





-
樱花是蔷薇科Rosaceae李亚科Prunoideae樱属Cerasus典型樱亚属植物(Subg. Cerasus)的泛称。全世界约150种,分布于亚洲、欧洲、北美洲等北半球温和地带,主要种类分布在中国、日本和朝鲜[1]。中国有38种8变种[2],其中:浙江省有10种,占全国资源的1/4[3]。樱花Cerasus是浙江省早春开花植物之一,花期一般在2-5月,株型优美,花型丰富,花色艳丽,有白色、粉色、玫红色、深红色等;花朵数量多,独树成景,是优良的园林观花乔木,具有较高的应用价值。樱花利用方面,中国尚处于较为落后的状态。目前栽培的品种大多引自于日本,且品种相对单一,以染井吉野C. × yedoensis ‘Somei-yoshino’,关山C. serrulata ‘Kanzan’和普贤象C. serrulata ‘Albo-rosea’等几个品种为主,多数品种适应性较差。浙江省樱花野生种类丰富,具有品种选育的基因资源优势;与栽培品种相比,由于长期适应环境条件的变化以及不断抵御生物和非生物胁迫,野生种在进化过程中形成并积累了丰富的遗传变异和有益基因资源,在育种的目标性状选择等遗传改良方面,具有十分重要的作用。然而,由于樱花资源分布广泛,花期相对集中且较短,仅依靠花部形态的收集很难在短时间内完成野生樱花种类的鉴定及优株筛选。植物枝干及叶片形态的变异以其自身的遗传为基础,是种质资源鉴定和筛选的重要依据。目前,关于樱花的研究主要集中在繁殖技术[4-8]、地理起源分布[9-11]、群落特征[12-17]、亲缘关系[18-21]等方面,关于其枝干、叶片研究的相关报道仅见于叶部腺体[22]、表皮毛被[23]、花粉结构[24]等微形态学的研究,缺乏对枝干、叶片形态变异全面系统的论述。因此,本研究以浙江省野生樱花资源为试验材料,通过对其枝干、叶片形态性状进行调查和分析,以揭示浙江省野生樱花种间及种内枝干、叶片形态的变异大小和遗传规律,为野生樱花物种鉴定和品种培育提供有力证据,也为科学合理地保护和利用浙江省现有的野生樱花资源提供理论依据。
-
浙江省位于中国东部沿海,处于欧亚大陆与西北太平洋的过渡地带。该地带属亚热带季风气候区。四季分明,气温适中,年平均气温15.0~18.0 ℃;光照较多,年平均日照时数1 710~2 100 h。雨量丰沛,年平均降水量为980~2 000 mm[25]。
-
根据浙江省野生樱花的分布情况,在全省范围内设计调查点,于2015-2016年连续2 a深入分布地进行调查。最终确定以主要分布的6种,2变种为试验材料,包括迎春樱C. discoidea,浙闽樱C. schneideriana,大叶早樱C. subhirtella,山樱花C. serrulata,毛叶山樱花C. serrulata var. pubescens,华中樱C. conradinae,毛萼华中樱C. conradinae var. trichocalyx和尾叶樱C. dielsiana,进行形态指标的调查与测量。
-
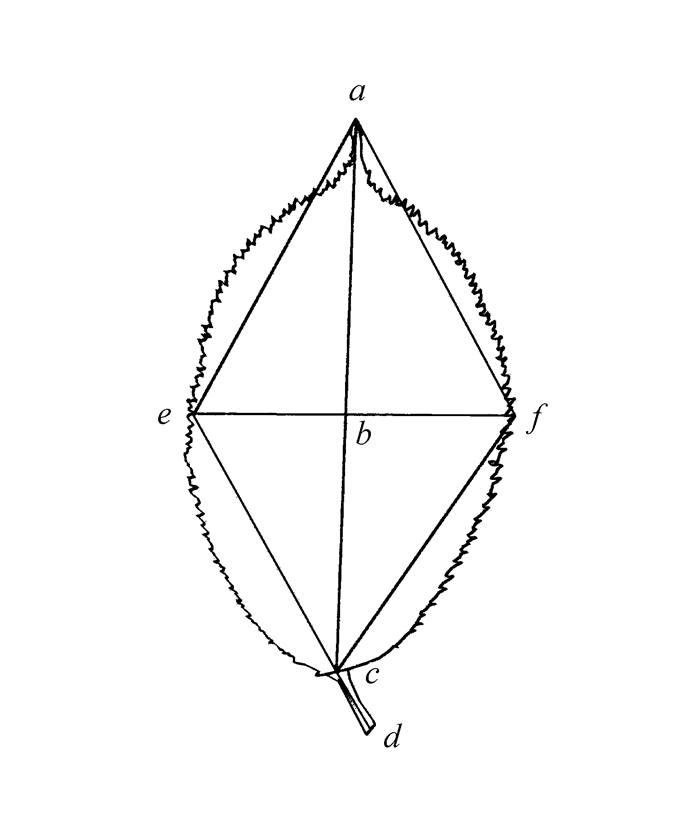
本研究选定了50个形态学性状进行观测,其中叶片及托叶性状34个(表 1,图 1),枝条的相关性状16个(表 2)。选取10~15株·种-1,取中部东西南北侧枝条各3个·株-1,并随机选择50片成熟叶片进行性状测量;颜色的测量依据英国园林园艺植物比色卡。

图 1 樱花叶片结构图
Figure 1. Leaf of Cerasus
表 1 叶片形态性状及编码
Table 1. Morphological traits of leaf and code
性状 形态学性状表述(单位)及编码 性状类型 叶长 叶基到叶尖的距离(mm),图 1中ac 数值性状 叶宽 叶片最宽处的长度(mm),图 1中ef 数值性状 叶形指数 叶长/叶宽,图 1中ac/ef 数值性状 叶片最宽处距叶尖距离 图 1中ab 数值性状 叶柄长 从枝干着生处到出现叶肉处的距离(mm),图 1中cd 数值性状 叶柄/叶片长 叶柄长/叶长,图 1中cd/ac 数值性状 叶缘齿数 叶片单侧裂齿数目(个) 数值性状 叶顶端夹角 (°),图 1中∠eaf 数值性状 叶基部夹角 (°),图 1中∠ecf 数值性状 叶面积 mm2 数值性状 托叶长 托叶基到托叶尖的距离(mm) 数值性状 托叶宽 托叶最宽处的直径(mm) 数值性状 幼叶颜色(正) 浅黄绿(1),浅灰黄、浅灰橙(2),浅灰紫、浅灰褐(3),黄绿(4),绿(5),灰橙(6),深绿(7) 多态性状 幼叶颜色(反) 浅灰橙、浅灰黄(1),浅灰红、浅灰褐(2),浅绿(3),黄绿(4),绿(5),灰橙(6),深黄绿(7),深红紫(8) 多态性状 幼叶毛被(正反)、成叶毛被(正反)、叶柄毛被 无(1),疏(2),中(3),密(4) 多态性状 成叶颜色(正) 黄绿(1),绿(2),深黄绿(3),深绿(4) 多态性状 成叶颜色(反) 浅绿、浅黄绿(1),黄绿(2),绿(3),深黄绿(4),深绿(5) 多态性状 叶片顶端形状 渐尖(1),短渐尖(2),尾尖(3),短尾尖(4),骤尖(5) 多态性状 叶片基部形状 近圆(1),楔形(2),微心形(3),圆钝、钝形(4),平截(5) 多态性状 叶形 椭圆(1),长椭圆(2),卵形、卵状椭圆(3),倒卵形(4) 多态性状 叶缘锯齿类型 单锯齿(1),重锯齿(2) 二元性状 叶缘锯齿形状 渐尖(1),急尖(2),圆钝(3) 多态性状 叶片齿端腺体 无(1),有(2) 二元性状 叶柄颜色 浅绿(1),浅灰褐(2),黄绿(3),绿(4),深绿(5),灰橙(6),深红紫、深褐色(7) 多态性状 叶柄腺体位置 叶基(1),上部(2),上部和叶基(3),上部和下部(4) 多态性状 叶柄腺体数量 Nmax=2(1),Nmax=3(2),Nmax=4(3),Nmax=5(4) 多态性状 叶柄腺体形状 盘状(1),头状(2) 二元性状 托叶形状 披针型羽裂(1),卵形羽裂(2),狭卵圆羽裂(3) 多态性状 托叶颜色 浅绿(1),黄绿(2),绿(3),深绿(4),深灰紫(5) 多态性状 托叶齿端腺体 无(1),有(2) 二元性状 表 2 枝条性状及编码
Table 2. Cerasus branch traits and code
性状 形态学性状表述(单位)及编码 性状类型 株型 灌木(< 3m)(l),小乔木(3~8m)(2),乔木(>8m)(3) 多态性状 树形 扁圆形(1),卵形(2),锥形(3),伞形(4),旗形(5) 多态性状 长势 弱(1),中(2),强(3) 多态性状 树皮颜色 浅褐色(1),灰褐色(2),褐色(3),深褐色(4) 多态性状 皮孔特征 横列(1),口唇状(2) 二元性状 小枝颜色 浅灰褐(1),灰橙(2),灰绿(3),褐色(4),深灰橙(5),深褐色(6) 多态性状 小枝毛被、嫩枝毛被 无(1),疏(2),中(3),密(4) 多态性状 嫩枝颜色 浅绿(1),浅橙红(2),灰褐(3),黄绿(4),绿(5),灰橙(6),灰紫红(7),深红紫(8) 多态性状 老枝颜色 浅褐色(1),灰褐(2),褐色(3),深灰橙(4),深褐色(5),黑褐色(6) 多态性状 株高 植株基部至主茎顶部的距离(m) 数值性状 枝下高 植物基部至第1个分枝点的距离(m) 数值性状 胸径 距植物基部1.3 m处的树木直径(cm) 数值性状 冠幅南北、冠幅东西 植物南北或者东西方向的宽度(m) 数值性状 分枝角度 第1个分枝与主茎的夹角(°) 数值性状 -
对数值性状进行描述性统计,计算其变异系数VC=标准差/均值×100%,进行单因素方差分析;二元和多态性状进行形态变异的频数统计,计算遗传多样性指数H′,H′=-Σpilnpi,pi为某一性状第i级内材料份数占总份数的百分比[26];对变异较大的性状进行主成分分析。本研究所有数据的统计分析均用SPSS 19.0软件。
-
