





-
陆地总初级生产力(GPP)是指在单位时间和空间上,绿色植物通过光合作用所固定的有机物总量。GPP不仅能够反映植被群落的生产能力,还是客观评价生态系统功能状况的重要指标[1]。光能利用率模型可以利用植被吸收的光合有效辐射(APAR)和光能利用率估算GPP,已经成为估算区域和全球尺度GPP的主流方法,包括CASA、MOD17、GLO-PEM、EC-LUE、TL-LUE、VPM等模型,其中VPM模型因其结构简单,计算效率高,精度好等特点而被广泛运用[2-3]。增强型植被指数(EVI)和陆表水指数(LSWI)是VPM模型的主要驱动因子,在估计GPP中起到重要的作用。EVI已被广泛用于指示植被覆盖状况,是估计GPP较好的参数[4-5]。刘丹等[6]认为:LSWI对土壤相对湿度很敏感。然而,叶昊天[7]研究发现:LSWI并不能很好地反映生态系统土壤含水量的大小,因此其准确性还有待考证。蒸散量是地球水循环的关键驱动因子,主要与温度和降水有关,也与叶面积有关,是影响干旱的重要因素[8-9]。干旱不仅会影响植被的形态特征和代谢系统,还会减少光合有效面积,影响植被GPP的大小[10-11]。因此,本研究利用MODIS及通量塔数据,对GPP的驱动因子进行了分析,利用潜在蒸散量(PET)和降水量(P)对站点进行干湿划分,探究LSWI参数的准确性及在干旱影响下不同光能利用率模型模拟GPP的能力,对准确模拟北半球常绿针叶林GPP提供科学依据。
-
研究对象为北半球常绿针叶林(ENF)的23个站点,时间从2001−2014年,纬度为38°53′43″~67°21′43″N(表1)。站点包括温带大陆性气候、高原山地气候、地中海气候等多种干湿类型的气候带。
表 1 北半球常绿针叶林通量塔站点分布情况
Table 1. Distribution of flux tower sites of evergreen needleleaved forests in Northern Hemisphere
站点名称 纬度 经度 海拔/m 站点名称 纬度 经度 海拔/m CA-Man 55°52′47″N 98°28′51″W 259 CZ-BK1 49°30′08″N 18°32′13″E 875 CA-Obs 53°59′14″N 105°07′04″W 629 DE-Obe 50°47′01″N 13°43′11″E 735 CA-Qfo 49°41′33″N 74°20′32″W 382 DE-Tha 50°57′49″N 13°34′01″E 380 FR-LBr 44°43′02″N 0°46′09″W 61 FI-Hyy 61°50′51″N 24°17′42″E 181 IT-Ren 46°35′13″N 11°26′01″E 1 730 FI-Sod 67°21′43″N 26°38′16″E 180 IT-SRo 43°43′40″N 10°17′04″E 6 IT-Lav 45°57′22″N 11°16′53″E 1 353 US-Blo 38°53′43″N 120°37′58″W 1 315 NL-Loo 52°10′00″N 5°44′37″E 25 US-GBT 41°21′57″N 106°14′23″W 3 191 RU-Fyo 56°27′41″N 32°55′20″E 265 CA-TP1 42°39′39″N 80°33′34″W 265 US-GLE 41°21′59″N 106°14′24″W 3 197 CA-TP3 42°42′24″N 80°20′54″W 184 US-Me2 44°27′08″N 121°33′27″W 1 253 CA-TP4 42°42′37″N 80°21′27″W 184 US-NR1 40°01′58″N 105°32′47″W 3 050 CH-Dav 46°48′55″N 9°51′21″E 1 639 -
MODIS数据来源于美国国家航空航天局(NASA)Terra卫星传感器。数据通过输入通量塔站点的经纬度下载(
https://modis.ornl.gov/globalsubset/ ),包括2001−2014年的光合有效辐射吸收比(MOD15A2H)、潜在蒸散量(MOD16A2)和地表反射率(MOD09A1),时间步长为8 d,空间分辨率为500 m。 -
涡流协方差测量数据采集自FLUXNET 2015数据集,该数据集可从FLUXNET通量观测网站下载,包括2001−2014年的GPP、温度、降水、短波辐射数据集。时间步长为每天。
-
为了与模型中遥感参数步长相匹配,对通量塔数据进行8 d的累加。建立通量塔数据与遥感数据相匹配的2001−2014年时间序列,删除遥感数据中的空缺值以及排除平均值的3倍标准差之外的异常值。本研究的陆表水指数(ILSW)、增强型植被指数(IEV)、光合有效辐射吸收比(FPAR)计算如下:
$$ {I}_{\mathrm{L}\mathrm{S}\mathrm{W}}=({{r}_{\mathrm{n}\mathrm{i}\mathrm{r}}-{r}_{\mathrm{d}}})/({{r}_{\mathrm{n}\mathrm{i}\mathrm{r}}+{r}_{\mathrm{d}}})\text{;} $$ (1) $$ {I}_{\mathrm{E}\mathrm{V}}=[{2.5 ({r}_{\mathrm{n}\mathrm{i}\mathrm{r}}-{r}_{\mathrm{R}})}]/{({r}_{\mathrm{n}\mathrm{i}\mathrm{r}}+6 {r}_{\mathrm{R}}-7.5 {r}_{\mathrm{b}}+1)}\text{;} $$ (2) $$ {F}_{\mathrm{P}\mathrm{A}\mathrm{R}}={I}_{\mathrm{E}\mathrm{V}}{\text{。}} $$ (3) 式(1)~(3)中:rnir为近红外波段的反射值;rd为短波红外波段的反射值;rR为红光波段的反射值;rb为蓝光波段的反射值。
