21]。杜鹃花属植物大多姿态优美、花径大而颜色娇艳,具有很高的观赏、审美和经济价值[22−23]。同时,杜鹃花属植物在自然状态下常常形成大规模的灌木丛,根系发达、保水能力强,在山地生态系统的稳定与流域保护等方面发挥着极其重要的作用[24]。目前对色季拉区域杜鹃花的研究主要集中在解剖结构[25−26]、凋落物分解[27]、亲缘关系等[28]多个方面,对杜鹃花属植物叶功能性状特征及其资源权衡的研究鲜有报道,因此本研究以色季拉山13种杜鹃花属植物为研究对象,测定其16个叶功能性状,分析杜鹃花属植物生长及资源分配过程中的生长策略,旨在丰富该区域植物叶功能性状研究,为该区域生物多样性保护和管理提供理论依据。
-
研究区色季拉山位于青藏高原东南部,是念青唐古拉山向南延伸的余脉,整体呈现西北—东南走向,海拔2 500~5 430 m。该研究区受印度洋季风的影响,干湿分明,植被垂直分布明显,植被类型丰富,人为干扰较少[29−30]。年降水量为1 134.0 mm,6—9月为雨季,约占全年总降水量的80%,年均相对湿度为78.8%,年平均气温为−0.7 ℃,7月最热月平均气温为9.2 ℃,1月最冷月平均气温为−14.0 ℃,年日照时数为1 151.0 h[31]。该区除杜鹃花属植物外,木本植物主要有急尖长苞冷杉Abies georgei var. smithii、方枝柏Juniperus saltuaria、林芝云杉Picea likiangensis var. linzhiensis、川滇高山栎Quercus aquifolioides等[29, 32]。石砾较多,土壤以山地棕壤和山地酸性棕壤为主,以酸性和强酸性为主[33−34]。
-
2024年5—6月和2025年3—6月,在全面踏查的基础上,在色季拉山选取典型杜鹃花属植物群落,在其中随机布设3个20 m×20 m样地,在每个样地内通过对角线法设置3个5 m×5 m的样方进行杜鹃花属植物调查[35−36]。植物群落常规调查时,记录样方中植物的名称、株高、盖度、数量等相关数据,同时利用便携式全球定位系统(GPS)记录大样方的经度、纬度和海拔(表1),同时采集所有杜鹃属植物标本带回实验室,由植物分类专业人员对照《中国植物志》和《西藏植物志》进行鉴定,确定植物学名,杜鹃花标本保存于西藏农牧大学高原生态学院(研究所)标本室。根据色季拉山杜鹃花属植物的调查结果,共选取其中13种杜鹃花属植物作为研究对象,分别是三花杜鹃R. triflorum、硬毛杜鹃R. hirtipes、栎叶杜鹃R. phaeochrysum、山育杜鹃R. oreotrephes、黄杯杜鹃R. wardii、雪层杜鹃R. nivale、林芝杜鹃R. nyingchiense、草莓花杜鹃R. fragariiflorum、白毛杜鹃R. vellereum、光蕊杜鹃R. coryanum、柳条杜鹃R. virgatum、鳞腺杜鹃R. lepidotum和鲁朗杜鹃花R. lulangense。