对野生樱花叶片和枝干的18个数值型性状进行单因素方差分析可知,株高在8个种(变种)间差异不显著,枝下高在种间差异显著(P<0.05),其余16个性状在种间差异均为极显著(P<0.01)。叶片和枝干数值型性状在种间的变异系数为14.05%~64.15%,其中,叶长的种间变异系数最小,胸径的最大。种间变异系数达30%以上的性状有10个,包括叶片性状5个,分别是托叶长、叶面积、托叶宽、叶柄/叶片长、叶柄长;其中,叶柄长的种间变异系数最大,达53.07%。除分枝角度外,其余枝干性状的变异系数均大于30%,说明枝干性状在不同种间变异幅度大。
樱花各性状的种内变异大部分均小于种间变异,如叶长、叶宽、叶柄长、叶面积、托叶长等。不同樱花叶长、叶面积的种内变异系数为9.15%~13.99%和18.09%~30.10%,其中,毛叶山樱花的叶长、叶面积变异最丰富,叶长为71.51~150.48 mm,叶面积为1 424.70~8 279.84 mm2;而浙闽樱的叶长和叶面积的数值较为集中。叶宽和东西冠幅的种内变异系数分别为12.62%~16.76%和7.14%~46.15%,华中樱叶宽变异程度较大,毛萼华中樱叶宽的离散程度最小。叶柄长的种内变异系数为20.20%~41.30%,其中大叶早樱的变异系数最大,山樱花的变异系数最小,还不足其种间变异系数的1/2。叶柄/叶片长、叶顶端夹角、胸径的种内变异系数分别为20.80%~45.99%,10.42%~15.88%和4.52%~51.88%,这3个性状中均为毛萼华中樱的变异系数最小,大叶早樱的种内变异系数最大。托叶长的种内变异系数以毛萼华中樱最小,仅为15.65%,毛叶山樱花最大,为28%;南北冠幅的种内变异系数以尾叶樱最大,毛萼华中樱最小。另外,在叶形指数、叶缘齿数、叶基部夹角、托叶宽、株高等性状中发现少数种的种内变异系数大于种间变异系数的现象,主要存在于华中樱和大叶早樱中。
-
野生樱花数值型性状间的相关系数达显著水平(P<0.05)的有70个,包含极显著水平(P<0.01)的有50个。其中,叶柄长与叶柄/叶片长的相关系数最大,为0.97,其次是叶宽与叶面积的相关性较高,达0.95。托叶宽与株高不相关,叶宽与叶形指数、叶形指数与叶基部夹角均表现出较高的负相关,相关系数分别为-0.71和-0.70。由此可见,大部分数值性状间彼此相关,甚至高度相关,包含了大量的重复信息,所以可以进行主成分分析。
-
对野生樱花叶片和枝干的32个二元及多态性状进行频率分布和多样性统计,各性状在樱花中的平均多样性指数为0.03~1.28(表 3~4)。其中,小枝毛被和托叶齿端腺体的平均多样性指数最低,幼叶反面颜色最高。6种樱花在野外的长势均中等,树干皮孔均横列;成熟叶片正面为深绿色,叶柄上部均有盘状腺体;托叶绿色、羽裂,齿端均有腺体。毛萼华中樱小枝上有中度的毛被,其余种小枝上均无毛,且该变种多态性状在变种内稳定,故表 4中不再单独列出。华中樱叶片边缘单锯齿分布频率为60%,其余种叶片边缘均为重锯齿。大叶早樱叶片齿端无腺体,其他均有。在这些性状中,叶形、幼叶颜色、成叶反面颜色、嫩枝颜色、小枝颜色和老枝颜色7个性状在樱花中平均多样性指数均在0.95以上,说明这些性状在各种内多样性丰富。
表 3 华中樱二元及多态性状的频率分布
Table 3. Frequence distribution of traits Cerasus conradinae
性状 性状占比例/% 幼叶颜色(正) 浅黄绿(5),浅灰黄、浅灰橙(20),浅灰紫、浅灰褐(5),黄绿(25),灰橙(45) 幼叶颜色(反) 浅灰橙、浅灰黄(30),浅灰红、浅灰褐(15),浅绿(10),黄绿(15),灰橙(25),深红紫(5) 幼叶毛被(正) 无(75),疏(15),中(5),密(5) 幼叶毛被(反) 无(75),疏(15),中(5),密(5) 成叶颜色(正) 黄绿(5),绿(15),深黄绿(10),深绿(70) 成叶颜色(反) 浅绿、浅黄绿(40),黄绿(5),绿(30),深黄绿(20),深绿(5) 成叶毛被(正) 无(85),疏(10),中(5) 成叶毛被(反) 无(90),疏(5),中(5) 叶片顶端形状 渐尖(10),短渐尖(10),尾尖(60),短尾尖(15),骤尖(5) 叶片基部形状 近圆(20),楔形(50),微心形(20),圆钝、钝形(10) 叶形 椭圆(35),长椭圆(15),卵形、卵状椭圆(15),倒卵形(35) 叶缘锯齿类型 单锯齿(60),重锯齿(40) 叶缘锯齿形状 渐尖(5),圆钝(95) 叶片齿端腺体 有(100) 叶柄毛被 无(90),疏(10) 叶柄颜色 绿(5),灰橙(5),深红紫、深褐色(90) 叶柄腺体位置 上部(100) 叶柄腺体数量 Nmax=2(90),Nmax=3(10) 叶柄腺体形状 盘状(100) 托叶形状 披针型羽裂(60),卵形羽裂(30),狭卵圆羽裂(10) 托叶颜色 浅绿(15),黄绿(10),绿(70),深灰紫(5) 托叶齿端腺体 有(100) 株型 小乔木(3〜8m)(70),乔木(>8m)(30) 树形 锥形(45),伞形(50),旗形(5) 长势 中(80),强(20) 树皮颜色 灰褐色(45),褐色(10),深褐色(45) 皮孔特征 横列(85),口唇状(15) 小枝颜色 浅灰褐(25),灰橙(15),灰绿(20),褐色(10),深灰橙(25),深褐色(5) 小枝毛被 无(100) 嫩枝颜色 浅橙红(10),灰褐(5),灰橙(15),灰紫红(5),深红紫(65) 嫩枝毛被 无(100) 嫩枝毛被 浅褐色(20),灰褐(25),褐色(20),深褐色(20),黑褐色(15) 表 4 野生樱花二元及多态性状的多样性
Table 4. Diversity index of Cerasus traits
性状 多样性指数H' 均值 华中櫻 尾叶櫻 迎春櫻 山櫻花 浙闽櫻 大叶早櫻 毛叶山櫻花 幼叶颜色(正) 1.33 1.45 0.68 1.56 0.69 0 1.19 0.99 幼叶颜色(反) 1.66 1.43 1.07 1.56 0.69 1.04 1.50 1.28 幼叶毛被(正) 0.80 0.73 0.58 0.64 0.69 1.04 0.64 0.73 幼叶毛被(反) 0.80 0.33 0.22 0.64 0 0 0.95 0.42 成叶颜色(正) 0.91 0.39 0.58 0.45 0 0 0 0.33 成叶颜色(反) 1.35 1.19 1.24 1.33 0 1.04 0.67 0.97 成叶毛被(正) 0.52 0.39 0.87 0 0.69 0.69 0.67 0.55 成叶毛被(反) 0.39 0.86 0.81 0.45 0.69 0.56 0.94 0.67 叶片顶端形状 1.20 0 0.47 0.45 0 0.69 0.80 0.52 叶片基部形状 1.22 1.26 0.65 1.01 0.69 0 1.03 0.84 叶形 1.30 1.29 1.03 1.01 0.69 0.56 0.80 0.95 叶缘锯齿类型 0.67 0 0.22 0 0 0 0 0.13 叶缘锯齿形状 0.20 0.79 0.87 0.64 0.69 0.56 0.67 0.63 叶片齿端腺体 0 0 0 0.64 0 0.56 0.61 0.26 叶柄毛被 0.33 0.33 0.87 0.64 0.69 0.69 1.17 0.67 叶柄颜色 0.39 1.45 1.53 0.69 0.69 0.56 0.80 0.87 叶柄腺体位置 0 0.39 0.22 0 0 0.69 0 0.19 叶柄腺体数量 0.33 0.93 0.97 0.87 0 0 1.28 0.63 叶柄腺体形状 0 0.50 0 0 0 0 0 0.07 托叶形状 0.90 0.34 0.58 0 0 0 0.56 0.34 托叶颜色 0.91 0.83 0.44 0 0.69 0 0 0.41 托叶齿端腺体 0 0 0.22 0 0 0 0 0.03 株型 0.61 0.58 0.55 0.64 0.69 0.56 0.53 0.59 树形 0.86 0.21 1.24 0.45 0.69 0.69 0.94 0.73 长势 0.50 0 0.47 0 0.69 0.69 0 0.34 树皮颜色 0.95 1.16 0.83 1.01 0 0.69 1.03 0.81 皮孔特征 0.42 0 0 0 0 0 0 0.06 小枝颜色 1.68 1.45 1.53 0.45 0.69 0.69 1.42 1.13 小枝毛被 0 0.20 0 0 0 0 0 0.03 嫩枝颜色 1.09 1.68 1.32 0.95 0.69 0.56 1.31 1.09 嫩枝毛被 0 0.52 0.68 0 0 1.04 0.53 0.40 嫩枝毛被 1.60 1.37 1.24 1.56 0.69 0 1.09 1.08 -