-
不同水热条件下植被的生长状况有明显差异,在研究GPP时,有必要针对具体的气候区域进行研究[12]。降水和蒸散分别是植被获取和失去水分的主要途径,用这2个变量构造干湿度指数,可以反映站点的水分情况。其中降水数据为实际观测值,而遥感数据潜在蒸散量与温度、光照和GPP具有很高的相关性,因此用于构造干湿度指数(ID)具有合理性,即1 a降水量和潜在蒸散量累加和的比值,其计算如下:
$$ {I}_{\mathrm{D}}=\displaystyle \sum\limits_{i = 1}^n {P_i}\left/{\displaystyle \sum\limits_{i = 1}^n {P}_{\mathrm{e}\mathrm{t}i}{\text{。}}}\right. $$ (4) 式(4)中:Pi为8 d的降水量(mm),
$ {P}_{\mathrm{e}\mathrm{t}i} $ 为8 d的潜在蒸散量(mm),i为站点数据的条号,i=1,2,3$,\cdots, $ n,n为站点数据的总条数。根据干湿度指数将站点分为[13]:干旱地区(ID为0.25~0.50)、半干旱地区(ID为0.50~0.67)、半湿润地区(ID为0.67~1.00)和湿润地区(ID为1.00~2.00)。 -
VPM模型是利用温度、水分等环境因子以及涡度观测碳通量数据,并且考虑了植被叶绿素吸收的光合有效辐射来估计GPP的1种光能利用率模型。VPM模型的一般形式有:
$${G_{{\rm{PP}}}} = \varepsilon {F_{\rm{T}}} {F_{\rm{W}}} {F_{\rm{P}}} {F_{{\rm{PAR}}}} {P_{{\rm{AR}}}};$$ (5) $${P_{{\rm{AR}}}} = 0.45 {S_{{\rm{Wrad}}}}_{};$$ (6) $${F_{\rm{T}}} = \left[{{\left( {T - {T_{{\rm{min}}}}} \right) \left( {T - {T_{{\rm{max}}}}} \right)}}]/[{{\left( {T - {T_{{\rm{min}}}}} \right) \left( {T - {T_{{\rm{max}}}}} \right) - {{( {T - {T_{{\rm{opt}}}}} )}^2}}}\right];$$ (7) $${F_{\rm{W}}} = ({{1 + {I_{{\rm{LSW}}}}}})/({{1 + {I_{{\rm{LSWmax}}}}}}){\text{。}}$$ (8) 式(5)~(8)中:
$ {G}_{\mathrm{P}\mathrm{P}} $ 表示总初级生产力;$ \varepsilon $ 表示最大的光能利用效率;$ {F}_{\mathrm{T}} $ 、$ {F}_{\mathrm{W}} $ 和$ {F}_{\mathrm{P}} $ 分别表示模拟温度、水、物候对GPP的影响,本研究FP取值为1[14];$ {F}_{\mathrm{P}\mathrm{A}\mathrm{R}} $ 表示植被吸收的光合有效辐射的比例;$ {P}_{\mathrm{A}\mathrm{R}} $ 表示光合有效辐射;$ {S}_{\mathrm{W}\mathrm{r}\mathrm{a}\mathrm{d}} $ 表示站点短波辐射;T表示温度;Topt表示植被生长所需温度最优值;Tmax表示温度最大值;Tmin表示温度最小值;当T小于Tmin时,$ {F}_{\mathrm{T}} $ 等于0,当T大于Tmax时,$ {F}_{\mathrm{T}} $ 等于1。在拟合时Tmin取值−5 ℃,Tmax取值40 ℃,Topt通常取值20 ℃[15],但在本研究中,$ \varepsilon $ 和Topt通过非线性拟合得到[16]。$ {I}_{\mathrm{L}\mathrm{S}\mathrm{W}} $ 为陆表水指数,$ {I}_{\mathrm{L}\mathrm{S}\mathrm{W}\mathrm{m}\mathrm{a}\mathrm{x}} $ 为生长季最大陆表水指数。 -
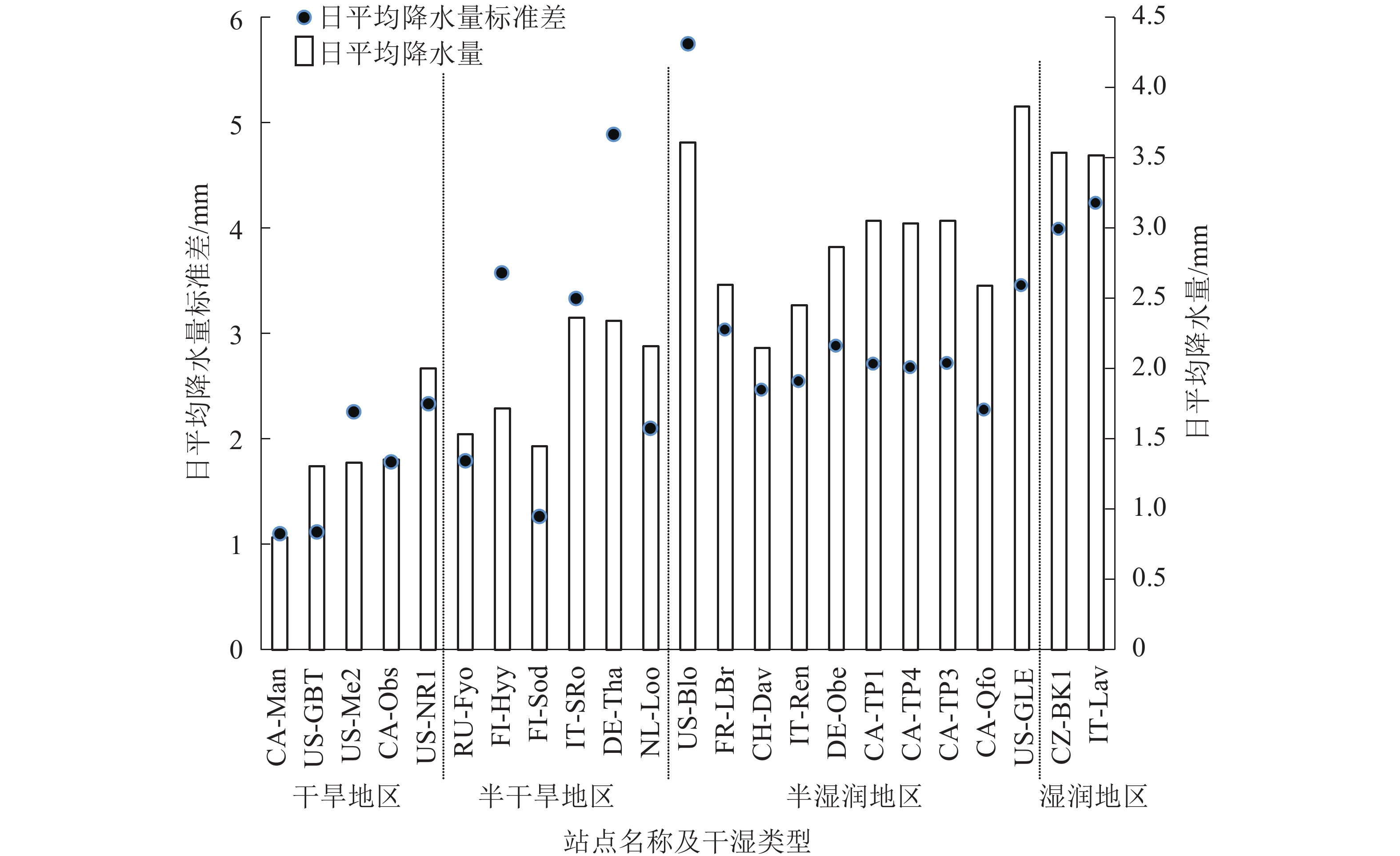
LSWI由MODIS遥感数据中的地表反射率计算得到,但由于常绿针叶林林冠层绿度季节变动性较弱,因此遥感信息提取存在较大的不确定性[17]。在不同干湿类型的23个站点上,分别作出各站点2001−2014年的年平均LSWI与年尺度上的日平均降水量(图1)。LSWI、日平均降水量分别由遥感数据和站点观测数据得到,发现LSWI在干旱和半干旱地区的数值高于半湿润地区,而降水量从干旱区到湿润区依次递增,两者相矛盾,说明LSWI可能在干旱和半干旱地区不能反映常绿针叶林的水分情况。为了提高干旱和半干旱地区GPP的估算精度,将VPM模型中的模拟水(FW)因子去除,生成改进的VPMsw模型,计算如下:

图 1 不同干湿类型站点陆表水指数和日平均降水量分布
Figure 1. Dstribution of land surface water index and daily mean precipitation at different dry and wet stations
