





-
随着公益林建设的不断深化,其改善环境、维持生态平衡的作用逐渐体现。然而公益林在封山育林的纯自然演替状态下群落结构恢复较慢,对不同立地条件下复杂多样的公益林类型至今也没有较为完善的差别化经营管理策略。公益林的生态效益和社会效益还存在较大的提升潜力。因此利用合适的方法来对公益林群落进行分类和排序,理解不同立地条件下公益林的分布类型以及两者之间的相互关系是十分必要的。对植被群落进行分类和排序能了解研究区域群落结构特征并揭示植物群落间以及群落与环境因子之间的联系[1]。分类能够在一定程度上探明植物群落的结构和组成[2],排序则能表明物种、群落分布与环境梯度上的关系[3]。群落生态学上目前运用最为常见的排序方法有除趋势对应分析(DCA)、典范对应分析(CCA)和除趋势典范对应分析(DCCA)等[4-6]。CCA分析中结合了物种数据和多个环境数据,且每一步计算都会与环境因子进行回归。CCA相比较于聚类法、多元回归树(MRT)具有更好的效果[7]。种间关联性指群落内不同物种在空间分布上的相互关联性[8-9],是植物群落重要的数量和结构指标[10-12]。数量分析中的多个分析方法和指标相结合对植物群落进行研究,能更全面地解读植被群落与环境之间的复杂关系及其群落结构特征[13-14]。本研究基于浙江省松阳县128个样地为研究对象,采用双向指示种分析(TWINSPAN)、CCA排序和多样性指数等方法,研究了松阳县公益林群落结构及其在环境梯度上的分异规律,以期为不同分类管理模式和不同立地条件下公益林生物多样性维持和提升提供理论依据。
-
松阳县(28°14′~28°36′N,119°10′~119°42′E)位于浙江省西南部,全县总面积为14.06 万hm2,公益林面积为6.38 万hm2,占林业用地面积的55.0%,森林覆盖率75.0%,林木绿化率75.9%[15-16]。气候属亚热带季风气候,温暖湿润,四季分明,无霜期长,具有一定的山地立体气候。林地土壤有红壤、黄壤、岩性土、潮土等4类,其中红壤和黄壤为主要的土壤类型。松阳县地带性植被属于中亚热带常绿阔叶林,现有森林的乔木林林分面积中马尾松Pinus massoniana林、杉木Cunninghamia lanceolata林、阔叶林、经济林的面积比为39∶36∶17∶8[17]。群落类型以阔叶林、针阔混交林和针叶林群落为主,优势树种有马尾松、杉木、枫香Liquidambar formosana、木荷Schima superba、青冈Cyclobalanopsis glauca、苦槠Castanopsis sclerophylla、麻栎Quercus acutissima等[18]。
-
以浙江省松阳县的公益林作为总体样本,按一定比例随机抽取监测小班,在每个小班中选取1个最能代表该小班状况的区域设置20 m×20 m的固定监测样地,共设128个。使用哈尔滨牌DQL-12Z型12倍正像森林罗盘仪进行打样地,闭合差控制在1/200以内。对样地内乔木树种(胸径≥5.0 cm)采用每木调查,记录树种名称、胸径、树高、冠幅、枝下高等数据。记录样地基本地理信息和环境因子,包括海拔、坡度、坡位、坡向、土壤类型、腐殖质厚度和凋落物厚度[19],并利用手持GPS仪记录样地经纬度坐标。该县被抽中的128个样地中包含马尾松林、杉木林、针阔混交林、阔叶林、竹林等林分类型。样地分布涵盖了中山、低山、丘陵、盆地等地形,海拔为115~1 278 m。
-
各个种群的重要值反映种群在群落中的作用和地位,计算公式[18]如下:乔木层重要值=(相对显著度+相对频度+相对多度)/3×100%;相对显著度=某一树高胸高断面积之和/所有树种胸高断面积之和×100%;相对多度=某一树种株数之和/所有树种株数之和×100%;相对频度=某一树种频度/所有树种的频度和×100%。
-
TWINSPAN是在指示种分析(indicator species analysis)的基础上改进而来的。它以二岐式的分割法来划分类型,其划分根据“指示种”(或称为指示指标)将样地类型与物种组成依次划分为各个等级的类型单位,完成物种分类和样地分类[20]。本研究依据物种重要值排序,选取重要值大于0.1的乔木层树种,利用WinTWINS 2.3软件分析[20]。重要值采用5个等级作为样地划分的指示,分别为0~0.05、0.05~0.10、0.10~0.30、0.30~0.50、>0.50。
CCA是一种非线性多元直接梯度分析方法,它把对应分析与多元回归结合起来,每一步计算结果都与环境因子进行回归,研究物种与环境的关系[21]。本研究选取坡位、坡度、坡向、海拔高度、土壤类型、腐殖质厚度、凋落物厚度为环境因子。将这些环境因子建立矩阵,在CANOCO 4.5中进行分析。此外,样地分布环境涵盖较广,也可以使得CCA排序的结果更加可靠。
-
采用Shannon-Wiener多样性指数、Simpson指数、Pielou均匀指数和物种丰富度作为衡量群落生物多样性的指标[22]。用SPSS单因素方差分析比较不同群丛的物种多样性差异。使用R 3.6.1的boxplert包绘制箱型图[23]。由于χ2检验可判断种间关联性程度的强弱,但是无法定量关联程度大小,因此采用联结系数和共同出现百分率对研究区11个优势种(重要值>1.1%)进行种间联结分析[24-27]。利用Excel 2019进行数据整理,使用R 3.6.1的spaa包进行种间关联分析,corrplot包进行种间关联半矩阵图输出。
-
表1列出了128个样地中重要值排名前20的物种,主要有杉木、马尾松、毛竹、木荷、青冈、甜槠、苦槠、石栎等树种。
表 1 松阳县公益林128个样地主要物种的重要值
Table 1. Importance value of 128 sample plots in Songyang County