将樱花18个数值型性状和7个遗传多样性指数较高的多态性状转化为25个主成分进行分析。结果显示:第1主成分的特征值为6.18,方差贡献率为24.71%,代表了樱花枝干、叶片性状信息的24.71%,是最重要的主成分;其中,叶宽、叶长、叶面积、叶柄长、托叶长宽、叶柄/叶片长、叶片最宽处距叶尖距离和叶基部夹角在第1主成分上具有较高载荷,均在0.50以上,说明第1主成分基本反映了这些指标的信息。冠幅、胸径、分枝角度、株高和枝下高在第2主成分具有较高载荷;叶形指数在第3主成分上载荷最高,为0.67,其次是老枝颜色;叶柄/叶片长、幼叶正反面颜色在第4主成分上载荷较高;前4个主成分的累积贡献率为54.87%。第5主成分与嫩枝颜色、幼叶颜色(正)、叶形相关性较高,第6主成分与叶形、成叶颜色(反)相关性较高,第7主成分与成叶颜色(反)、叶缘齿数的相关系数均在0.4以上,第8主成分与枝下高的相关性最高,相关系数为0.49。前8个主成分的累积方差贡献率为76.85%,已经将樱花枝干和叶片性状70%以上的信息反映出来;因此,可以提取前8个主成分作为野生樱花枝干和叶片形态性状评价的综合指标(表 5)。
表 5 樱花枝干及叶片性状的主成分的贡献率和因子载荷矩阵
Table 5. Contributive percentage and factor loading matrix from principal components analysis
性状 第1主成分 第2主成分 第3主成分 第4主成分 第5主成分 第6主成分 第7主成分 第8主成分 叶长 0.83 0.03 0.26 -0.38 0.04 -0.01 0.01 -0.03 叶宽 0.92 -0.07 -0.25 -0.17 -0.05 0.12 -0.12 -0.01 叶形指数 -0.54 0.18 0.67 -0.13 0.15 -0.23 0.24 0.00 叶片最宽处距叶尖距离 0.67 0.11 0.02 -0.62 -0.09 -0.09 -0.05 -0.01 叶柄长 0.80 -0.21 -0.01 0.37 -0.01 -0.19 0.00 -0.01 叶柄/叶片长 0.67 -0.24 -0.06 0.50 -0.03 -0.23 -0.01 0.02 叶缘齿数 0.08 0.09 -0.34 -0.47 -0.12 0.12 0.41 0.14 叶顶端夹角 0.05 0.13 -0.81 0.00 -0.34 0.00 0.18 -0.15 叶基部夹角 0.61 -0.31 -0.13 0.21 0.13 0.30 -0.43 0.16 叶面积 0.92 -0.02 -0.08 -0.26 -0.04 0.06 0.00 -0.06 托叶长 0.77 -0.10 0.23 0.05 0.15 -0.29 0.29 -0.09 托叶宽 0.53 -0.50 0.29 -0.11 0.02 -0.26 0.30 -0.14 幼叶颜色(正) 0.22 0.08 -0.25 0.40 0.42 -0.25 0.37 0.36 幼叶颜色(反) 0.04 0.24 -0.22 0.40 0.35 0.15 0.17 -0.24 成叶颜色(反) 0.01 -0.02 0.29 0.23 -0.40 0.48 0.45 -0.01 叶形 0.11 0.06 -0.13 -0.07 0.41 0.56 0.28 0.25 小枝颜色 0.28 -0.35 0.10 0.21 -0.15 0.30 0.34 -0.19 嫩枝颜色 -0.09 0.22 -0.35 -0.26 0.47 -0.26 0.07 0.31 老枝颜色 0.24 -0.19 0.40 -0.21 0.27 0.39 -0.12 0.32 株高 0.35 0.54 0.25 0.27 -0.22 -0.04 0.06 0.39 枝下高 0.19 0.40 0.07 0.15 -0.59 -0.08 -0.03 0.49 胸径 0.37 0.74 0.10 0.18 -0.12 0.01 -0.16 -0.05 冠幅南北 0.31 0.78 0.01 0.08 0.19 0.12 -0.02 -0.28 冠幅东西 0.32 0.73 0.22 0.14 0.28 0.13 -0.10 -0.25 分枝角度 0.08 0.61 -0.05 -0.30 -0.12 -0.10 0.20 -0.10 特征值 6.18 3.32 2.15 2.08 1.67 1.41 1.30 1.12 贡献率/% 24.71 13.27 8.59 8.30 6.68 5.65 5.18 4.46 累积贡献率/% 24.71 37.98 46.57 54.87 61.55 67.20 72.38 76.85 第1主成分反映了叶片的主要性状,第2主成分反映了枝干的主要性状,前2个主成分在8个主成分中所占的比例最大;以第1主成分值为横坐标,第2主成分值为纵坐标,对樱花种质资源进行聚类。由图 2可知:91份野生樱花单株可分为5大类,第Ⅰ类为大叶早樱类;第Ⅱ类为华中樱类,其变种毛萼华中樱聚在一起;第Ⅲ类为迎春樱类,第Ⅳ类主要为尾叶樱类;第Ⅴ类为山樱花类,其变种毛叶山樱花聚在一起。其中,浙闽樱并没有成为独立的一类,而是位于尾叶樱和毛叶山樱花的中间。此结果说明:通过前2个主成分可将5类樱花大体区分出来,但仅依靠枝干和叶片的性状差异,无法将浙闽樱从其他野生樱花中区分出来;若要进一步分类,还需加入更多其他性状。

图 2 基于形态性状的樱花种质资源主成分分析
Figure 2. Principal component analysis of Cerasus based on morphologic traits
-
形态性状是指那些肉眼可以观察或仪器可以直接测量的植物表观特征,如枝干不同时期的颜色、有无被毛,叶片的长宽、面积、锯齿数、齿端腺体,托叶的形状、长宽、有无腺体等;具有简单、便于识别、快速等特点。形态性状研究是探讨植物遗传变异和多样性最直接、最基础的方法[27-28],是在短期内对种质资源变异进行评价的重要手段;同时也是大多数植物种质鉴定的基础[29]。作为中国主要分布的观花乔木,樱花分布广泛,花期相对集中且较短,仅依靠花部形态的收集很难在短时间内完成野生樱花种类的鉴定及优良单株的筛选;而叶片和枝干作为樱花重要的同化器官,其形态特征直接影响到植物的基本行为和功能[30],其性状具有一定的观赏特性,且性状稳定、持续时间长,便于观测[31];因此,樱花枝干和叶片性状的研究在樱花初期研究中具有重要的意义。本研究选取了樱花枝干和叶片的50个形态学性状,数值型性状18个,除株高外,其余17个性状在种间差异显著,其中16个极为显著。这些性状在不同种间的变异系数为14.05%~64.15%。变异系数表示性状离散程度,可在消除量纲的情况下间接反映不同个体或群体在不同性状上的变异程度,变异系数越大,则性状值离散程度越大[32]。叶片性状中托叶长、叶面积、托叶宽、叶柄/叶片长、叶柄长的种间变异系数均大于30%;枝干性状中除分枝角度外,其余性状的变异系数均大于30 %;说明这些性状在不同种间变异幅度较大,性状多样性丰富。另外,各种内个体之间也存在着各种形态变异。研究发现:樱花各性状的种内变异大部分均小于种间变异,这与红榉Zelkova schneideriana[33],印楝属Azadirachta[34],杜鹃红山茶Camellia azalea[27],云南樱花Cerasus yunnannensis[35]等植物的研究结果一致。同时,对野生樱花叶片和枝干的32个二元及多态性状进行频率分布和多样性统计表明,叶形、幼叶颜色、成叶反面颜色、嫩枝颜色、小枝颜色和老枝颜色7个性状在樱花中平均多样性指数均在0.95以上,说明这些性状在各种内多样性丰富,有较大的改良潜力,可为优良种质资源的筛选提供依据。