序号 物种 北纬(N) 东经(E) 海拔/m 1 光蕊杜鹃 29°53′08.03″ 94°47′12.84″ 2782 2 三花杜鹃 29°51′25.61″ 94°46′12.83″ 2911 3 鲁朗杜鹃花 29°37′15.07″ 94°42′33.44″ 4208 4 硬毛杜鹃 29°38′05.96″ 94°42′56.03″ 4105 5 栎叶杜鹃 29°38′08.53″ 94°42′47.53″ 4096 6 山育杜鹃 29°39′56.76″ 94°42′55.21″ 3720 7 黄杯杜鹃 29°39′31.84″ 94°43′25.55″ 3804 8 雪层杜鹃 29°41′49.84″ 94°43′45.34″ 3434 9 林芝杜鹃 29°37′24.57″ 94°37′49.00″ 4177 10 白毛杜鹃 29°33′32.74″ 94°34′25.72″ 3830 11 草莓花杜鹃 29°36′56.89″ 94°39′36.24″ 4462 12 柳条杜鹃 29°52′34.75″ 94°46′40.06″ 2805 13 鳞腺杜鹃 29°38′44.23″ 94°42′10.99″ 3937 Table 1. Sampling site information for Rhododendron in the Shergyla Mountain
对样地内杜鹃花属植物叶片进行采集时,在每种杜鹃花(指同一物种不同个体,不含变种)中选择5株生长均一、株高相近的成熟健康植株作为标准株,在冠层中部相同高度不同方位采集生长均匀、完全展开、无病虫害、无损伤的25片叶片,其中15~20片用湿润滤纸覆盖并装入提前做好标记的自封袋中,带回实验室处理[4]。另外5~10片叶片,用剪刀修剪后立即放入事先准备好的乙醇醋酸福尔马林(FAA)混合固定液(武汉赛维尔生物科技有限公司)中保存,带回实验室备用[21]。同时采集400~500 g叶片混合样品,用于后续叶片碳、氮、磷质量分数测定。本研究测定的植物叶功能性状指标:叶面积(mm)、叶厚度(mm)、叶干质量(g)、叶组织密度(g·cm−3)、叶干物质质量分数(g·g−1)、叶形指数、叶片含水量(%)、比叶面积(cm2·g−1)、叶碳质量分数(g·kg−1)、叶氮质量分数(g·kg−1)、叶磷质量分数(g·kg−1)、叶碳氮比、叶碳磷比、叶氮磷比、栅栏组织厚度(μm)、海绵组织厚度(μm)。
-
每种杜鹃花属植物随机选取8片叶片,选用电子数显游标卡尺(精度为0.01 mm)在叶片沿主脉方向,避开叶片主脉,随机选择3个部位进行测量,取3次测量的平均值作为叶厚度。使用叶面积仪(LA-S系列植物图像分析仪系统,杭州万深检测科技有限公司)测定杜鹃花属植物的叶面积、叶长和叶宽。将扫描完的叶片用电子天平(精确到0.000 1 g)称量,得到叶鲜质量,随后在120 ℃下杀青30 min,65 ℃烘干72 h至质量恒定,称取叶干质量[37]。最后参考文献[38−39]计算比叶面积、叶干物质质量分数、叶组织密度、叶形指数和叶含水量。
-
取出经FAA固定液固定后的杜鹃花属植物叶片,经蒸馏水冲洗数次后,用刀片切取宽约0.5 cm × 0.5 cm的叶片,经不同浓度梯度的乙醇脱水、二甲苯透明、石蜡浸蜡、石蜡包埋、全自动石蜡切片机(ERM 4000)切割叶片(厚度为10 µm)、番红-固绿染色、中性树胶封片[40]。每个物种制作封片5张。待封片干燥后用显微镜(尼康ECLIPSE 80i)随机拍摄5张照片,用Leica Application Suite X 软件测定栅栏组织厚度和海绵组织厚度。每项指标重复测定5次。
-