排序号 物种 重要值/% 1 杉木 Cunninghamia lanceolata 30.83 2 马尾松 Pinus massoniana 12.78 3 毛竹 Phyllostachys edulis 9.28 4 木荷 Schima superba 5.59 5 青冈 Cyclobalanopsis glauca 3.70 6 甜槠 Castanopsis eyrei 3.07 7 苦槠 Castanopsis sclerophylla 2.19 8 石栎 Lithocarpus glaber 1.88 9 白栎 Quercus fabri 1.49 10 黄山松 Pinus taiwanensis 1.41 11 赤杨叶 Alniphyllum fortunei 1.10 12 樟树 Cinnamomum camphora 1.03 13 板栗 Castanea mollissima 1.02 14 枫香 Liquidambar formosana 1.01 15 杨梅 Myrica rubra 0.94 16 山合欢 Albizia kalkora 0.93 17 米槠 Castanopsis carlesii 0.90 18 南酸枣 Choerospondias axillaris 0.74 19 漆树 Toxicodendron verniciflnum 0.68 20 野樱桃 Cerasus pseudocerasus 0.68 -
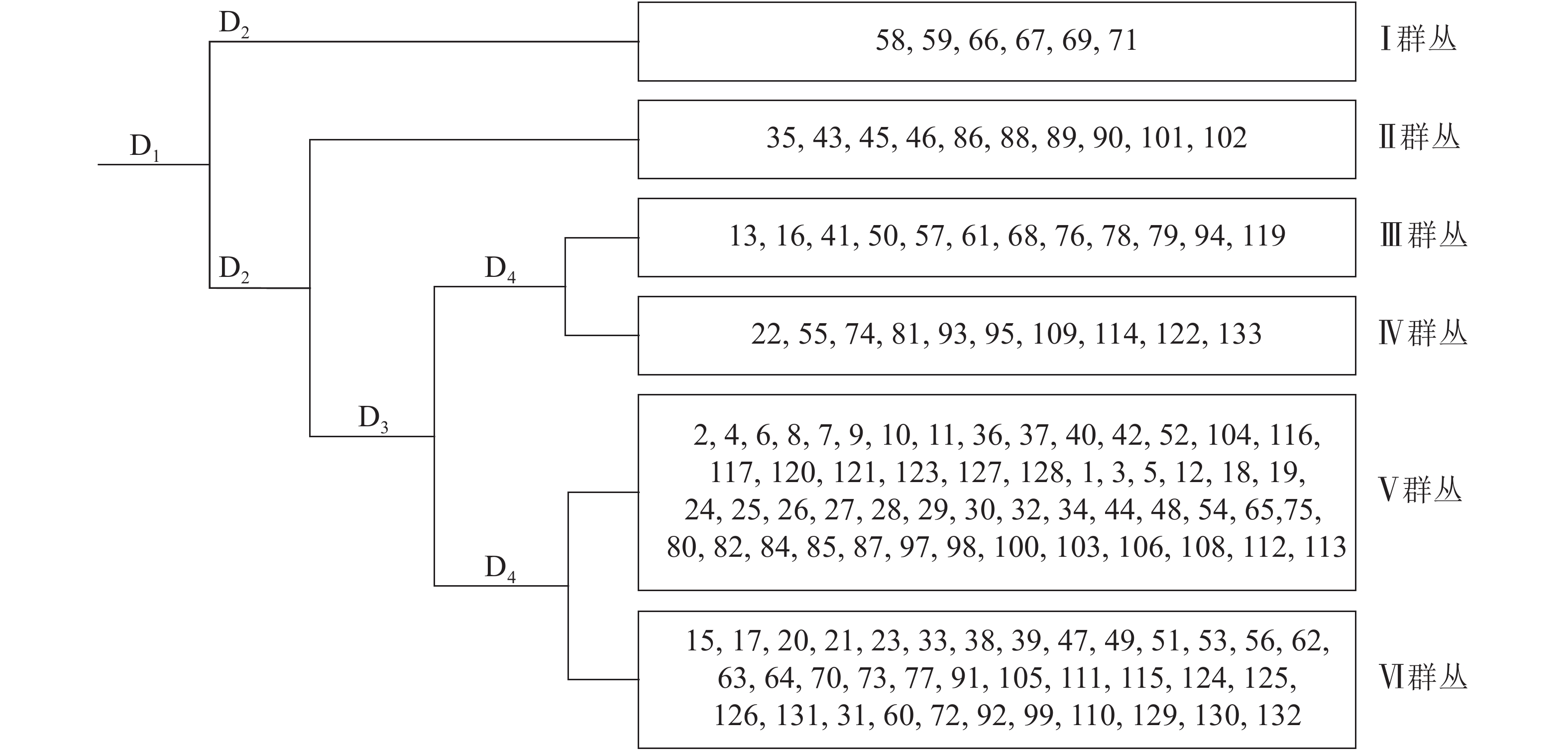
采用研究区128个样地重要值矩阵数据在TWINSPAN中进行群丛划分,得到6个群丛类型,每个群丛的样地及环境信息如图1和表2所示。其中:群丛Ⅰ为板栗+杉木+杨梅群丛,群丛Ⅱ为毛竹+杉木+马尾松群丛,群丛Ⅲ为马尾松+杉木+油茶Camellia oleifera群丛,群丛Ⅳ为马尾松+木荷+杉木群丛,群丛Ⅴ为杉木+木荷+马尾松群丛,群丛Ⅵ为杉木+马尾松+木荷群丛。
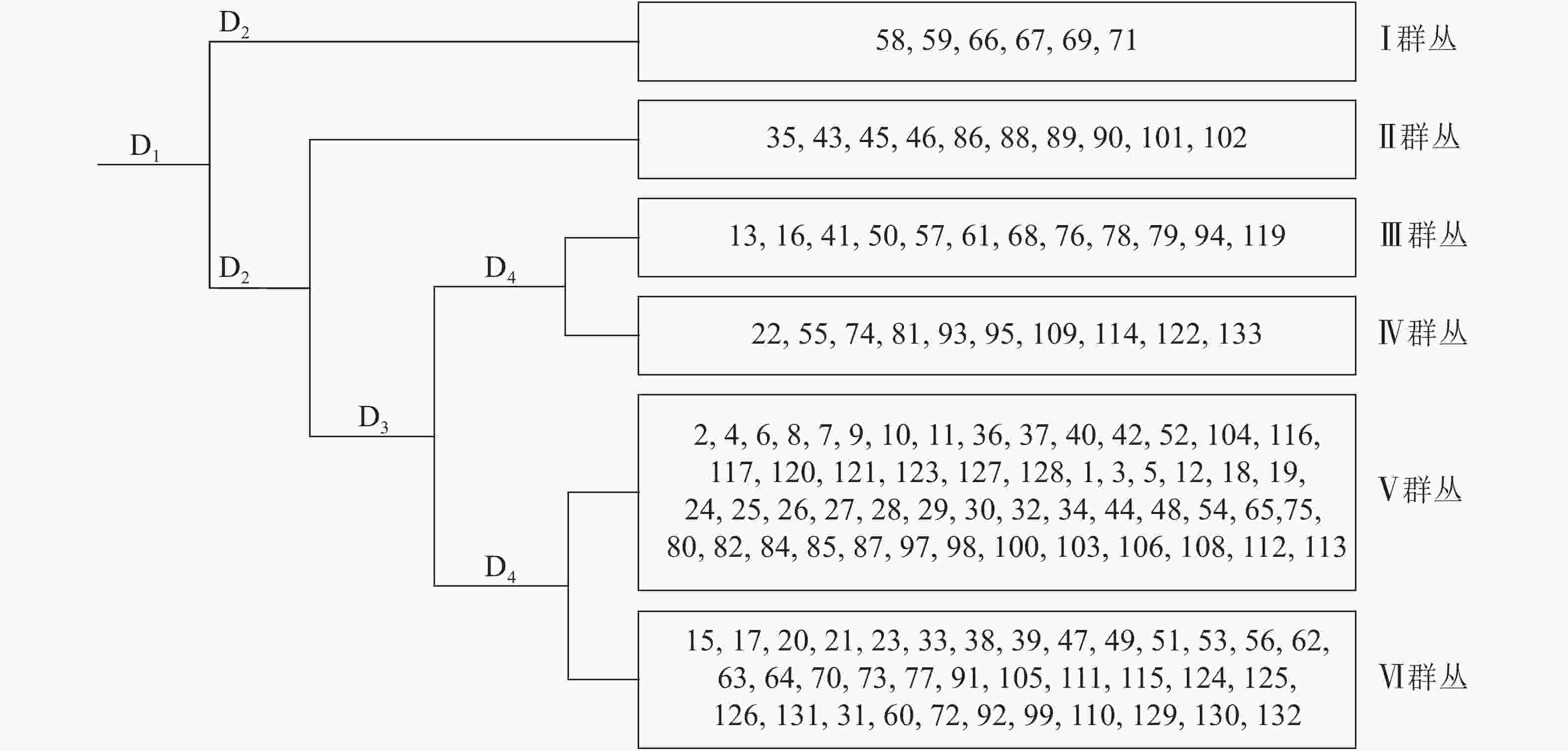
图 1 松阳县公益林128个样方TWINSPAN分类等级
Figure 1. Dendrogram illustrating the presence of six vegetation associations using TWINSPAN analysis of 128 plots in Songyang non-commercial forest
表 2 松阳县公益林TWINSPAN分类群丛及其环境因子
Table 2. Community type identified by TWINSPAN and the environment variables in Songyang non-commercial forest
群丛 海拔/m 土壤类型 坡向 坡度/(°) 腐殖质厚度/cm 凋落物厚度/cm Ⅰ. 板栗+杉木+杨梅 119~390 红壤 西和西南 25~35 12~18 3~5 Ⅱ. 毛竹+杉木+马尾松 350~1 080 红壤和黄壤 全坡 10~40 2~20 2~12 Ⅲ. 马尾松+杉木+油茶 117~511 红壤 全坡 15~41 1~15 1~10 Ⅳ. 马尾松+木荷+杉木 142~516 红壤 南 20~41 2~15 1~16 Ⅴ. 杉木+木荷+马尾松 145~1 278 红壤和黄壤 全坡 27~49 1~20 1~15 Ⅵ. 杉木+马尾松+木荷 127~1 074 红壤和黄壤 全坡 13~42 1~40 1~20 -
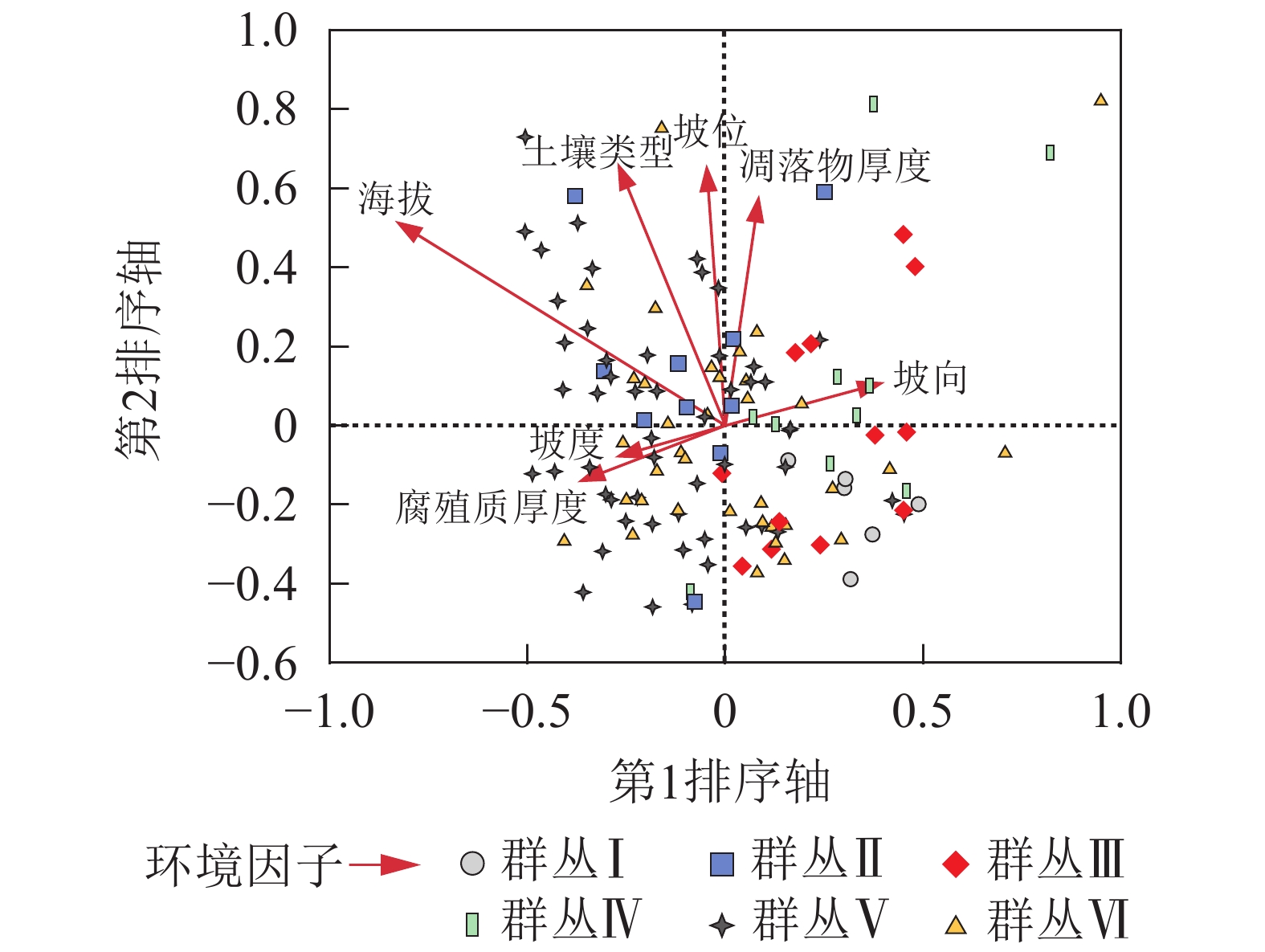
第1轴和第2轴与环境因子的特征值分别为0.302和0.167,物种环境相关性分别为0.718和0.555,包含了65.7%的生态信息。因此本研究使用第1轴和第2轴的数据来分析样地中物种和周围环境之间的关系(表3)。CCA排序图的每个箭头代表1种环境因子,箭头的长度表示该环境因子对物种的影响大小[28-29]。从图2可看出:CCA排序第1轴与海拔高度呈反比,相关系数为−0.583,与坡向呈正相关0.271(表3)。第2轴与土壤类型、坡位和凋落物厚度相关性较高。
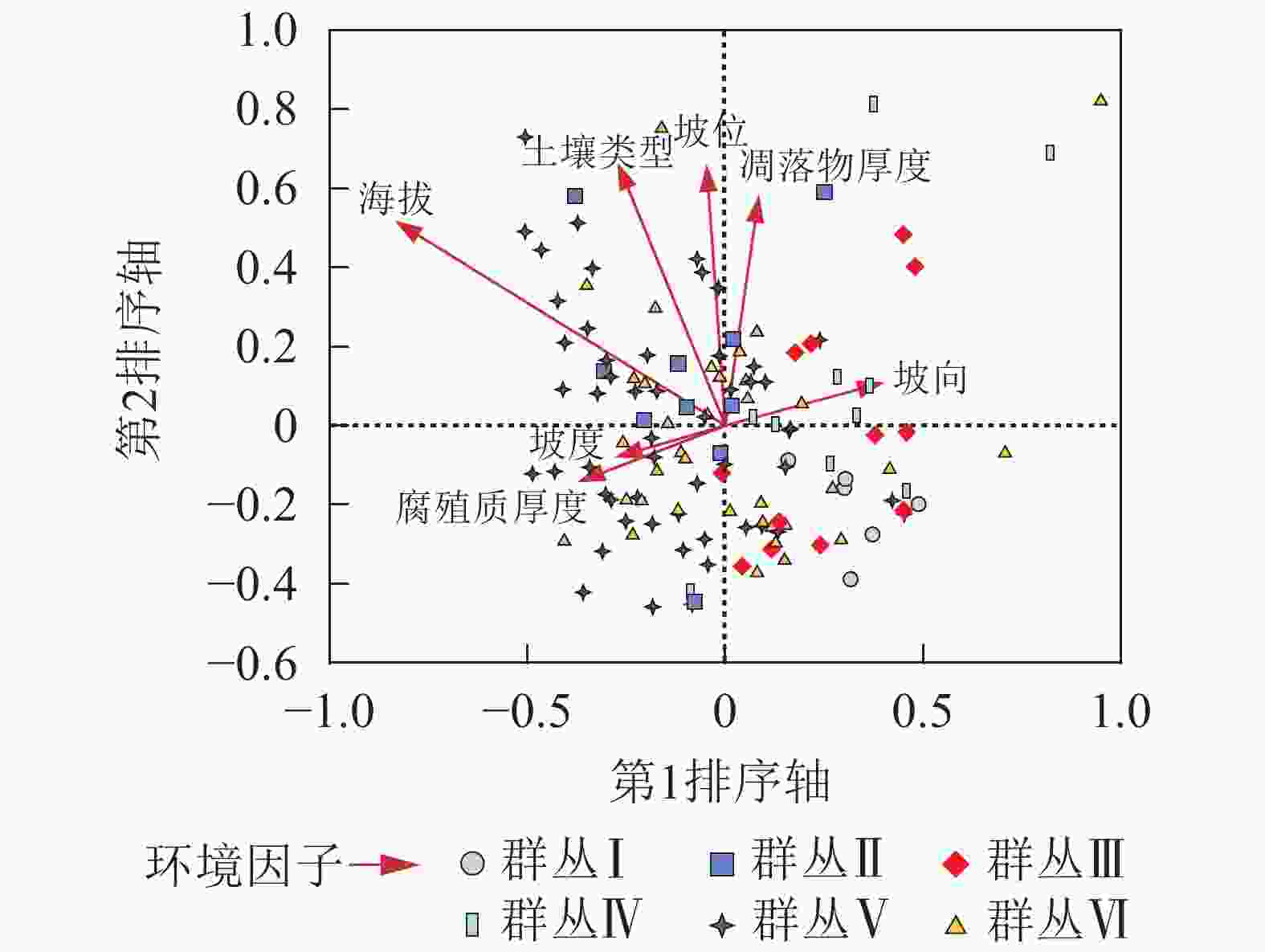
图 2 松阳公益林128个样方的CCA二维排序
Figure 2. CCA ordination diagram of 128 plots in Songyang non-commercial forest
表 3 松阳县公益林128个样地CCA排序前4轴系数与环境因子的相关性
Table 3. Correlations between environmental variables and CCA ordination axes of 128 plots in Songyang non-commercial forest
环境因子 轴1 轴2 轴3 轴4 坡位 −0.033 0.352 0.233 −0.158 坡度 −0.182 −0.040 0.064 0.100 坡向 0.271 0.057 −0.144 0.004 海拔高度 −0.583 0.280 −0.093 0.029 土壤类型 −0.189 0.356 −0.189 0.159 腐殖质厚度 −0.250 −0.074 0.028 0.123 枯落物厚度 0.058 0.309 0.044 −0.081 特征值 0.302 0.167 0.082 0.062 物种环境相关性 0.718 0.555 0.402 0.378 物种环境关系变异累计百分数/% 42.3 65.7 77.2 85.9 -
从图3可见:6个群丛在物种多样性指数分析中存在显著差异(P<0.001)。其中多样性指数(Shannon-Wiener指数和Simpson指数)变化趋势基本一致,群丛Ⅴ的Shannon-Wiener指数和Simpson指数显著高于其他群丛(P<0.001),且群丛Ⅰ和Ⅱ的Simpson指数与群丛Ⅲ差异显著(P<0.001);群丛Ⅲ的Pielou指数与除群丛Ⅴ之外的群丛差异显著(P<0.001),群丛Ⅴ与群丛Ⅰ、Ⅱ存在显著差异(P<0.001);物种丰富度显示群丛Ⅴ显著高于其他群丛(P<0.001)。
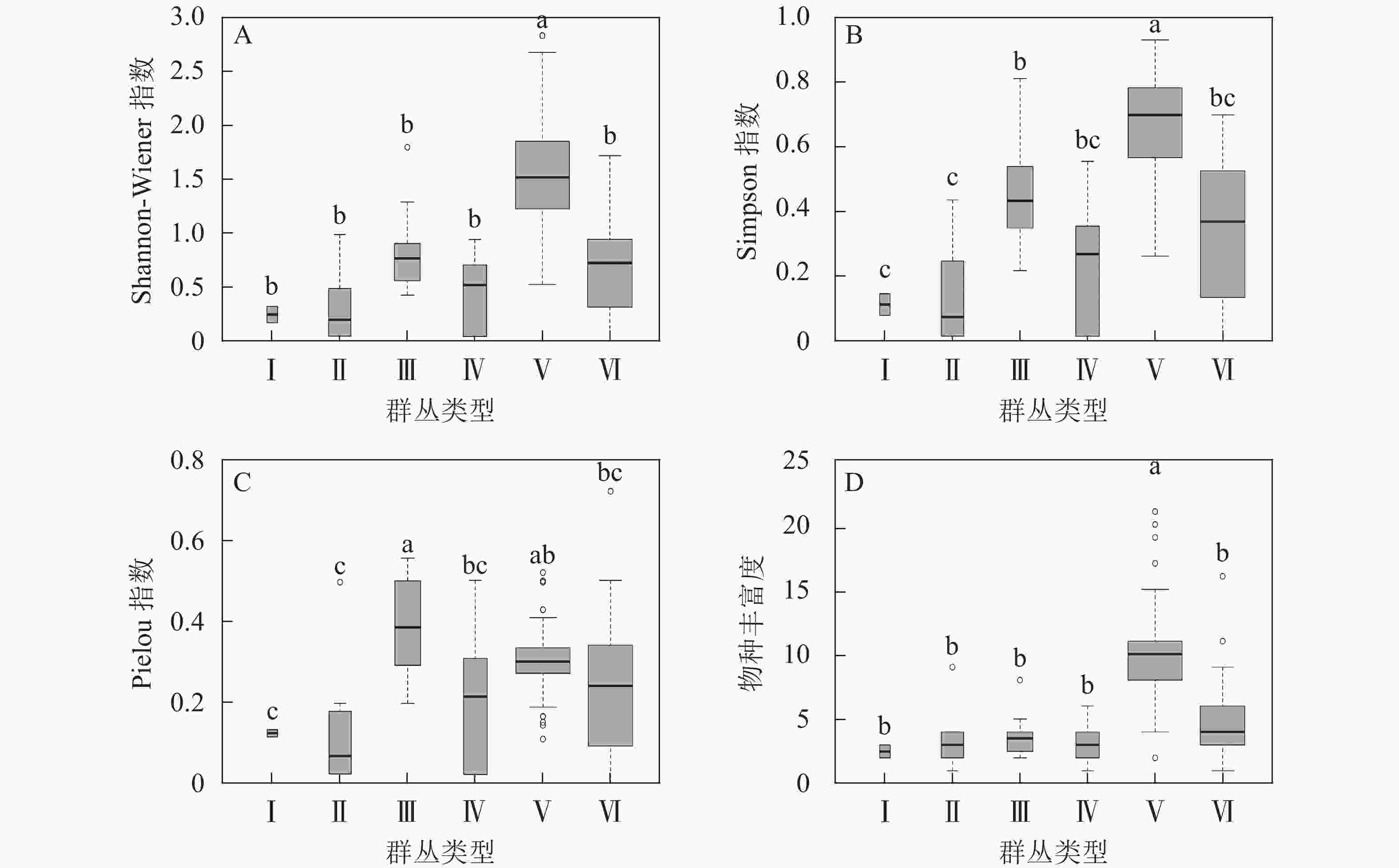
图 3 6个群丛的物种多样性指数变化规律
Figure 3. Changes of species diversity index among six associations
-