植物在适应和演化过程中,不同形态性状之间往往通过相互调节达到适应环境的目的。一个形态性状的改变亦直接或者间接导致其他性状的变化,因此,形态性状间相关性是植物资源研究和利用的重要内容[28, 36]。本研究发现:樱花各数值性状间彼此相关,达到显著水平的有70个,包含了大量的重复信息,可利用主成分分析对众多的性状进行降维,从而使分析结果更加准确和客观。通过主成分分析方法将18个数值型性状和7个多样性指数较高的多态性状转化为25个主成分,最终综合为8个主成分,综合了枝干和叶片的主要特性,其代表性达76.85%。根据前2个主成分值,对91份樱花种质资源进行聚类,可分为大叶早樱类、华中樱类、迎春樱类、尾叶樱、山樱花类,说明主成分分析的方法在这5种樱花枝干、叶片的性状分析中是可行的,可将大量的性状综合为8个主成分,以便在今后的选育过程中利用少数几个因子进行优良单株筛选,从而提高选择效率。
樱花种质资源枝干、叶片形态特征和多样性的研究说明浙江省主要分布的8种(变种)樱花枝干、叶片性状差异显著,变异幅度大,多样性丰富,说明这些性状在遗传改良中具有较大的潜力,可为今后樱花物种鉴定、优良亲本选择、遗传改良及新品种选育提供有效的信息。
Morphological variation analysis of leaf and branch traits of wild Cerasus spp. in Zhejiang
-
摘要: 为科学合理地保护和利用浙江省野生樱花资源,挖掘其优良性状,以野生樱花种质资源为研究材料,对枝干、叶片的50个形态学性状进行测定,采用方差分析、主成分分析等方法研究其种间及种内各性状的变异,探明不同性状间的相关性。结果表明:①除株高外,其余17个数值型性状在8个种(变种)间差异显著(P < 0.05),其中16个性状为极显著(P < 0.01)。各性状在种间的变异系数为14.05%~64.15%,其中,叶长的种间变异系数最小,胸径的最大。叶片性状中托叶长、叶面积、托叶宽、叶柄/叶片长、叶柄长的种间变异系数均大于30%;除分枝角度外,其他枝干性状的变异系数均大于30 %;大部分性状的种内变异小于种间变异。②二元及多态性状的平均多样性指数为0.03~1.28,其中,叶形、幼叶颜色、成叶反面颜色、嫩枝、小枝和老枝颜色7个性状的平均多样性指数均在0.95以上。③叶片、枝干各性状间彼此相关,利用主成分分析将数值型性状和7个多样性指数较高的多态性状综合为8个主成分,其累计贡献率可达76.85 %。④根据主成分值将91份樱花种质资源分为5大类,分别为大叶早樱Cerasus subhirtella类,华中樱C. conradinae类,迎春樱C. discoidea类,尾叶樱C. dielsiana和山樱花C. serrulata类,其中2个变种也分别聚在了相应的种内,而浙闽樱C. schneideriana并没有成为独立的一类。研究认为:野生樱花种间及种内各性状的变异幅度大,差异显著,多样性丰富,具有较大的遗传改良潜力;主成分分析的方法在枝干、叶片性状分析中是可行的,可提高筛选效率。Abstract: To effectively protect and utilize the wild germplasm resources of Cerasus, 50 morphological traits of leaves and branches from eight Cerasus species (six species and two varieties) were investigated. Based on morphological data which was randomly measured fifty leaves and twelve branches from ten plants of each species, the interspecific and intraspecific variation of morphological traits and the relationship between them were analyzed by ANOVA and principal component analysis (PCA). Results showed that the differences for 17 quantitative characters among the eight species were highly significant (P < 0.01). The interspecific coefficient of variation (CV) of quantitative characters ranged from 14.05% to 64.15% with the largest value being diameter at breast height (DHB) and the smallest being leaf length. The CVs for stipule length, leaf area, stipule width, petiole length/leaf length, and petiole length were > 30% as were branch traits except for branch angle. Most of the intraspecific CVs were less than interspecific variations. The average diversity indexes of qualitative characters ranged from 0.03 to 1.28 with the average diversity indexes of leaf index, young leaf color, mature leaf color (abaxial), branchlet color, twig color, and branch color being > 0.95. Based on the PCA, the quantitative characters and seven qualitative characters with high diversity indexes were divided into eight principle components, with the cumulative contribution rate reaching 76.85%. Also, 91 wild germplasm resources were divided into 5 groups, namely Cerasus subhirtella, C. conradinae, C. discoidea, C. dielsiana, and C. serrulata. The two varieties were clustered into relevant species, and Cerasus schneideriana was not considered as an independent group. This research indicated strong differences in the traits of leaves and branches of Cerasus species, and the PCA was suitable for analyzing leaf and branch traits, which can screen and exploit superior germplasm resources efficiently.