$${G_{{\rm{PP}}}} = \varepsilon {F_{\rm{T}}} {F_{\rm{P}}} {F_{{\rm{PAR}}}} {P_{{\rm{AR}}}}{\text{。}}$$ (9) -
通过皮尔逊相关性分析,分析温度、陆表水指数、光合有效辐射和潜在蒸散量对GPP影响的重要性。本研究使用随机森林算法分析温度、光照和水分对GPP的重要性,通过打乱其中1个因子的取值造成随机森林前后结果的错误率,错误率越高说明该因子越重要[18]。该方法对异常值具有较好的容忍度,并且不容易出现过拟合现象。
-
采用均方根误差和决定系数来检验模型的精度,均方根误差越小,偏离程度越小,表现力越好。决定系数越大,解释程度越高,效果越好[19]。
-
由皮尔逊相关性分析可知:GPP与温度、潜在蒸散量和光合有效辐射相关系数较高,而与陆表水指数相关系数较低(表2)。
表 2 各因子之间皮尔逊相关性
Table 2. Pearson correlation among the factors
因子 陆表水指数 光合有效辐射 温度 潜在蒸散量 GPP 陆表水指数 1 −0.31** −0.45** −0.36** −0.28** 光合有效辐射 −0.31** 1 0.68** 0.86** 0.60** 温度 −0.45** 0.68** 1 0.74** 0.74** 潜在蒸散量 −0.36** 0.86** 0.74** 1 0.66** GPP −0.28** 0.60** 0.74** 0.66** 1 说明:**表示极显著相关(P<0.01) 随机森林分析结果表明:在23个站点中,17个站点以温度为主导因子,重要性达70%以上;5个站点以光照为主导因子,重要性超过50%;仅1个站点(US-Blo)是以水分为主导因子的,重要性达60%。
-
对所有站点区分干湿类型后,使用VPM和VPMsw模型模拟GPP,计算均方根误差和决定系数(R2),结果见表3。在干旱和半干旱地区,VPMsw模型的R2比VPM模型高,但均方根误差比VPM模型低。VPMsw模型在干旱站点上均方根误差平均减少了6.5%,在半干旱站点上均方根误差平均减少了23.4%,在半干旱地区精度提高的效果较好;而在半湿润和湿润地区,VPM模型的精度比VPMsw模型高。基于VPMsw模型在干旱和半干旱地区较好的模拟效果,将干旱和半干旱站点所有数据实测值与估计值进行线性拟合(图2)发现:相比VPM模型,VPMsw模型在干旱地区模拟GPP的精度高于半干旱地区。
表 3 站点干湿度指数及模型均方根误差对比
Table 3. Comparison of dry humidity index and model root mean square error
干湿
类型站点 干湿度
指数均方根误差 决定系数 干湿
类型站点 干湿度
指数均方根误差 决定系数 VPMsw
模型VPM
模型VPMsw
模型VPM
模型VPMsw
模型VPM
模型VPMsw
模型VPM
模型干旱 CA-Man 0.28 1.28 1.58 0.71 0.77 半湿润 FR-LBr 0.73 2.27 2.15 0.49 0.47 US-GBT 0.29 1.29 1.70 0.83 0.66 CH-Dav 0.76 2.17 2.38 0.46 0.47 US-Me2 0.35 1.91 1.96 0.51 0.51 IT-Ren 0.76 2.09 1.85 0.63 0.72 CA-Obs 0.41 1.23 1.44 0.78 0.68 DE-Obe 0.79 3.69 3.53 0.62 0.70 US-NR1 0.47 1.58 1.72 0.77 0.72 CA-TP1 0.80 3.84 3.73 0.31 0.39 CA-TP4 0.81 2.46 2.20 0.6 0.66 半干旱 RU-Fyo 0.50 2.48 2.77 0.65 0.53 CA-TP3 0.82 2.46 2.15 0.61 0.69 FI-Hyy 0.56 2.17 2.53 0.69 0.49 CA-Qfo 0.94 1.39 1.28 0.61 0.71 FI-Sod 0.62 1.55 1.90 0.63 0.38 US-GLE 0.95 1.39 1.24 0.71 0.76 IT-SRo 0.62 3.19 3.00 0.36 0.36 DE-Tha 0.66 2.93 3.03 0.67 0.57 湿润 CZ-BK1 1.00 3.23 3.28 0.59 0.49 NL-Loo 0.67 2.49 2.72 0.71 0.61 IT-Lav 1.04 3.59 3.48 0.61 0.6 半湿润 US-Blo 0.72 1.75 2.48 0.42 0.49 
图 2 干旱和半干旱站点GPP拟合精度对比
Figure 2. Comparison of fitting accuracy of GPP between arid and semi-arid sites
-
从图3可见:从干旱地区到湿润地区,日平均降水量总体呈逐渐增大趋势,但也存在部分站点降水量突增的情况,如US-NR1、IT-SRo、DE-Tha、US-Blo和US-GLE站点。而日平均降水量标准差也从干旱地区到湿润地区呈逐渐增大趋势,其中US-Me2、FI-Hyy、IT-SRo、DE-Tha和US-Blo站点的日平均降水量标准差异常大,且这些站点的日平均降水量都处于所在干湿区降水量的较高水平。日平均降水量标准差异常大的站点在干旱地区有4个,湿润地区仅1个。
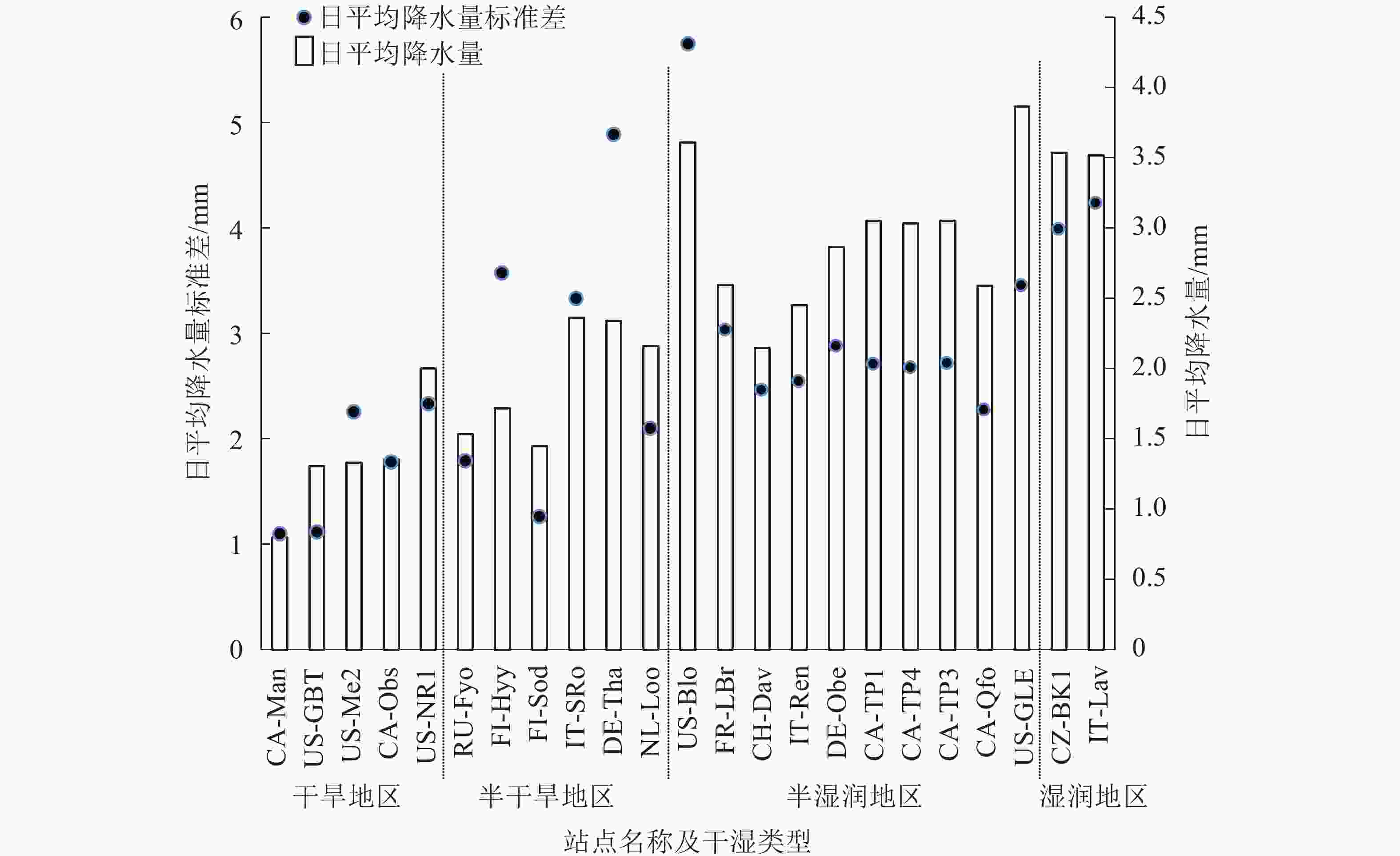