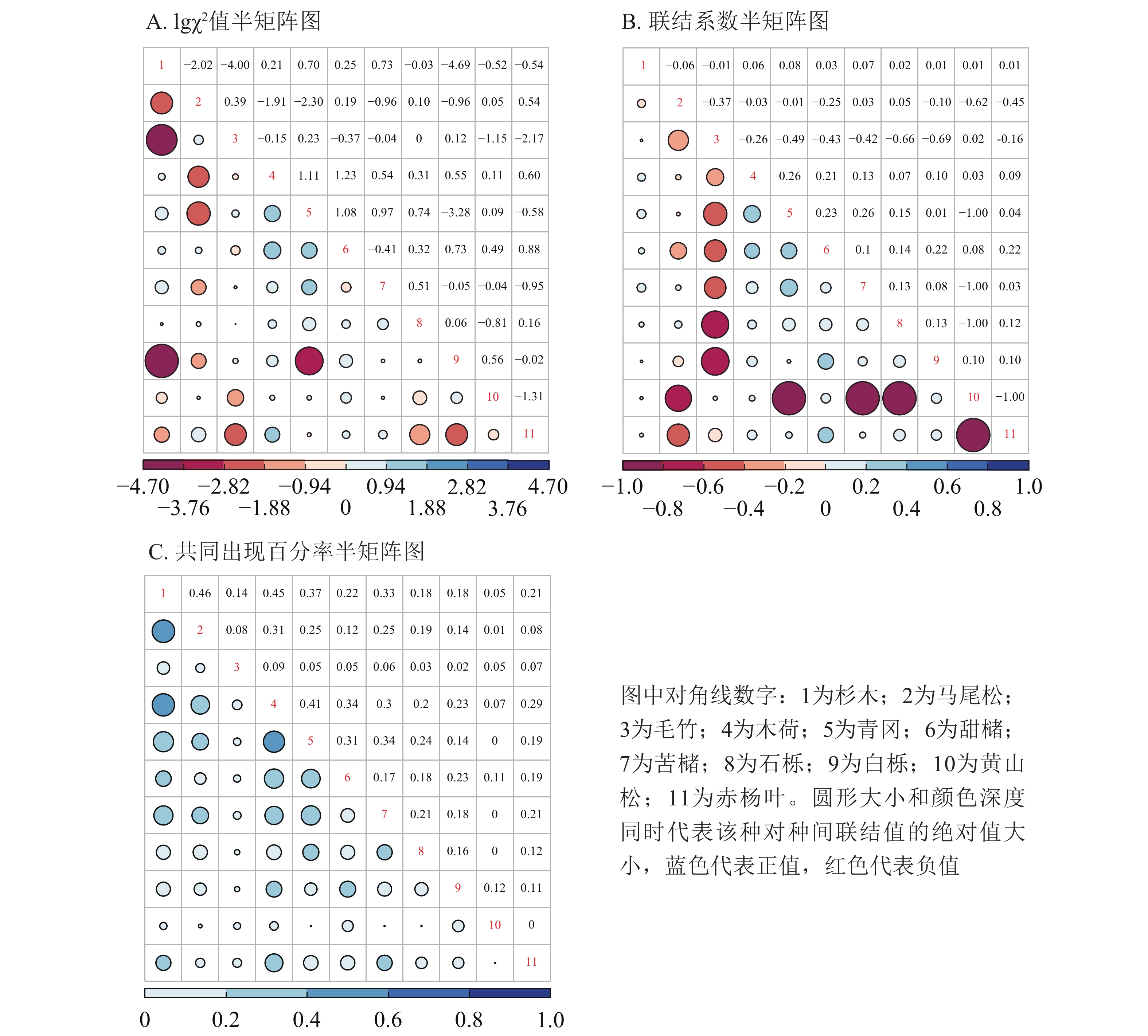
选择重要值排名前11位的树种,分别计算其两两之间的χ2值、联结系数和共同出现百分率,并分别做半矩阵图。将χ2结果对数化,lg3.841=0.584,lg6.635=0.822。如图4A所示:4个种对lgχ2处于[0.584,0.822]内,6个种对lgχ2>0.822,表明有4个(7.2%)种对呈显著关联,6个(10.9%)种对间关联极显著。表现极显著联结的种对有:马尾松对赤杨叶、木荷对甜槠、木荷对青冈、木荷对赤杨叶、青冈对苦槠、青冈对甜槠。表现显著联结的种对有:杉木对青冈、杉木对苦槠、青冈对石栎、青冈对白栎。图4B表明:优势物种种间关系有36对(65.5%)呈正关联,19对(34.5%)呈负关联,正负关联比为1.9。其中,联结系数
处于[0.5,1.0]值域的有0对,处于[0.2,0.5]值域的有6对(10.9%):木荷对青冈、木荷对甜槠、青冈对甜槠、青冈对苦槠、甜槠对白栎、甜槠对赤杨叶。而处于[−1.0,−0.5]值域的有7对(12.7%):马尾松对黄山松、毛竹对石栎、毛竹对白栎、青冈对黄山松、苦槠对黄山松、石栎对黄山松、黄山松对赤杨叶,处于[−0.5,−0.2]值域的有6对(10.9%):马尾松对毛竹、马尾松对甜槠、马尾松对赤杨叶、毛竹对木荷、毛竹对青冈、毛竹对甜槠。根据共同出现百分率矩阵图(图4C)可知:共同出现百分率 处于[0.4,0.7]的种对有3对,占总种对数的5.5%,这些种对呈显著正联结。共同出现百分率 处于[0.2,0.4]的种对有17对,占总种对数的30.9%,这些种对为不显著正联结。处于[0,0.2]的种对有35对,占总种对数的63.6%,这些种对几乎无联结。 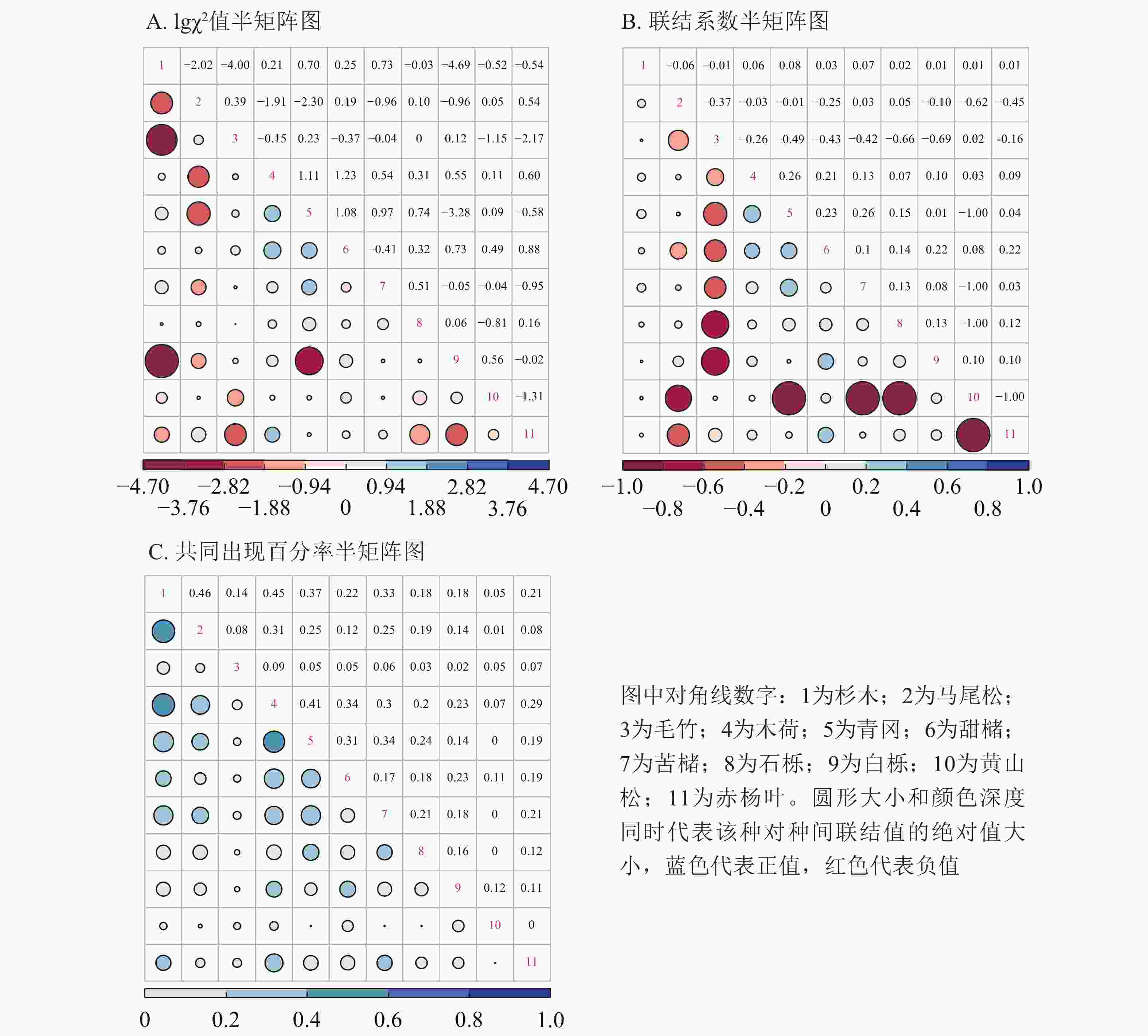
图 4 研究区11个优势种种间关联半矩阵图
Figure 4. Semi-matrices graphs of interspecific association of 11 dominant species
-
CCA第1轴排序主要与海拔、坡向以及土壤有关,本质上反映植物对水热和阳光需求的变化,这与钟娇娇等[12]对秦岭山地天然次生林进行排序分类的结论一致。群落的形成与环境因子密不可分,通常是气候、地形和土壤等因子的综合作用,而这些环境因子在不同地理空间上对于植物群落格局形成的影响也不同[30]。本研究结果中影响群落分布的第1主导因子是海拔,第2主导因子是土壤类型。陈云等[31]在小秦岭的研究表明:森林群落物种组成和分布格局的前2个主要环境因子是分别是海拔和坡度,其中第2主导因子与本研究结果存在差异,这种差异在与叶诺楠等[32]的研究比较中也得到了体现。可见在不同的地理和气候区划中,环境因子对植被群落分布和类型的影响是不同的。
-
在不同的立地条件下松阳县公益林的群落组成存在显著差异,同时物种多样性在环境梯度上也呈现一定分异规律,这与早期的研究也较为接近[32]。本研究中,杉木+木荷+马尾松群丛(V)的Shannon-Wiener指数高于其他群丛,可能由于群丛Ⅴ在各海拔均有分布且林分为混交林,因此群落物种结构复杂、多样性高。而其他群丛有些在单一环境下分布,群落组成简单、结构单一,故群落物种多样性低。此外,毛竹入侵造成群落中的光照和土壤等环境条件变化[33],这可能将造成乔木和灌木多样性降低[34],这也是毛竹为主导的群丛多样性较低的主要原因之一。
-
研究区大多数乔木树种呈现不显著联结,说明乔木层的物种间联系不明显,趋于独立分布,这也有可能是生境异质性导致的[35]。同时,χ2检验难以定量确定种间联结程度的大小,因此再采用AC值和PC值同时进行种间联结强度测定能够从不同角度阐明物种种间关联特性。样地内呈现正联结的种对数多于负联结,说明乔木层群落结构尚不稳定[36]。物种入侵将影响群落间的种间关联性,一些入侵的物种会降低群落的正关联性,增加负联结性[37]。
-
CCA结果显示群丛Ⅰ的样地数量不多,主要位于低海拔的阳坡,早期为杉木林内伐除杉木后人为种植的板栗和杨梅等经济林,为便于经营管理,近年来被划入公益林内。该类林分结构单一,生物多样性低,但立地条件尚可,可以较大强度地间伐板栗和杉木,补植木荷、青冈等珍贵树种,逐步改造为常绿落叶阔叶混交林。需要引起高度重视的是群丛Ⅱ,即毛竹群丛,样地数量不少,但生物多样性是研究区所有类型中最低的。以毛竹为代表的鞭生竹克隆生长使其能够入侵相邻林分,并逐渐发展为新的“优势种”,成为一种“局部入侵”物种,影响周围森林群落结构、土壤质量、幼苗更新及幼树生长,从而影响整个森林生态系统。因此,加强公益林内毛竹群丛的林分改造,防止竹林的无序扩张是今后公益林管理的重要问题。在CCA排序图中,样地众多的群丛Ⅴ和群丛Ⅵ主要分布在海拔500~1 000 m且土层厚度较好的生境中,林内物种较为丰富,以木荷为代表的阔叶树种在乔木层占有一定的优势,今后可以适当间伐杉木、马尾松等针叶树,对阔叶树种实施定株抚育或目标树经营,促进阔叶树的生长,加速形成以阔叶树为主的林分类型。
-
本研究对松阳县生态公益林群落进行数量分类与排序,分析了多样性指数和种间关系,提出了公益林的差别管理措施建议。在日后公益林管理工作中,可考虑对不同立地条件不同林分的公益林进行不同的补植措施,对毛竹林进行边缘控制,对一些能适应各类环境的群落组合进行适当推广等措施。
Classification, ordination and correlation analysis of dominant species of ecological non-commercial forests in Songyang, Zhejiang
-
摘要:
目的 对浙江松阳县生态公益林群落进行分类和排序,探讨多样性变化规律及种间关系,有助于揭示植物群落和多样性在复杂环境梯度上的分布规律。 方法 以松阳县重点公益林128个固定监测样地为研究对象,利用双向指示种分析(TWINSPAN)、典范对应分析(CCA)、物种多样性指数及种间联结分析等对该县公益林群落特征展开研究。 结果 TWINSPAN结果显示:松阳公益林群落可分为6种群丛类型。CCA结果显示:松阳县公益林群落分布和类型在环境梯度上呈现明显的分异规律,其主要受到海拔、土壤类型和坡向的影响。物种多样性结果表明:针阔混交林和阔叶林为主导的群丛Shannon-Wiener指数和Simpson指数显著高于其他群丛,以毛竹Phyllostachys edulis为主导的群丛多样性显著低于其他群丛。种间联结分析显示:样地内大多数乔木树种呈现不显著联结,正负关联比为1.9,说明群落整体稳定。 结论 松阳县公益林应进行差别化管理,积极推广针叶林混交化和阔叶化。可考虑对不同立地条件不同林分的公益林进行不同的补植措施,对毛竹林进行边缘控制,对一些能适应各类环境的群落组合进行适当推广。图4表3参37 Abstract:Objective This study aims to indicate the distribution of forest community and species diversity on the gradients of complex environment by classifying and sorting the community of ecological non-commercial forest in Songyang County, Zhejiang Province, and discussing the change law of diversity and interspecific relationship. Method Taking 128 monitoring plots as research projects, two-way indicator species analysis (TWINSPAN), canonical correspondence analysis (CCA), species diversity index and interspecific association analysis were used to study the community characteristics of non-commercial forest in Songyang County. Result TWINSPAN results showed that the forest community could be divided into 6 types. CCA results demonstrated that the community distribution and types of non-commercial forest in Songyang County showed obvious differentiation on environmental gradient, which was mainly affected by altitude, soil type and slope aspect. The results of multiple comparison of species diversity showed that the Shannon-Wiener index and Simpson index of associations with mixed forest or broad-leaved forest were significantly higher than those of other associations, and the diversity of association dominated by Phyllostachys edulis was significantly lower than that of other associations. Interspecific Association Analysis illustrated that most tree species in sample plots had no significant correlations. The positive and negative correlation ratio was 1.9, indicating the overall stability of the community. Conclusion Differential management should be carried out for the non-commercial forest in Songyang County, and mixed and broad-leaved forests should be actively promoted. It is possible to consider different supplementing measures for the non-commercial forests with different site conditions and different stands. The edge control of bamboo forests and community combinations that can adapt to various environments should be properly promoted. [Ch, 4 fig. 3 tab. 37 ref.] -
图 1 松阳县公益林128个样方TWINSPAN分类等级
图中1,2,3等代表样地号,D1,D2,D3和D4代表分类层数
Figure 1 Dendrogram illustrating the presence of six vegetation associations using TWINSPAN analysis of 128 plots in Songyang non-commercial forest
图 2 松阳公益林128个样方的CCA二维排序
Figure 2 CCA ordination diagram of 128 plots in Songyang non-commercial forest
图 3 6个群丛的物种多样性指数变化规律
字母组合表示各群丛间多样性差异,群丛间无相同字母表示差异显著(P<0.001),群丛间出现1个或以 上相同字母时则差异不显著。箱体宽度越大表示样地数量越多