-
Key words:
- botany /
- Cerasus /
- morphological trait /
- principal component analysis
-
图 2 基于形态性状的樱花种质资源主成分分析
Figure 2 Principal component analysis of Cerasus based on morphologic traits
表 1 叶片形态性状及编码
Table 1. Morphological traits of leaf and code
性状 形态学性状表述(单位)及编码 性状类型 叶长 叶基到叶尖的距离(mm),图 1中ac 数值性状 叶宽 叶片最宽处的长度(mm),图 1中ef 数值性状 叶形指数 叶长/叶宽,图 1中ac/ef 数值性状 叶片最宽处距叶尖距离 图 1中ab 数值性状 叶柄长 从枝干着生处到出现叶肉处的距离(mm),图 1中cd 数值性状 叶柄/叶片长 叶柄长/叶长,图 1中cd/ac 数值性状 叶缘齿数 叶片单侧裂齿数目(个) 数值性状 叶顶端夹角 (°),图 1中∠eaf 数值性状 叶基部夹角 (°),图 1中∠ecf 数值性状 叶面积 mm2 数值性状 托叶长 托叶基到托叶尖的距离(mm) 数值性状 托叶宽 托叶最宽处的直径(mm) 数值性状 幼叶颜色(正) 浅黄绿(1),浅灰黄、浅灰橙(2),浅灰紫、浅灰褐(3),黄绿(4),绿(5),灰橙(6),深绿(7) 多态性状 幼叶颜色(反) 浅灰橙、浅灰黄(1),浅灰红、浅灰褐(2),浅绿(3),黄绿(4),绿(5),灰橙(6),深黄绿(7),深红紫(8) 多态性状 幼叶毛被(正反)、成叶毛被(正反)、叶柄毛被 无(1),疏(2),中(3),密(4) 多态性状 成叶颜色(正) 黄绿(1),绿(2),深黄绿(3),深绿(4) 多态性状 成叶颜色(反) 浅绿、浅黄绿(1),黄绿(2),绿(3),深黄绿(4),深绿(5) 多态性状 叶片顶端形状 渐尖(1),短渐尖(2),尾尖(3),短尾尖(4),骤尖(5) 多态性状 叶片基部形状 近圆(1),楔形(2),微心形(3),圆钝、钝形(4),平截(5) 多态性状 叶形 椭圆(1),长椭圆(2),卵形、卵状椭圆(3),倒卵形(4) 多态性状 叶缘锯齿类型 单锯齿(1),重锯齿(2) 二元性状 叶缘锯齿形状 渐尖(1),急尖(2),圆钝(3) 多态性状 叶片齿端腺体 无(1),有(2) 二元性状 叶柄颜色 浅绿(1),浅灰褐(2),黄绿(3),绿(4),深绿(5),灰橙(6),深红紫、深褐色(7) 多态性状 叶柄腺体位置 叶基(1),上部(2),上部和叶基(3),上部和下部(4) 多态性状 叶柄腺体数量 Nmax=2(1),Nmax=3(2),Nmax=4(3),Nmax=5(4) 多态性状 叶柄腺体形状 盘状(1),头状(2) 二元性状 托叶形状 披针型羽裂(1),卵形羽裂(2),狭卵圆羽裂(3) 多态性状 托叶颜色 浅绿(1),黄绿(2),绿(3),深绿(4),深灰紫(5) 多态性状 托叶齿端腺体 无(1),有(2) 二元性状  下载: 导出CSV
下载: 导出CSV
表 2 枝条性状及编码
Table 2. Cerasus branch traits and code
性状 形态学性状表述(单位)及编码 性状类型 株型 灌木(< 3m)(l),小乔木(3~8m)(2),乔木(>8m)(3) 多态性状 树形 扁圆形(1),卵形(2),锥形(3),伞形(4),旗形(5) 多态性状 长势 弱(1),中(2),强(3) 多态性状 树皮颜色 浅褐色(1),灰褐色(2),褐色(3),深褐色(4) 多态性状 皮孔特征 横列(1),口唇状(2) 二元性状 小枝颜色 浅灰褐(1),灰橙(2),灰绿(3),褐色(4),深灰橙(5),深褐色(6) 多态性状 小枝毛被、嫩枝毛被 无(1),疏(2),中(3),密(4) 多态性状 嫩枝颜色 浅绿(1),浅橙红(2),灰褐(3),黄绿(4),绿(5),灰橙(6),灰紫红(7),深红紫(8) 多态性状 老枝颜色 浅褐色(1),灰褐(2),褐色(3),深灰橙(4),深褐色(5),黑褐色(6) 多态性状 株高 植株基部至主茎顶部的距离(m) 数值性状 枝下高 植物基部至第1个分枝点的距离(m) 数值性状 胸径 距植物基部1.3 m处的树木直径(cm) 数值性状 冠幅南北、冠幅东西 植物南北或者东西方向的宽度(m) 数值性状 分枝角度 第1个分枝与主茎的夹角(°) 数值性状
下载: 导出CSV
表 3 华中樱二元及多态性状的频率分布
Table 3. Frequence distribution of traits Cerasus conradinae
性状 性状占比例/% 幼叶颜色(正) 浅黄绿(5),浅灰黄、浅灰橙(20),浅灰紫、浅灰褐(5),黄绿(25),灰橙(45) 幼叶颜色(反) 浅灰橙、浅灰黄(30),浅灰红、浅灰褐(15),浅绿(10),黄绿(15),灰橙(25),深红紫(5) 幼叶毛被(正) 无(75),疏(15),中(5),密(5) 幼叶毛被(反) 无(75),疏(15),中(5),密(5) 成叶颜色(正) 黄绿(5),绿(15),深黄绿(10),深绿(70) 成叶颜色(反) 浅绿、浅黄绿(40),黄绿(5),绿(30),深黄绿(20),深绿(5) 成叶毛被(正) 无(85),疏(10),中(5) 成叶毛被(反) 无(90),疏(5),中(5) 叶片顶端形状 渐尖(10),短渐尖(10),尾尖(60),短尾尖(15),骤尖(5) 叶片基部形状 近圆(20),楔形(50),微心形(20),圆钝、钝形(10) 叶形 椭圆(35),长椭圆(15),卵形、卵状椭圆(15),倒卵形(35) 叶缘锯齿类型 单锯齿(60),重锯齿(40) 叶缘锯齿形状 渐尖(5),圆钝(95) 叶片齿端腺体 有(100) 叶柄毛被 无(90),疏(10) 叶柄颜色 绿(5),灰橙(5),深红紫、深褐色(90) 叶柄腺体位置 上部(100) 叶柄腺体数量 Nmax=2(90),Nmax=3(10) 叶柄腺体形状 盘状(100) 托叶形状 披针型羽裂(60),卵形羽裂(30),狭卵圆羽裂(10) 托叶颜色 浅绿(15),黄绿(10),绿(70),深灰紫(5) 托叶齿端腺体 有(100) 株型 小乔木(3〜8m)(70),乔木(>8m)(30) 树形 锥形(45),伞形(50),旗形(5) 长势 中(80),强(20) 树皮颜色 灰褐色(45),褐色(10),深褐色(45) 皮孔特征 横列(85),口唇状(15) 小枝颜色 浅灰褐(25),灰橙(15),灰绿(20),褐色(10),深灰橙(25),深褐色(5) 小枝毛被 无(100) 嫩枝颜色 浅橙红(10),灰褐(5),灰橙(15),灰紫红(5),深红紫(65) 嫩枝毛被 无(100) 嫩枝毛被 浅褐色(20),灰褐(25),褐色(20),深褐色(20),黑褐色(15)
下载: 导出CSV
表 4 野生樱花二元及多态性状的多样性
Table 4. Diversity index of Cerasus traits