图 3 每个站点的日平均降水量及其标准差
Figure 3. Average daily precipitation and standard deviation in each site
-
常绿针叶林GPP的主要驱动因子是温度、潜在蒸散量,LSWI与GPP的相关性最低。干旱和半干旱地区,LSWI不能较好地反映常绿针叶林的水分情况,所以在干旱和半干旱地区,VPMsw模型拟合GPP的精度较高。
-
温度与GPP的相关性最高,这与贺忠华等[20]的研究结果一致。温度的适量增加有助于延长植被生长季的长度,对GPP增加效果明显,是影响植被GPP的主要驱动因子[21-22]。
水分与GPP的相关性最低。由表3可知:在不同干湿类型地区,LSWI对GPP估计的影响从大到小依次为干旱区、半干旱地区、半湿润、湿润地区。也有研究表明:LSWI不能反映近岸地区潮汐作用对植被光合作用的影响,导致VPM模型的拟合效果变差[23]。
本研究发现:温度为主导因子的站点最多且重要性最大,认为温度的影响力大于水分。温度是植被生长的主要驱动因子,水分只是植被生长的限制因子。在干旱和半干旱地区常年降水量较低,降水主要来自于夏季。从干旱和半干旱部分站点过高的日降水量标准差可知:这些站点受到高温胁迫导致夏季降水的天数减少,而半湿润和湿润地区日降水量较多,能有效缓解高温对植被造成的生产力下降的负面影响[24],其降水量标准差大是由于日降水量分布范围大。在干旱和半干旱地区植被更易受到高温和缺水的影响,因此植被在干旱和半干旱地区更易受到水分的限制作用。其他研究也表明:高温和缺水会导致VPM模型拟合的GPP不准确[25],这是因为遥感数据不能准确地反映水分的变化。由于温度变化比较稳定,且遥感数据获取的温度比较准确,因此为了更准确地使用VPM模型拟合GPP,在干旱和半干旱地区需要更准确的遥感数据来反映水分因子。
本研究发现:LSWI参数在半干旱和干旱的常绿针叶林区域不能很好地表现水分情况,从而影响了VPM模型拟合GPP的准确性。本研究构造的干湿度指数可在长时间尺度上区分站点的水分情况;对干湿度指数进行变换处理,使之成为可直接带入模型的水分参数还有待研究。未来在干旱和半干旱地区找到准确反映水分状况的遥感数据是提高模型拟合GPP的关键。
A comparison of different radiation use efficiency models in gross primary production accuracy under drought
-
摘要:
目的 研究干旱影响下不同光能利用率模型估算常绿针叶林总初级生产力(GPP)的能力,为精准模拟北半球常绿针叶林的GPP提供参考依据。 方法 采用MODIS数据和通量站点观测数据,在8 d尺度上对常绿针叶林的GPP进行参数化建模。首先利用皮尔逊相关系数和随机森林因子评价方法分析GPP各个驱动因子的重要性。使用降水量和潜在蒸散量(PET)构造干湿度指数,对各个站点进行干湿分类。其次在植被光合模型(VPM)的基础上,去除其中的水分参数,得到改进的植被光合模型(VPMsw)。最后定量比较各站点在不同干湿类型下,VPM模型与VPMsw模型拟合GPP的精度。 结果 ①由驱动因子分析可知:GPP与温度、PET相关性较强,与陆表水指数(LSWI)相关性较弱,温度对GPP的重要性大于水分。②与VPM模型相比,VPMsw模型在干旱和半干旱站点拟合GPP的精度有所提高,均方根误差分别减少6.5%和23.4%,在半干旱地区精度提升效果更明显。 结论 干旱影响下,LSWI不能较好地反映干旱和半干旱地区常绿针叶林的水分情况,所以在干旱和半干旱地区,VPMsw模型拟合GPP的精度更高。图3表3参25 Abstract:Objective With an investigation of different radiation use efficiency models estimating the gross primary production (GPP) of evergreen needle-leaved forest (ENF) under the influence of drought, this paper is aimed to provide basis for accurately simulating GPP of ENF in Northern Hemisphere. Method MODIS data and flux site observation data at 8-day scale were used to simulate the GPP of ENF. Firstly, Pearson correlation coefficient and random forest factor evaluation method were used to analyze the correlation and importance of each driving factor of GPP while precipitation and potential evapotranspiration (PET) were used to calculate the dry-wet index to classify the dry-wet type of each site. Secondly, the VPMsw model was constructed by deleting the moisture parameter in the vegetation photosynthesis model (VPM). Finally, the accuracy