Figure 3 Changes of species diversity index among six associations
图 4 研究区11个优势种种间关联半矩阵图
Figure 4 Semi-matrices graphs of interspecific association of 11 dominant species
表 1 松阳县公益林128个样地主要物种的重要值
Table 1. Importance value of 128 sample plots in Songyang County
排序号 物种 重要值/% 1 杉木 Cunninghamia lanceolata 30.83 2 马尾松 Pinus massoniana 12.78 3 毛竹 Phyllostachys edulis 9.28 4 木荷 Schima superba 5.59 5 青冈 Cyclobalanopsis glauca 3.70 6 甜槠 Castanopsis eyrei 3.07 7 苦槠 Castanopsis sclerophylla 2.19 8 石栎 Lithocarpus glaber 1.88 9 白栎 Quercus fabri 1.49 10 黄山松 Pinus taiwanensis 1.41 11 赤杨叶 Alniphyllum fortunei 1.10 12 樟树 Cinnamomum camphora 1.03 13 板栗 Castanea mollissima 1.02 14 枫香 Liquidambar formosana 1.01 15 杨梅 Myrica rubra 0.94 16 山合欢 Albizia kalkora 0.93 17 米槠 Castanopsis carlesii 0.90 18 南酸枣 Choerospondias axillaris 0.74 19 漆树 Toxicodendron verniciflnum 0.68 20 野樱桃 Cerasus pseudocerasus 0.68  下载: 导出CSV
下载: 导出CSV
表 2 松阳县公益林TWINSPAN分类群丛及其环境因子
Table 2. Community type identified by TWINSPAN and the environment variables in Songyang non-commercial forest
群丛 海拔/m 土壤类型 坡向 坡度/(°) 腐殖质厚度/cm 凋落物厚度/cm Ⅰ. 板栗+杉木+杨梅 119~390 红壤 西和西南 25~35 12~18 3~5 Ⅱ. 毛竹+杉木+马尾松 350~1 080 红壤和黄壤 全坡 10~40 2~20 2~12 Ⅲ. 马尾松+杉木+油茶 117~511 红壤 全坡 15~41 1~15 1~10 Ⅳ. 马尾松+木荷+杉木 142~516 红壤 南 20~41 2~15 1~16 Ⅴ. 杉木+木荷+马尾松 145~1 278 红壤和黄壤 全坡 27~49 1~20 1~15 Ⅵ. 杉木+马尾松+木荷 127~1 074 红壤和黄壤 全坡 13~42 1~40 1~20
下载: 导出CSV
表 3 松阳县公益林128个样地CCA排序前4轴系数与环境因子的相关性
Table 3. Correlations between environmental variables and CCA ordination axes of 128 plots in Songyang non-commercial forest
环境因子 轴1 轴2 轴3 轴4 坡位 −0.033 0.352 0.233 −0.158 坡度 −0.182 −0.040 0.064 0.100 坡向 0.271 0.057 −0.144 0.004 海拔高度 −0.583 0.280 −0.093 0.029 土壤类型 −0.189 0.356 −0.189 0.159 腐殖质厚度 −0.250 −0.074 0.028 0.123 枯落物厚度 0.058 0.309 0.044 −0.081 特征值 0.302 0.167 0.082 0.062 物种环境相关性 0.718 0.555 0.402 0.378 物种环境关系变异累计百分数/% 42.3 65.7 77.2 85.9
下载: 导出CSV
-
[1] 李晨晨, 周再知, 梁坤南, 等. 不同林药复合经营模式对杉木生态公益林土壤理化性质的改良效果[J]. 浙江农林大学学报, 2018, 35(1): 51 − 59. LI Chenchen, ZHOU Zaizhi, LIANG Kunnan, et al. Physical and chemical properties of ecological forest soils using different agroforestry patterns of Chinese fir with medicinal plants [J]. J Zhejiang A&F Univ, 2018, 35(1): 51 − 59. [2] 张文静, 张钦弟, 王晶, 等. 多元回归树与双向指示种分析在群落分类中的应用比较[J]. 