性状 多样性指数H' 均值 华中櫻 尾叶櫻 迎春櫻 山櫻花 浙闽櫻 大叶早櫻 毛叶山櫻花 幼叶颜色(正) 1.33 1.45 0.68 1.56 0.69 0 1.19 0.99 幼叶颜色(反) 1.66 1.43 1.07 1.56 0.69 1.04 1.50 1.28 幼叶毛被(正) 0.80 0.73 0.58 0.64 0.69 1.04 0.64 0.73 幼叶毛被(反) 0.80 0.33 0.22 0.64 0 0 0.95 0.42 成叶颜色(正) 0.91 0.39 0.58 0.45 0 0 0 0.33 成叶颜色(反) 1.35 1.19 1.24 1.33 0 1.04 0.67 0.97 成叶毛被(正) 0.52 0.39 0.87 0 0.69 0.69 0.67 0.55 成叶毛被(反) 0.39 0.86 0.81 0.45 0.69 0.56 0.94 0.67 叶片顶端形状 1.20 0 0.47 0.45 0 0.69 0.80 0.52 叶片基部形状 1.22 1.26 0.65 1.01 0.69 0 1.03 0.84 叶形 1.30 1.29 1.03 1.01 0.69 0.56 0.80 0.95 叶缘锯齿类型 0.67 0 0.22 0 0 0 0 0.13 叶缘锯齿形状 0.20 0.79 0.87 0.64 0.69 0.56 0.67 0.63 叶片齿端腺体 0 0 0 0.64 0 0.56 0.61 0.26 叶柄毛被 0.33 0.33 0.87 0.64 0.69 0.69 1.17 0.67 叶柄颜色 0.39 1.45 1.53 0.69 0.69 0.56 0.80 0.87 叶柄腺体位置 0 0.39 0.22 0 0 0.69 0 0.19 叶柄腺体数量 0.33 0.93 0.97 0.87 0 0 1.28 0.63 叶柄腺体形状 0 0.50 0 0 0 0 0 0.07 托叶形状 0.90 0.34 0.58 0 0 0 0.56 0.34 托叶颜色 0.91 0.83 0.44 0 0.69 0 0 0.41 托叶齿端腺体 0 0 0.22 0 0 0 0 0.03 株型 0.61 0.58 0.55 0.64 0.69 0.56 0.53 0.59 树形 0.86 0.21 1.24 0.45 0.69 0.69 0.94 0.73 长势 0.50 0 0.47 0 0.69 0.69 0 0.34 树皮颜色 0.95 1.16 0.83 1.01 0 0.69 1.03 0.81 皮孔特征 0.42 0 0 0 0 0 0 0.06 小枝颜色 1.68 1.45 1.53 0.45 0.69 0.69 1.42 1.13 小枝毛被 0 0.20 0 0 0 0 0 0.03 嫩枝颜色 1.09 1.68 1.32 0.95 0.69 0.56 1.31 1.09 嫩枝毛被 0 0.52 0.68 0 0 1.04 0.53 0.40 嫩枝毛被 1.60 1.37 1.24 1.56 0.69 0 1.09 1.08
下载: 导出CSV
表 5 樱花枝干及叶片性状的主成分的贡献率和因子载荷矩阵
Table 5. Contributive percentage and factor loading matrix from principal components analysis
性状 第1主成分 第2主成分 第3主成分 第4主成分 第5主成分 第6主成分 第7主成分 第8主成分 叶长 0.83 0.03 0.26 -0.38 0.04 -0.01 0.01 -0.03 叶宽 0.92 -0.07 -0.25 -0.17 -0.05 0.12 -0.12 -0.01 叶形指数 -0.54 0.18 0.67 -0.13 0.15 -0.23 0.24 0.00 叶片最宽处距叶尖距离 0.67 0.11 0.02 -0.62 -0.09 -0.09 -0.05 -0.01 叶柄长 0.80 -0.21 -0.01 0.37 -0.01 -0.19 0.00 -0.01 叶柄/叶片长 0.67 -0.24 -0.06 0.50 -0.03 -0.23 -0.01 0.02 叶缘齿数 0.08 0.09 -0.34 -0.47 -0.12 0.12 0.41 0.14 叶顶端夹角 0.05 0.13 -0.81 0.00 -0.34 0.00 0.18 -0.15 叶基部夹角 0.61 -0.31 -0.13 0.21 0.13 0.30 -0.43 0.16 叶面积 0.92 -0.02 -0.08 -0.26 -0.04 0.06 0.00 -0.06 托叶长 0.77 -0.10 0.23 0.05 0.15 -0.29 0.29 -0.09 托叶宽 0.53 -0.50 0.29 -0.11 0.02 -0.26 0.30 -0.14 幼叶颜色(正) 0.22 0.08 -0.25 0.40 0.42 -0.25 0.37 0.36 幼叶颜色(反) 0.04 0.24 -0.22 0.40 0.35 0.15 0.17 -0.24 成叶颜色(反) 0.01 -0.02 0.29 0.23 -0.40 0.48 0.45 -0.01 叶形 0.11 0.06 -0.13 -0.07 0.41 0.56 0.28 0.25 小枝颜色 0.28 -0.35 0.10 0.21 -0.15 0.30 0.34 -0.19 嫩枝颜色 -0.09 0.22 -0.35 -0.26 0.47 -0.26 0.07 0.31 老枝颜色 0.24 -0.19 0.40 -0.21 0.27 0.39 -0.12 0.32 株高 0.35 0.54 0.25 0.27 -0.22 -0.04 0.06 0.39 枝下高 0.19 0.40 0.07 0.15 -0.59 -0.08 -0.03 0.49 胸径 0.37 0.74 0.10 0.18 -0.12 0.01 -0.16 -0.05 冠幅南北 0.31 0.78 0.01 0.08 0.19 0.12 -0.02 -0.28 冠幅东西 0.32 0.73 0.22 0.14 0.28 0.13 -0.10 -0.25 分枝角度 0.08 0.61 -0.05 -0.30 -0.12 -0.10 0.20 -0.10 特征值 6.18 3.32 2.15 2.08 1.67 1.41 1.30 1.12 贡献率/% 24.71 13.27 8.59 8.30 6.68 5.65 5.18 4.46 累积贡献率/% 24.71 37.98 46.57 54.87 61.55 67.20 72.38 76.85
下载: 导出CSV
-
[1] WU Zhengyi, RAVEN P H, HONG Deyuan. Flora of China:Volume 9[M]. Beijing:Science Press, 2003:404-420. [2] 俞德浚.中国植物志:第38卷[M].北京:科学出版社, 1986:41-87. [3] 韦直, 何业祺.浙江省植物志[M].杭州:浙江科学技术出版社, 1993:239-252. [4] HAMADA Y, OGAWA T, TANAKA H. Effects of low temperature and storage periods for seed dormancy release on Prunus lannesiana (Carr.) var. speciosa[J]. Acta Hortic, 1988, 226(2):541-548. [5] 王宇萍, 王朝文, 杨洪涛, 等.冬樱花种子育苗技术研究[J].种子, 2015, 34(3):117-119. WANG Yuping, WANG Chaowen, YANG Hongtao, et al. Study on the seeds seedling-raising technology of winter cherry[J]. Seed, 2015, 34(3):117-119. [6] 聂超仁, 许小过, 段庆明, 等. 2种方法提高山樱花种子发芽率试验[J].安徽农业科学, 2014, 42(13):3835-3838. NIE Chaoren, XU Xiaoguo, DUAN Qingming, et al. Two methods of improving germination rate of Cerasus serrualta(Lindl.) G. Don ex London[J]. J Anhui Agric Sci, 2014, 42(13):3835-3838. [7] SARROPOULOU V, DIMASSI-THERIOU K, THERIOS I. In vitro plant regeneration from leaf explants of the cherry rootstocks CAB-6P, Gisela 6, and MxM 14 using sodium nitroprusside[J]. In Vitro Cell Develop Biol-Plant, 2014, 50(2):226-234. [8] WANG Ying, PIJUT P M. Improvement of agrobacterium-mediated transformation and rooting of black cherry[J]. In Vitro Cell Develop Biol-Plant, 2014, 50(3):307-316. [9] 李苗苗.