of GPP estimated from VPM model and VPMsw model were quantitatively compared under different dry and wet types at each site. Result (1) GPP had strong correlation with temperature and PET, but weak correlation with Land Surface Water Index (LSWI). (2) compared with the VPM model, the accuracy of VPMsw model in fitting GPP was improved in arid and semi-arid sites, and the root mean square error (RMSE) decreased by 6.5% and 23.4%, respectively. the accuracy of GPP estimates in semi-arid sites was significantly improved by the VPMsw model. Conclusion VPMsw model was more accurate in simulating GPP in arid and semi-arid areas because LSWI could not well reflect the water content of the ENF in arid and semi-arid areas under the influence of drought. [Ch, 3 fig. 3 tab. 25 ref.] -
图 1 不同干湿类型站点陆表水指数和日平均降水量分布
Figure 1 Dstribution of land surface water index and daily mean precipitation at different dry and wet stations
图 2 干旱和半干旱站点GPP拟合精度对比
Figure 2 Comparison of fitting accuracy of GPP between arid and semi-arid sites
图 3 每个站点的日平均降水量及其标准差
Figure 3 Average daily precipitation and standard deviation in each site
表 1 北半球常绿针叶林通量塔站点分布情况
Table 1. Distribution of flux tower sites of evergreen needleleaved forests in Northern Hemisphere
站点名称 纬度 经度 海拔/m 站点名称 纬度 经度 海拔/m CA-Man 55°52′47″N 98°28′51″W 259 CZ-BK1 49°30′08″N 18°32′13″E 875 CA-Obs 53°59′14″N 105°07′04″W 629 DE-Obe 50°47′01″N 13°43′11″E 735 CA-Qfo 49°41′33″N 74°20′32″W 382 DE-Tha 50°57′49″N 13°34′01″E 380 FR-LBr 44°43′02″N 0°46′09″W 61 FI-Hyy 61°50′51″N 24°17′42″E 181 IT-Ren 46°35′13″N 11°26′01″E 1 730 FI-Sod 67°21′43″N 26°38′16″E 180 IT-SRo 43°43′40″N 10°17′04″E 6 IT-Lav 45°57′22″N 11°16′53″E 1 353 US-Blo 38°53′43″N 120°37′58″W 1 315 NL-Loo 52°10′00″N 5°44′37″E 25 US-GBT 41°21′57″N 106°14′23″W 3 191 RU-Fyo 56°27′41″N 32°55′20″E 265 CA-TP1 42°39′39″N 80°33′34″W 265 US-GLE 41°21′59″N 106°14′24″W 3 197 CA-TP3 42°42′24″N 80°20′54″W 184 US-Me2 44°27′08″N 121°33′27″W 1 253 CA-TP4 42°42′37″N 80°21′27″W 184 US-NR1 40°01′58″N 105°32′47″W 3 050 CH-Dav 46°48′55″N 9°51′21″E 1 639  下载: 导出CSV
下载: 导出CSV
表 2 各因子之间皮尔逊相关性
Table 2. Pearson correlation among the factors
因子 陆表水指数 光合有效辐射 温度 潜在蒸散量 GPP 陆表水指数 1 −0.31** −0.45** −0.36** −0.28** 光合有效辐射 −0.31** 1 0.68** 0.86** 0.60** 温度 −0.45** 0.68** 1 0.74** 0.74** 潜在蒸散量 −0.36** 0.86** 0.74** 1 0.66** GPP −0.28** 0.60** 0.74** 0.66** 1 说明:**表示极显著相关(P<0.01)
下载: 导出CSV
表 3 站点干湿度指数及模型均方根误差对比