植物生态学报, 2015, 39(6): 586 − 592. ZHANG Wenjing, ZHANG Qindi, WANG Jing, et al. A comparison of multivariate regression tree and two-way indicator species analysis in plant community classification [J]. Chin J Plant Ecol, 2015, 39(6): 586 − 592. [3] VIRTANEN R, LUOTO M, RÄMÄ T, et al. Recent vegetation changes at the high-latitude tree line ecotone are controlled by geomorphological disturbance, productivity and diversity [J]. Global Ecol Biogeogr, 2010, 19(6): 810 − 821. [4] KATZ V S. A model university/community-based intervention strategy to prevent harmful substance use [J]. Int J Ment Health Addict, 2018, 16(4): 860 − 863. [5] ZHAO Hui, WANG Qirui, FAN Wei, et al. The relationship between secondary forest and environmental factors in the southern Taihang Mountains [J]. Sci Rep, 2017, 7(1): 16431. [6] VITTOZ P, BAYFIELD N, BROOKER R, et al. Reproducibility of species lists, visual cover estimates and frequency methods for recording high-mountain vegetation [J]. J Veg Sci, 2010, 21(6): 1035 − 1047. [7] DE’ATH G. Multivariate regression trees: a new technique for modeling species-environment relationships [J]. Ecology, 2002, 83(4): 1105 − 1117. [8] 董晨, 夏凯. 基于Apriori算法的浙西杉木用材林立地及生长因子关联分析[J]. 浙江农林大学学报, 2019, 36(4): 741 − 748. DONG Chen, XIA Kai. Chinese fir forest site association analysis and growth factors in western Zhejiang based on the Apriori algorithm [J]. J Zhejiang A&F Univ, 2019, 36(4): 741 − 748. [9] SFENTHOURAKIS S, TZANATOS E, GIOKAS S. Species co-occurrence: the case of congeneric species and a causal approach to patterns of species association [J]. Global Ecol Biogeogr, 2006, 15(1): 39 − 49. [10] 邓贤兰, 刘玉成, 吴杨. 井冈山自然保护区栲属群落优势种群的种间联结关系研究[J]. 植物生态学报, 2003, 27(4): 531 − 536. DENG Xianlan, LIU Yucheng, WU Yang. Interconnection among dominant plant populations of Castanopsis community in Jinggang Mountain Nature Reserve [J]. Acta Phytoecol Sin, 2003, 27(4): 531 − 536. [11] LIU Liting, WANG Xiaodong, WEN Qiang, et al. Interspecific associations of plant populations in rare earth mining wasteland in southern China [J]. Int Biodeterior Biodegradation, 2017, 118: 82 − 88. [12] 钟娇娇, 陈杰, 陈倩, 等. 秦岭山地天然次生林群落MRT数量分类、CCA排序及多样性垂直格局[J]. 生态学报, 2019, 39(1): 277 − 285. ZHONG Jiaojiao, CHEN Jie, CHEN Qian, et al. Quantitative classification of MRT, CCA ordination, and species diversity along elevation gradients of a natural secondary forest in the Qinling mountains [J]. Acta Ecol Sin, 2019, 39(1): 277 − 285. [13] 朱彦鹏, 梁军, 孙志强, 等. 昆嵛山森林群落数量分类、排序及多样性垂直格局[J]. 林业科学, 2013, 49(4): 54 − 61. ZHU Yanpeng, LIANG Jun, SUN Zhiqiang, et al. Numerical classification, ordiantion and species diversity along elevation gradients of the forest community in Kunyu Moutain [J]. Sci Silv Sin, 2013, 49(4): 54 − 61. [14] MONNAHAN C C, THORSON J T, BRANCH T A. Faster estimation of Bayesian models in ecology using Hamiltonian Monte Carlo [J]. Methods Ecol Evol, 2017, 8(3): 339 − 348. [15] 潘永柱, 阙建勇, 叶连宝. 松阳县生物防火林带布设初探[J]. 浙江林业科技, 2012, 32(3): 50 − 54. PAN Yongzhu, QUE Jianyong, YE Lianbao. Layout biological fire belt in Songyang County [J]. J Zhejiang For Sci Technol, 2012, 32(3): 50 − 54. [16] 毛朝明, 蒋灵华, 钱龙福. 