樱亚属植物分子亲缘地理及中国樱桃自然居群遗传多样性研究[D].西安: 西北大学, 2009. LI Miaomiao. Phylogeography of Prunus Subgenus Cerasus and Genetic Diversity of P. pseudocerasus[D]. Xi'an: Northwest University, 2009. [10] 王贤荣.中国樱花品种图志[M].北京:科学出版社, 2014:80-82. [11] 李蒙, 伊贤贵, 王华辰, 等.山樱花地理分布与水热环境因子的关系[J].南京林业大学学报(自然科学版), 2014, 38(增刊1):74-80. LI Meng, YIN Xiangui, WANG Huachen, et al. Studies on the relationship between Cerasus serrulata distribution region and the environmental factors[J]. J Nanjing For Univ Nat Sci Ed, 2014, 38(suppl 1):74-80. [12] 李蒙, 严邦祥, 赵昌高, 等.大仰山高山湿地山樱花种群数量结构特征[J].南京林业大学学报(自然科学版), 2013, 37(5):40-44. LI Meng, YAN Bangxiang, ZHAO Changgao, et al. Numeric and structural characteristics of Cerasus serrulata population around the high-elevation wetlands of Dayangshan[J]. J Nanjing For Univ Nat Sci Ed, 2013, 37(5):40-44. [13] 王贤荣, 谢春平, 伊贤贵, 等.不同居群野生早樱形态变异研究[J].植物研究, 2007, 27(6):746-752. WANG Xianrong, XIE Chunping, YIN Xiangui, et al. Study on the morphological variations of Cerasus subhirtella var. ascendens in different populations[J]. Bull Botan Res, 2007, 27(6):746-752. [14] 商韬, 王贤荣, 南程慧, 等.基于SSR标记的迎春樱自然居群遗传多样性分析[J].甘肃农大学报, 2013, 48(6):104-109. SHANG Tao, WANG Xianrong, NAN Chenghui, et al. Genetic diversity in natural populations of Cerasus discoidea based on SSR markers[J]. J Gansu Agric Univ, 2013, 48(6):104-109. [15] TSUDA Y, KIMURA M, KATO S, et al. Genetic structure of Cerasus jamasakura, a Japanese flowering cherry, revealed by nuclear SSRs:implications for conservation[J]. J Plant Res, 2009, 122(4):367-375. [16] NAS M N, BOLEK Y, BARDAK A. Genetic diversity and phylogenetic relationships of Prunus microcarpa C.A. Mey. subsp. tortusa analyzed by simple sequence repeats (SSRs)[J]. Sci Hortic, 2011, 127(3):220-227. [17] ANTONIUS K, AALTONEN M, UOSUKAINEN M, et al. Genotypic and phenotypic diversity in finnish cultivated sour cherry (Prunus cerasus L.)[J]. Gen Resour Crop Evol, 2012, 59(3):375-388. [18] 赵庆杰, 李海波, 屈燕, 等.基于SCAR标记的20个樱花品种的分子鉴别[J].南京林业大学学报(自然科学版), 2016, 40(5):34-40. ZHO Qingjie, LI Haibo, QU Yan, et al. SCAR markers-based molecular identification of 20 flowering cherries(Cerasus) cultivars[J]. J Nanjing For Univ Nat Sci Ed, 2016, 40(5):34-40. [19] SHI Shuo, LI Jinlu, SUN Jiahui, et al. Phylogeny and classification of Prunus sensulato(Rosaceae)[J]. J Integrative Plant Biol, 2013, 55(11):1069-1079. [20] 付涛, 王志龙, 林立, 等.樱属植物种质资源系统鉴定方法的研究[J].园艺学报, 2015, 42(12):2455-2468. FU Tao, WANG Zhilong, LIN Li, et al. Study on the identification method of a system of the Cerasus plants germplasm resources[J]. Acta Hortic Sin, 2015, 42(12):2455-2468. [21] 顾宇, 史港影, 李祯, 等.樱属植物核型参数及种间亲缘关系分析[J].南京林业大学学报(自然科学版), 2014, 38(增刊1):25-29. GU Yu, SHI Gangying, LI Zhen, et al. Karyotype parameters analysis and genetic relationship discussion of Cerasus (Rosaceae)[J]. J Nanjing For Univ Nat Sci Ed, 2014, 38(suppl 1):25-29. [22] 王贤荣, 向其柏.樱属植物叶腺体形态研究及其在分类中的意义[J].南京林业大学学报(自然科学版), 1997, 21(4):63-67. WANG Xianrong, XIANG Qibai. Studies on morphology of foliar glands and its taxonomical significance of Cerasus(Rosaceae)[J]. J Nanjing For Univ Nat Sci Ed, 1997, 21(4):63-67. [23] 伊贤贵, 王贤荣, 谢春平, 等.武夷山樱属植物叶表皮形态扫描电镜观察[G]//中国植物学会.中国植物学会七十五周年年会论文摘要汇编(1933-2008).