Table 3. Comparison of dry humidity index and model root mean square error
干湿
类型站点 干湿度
指数均方根误差 决定系数 干湿
类型站点 干湿度
指数均方根误差 决定系数 VPMsw
模型VPM
模型VPMsw
模型VPM
模型VPMsw
模型VPM
模型VPMsw
模型VPM
模型干旱 CA-Man 0.28 1.28 1.58 0.71 0.77 半湿润 FR-LBr 0.73 2.27 2.15 0.49 0.47 US-GBT 0.29 1.29 1.70 0.83 0.66 CH-Dav 0.76 2.17 2.38 0.46 0.47 US-Me2 0.35 1.91 1.96 0.51 0.51 IT-Ren 0.76 2.09 1.85 0.63 0.72 CA-Obs 0.41 1.23 1.44 0.78 0.68 DE-Obe 0.79 3.69 3.53 0.62 0.70 US-NR1 0.47 1.58 1.72 0.77 0.72 CA-TP1 0.80 3.84 3.73 0.31 0.39 CA-TP4 0.81 2.46 2.20 0.6 0.66 半干旱 RU-Fyo 0.50 2.48 2.77 0.65 0.53 CA-TP3 0.82 2.46 2.15 0.61 0.69 FI-Hyy 0.56 2.17 2.53 0.69 0.49 CA-Qfo 0.94 1.39 1.28 0.61 0.71 FI-Sod 0.62 1.55 1.90 0.63 0.38 US-GLE 0.95 1.39 1.24 0.71 0.76 IT-SRo 0.62 3.19 3.00 0.36 0.36 DE-Tha 0.66 2.93 3.03 0.67 0.57 湿润 CZ-BK1 1.00 3.23 3.28 0.59 0.49 NL-Loo 0.67 2.49 2.72 0.71 0.61 IT-Lav 1.04 3.59 3.48 0.61 0.6 半湿润 US-Blo 0.72 1.75 2.48 0.42 0.49
下载: 导出CSV
-
[1] 张方敏, 居为民, 陈镜明, 等. 基于遥感和过程模型的亚洲东部陆地生态系统初级生产力分布特征[J]. 应用生态学报, 2012, 23(2): 307 − 318. ZHANG Fangmin, JU Weimin, CHEN Jingming, et al. Characteristics of terrestrial ecosystem primary productivity in East Asia based on remote sensing and process-based model [J]. Chin J Appl Ecol, 2012, 23(2): 307 − 318. [2] 陈静清, 闫慧敏, 王绍强, 等. 中国陆地生态系统总初级生产力VPM遥感模型估算[J]. 第四纪研究, 2014, 34(4): 732 − 742. CHEN Jingqing, YAN Huimin, WANG Shaoqiang, et al. Estimation of gross primary productivity in Chinese terrestrial ecosystems by using VPM model [J]. Quaternary Sci, 2014, 34(4): 732 − 742. [3] WANG Zheng, XIAO Xiangming, YAN Xiaodong. Modeling gross primary production of maize cropland and degraded grassland in northeastern China [J]. Agric For Meteorol, 2010, 150(9): 1160 − 1167. [4] 刘世梁, 董玉红, 安南南, 等. 基于增强型植被指数序列和景观格局分析的松嫩平原盐碱地动态——以大安市为例[J]. 应用生态学报, 2014, 25(11): 3263 − 3269. LIU Shiliang, DONG Yuhong, AN Nannan, et al. Dynamics of salinization land based on EVI series data and landscape pattern analysis in Songnen Plain: a case study in Da’an City [J]. Chin J Appl Ecol, 2014, 25(11): 3263 − 3269. [5] NEMANI R R, KEELING C D, HASHIMOTO H, et al. Climate-driven increases in global terrestrial net primary production from 1982 to 1999 [J]. Science, 2003, 300(5625): 1560 − 1563. [6] 刘丹, 冯锐, 于成龙, 等. 基于植被指数的春玉米干旱响应遥感监测[J]. 农业工程学报, 2019, 35(20): 152 − 161. LIU Dan, FENG Rui, YU Chenlong, et al. Remote sensing monitoring of drought response of spring maize based on vegetation indexes [J]. Trans Chin Soc Agric Eng, 2019, 35(20): 152 − 161. [7] 叶昊天. 