森林资源二类调查小班区划方法探讨:以浙江省松阳县为例[J]. 华东森林经理, 2013, 27(3): 25 − 26. MAO Chaoming, JIANG Linghua, QIAN Longfu. Discussion on subdivision method of second-class forest resources investigation: a case study of Songyang County, Zhejiang Province [J]. East China For Manage, 2013, 27(3): 25 − 26. [17] 占拥法, 洪枢勇, 吕律英. 松阳县森林资源特点分析及可持续发展对策[J]. 中国林业经济, 2016(2): 83 − 84. ZHAN Yongfa, HONG Shuyong, LÜ Lüying. Characteristics analysis and sustainable development of forest resources in Songyang County [J]. China For Econ, 2016(2): 83 − 84. [18] 孙杰杰, 沈爱华, 黄玉洁, 等. 浙江省大叶榉树生境地群落数量分类与排序[J]. 南京林业大学学报(自然科学版), 2019, 43(4): 85 − 93. SUN Jiejie, SHEN Aihua, HUANG Yujie, et al. Quantitative classification and ordination of Zelkova schneideriana habitat in Zhejiang Province [J]. J Nanjing For Univ Nat Sci Ed, 2019, 43(4): 85 − 93. [19] 孙杰杰. 浙江檫木群落生境特征及区划研究[D]. 杭州: 浙江农林大学, 2019. SUN Jiejie. Study on Habitat Characterization and Regionalization of Sassafras tzumu (Hemsl.) Hemsl. Community in Zhejiang[D]. Hangzhou: Zhejiang A&F University, 2019. [20] BORCARD D, GILLET F, LEGENDRE P. Numerical Ecology with R[M]. Montreal: Springer, 2018. [21] Ter BRAAK C J F. The analysis of vegetation-environment relationships by canocical correspondence analysis [J]. Vegetatio, 1987, 69(1/3): 69 − 77. [22] 方精云, 王襄平, 沈泽昊, 等. 植物群落清查的主要内容、方法和技术规范[J]. 生物多样性, 2009, 17(6): 533 − 548. FANG Jingyun, WANG Xiangping, SHEN Zehao, et al. Methods and protools for plant community inventory [J]. Biodiversity Sci, 2009, 17(6): 533 − 548. [23] WALLERIUS M L, NÄSLUND J, KOECK B, et al. Interspecific association of brown trout (Salmo trutta) with non-native brook trout (Salvelinus fontinalis) at the fry stage [J]. Ethology, 2017, 123(12): 933 − 941. [24] 郭屹立, 卢训令, 丁圣彦. 伊洛河河岸带生态系统草本植物功能群划分[J]. 生态学报, 2012, 32(14): 4434 − 4442. GUO Yili, LU Xunling, DING Shengyan. The classification of plant functional types based on the dominant herbaceous species in the riparian zone ecosystems in the Yiluo River [J]. Acta Ecol Sin, 2012, 32(14): 4434 − 4442. [25] HURLBERT S H. A coefficient of interspecific assciation [J]. Ecology, 1969, 50(1): 1 − 9. [26] WHITTAKER R H, FAIRBANKS C W. A study of plankton copepod communities in the Columbia Basin, Southeastern Washington [J]. Ecology, 1958, 39(1): 46 − 65. [27] GRIFFITH D M, VEECH J A, MARSH C J. Cooccur: probabilistic species co-occurrence analysis in R [J]. J Stat Software, 2016, 69(2): 1 − 17. [28] 苏日古嘎, 张金屯, 张斌, 等. 松山自然保护区森林群落的数量分类和排序[J]. 