用球磨仪将烘干后的植物叶片磨碎,过100目筛,采用元素分析仪(Vario Max CN Element analyzer,Elementar)测定叶碳质量分数和叶氮质量分数,采用钼锑比色法测定叶磷质量分数[8,15]。
-
使用Excel 2010对杜鹃花属植物的各项功能性状数据进行统计整理。首先使用SPSS 26对各项功能性状求平均值和标准差,计算变异系数(VC)=(标准差/平均值×100%),评估各功能性状的变异程度。用Pearson相关性分析法评估各功能性状间的关系。采用主成分分析法(PCA)对16个叶功能性状进行综合分析。利用Origin 2021软件绘图。
-
由表2可知:色季拉山杜鹃花属植物叶功能性状的变异系数为10.67%~120.29%,变异系数由大到小依次为叶干质量(120.29%)、叶面积(96.28%)、比叶面积(48.81%)、海绵组织厚度(47.31%)、栅栏组织厚度(33.68%)、叶磷质量分数(27.25%)、叶氮质量分数(27.00%)、叶形指数(25.73%)、叶碳磷比(25.22%)、叶碳氮比(24.22%)、叶厚度(22.93%)、叶氮磷比(20.20%)、叶组织密度(18.88%)、叶干物质质量分数(16.22%)、叶含水量(14.61%)、叶碳质量分数(10.67%)。其中叶干质量和叶面积属于强变异,比叶面积、海绵组织厚度、栅栏组织厚度、叶磷质量分数、叶氮质量分数、叶形指数、叶碳磷比、叶碳氮比、叶厚度、叶氮磷比属于中等变异,其余指标属于弱变异。杜鹃花属植物叶性状种间变异普遍高于种内变异。
项目 叶面积/mm2 叶厚度/mm 叶干质量/g 叶组织密度/
(g·cm−3)叶干物质/(g·g−1) 叶形指数 叶含水量/% 比叶面积/(cm2·g−1) 平均值±标准差 1 959.13±1 886.28 0.31±0.07 0.340 0±0.410 0 0.47±0.09 0.47±0.08 2.20±0.56 52.61±7.69 74.43±36.33 最大值 8 695.56 0.47 2.320 0 0.63 0.63 4.48 79.44 269.28 最小值 36.88 0.17 0.003 8 0.22 0.21 1.31 36.97 31.87 变异系数/% 96.28 22.93 120.290 0 18.88 16.22 25.73 14.61 48.81 变异强度 强变异 中等变异 强变异 弱变异 弱变异 中等变异 弱变异 中等变异 项目 叶碳/(g·kg−1) 叶氮/(g·kg−1) 叶磷/(g·kg−1) 叶碳氮比 叶碳磷比 叶氮磷比 栅栏组织厚度/μm 海绵组织厚度/μm 平均值±标准差 535.91±57.16 15.04±4.06 0.88±0.24 37.68±9.13 647.73±163.38 17.47±3.53 116.44±39.21 132.80±62.83 最大值 686.64 21.79 1.36 62.60 1 059.09 25.35 196.03 371.07 最小值 401.20 9.49 0.53 24.92 401.89 10.70 49.79 35.55 变异系数/% 10.67 27.00 27.25 24.22 25.22 20.20 33.68 47.31 变异强度 弱变异 中等变异 中等变异 中等变异 中等变异 中等变异 中等变异 中等变异 说明:变异系数≥50%为强变异,20%≤变异系数<50%为中等变异,变异系数<20%为弱变异[41]。 Table 2. Statistical characteristics of leaf functional traits for Rhododendron on Shergyla Mountain