锡林郭勒半干旱针茅草原夜间生态系统呼吸作用及生态系统呼吸模型研究[D]. 南京: 南京信息工程大学, 2019. YE Haotian. A Study of Nighttime Ecosystem Respiration and Ecosystem Respiration Model in a Semiarid Stipa Grassland of Xilin Gol[D]. Nanjing: Nanjing University of Information Science and Technology, 2019. [8] CUI Mingyue, WANG Junbang, WANG Shaoqiang, et al. Temporal and spatial distribution of evapotranspiration and its influencing factors on Qinghai-Tibet Plateau from 1982 to 2014 [J]. J Resour Ecol, 2019, 10(2): 213 − 224. [9] 宋成刚, 李红琴, 王军邦, 等. 青海海北高寒嵩草草甸系统水分利用效率特征[J]. 干旱区资源与环境, 2017, 31(6): 90 − 96. SONG Chenggang, LI Hongqin, WANG Junbang, et al. Characterizing ecosystem water use efficiency of alpine Kobresia meadow over the northeastern Qinghai-Tibetan Plateau [J]. J Arid Land Resour Environ, 2017, 31(6): 90 − 96. [10] BOVARD B D, CURTIS P S, VOGEL C S, et al. Environmental controls on sap flow in a northern hardwood forest [J]. Tree Physiol, 2005, 25(1): 31 − 38. [11] 汪本福, 王晴芳, 李阳, 等. 干旱胁迫对水稻叶片生理生化特性的影响综述[J]. 湖北农业科学, 2019, 58(23): 5 − 9. WANG Benfu, WANG Qingfang, LI Yang, et al. Review on the effects of drought stress on physiological and biochemical characteristics of rice leaves [J]. Hubei Agric Sci, 2019, 58(23): 5 − 9. [12] 袁文平, 蔡文文, 刘丹, 等. 陆地生态系统植被生产力遥感模型研究进展[J]. 地球科学进展, 2014, 29(5): 541 − 550. YUAN Wenping, CAI Wenwen, LIU Dan, et al. Satellite-based vegetation production models of terrestrial ecosystem: an overview [J]. Adv Earth Sci, 2014, 29(5): 541 − 550. [13] 孔令颖, 扶松林, 韩晓阳, 等. 基于传统干湿指数的省域长历时气象干旱变化特征及其对旱作粮食单产的影响[J]. 水土保持研究, 2020, 27(3): 159 − 167. KONG Lingying, FU Songlin, HAN Xiaoyang, et al. Spatiotemporal variation of long-term meteorological drought and its impacts on grain yield in Shaanxi Province based on the traditional arid-wet index [J]. Res Soil Water Conserv, 2020, 27(3): 159 − 167. [14] 牛忠恩, 闫慧敏, 陈静清, 等. 基于VPM与MOD17产品的中国农田生态系统总初级生产力估算比较[J]. 农业工程学报, 2016, 32(4): 191 − 198. NIU Zhong’en, YAN Huimin, CHEN Jingqing, et al. Comparison of crop gross primary productivity estimated with VPM model and MOD17 product in field ecosystem of China [J]. Trans Chin Soc Agric Eng, 2016, 32(4): 191 − 198. [15] XIAO Xiangming, ZHANG Qingyuan, BRASWELL B, et al. Modeling gross primary production of temperate deciduous broadleaf forest using satellite images and climate data [J]. Remote Sensing Environ, 2004, 91(2): 256 − 270. [16] 刘正佳, 刘纪远, 邵全琴. 不同土地覆盖类型上植被生长的最适温度[J]. 