生态学报, 2010, 30(10): 2621 − 2629. Suriguga, ZHANG Jintun, ZHANG Bin, et al. Numerical classification and ordination of forest communities in the Songshan National Nature Reserve [J]. Acta Ecol Sin, 2010, 30(10): 2621 − 2629. [29] 刘梦婷, 王振锡, 王雅佩, 等. 新疆天山云杉林群落分布格局及环境解释[J]. 林业科学研究, 2019, 32(6): 90 − 98. LIU Mengting, WANG Zhenxi, WANG Yapei, et al. Plant communities pattern of Picea tianschanica forest and their interrelations with environmental factors in Tianshan Area [J]. For Res, 2019, 32(6): 90 − 98. [30] SIEFERT A, RAVENSCROFT C, ALTHOFF D, et al. Scale dependence of vegetation-environment relationships: a meta-analysis of multivariate data [J]. J Veg Sci, 2012, 23(5): 942 − 951. [31] 陈云, 王海亮, 韩军旺, 等. 小秦岭森林群落数量分类、排序及多样性垂直格局[J]. 生态学报, 2014, 34(8): 2068 − 2075. CHEN Yun, WANG Hailiang, HAN Junwang, et al. Numerical classification, ordination and species diversity along elevation gradients of the forest community in Xiaoqinling [J]. Acta Ecol Sin, 2014, 34(8): 2068 − 2075. [32] 叶诺楠, 沈娜娉, 商天其, 等. 浙江瑞安公益林群落结构及其与环境的相关性[J]. 植物学报, 2017, 52(4): 496 − 510. YE Nuonan, SHEN Napin, SHANG Tianqi, et al. Vegetation structure and internal relationship between distribution patterns of vegetation and environment in ecology service forest of Rui’an City in Zhejiang Province [J]. Chin Bull Bot, 2017, 52(4): 496 − 510. [33] 刘烁, 周国模, 白尚斌. 基于光照强度变化的毛竹扩张对杉木影响的探讨[J]. 浙江农林大学学报, 2011, 28(4): 550 − 554. LIU Shuo, ZHOU Guomo, BAI Shangbin. Light intensity changes on Cunninghamia lanceolata in mixed stands with different concentrations of Phyllostachys pubescens [J]. J Zhejiang A&F Univ, 2011, 28(4): 550 − 554. [34] 宋庆妮. 毛竹扩张对常绿阔叶林氮磷分配格局与过程的影响[D]. 北京: 清华大学, 2017. SONG Qingni. The Effects for Phyllostachys pubscens Expansion on Nitrogen and Phosphorus Distribution Pattern and Process of Evergreen Broadleaved Forests[D]. Beijing: Tsinghua University, 2010. [35] 沈国春. 生境异质性和扩散限制对亚热带和热带森林物种多样性维持的作用[D]. 杭州: 浙江大学, 2010. SHEN Guochun. The Effects of Habitat Heterogeneity and Dispersal Limitation on Species Diversity in Subtropical and Tropical Forests[D]. Hangzhou: Zhejiang University, 2010. [36] 孙杰杰, 江波, 吴初平, 等. 浙江省檫木林生境与生态位研究[J]. 生态学报, 2019, 39(3): 131 − 141. SUN Jiejie, JIANG Bo, WU Chuping, et al. Study on the habitat and niche of Sassafras tzumu (Hemsl.) Hemsl. in Zhejiang Province [J]. Acta Ecol Sin, 2019, 39(3): 131 − 141. [37] 许金石, 陈煜, 王国勋, 等. 陕北桥山林区主要木本植物群落种间联结性[J]. 西北植物学报, 2014, 34(7): 1467 − 1475. XU Jinshi, CHEN Yu, WANG Guoxun, et al. Interspecific association of dominant woody plant communities in Qiaoshan Forest Region, Shaanxi [J]. Acta Bot Boreali-Occident Sin, 2014, 34(7): 1467 − 1475. -

-
链接本文:
https://zlxb.zafu.edu.cn/article/doi/10.11833/j.issn.2095-0756.20190514
 点击查看大图
点击查看大图
计量
- 文章访问数: 2482
- HTML全文浏览量: 580
- PDF下载量: 62
- 被引次数: 0