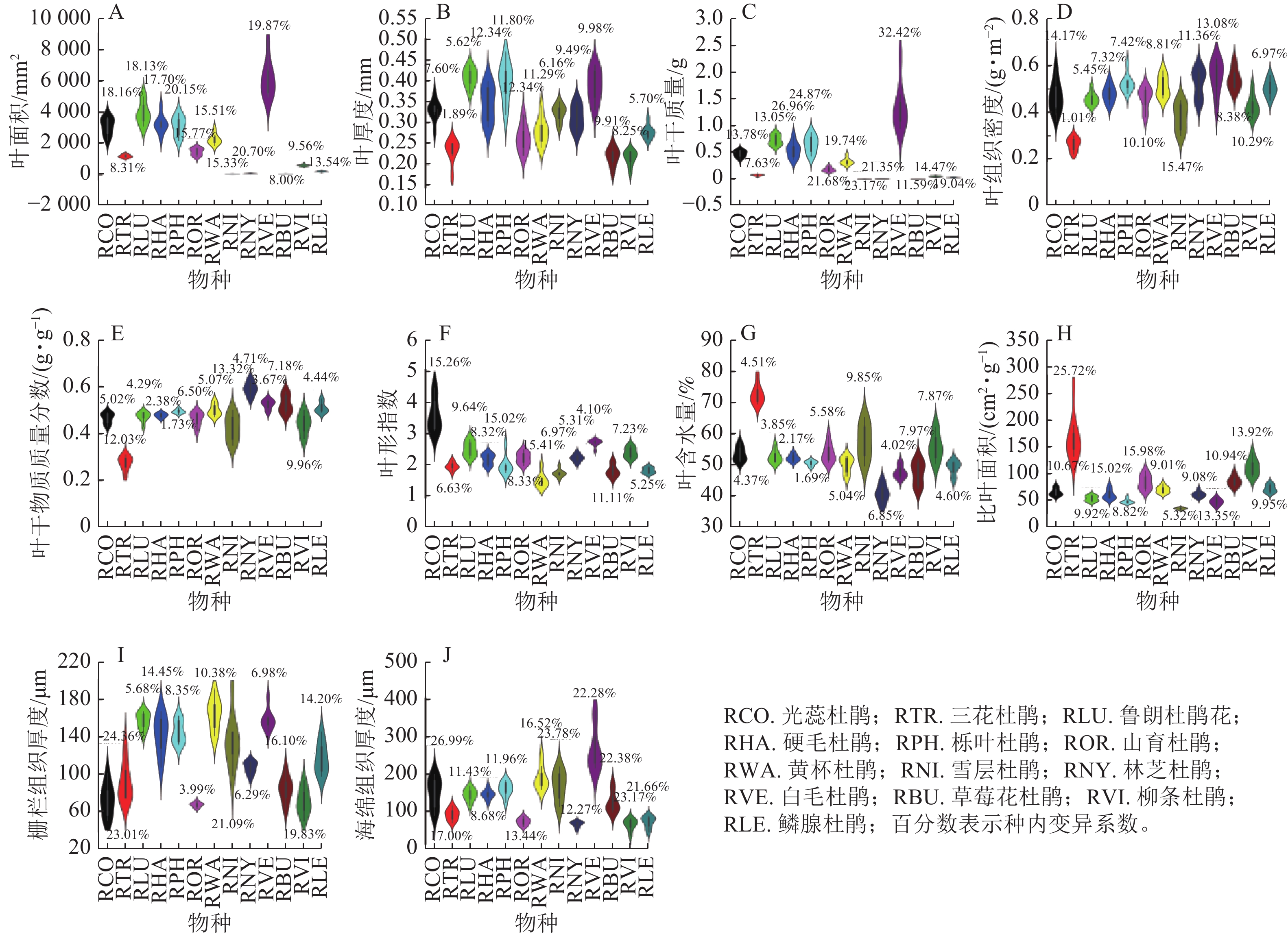
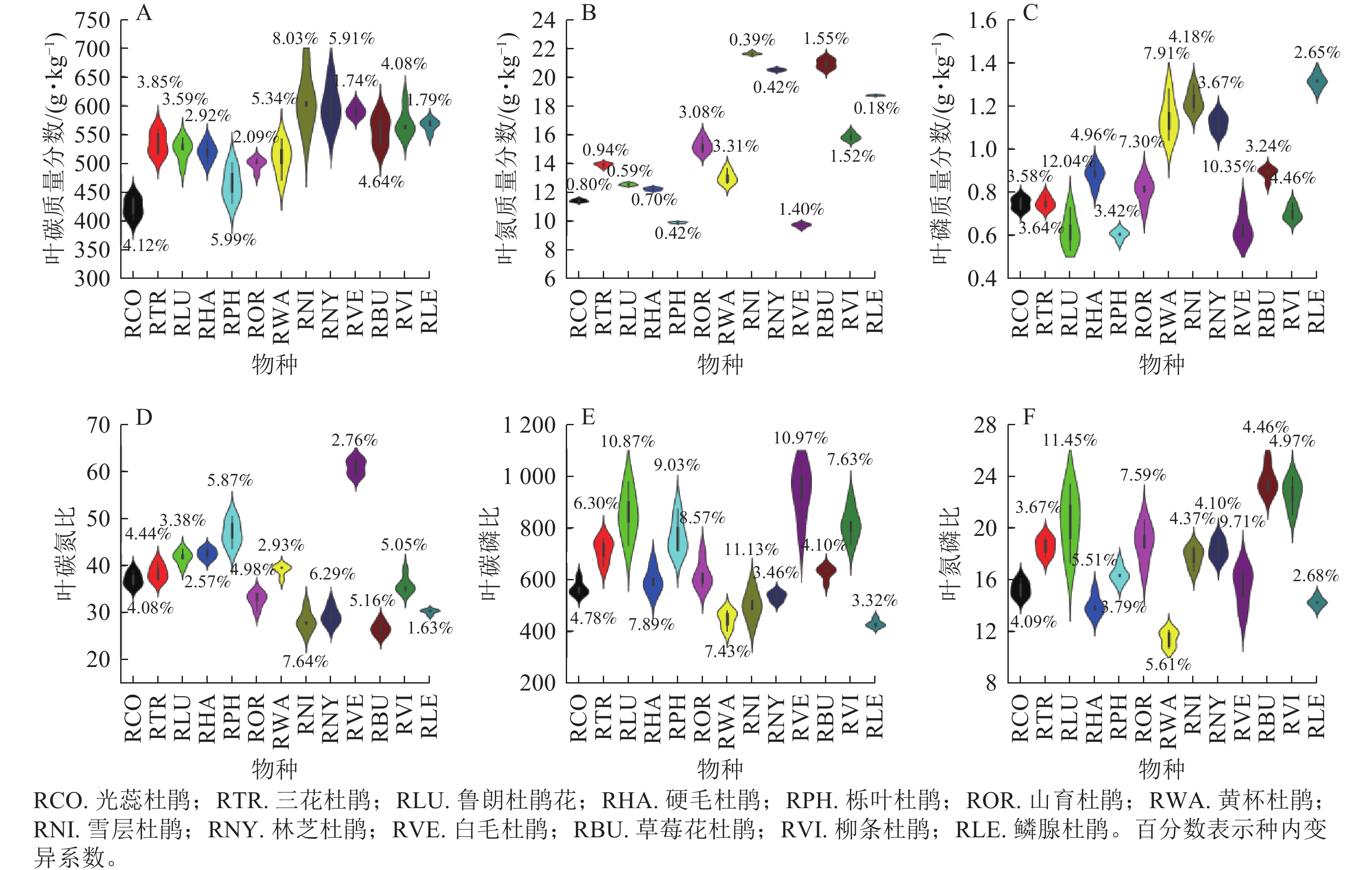
在物种水平上,色季拉山13种杜鹃花属植物的叶功能性状差异明显。在结构性状方面(图1),白毛杜鹃叶面积最大(6 178.94 mm2,变异系数为19.87%),林芝杜鹃叶面积为65.26 mm2,变异系数为20.70%。鲁朗杜鹃花叶厚度(0.41 mm)最大,栎叶杜鹃次之(0.40 mm),所有物种叶厚度的变异系数均小于20.00%,属于弱变异。白毛杜鹃叶干质量最大(1.340 0 g,变异系数为32.42%),草莓花杜鹃和雪层杜鹃叶干质量最小,分别为0.006 1 g (变异系数为11.59%)和0.005 6 g (变异系数为23.17%)。白毛杜鹃叶组织密度最大,为0.55 g·cm−3,而雪层杜鹃叶组织密度变异系数最大,为15.47%。林芝杜鹃叶干物质质量分数最大,为0.59 g·g−1,光蕊杜鹃和鲁朗杜鹃花叶干物质质量分数均为0.47 g·g−1,雪层杜鹃叶干物质质量分数变异系数最大,为13.32%。光蕊杜鹃叶形指数最大,达到3.59,山育杜鹃和林芝杜鹃叶形指数均为2.22。三花杜鹃叶含水量最大,为72.73%,林芝杜鹃叶含水量最小,为40.76%,雪层杜鹃叶含水量变异系数最大,为9.85%。三花杜鹃比叶面积最大(169.75 cm2·g−1,变异系数为25.72%),是比叶面积最小的雪层杜鹃的3.89倍。黄杯杜鹃栅栏组织厚度最大,为164.56 μm,山育杜鹃最小,为67.58 μm,且山育杜鹃栅栏组织的变异系数最小,为3.99%。白毛杜鹃海绵组织厚度最大,为267.33 μm,柳条杜鹃海绵组织厚度最小,为66.02 μm,光蕊杜鹃海绵组织厚度变异系数最大,硬毛杜鹃海绵组织厚度变异系数最小,为8.68%。
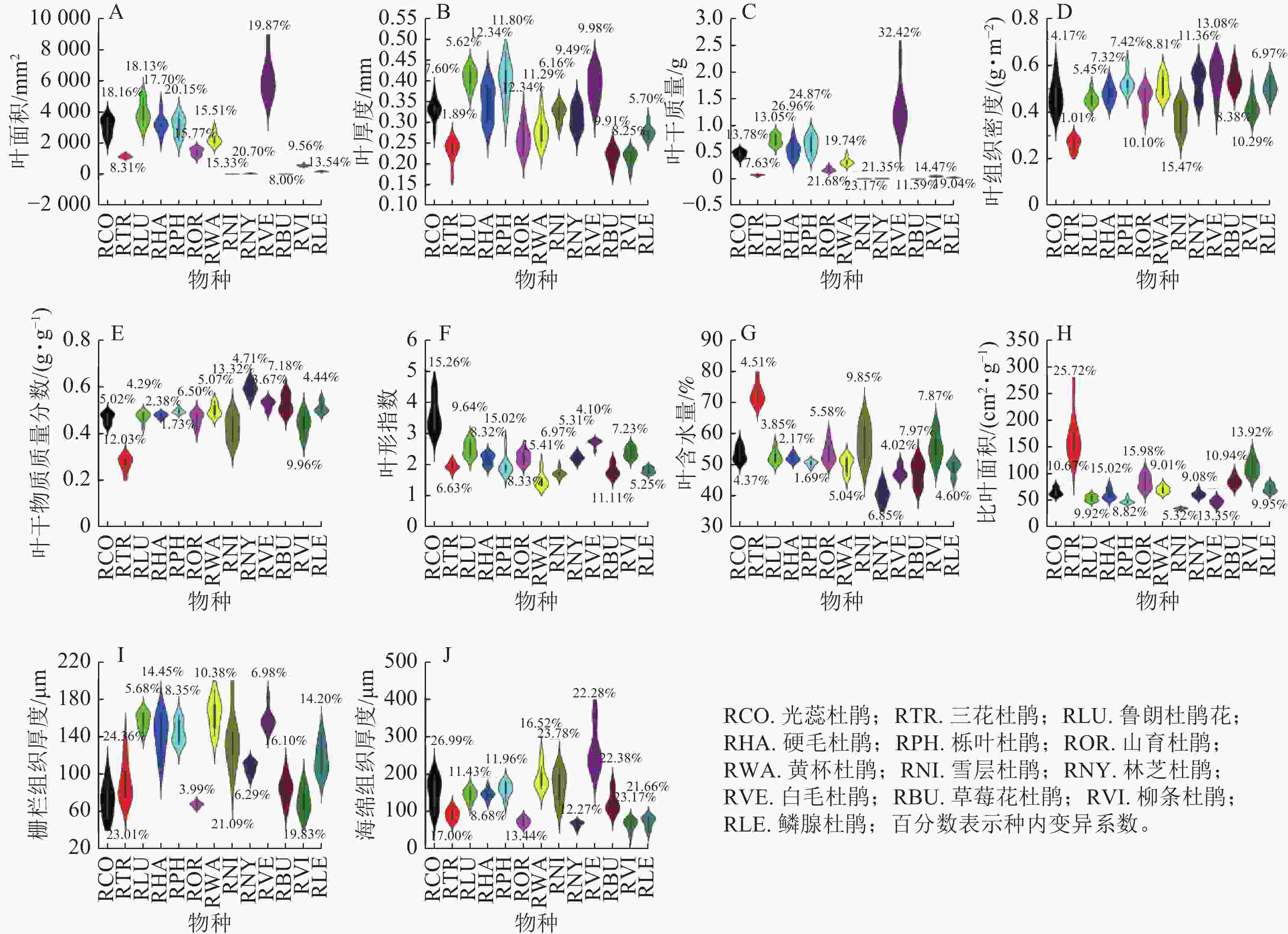
Figure 1. Leaf structural traits of Rhododendron
在化学性状方面(图2),雪层杜鹃叶碳质量分数最大(609.67 g·kg−1),所有变异系数均小于10.00%。雪层杜鹃叶氮质量分数最大(21.65 g·kg−1),是叶氮质量分数最小的白毛杜鹃的1.23倍;黄杯杜鹃叶氮质量分数变异系数最大,为5.44%。鳞腺杜鹃叶磷质量分数最大(1.31 g·kg−1),光蕊杜鹃和三花杜鹃叶磷质量分数均为0.75 g·kg−1,鲁朗杜鹃花叶磷质量分数变异系数最大,为12.04%。栎叶杜鹃叶碳氮比最大(46.95,变异系数为5.87%),草莓花杜鹃叶碳氮比最小,为26.60。白毛杜鹃叶碳磷比最大,为942.98,是叶碳磷比最小的鳞腺杜鹃的1.19倍,雪层杜鹃叶碳磷比变异系数最大,为11.13%。草莓花杜鹃叶氮磷比最大,为23.65,而鳞腺杜鹃叶氮磷比变异系数最小,为2.68%。
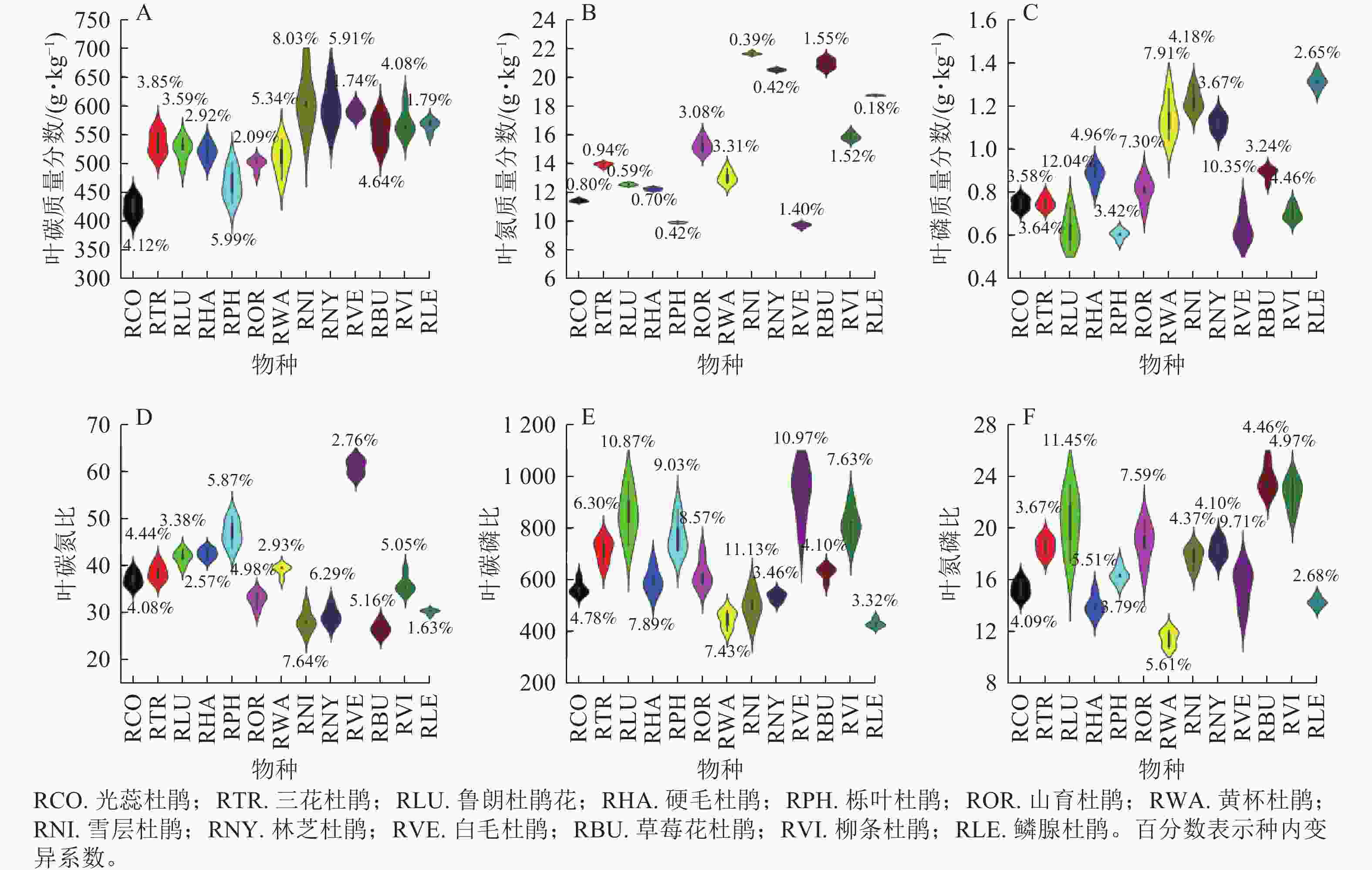