兰州: 兰州大学出版社, 2008: 316. YIN Xiangui, WANG Xianrong, XIE Chunping, et al. Observation of leaf epidermis morphology of Cerasus campanulata var.wuyiensis by scanning electron microscopy[G]//Botanical Society of China. Summary Compilation of Papers for the 75th Anniversary Annual Meeting of Botanical Society of China (1933-2008). Lanzhou: Lanzhou University Press, 2008: 316. [24] 雷海清.樱属花粉形态研究[J].亚热带植物科学, 2001, 30(4):14-17. LEI Haiqing. A study on pollen morphology of Cerasus[J]. Subtrop Plant Sci, 2001, 30(4):14-17. [25] 叶宏宝, 石晓燕, 李冬, 等.浙江省农业气候资源时空变化特征研究[J].浙江农业学报, 2014, 26(4):1021-1030. YE Hongbao, SHI Xiaoyan, LI Dong, et al. Spatiotemporal characteristics of agricultural climate resources in Zhejiang Province[J]. Acta Agric Zhejiang, 2014, 26(4):1021-1030. [26] 尚建立, 王吉明, 郭琳琳, 等.西瓜种质资源主要植物学性状的遗传多样性及相关性分析[J].植物遗传资源学报, 2012, 13(1):11-15. SHANG Jianli, WANG Jiming, GUO Linlin, et al. Genetic diversity and correlation analysis of main botany characters in watermelon genetic resources[J]. J Plant Gen Resour, 2012, 13(1):11-15. [27] 徐斌, 彭莉霞, 杨会肖, 等.杜鹃红山茶叶片主要性状的遗传多样性分析[J].植物研究, 2015, 35(5):730-734. XU Bin, PENG Lixia, YANG Huixiao, et al. Genetic diversity analysis for leaf main traits of Camellia azalea[J]. Bull Bot Res, 2015, 35(5):730-734. [28] 王庆芬.轮叶百合形态特征多样性调查研究[J].北方园艺, 2014(21):78-82. WANG Qingfen. Research on morphological characteristics diversity of the Lilium distichum[J]. Northern Hortic, 2014(21):78-82. [29] 唐军荣, 李斌, 朱丽娜, 等.滇杨多倍体苗期叶片形态及光合生理比较分析[J].林业科学研究, 2016, 29(1):103-109. TANG Junrong, LI Bin, ZHU Lina, et al. Analysis on leaf morphology and photosynthesis physiology of polyploidized Populus yunnanensis seedlings[J]. For Res, 2016, 29(1):103-109. [30] 张林, 罗天祥.植物叶寿命及其相关叶性状的生态学研究进展[J].植物生态学报, 2004, 28(6):844-852. ZHANG Lin, LUO Tianxiang. Advances in ecological studies on leaf lifespan and associated leaf traits[J]. Acta Phytoecol Sin, 2004, 28(6):844-852. [31] 于万里, 张博.新疆昭苏野生黄花苜蓿果实形态变异研究[J].草业学报, 2012, 21(2):249-255. YU Wanli, ZHANG Bo. A study on morphological variation of pods and seeds of Medicago falcata collected from Zhaosu, Xinjiang[J]. Acta Pratac Sin, 2012, 21(2):249-255. [32] 顾晓燕, 郭志慧, 张新全, 等.短芒披碱草异位保护群体的表型多样性研究[J].草业学报, 2015, 24(5):141-152. GU Xiaoyang, GUO Zhihui, ZHANG Xinquan, et al. Phenotypic variations in seven ex-situ conservation populations of Elymus breviaristatus[J]. Acta Pratac Sin, 2015, 24(5):141-152. [33] 王旭军, 程勇, 吴际友, 等.红榉不同种源叶片形态性状变异[J].福建林学院学报, 2013, 33(3):284-288. WANG Xujun, CHENG Yong, WU Jiyou, et al. Variation of leaf morphologic characters of Zelkova schneideriana Hand-Mazz. from different provenances[J]. J Fujian For Coll, 2013, 33(3):284-288. [34] 朱利利, 吴疆翀, 彭兴民, 等.印楝属植物表型变异分析[J].林业科学研究, 2016, 29(2):162-166. ZHU Lili, WU Jiangchong, PENG Xingmin, et al. Phenotypic difference among species and a variation type of Azadirachta[J]. For Res, 2016, 29(2):162-166. [35] 董文婧, 张雪, 申仕康, 等.都市园林植物云南樱花的表型多样性研究[J].广西植物, 2016, 36(3):349-354. DONG Wenjing, ZHANG Xue, SHEN Shikang, et al. Phenotypic diversity of urban landscape plant Prunus yunnanensis[J]. Guihaia, 2016, 36(3):349-354. [36] 梁振旭, 张延龙, 牛立新, 等.秦巴山区野百合表型多样性[J].广西植物, 2014, 34(6):727-733. LIANG Zhenxu, ZHANG Yanlong, NIU Lixin, et al. Phenotypic diversity of Lilium brownii native to Qinba mountainous area[J]. Guihaia, 2014, 34(6):727-733. -

-
链接本文:
https://zlxb.zafu.edu.cn/article/doi/10.11833/j.issn.2095-0756.2019.04.012
 点击查看大图
点击查看大图
计量
- 文章访问数: 5017
- HTML全文浏览量: 1070
- PDF下载量: 70
- 被引次数: 0