地球信息科学学报, 2014, 16(1): 1 − 7. LIU Zhengjia, LIU Jiyuan, SHAO Quanqin. Optimum temperature of vegetation growth for various land cover types in China [J]. J Geo-inf Sci, 2014, 16(1): 1 − 7. [17] JÖNSSON A M, EKLUNDH L, HELLSTRÖM M. Annual changes in MODIS vegetation indices of Swedish coniferous forests in relation to snow dynamics and tree phenology [J]. Remote Sensing Environ, 2010, 114(11): 2719 − 2730. [18] LIAW A, WIENER M. Classification and regression by randomForest [J]. R News, 2002, 2/3: 18 − 22. [19] 郭含茹, 张茂震, 徐丽华, 等. 基于地理加权回归的区域森林碳储量估计[J]. 浙江农林大学学报, 2015, 32(4): 497 − 508. GUO Hanru, ZHANG Maozhen, XU Lihua, et al. Geographically weighted regression based on estimation of regional forest carbon storage [J]. J Zhejiang A&F Univ, 2015, 32(4): 497 − 508. [20] 贺忠华, 张育慧, 何月, 等. 浙江省近20年植被变化趋势及驱动因子分析[J]. 生态环境学报, 2020, 29(8): 1530 − 1539. HE Zhonghua, ZHANG Yuhui, HE Yue, et al. Trends of vegetation change and driving factor analysis in recent 20 years over Zhejiang Province [J]. Ecol Environ Sci, 2020, 29(8): 1530 − 1539. [21] 陈正华, 麻清源, 王建, 等. 利用CASA模型估算黑河流域净第一性生产力[J]. 自然资源学报, 2008, 23(2): 263 − 273. CHEN Zhenghua, MA Qingyuan, WANG Jian, et al. Estimation of Heihe basin net primary producivity using the CASA model [J]. J Nat Resour, 2008, 23(2): 263 − 273. [22] 杨明兴, 代侦勇, 杜启勇, 等. 遥感GPP模型在亚热带常绿林的应用比较[J]. 测绘地理信息, 2019, 44(1): 69 − 73. YANG Mingxing, DAI Zhenyong, DU Qiyong, et al. Comparison of different GPP models in subtropical evergreen forest sites [J]. J Geomatics, 2019, 44(1): 69 − 73. [23] 郭海强. 长江河口湿地碳通量的地面监测及遥感模拟研究[D]. 上海: 复旦大学, 2010. GUO Haiqiang. Carbon Fluxes over an Estuarine Wetland: In Situ Measurement and Modeling[D]. Shanghai: Fudan University, 2010. [24] NIU Shuli, LI Zhixiong, XIA Jianyang, et al. Climatic warming changes plant photosynthesis and its temperature dependence in a temperate steppe of northern China [J]. Environ Exp Bot, 2007, 63(1): 91 − 101. [25] 伍卫星, 王绍强, 肖向明, 等. 利用MODIS影像和气候数据模拟中国内蒙古温带草原生态系统总初级生产力[J]. 中国科学: 地球科学, 2008, 38(8): 993 − 1004. WU Weixing, WANG Shaoqiang, XIAO Xiangming, et al. Using MODIS images and climate data to simulate the total primary productivity of temperate grassland ecosystem in Inner Mongolia, China [J]. Sci Sin Terrae, 2008, 38(8): 993 − 1004. -

-
链接本文:
https://zlxb.zafu.edu.cn/article/doi/10.11833/j.issn.2095-0756.20200751
 点击查看大图
点击查看大图
计量
- 文章访问数: 1632
- HTML全文浏览量: 378
- PDF下载量: 54
- 被引次数: 0
