Figure 2. Leaf chemical traits of Rhododendron
-
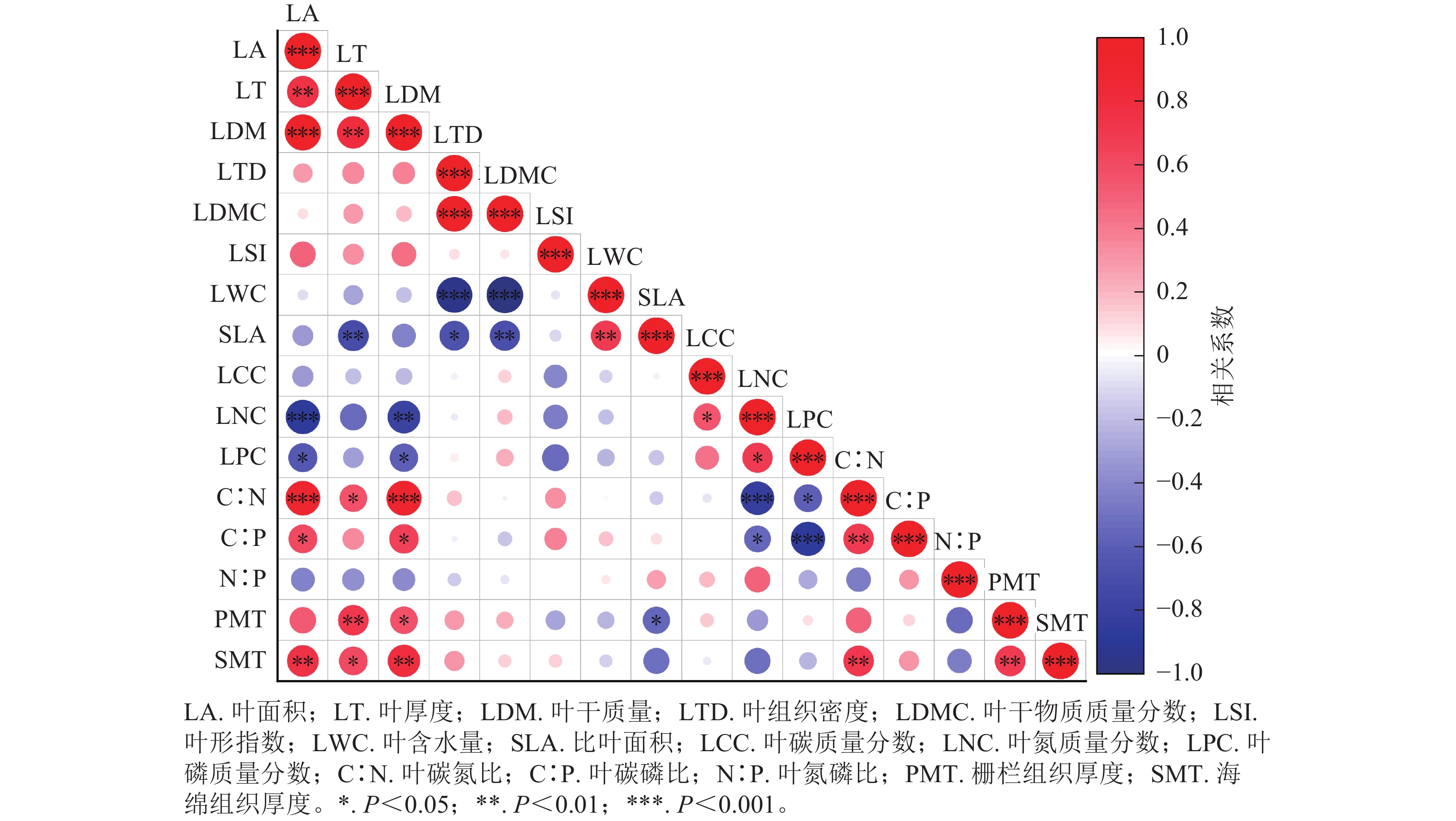
相关性矩阵结果如图3所示。杜鹃花属植物不同的叶功能性状间存在着不同的相关关系,其中比叶面积与叶干物质质量分数、叶厚度呈极显著负相关(P<0.01),与叶组织密度呈显著负相关(P<0.05),与叶含水量呈极显著正相关(P<0.01)。比叶面积与叶面积呈极显著正相关(P<0.01),叶干质量与叶面积呈极显著正相关(P<0.001),叶含水量与叶干物质质量分数、叶组织密度呈极显著负相关(P<0.001)。叶氮质量分数与叶磷质量分数呈显著正相关(P<0.05),叶碳氮比与叶面积、叶干质量呈极显著正相关(P<0.001)。栅栏组织厚度与叶厚度呈极显著正相关(P<0.01)。
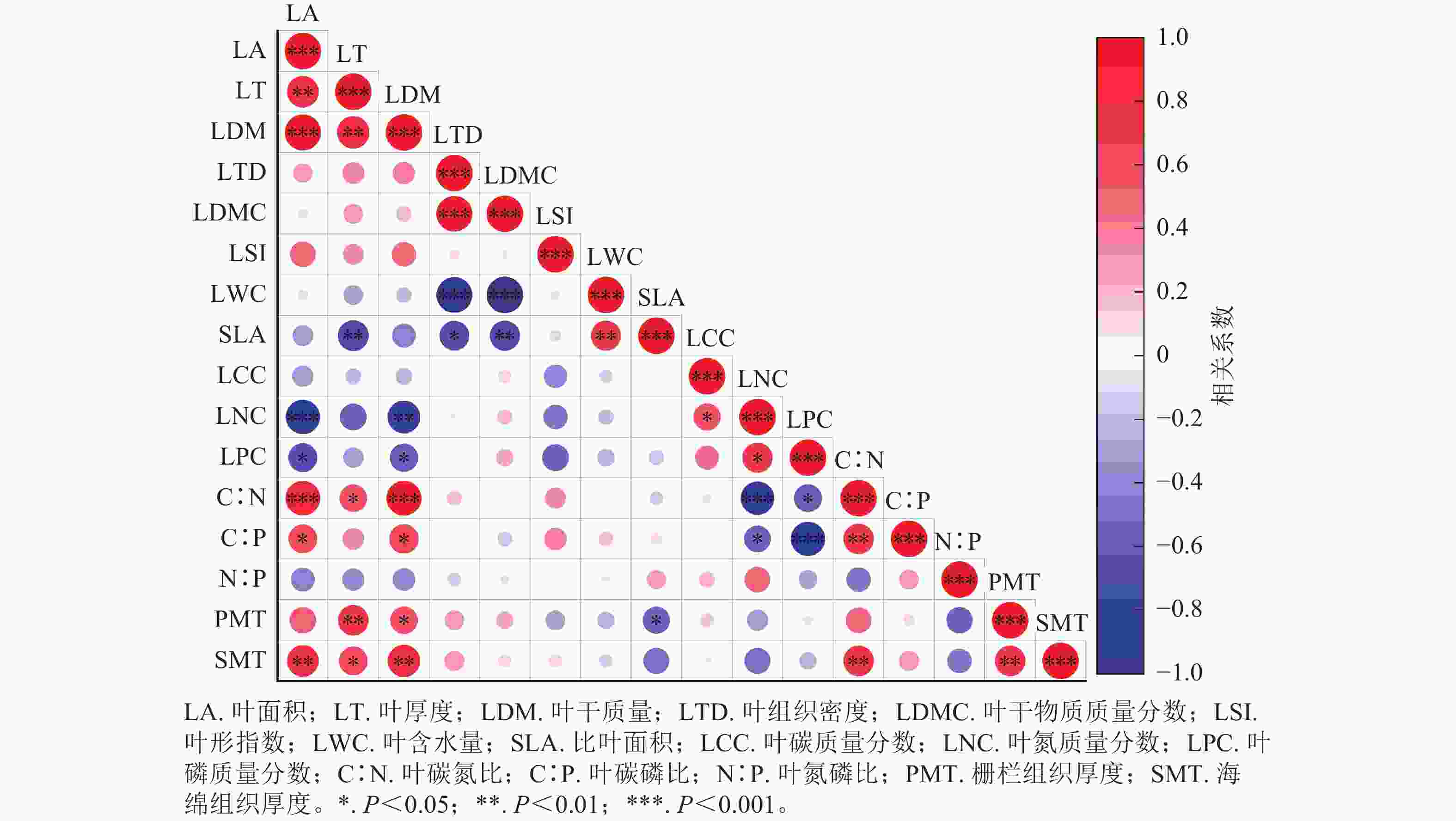
Figure 3. Correlation between leaf functional traits of Rhododendron
-
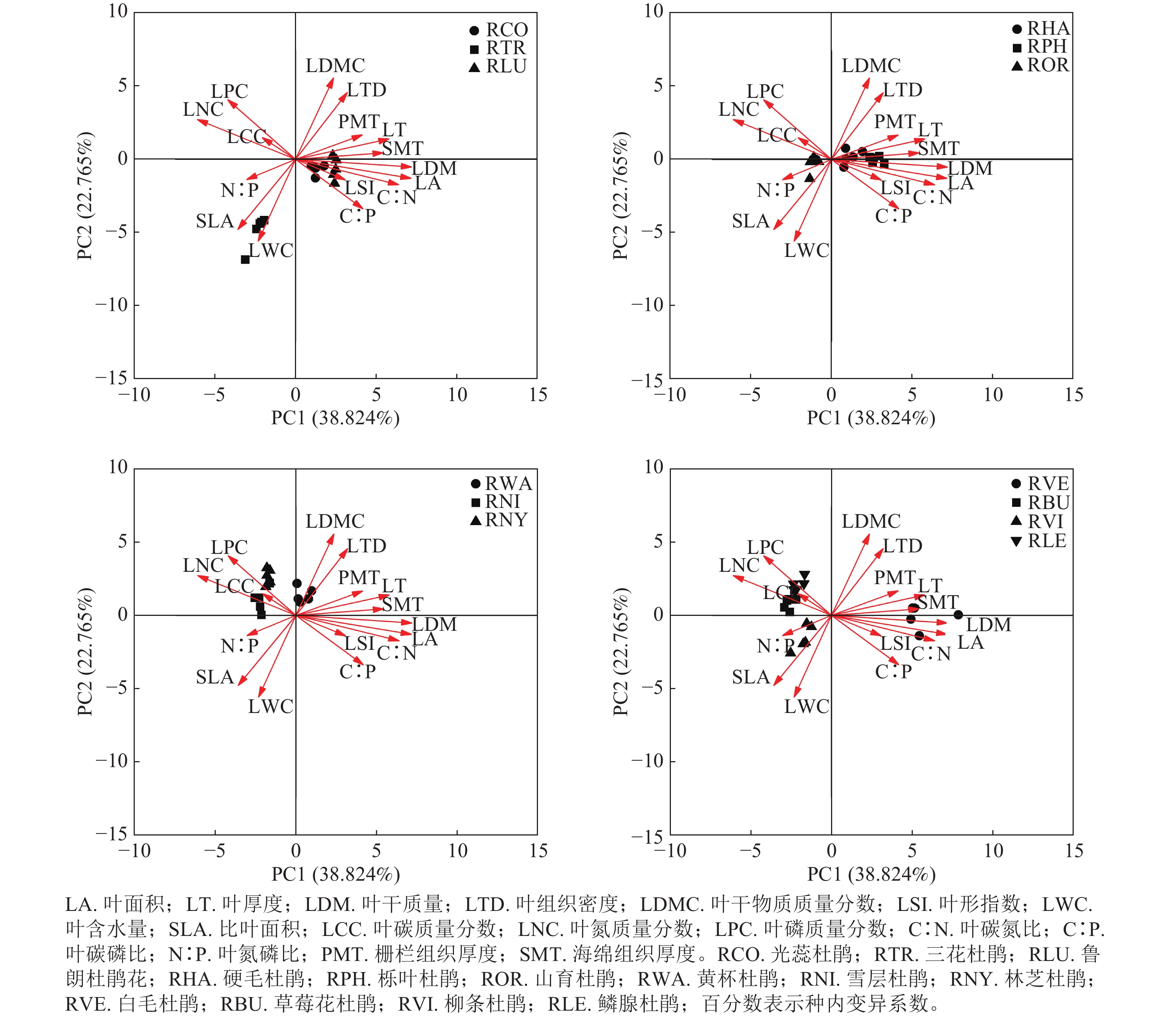
主成分分析表明:13种杜鹃花属植物的叶功能性状之间存在一定的相关性(表3和图4)。杜鹃花属植物叶功能性状前4个主成分的特征值分别为6.212、3.642、2.014和1.493,贡献率分别为38.824%、22.765%、12.585%和9.328%,累计贡献率为83.502%,表明这4个主成分是影响杜鹃花叶片功能性状变化的主要因素。主成分1与叶干质量、叶面积、叶碳氮比、叶厚度、叶氮质量分数相关性较强,主成分2与叶干物质质量分数、叶组织密度、叶含水量、比叶面积相关性较强,主成分3与叶氮磷比、栅栏组织厚度相关性较强,主成分4与叶碳质量分数、叶碳磷比相关性较强。在本研究中,山育杜鹃、草莓花杜鹃、鳞腺杜鹃、雪层杜鹃和林芝杜鹃位于第1主成分轴的负向区域,具有较大的叶磷质量分数、叶氮质量分数和较小的叶厚度、海绵组织厚度、栅栏组织厚度;白毛杜鹃、鲁朗杜鹃花、光蕊杜鹃、硬毛杜鹃和栎叶杜鹃位于第1主成分轴的正向区域,具有较大的叶厚度、海绵组织厚度、栅栏组织厚度和较小的叶磷质量分数、叶氮质量分数;柳条杜鹃和三花杜鹃位于第2主成分轴的负向区域,具有较大的比叶面积、叶含水量和较小的叶干物质质量分数、叶组织密度;黄杯杜鹃位于第2主成分轴的正向区域,具有较大的叶干物质质量分数、叶组织密度和较小的比叶面积、叶含水量。
性状 第1主成分 第2主成分 第3主成分 第4主成分 性状 第1主成分 第2主成分 第3主成分 第4主成分 叶面积 0.381 −0.104 −0.027 −0.008 叶磷质量分数 −0.224 0.325 −0.344 −0.068 叶厚度 0.308 0.111 −0.046 −0.020 叶碳氮比 0.340 −0.138 −0.104 0.167 叶干质量 0.383 −0.042 0.002 0.074 叶碳磷比 0.222 −0.270 0.270 0.416 叶组织密度 0.171 0.363 0.239 −0.030 叶氮磷比 −0.162 −0.110 0.477 0.350 叶干物质质量分数 0.125 0.443 0.256 0.025 栅栏组织厚度 0.220 0.132 −0.421 0.184 叶形指数 0.164 −0.111 0.357 −0.352 海绵组织厚度 0.289 0.035 −0.238 0.123 叶含水量 −0.123 −0.446 −0.256 −0.028 特征根 6.212 3.642 2.014 1.493 比叶面积 −0.191 −0.382 −0.037 0.015 方差贡献率/% 38.824 22.765 12.585 9.328 叶碳质量分数 −0.110 0.118 −0.112 0.665 累积贡献率/% 38.824 61.589 74.174 83.502 叶氮质量分数 −0.324 0.217 0.066 0.220 Table 3. Factor loading matrix of principal component analysis of leaf functional traits of Rhododendron
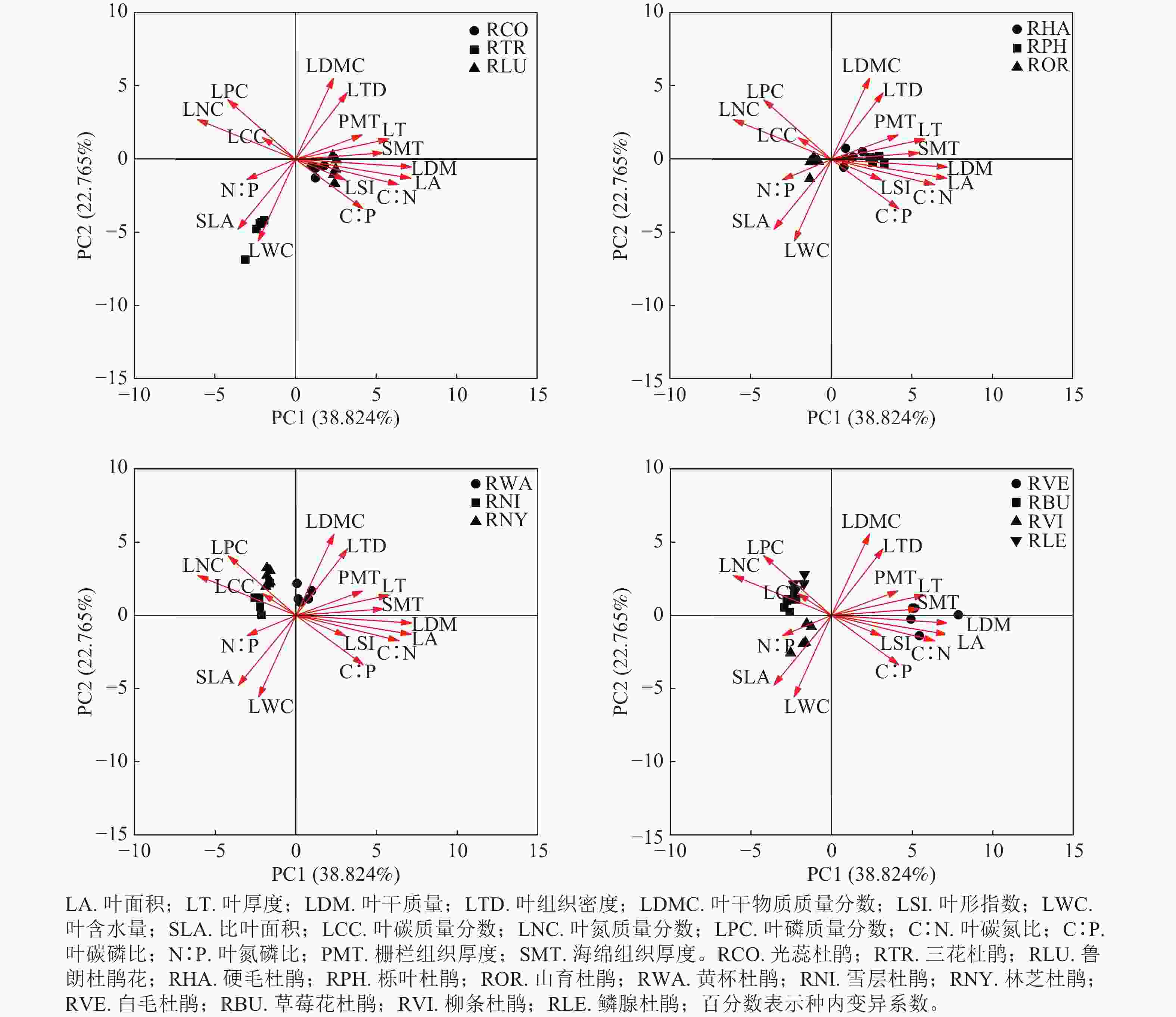
Figure 4. Principal component analysis of leaf functional traits of Rhododendron
-
植物叶功能性状变异是物种适应环境异质性,实现生态位分化的关键生长策略,变异幅度在一定程度上反映了植物生态适应性范围[42]。相关研究发现:不同叶性状的种间变异幅度差异非常大,即使是同一性状,在时空尺度上的变异程度差异也很大[43−44]。本研究结果显示:色季拉山13种杜鹃花属植物的16个叶功能性状的变异幅度为10.67%~120.29%。变异的程度代表了植物种质资源的固有特征及个体差异范围,变异系数越大,在资源选择时其选择空间也越大[45]。本研究中,杜鹃花属植物的叶干质量的变异系数最大,为120.29%,属于强变异,表明其选择空间较大;叶面积、比叶面积、叶碳氮比和叶磷质量分数的变异程度均较大,其中叶面积的变异系数达到96.28%。这些性状与植物对光、水和土壤养分的竞争密切相关[46]。植物叶片大小直接影响植物对光的截取和对碳的获取能力,并且通过影响叶片边界层厚度进而影响叶片温度和蒸腾速率,最终影响叶片热量和水平衡[47]。相关研究结果表明:植物叶干物质质量分数的变异幅度较低,是资源获取轴上较稳定的变量[48],这与本研究的结果相似。本研究中杜鹃花属植物叶碳质量分数的变异系数最小,与已有研究结果一致[49],属于稳定的叶功能性状指标。
本研究中,杜鹃花属植物叶功能性状的种间变异普遍高于种内变异,表明种间变异在植物性状变异中占据着主导地位,对群落构建的影响更显著[50]。然而,越来越多的证据表明:种内变异也是不可忽视的,可达性状总变异的28.00%~52.00%[51]。色季拉山杜鹃花属植物叶功能性状的种内变异平均达9.38%,相较于其他植被类型,其性状种内变异幅度较低,这可能反映了严酷生境(高寒、强紫外线、缺氧)条件下较小的形态可塑性[52]。碳、氮、磷元素是植物生长发育过程中所必需的元素,不仅影响植物蛋白质、酶和核酸等的合成,而且对植物光合作用、呼吸作用、能量传递等过程至关重要[53−54]。本研究结果显示:色季拉山杜鹃花属植物叶氮质量分数、叶磷质量分数、叶氮磷比均属于中等变异,分别为15.04 g·kg−1、0.88 g·kg−1、17.47。与全球陆生植物叶氮质量分数(18.74 g·kg−1)、叶磷质量分数(1.21 g·kg−1)和叶氮磷比(15.55)[55−56]相比,色季拉山杜鹃花属植物叶磷质量分数相对较低。相关研究结果表明:叶氮磷比≥16.00表示植被生长受磷限制,叶氮磷比<14.00表示受氮限制,14.00≤叶氮磷比<16.00时受氮磷的共同限制[43]。色季拉山杜鹃花属植物叶氮磷比为17.47,表明杜鹃花生长受磷限制。
-
植物在生长过程中受到环境、生理、系统发育等因素的共同作用,各性状间存在一定的相关性。这是在漫长进化历程中,植物通过不断协同与权衡,经自然筛选最终形成的,与周围环境相适应的性状组合[57−59]。本研究发现:色季拉山杜鹃花属植物比叶面积与叶厚度均呈显著负相关,这与前人的研究结果一致[60]。通常来说,较厚的叶片伴随较低的比叶面积,且叶片增厚有利于增加叶片内部水分向叶片表面扩散的阻力或距离,增强抵抗水分丧失的能力。这类叶片虽然光合作用效率较低,但能增强植物适应恶劣环境压力的能力[61]。资源分配假说认为:当植物将较多资源分配给叶片物理防御结构时,对叶片光合能力的投资则会降低[62]。本研究中,杜鹃花属植物叶功能性状反映植物光合作用的比叶面积与反映防御能力的叶干物质质量分数呈显著负相关,分析原因可能是色季拉山常年温度较低、紫外线较强,植物需要更多结构物质和更厚的叶片来保护光合器官。这也充分证明了资源分配假说,同时也表明植物在生长和防御能力之间存在一种“此消彼长”的权衡关系,达到平衡植物生存、生长、繁殖的目的[63]。
本研究中,叶含水量与叶干物质质量分数、叶组织密度呈极显著负相关,这说明杜鹃花属植物在低温强辐射的环境中,优先积累干物质来构建增强叶片的结构强度,且因“空间被挤占”和“代谢需求适配”,表现出更高的水分亏缺;同时更高的叶干物质质量分数常伴随叶组织密度的提高,这进一步降低了叶片内部活跃水分的含量[64]。本研究中,叶碳氮比与叶面积呈极显著正相关,这是因为杜鹃花扩大叶面积时,会优先分配资源合成坚固的结构框架来避免低温冻裂和强风损伤,导致叶碳质量分数随叶面积增加而上升,而叶氮质量分数的增长速度远慢于叶碳质量分数,最终导致叶碳氮比随叶面积的增大而升高[65],反映了植物在长期进化过程中,通过外部形态及内部结构特征的调节来适应环境的变化,最终形成一系列适应特定生境的最佳性状组合[59]。此外,色季拉山杜鹃花属植物的叶氮质量分数与叶磷质量分数呈显著正相关。这表明杜鹃花在相似生境中对氮、磷营养元素的需求基本一致,两者协同作用促进植物的生长[56]。这与WANG等[66]、闫风辰等[67]的研究结论一致。
-
根据植物叶经济谱理论,植物为适应特定环境变化所产生的资源权衡策略可以通过植物功能性状组合来反映[68]。WRIGHT等[10]提出了“叶经济谱”的概念,揭示了植物2种不同的生态策略,即“快速投资-收益型”策略和“缓慢投资-收益型”策略。在植物叶功能性状参数中,叶干物质质量分数、比叶面积、叶面积、叶氮质量分数、叶厚度、叶磷质量分数、叶组织密度与叶含水量等指标是众多植物在资源利用分类轴上的最佳变量,具有很好的可预测性[13]。本研究中色季拉山13种杜鹃花属植物处于叶经济谱不同位置,说明属内不同物种在相似环境中可能具有不同的生长策略[69]。山育杜鹃、草莓花杜鹃、鳞腺杜鹃、雪层杜鹃和林芝杜鹃位于第1主成分轴的负向区域,倾向于高的叶氮质量分数、叶磷质量分数和较小的叶厚度,倾向于“快速投资-收益型”策略;白毛杜鹃、鲁朗杜鹃花、光蕊杜鹃、硬毛杜鹃和栎叶杜鹃位于第1主成分轴的正向区域,倾向于较大的叶厚度和较小的叶氮质量分数、叶磷质量分数,倾向于“缓慢投资-收益型”策略;柳条杜鹃和三花杜鹃位于第2主成分轴的负向区域,倾向于高的比叶面积、叶含水量和较小的叶干物质质量分数、叶组织密度,在叶经济谱上倾向于“快速投资-收益型”策略;黄杯杜鹃位于第2主成分轴的正向区域,具有较大的叶干物质质量分数、叶组织密度和较小的比叶面积、叶含水量,在叶经济谱上倾向于“缓慢投资-收益型”策略[70]。在自然状态下,植物群落中资源环境在水平和垂直层面上都存在明显的异质性,这种异质性为多物种共存提供了环境基础[71],而植物功能性状间的资源权衡关系使不同物种表现出不同的性状组合,从而充分利用资源环境来降低共存物种间生态位重叠,促进物种间稳定共存[72]。本研究中杜鹃花属植物不同物种沿性状轴的变化不仅表明了属内物种间生态策略的差异,同时也反映了植物功能性状间资源权衡关系以及不同物种生态位的分化[73]。为了更准确地揭示功能性状权衡和环境变量之间的关系,未来的研究应尽可能广泛地收集地形、土壤、光照强度、海拔等信息,以获得最可靠的研究结果。
-
本研究以青藏高原色季拉山为研究区域,通过对13种杜鹃花属植物的叶功能性状差异及其权衡关系的分析,揭示了杜鹃花属植物的生长策略和生态适应机制。研究发现:杜鹃花属植物16个叶功能性状的变异幅度为10.67%~120.29%,叶干质量变异系数最大,为120.29%,说明杜鹃花属植物对环境适应能力强,且叶干质量是该属植物适应环境的核心维度。色季拉山杜鹃花属植物通过调整叶片功能性状,形成了2种不同的资源权衡策略,反映了植物功能性状间的资源权衡关系以及不同物种生态位的分化。研究结果可为探明该区域杜鹃花属植物资源权衡策略提供理论依据。
Leaf functional traits and resource balance analysis of Rhododendron in Shergyla Mountain
doi: 10.11833/j.issn.2095-0756.20260119
- Received Date: 2026-01-11
- Accepted Date: 2026-03-23
- Rev Recd Date: 2026-03-04
-
Key words:
- leaf functional trait /
- leaf economic spectrum /
- balancing strategy /
- Rhododendron /
- Shergyla Mountain
Abstract:
| Citation: | GUO Wenwen, BAI Youzhi, ZHUO Mecao, et al. Leaf functional traits and resource balance analysis of Rhododendron in Shergyla Mountain[J]. Journal of Zhejiang A&F University, 2026, 43(X): 1−14 doi: 10.11833/j.issn.2095-0756.20260119

|
 DownLoad:
DownLoad: