




-
植物的蒸腾作用是其水分利用的主要方式,拉动水分在“土壤—植物—大气”连续体体系中不断循环迁移。蒸腾耗水与植物生命表征直接联系,决定着植物的水分盈缺和灌溉与否[1]。对活立木蒸腾耗水的准确测定,可以为低耗水树种选择、合理密度配置以及城市园林绿化等工作提供理论依据和参考[2]。树木蒸腾耗水产生的水势差会拉动水分通过木质部向上运输进而形成液流,因此树干液流可作为评估树木蒸腾耗水能力的一项重要指标[3-4]。目前,有多种方法可以评估树木蒸腾耗水能力,多数是通过测定树木液流速率来估算蒸腾耗水量和耗水能力。不同树干液流测定方法测量精度不同,在选择树干液流速率测量方法时需要考虑实验研究目的、活立木树种的生理条件和实验研究所处的自然环境等因素。目前液流速率的测量主要有同位素示踪法和热技术法[5] 2类。其中同位素示踪法通过将化学同位素作为示踪剂注射到树木木质部,从而检测树木液流速率;但该方法在野外应用不便,且测定精度较低,有待改进[6]。利用热技术法测定树干液流不受外界环境和树木自身结构影响,安装布置操作相对简易,并且对树木组织结构损伤较小,具有一定的应用优势[7],因此被广泛应用于树干液流测定、液流速率与环境因子的关系研究中[8-10]。如王檬檬等[11]应用热技术法研究了晋西黄土区苹果Malus pumila树液流速率与太阳辐射、大气水份等的关系;温淑红等[12]应用热技术法分析了宁南黄土陵区山桃Amygdalus davidiana树树干液流速率与太阳辐射、温度、风速的关系;杨洁等[13]应用热技术法研究了树干液流时滞效应,并精确估算树木的蒸腾耗水;还有学者[14-15]应用热技术法探究树干木质部径向不同深度的液流速率和不同时间尺度下的液流速率特征等。目前常用的测量树干液流的热技术法主要有热脉冲法(Heat Pluse Velocity Method,HPVM)、热平衡法、热扩散法、热场变形法以及外热比法等,前3种方法在国内应用较多,后2种方法在国外有较为详细的应用描述,但在国内的研究有限。鉴于此,本研究综述了现有的树干液流无损检测方法,阐述这些方法的基本原理、装置布置、应用领域和最新研究案例,对不同热技术方法的测量精度、适用范围、潜在优势以及今后改进方向等方面进行讨论和比较,并对不同研究目标和实验条件下适用的测定方法给出建议,展望各自在液流研究方面的应用前景。
-
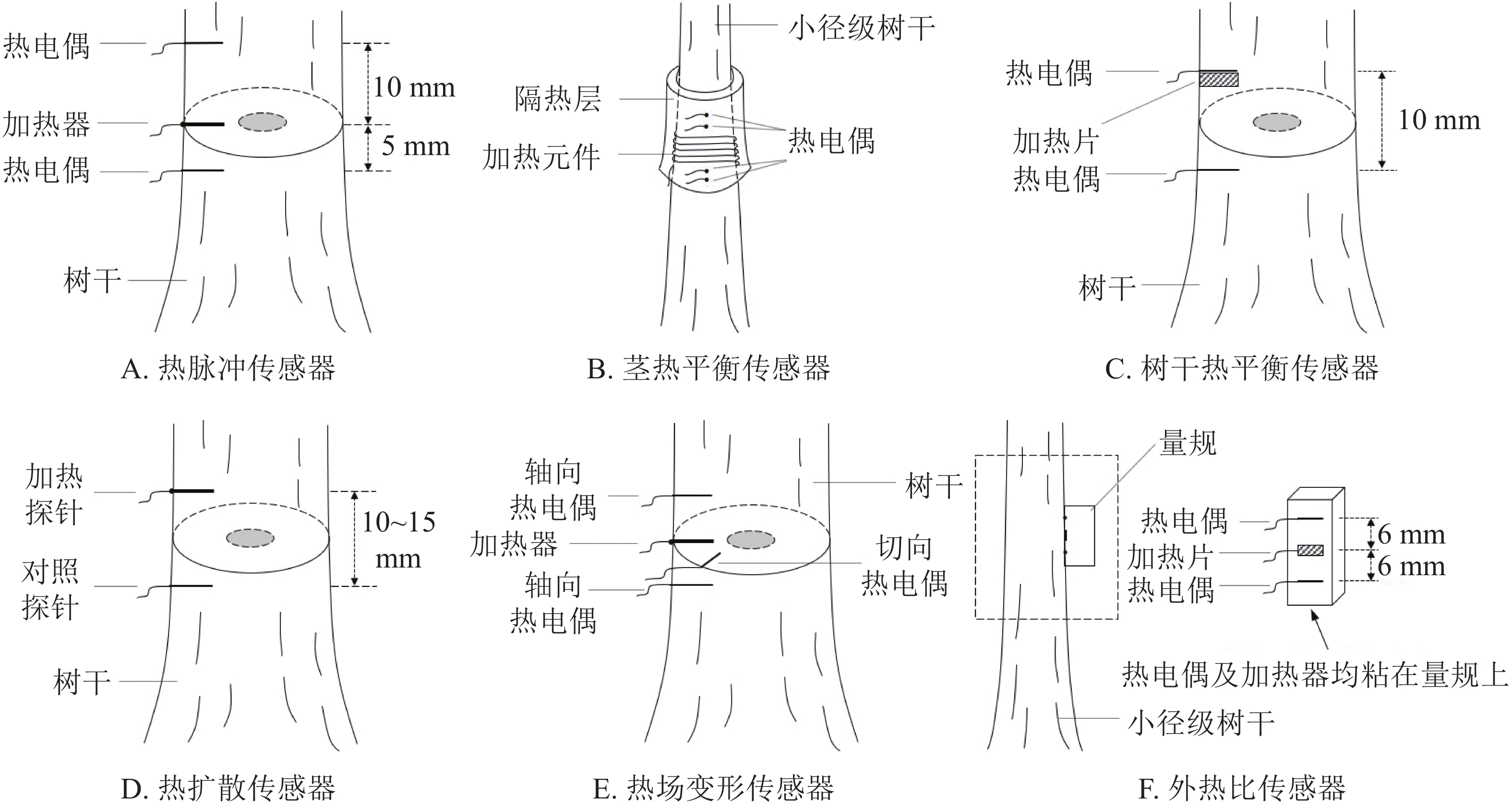
热脉冲法最早由HUBER等[16]提出,首次用热作为液体流动的示踪剂,利用热脉冲测量植物液流速率。该装置由加热元件和2个热电偶组成探针块,通过测量加热器发出热脉冲随着液流上升到达热电偶处所需的时间,计算液流速率(图1A)。由于热传导和对流会使得测量结果偏高,MARSHALL[17]利用热流方程建立了热脉冲法的模型框架,为热脉冲法的进一步发展提供了理论基础。

图 1 不同类型传感器示意图
Figure 1. Different kinds of heat-ratio sensor
基于脉冲加热的方法包括补偿热脉冲法、最大温差法(T-max法)以及热比率法。其中补偿热脉冲法(Compensation Heat Pulse Method,CHPM)通过测量2个对称放置在线性加热器两侧的温度传感器达到相同温度时的时间来计算液流密度,该装置安装探针时会对周围木材组织造成损伤而导致液流速率失真,因此需要根据不同探针间距设置校正参数[18],从而使液流速率的测量值更接近真实值。T-max法[19]的装置由加热器和1个温度探针组成,通过记录从发出热脉冲至温度探针到达最大温度时的时间,再根据MARSHALL的基础理论计算液流速率。该方法设备简单,仅需确定被测树干边材的导热系数,即可计算树干液流密度。热比率法(Heat Ratio Method,HRM)基于补偿热脉冲法提出[20],测量范围可以达到零值甚至延伸至负值,其装置由1个加热器和2个安装在加热器上下游的温度探针组成,通过测定2个探针的增温比即可计算液流速率。
-
热平衡法(Heat Balance Method,HBM)的原理是当树干内通过一定量液流时,加热元件作为热源会向树干提供已知的热量,直至树干温度趋于稳定。若不考虑热传导以及隔热层损失热量,热源提供的热量应与被液流带走的热量相等,可根据这种热平衡关系计算液流速率。热平衡法可分为茎热平衡法和树干热平衡法[21]。
-
茎热平衡法(Stem Heat Balance,SHB)[22]以环形加热元件作为热源,提供稳定的热量,热量散失途径包括树干液流带走、热传导向树干上下方散失和对流散失。装置(图1B)设计为包裹式,利用包裹式隔热层(通常为聚苯乙烯泡沫)防止热辐射造成的热量散失(树干周围的热辐射忽略不计)。加热元件上下方安装2对热电偶,用来测定液流通过后的温差,依据热量平衡关系计算液流。SHB法适用于测定胸径较小的树干,其优点是检测时不需要标定,不需要将热电偶插入树干中,对树木无直接损伤。
-
树干热平衡法(Trunk Heat Balance,THB)[23]的原理与茎热平衡法类似,均通过热量平衡关系计算液流,不同点是THB法测量装置(图1C)由插入树干的加热片和1对热电偶组成,2个热电偶分别安装在紧挨加热片上端(温度场最大值)和加热片下端(不受温度场影响)位置,通过记录液流通过前后树干温度差来计算液流。THB法同样不需要标定,并且可测定胸径较大的树干。但THB法设备较多,安装相对复杂,易对树干造成微损伤。目前热平衡法的应用较为广泛,通常用来研究环境因子与液流速率的关系以及耗水特性。
-
热扩散法(Thermal Dissipation Method,TDM)[24]又称Granier法,是目前应用最广泛的液流测定方法。该装置(图1D)包含2个传感器探针,沿液流方向插入树干中。下游(上部)探针包括加热元件(长约20 mm),并缠绕在装有热电偶的钢针上,热电偶尖端正对加热元件的中间;下游(上部)探针不加热,用作参考探头,以测量木质部的环境温度。工作时下游探针以恒定功率(0.2 W)连续加热,受液流的散热影响,2个探头间存在温度差异,因此可通过温差与液流速率间的关系计算液流速率。
TDM法常用来研究树干液流与环境因子的关系。万艳芳等[25]应用热扩散技术测定并分析了青海云杉Picea crassifolia树干液流密度与环境因子的关系,确定液流密度的主要环境影响因子是太阳辐射。朱敏捷等[26]利用热扩散法测定了尾叶桉Eucalyptus urophylla树干液流,研究了树干液流的方位差异以及与环境因子的关系。姚增旺等[27]应用热扩散探针测定梭梭Haloxylon ammodendron树干液流,研究了树干液流与环境因子之间的时滞效应。另外,通过测定单株树干液流还可以推算林分蒸腾量。王志超等[28] 研究了林分蒸腾耗水规律后发现:忽略夜间林分蒸腾耗水量会导致对林分蒸腾耗水量的估计不准确。
-
基于热脉冲法测量树木液流密度时,需要测量热脉冲前后温度,获得温度差,这就要求木材具有较高的热稳定性;热脉冲法两侧测量需要时间间隔,可以测得液流密度的最大值为45 cm3·cm−2·h−1,说明该方法具有一定的局限性。为解决以上问题,NADEZHDINA等[29]提出了热场变形法(Heat Field Deformation,HFD),通过记录线性加热器周围的木质部中不同径向位置的热场变化,将热场变形与树干木质部的液流密度联系起来。热场变形法液流检测系统(图1E)包括3个探针和1个加热器,其中2个探针沿轴向对称安装在加热器的上游和下游,另1个探针沿切向平行于加热器水平安装在加热器侧边,轴向探针测量对称温差,切向探针测量不对称温差。通过测定加热器周围轴向和切向的温度差来表征由树液流动而产生的热场变化,进而确定液流密度。液流密度q (cm3·cm−2·h−1)的一般计算公式为:
$$ q=3\; 600D_{{\rm{st}}}(K+T_{{\rm{s}}-{\rm{a}}})/T_{{\rm{as}}}Z_{{\rm{ax}}}Z_{{\rm{tg}}}L_{{\rm{sw}}}。 $$ 其中:Dst表示树干边材热扩散率(m2·s−1);(K+Ts-a)/Tas表示温差比率;ZaxZtg表示传感器探针间距离的校正因子;Lsw表示边材深度。K表示零液流下Ts-a的绝对值,其中Ts-a为Tsym与Tas的差。Tsym表示对称探针间的温差;Tas表示非对称探针间的温差;Zax表示轴向上游探针与加热器的距离;Ztg表示切向探针与加热器的距离。
HFD传感器也可以记录反向流量,即将Tsym改为负值。因此,计算公式转换为:
$$ q=-3 \;600D_{{\rm{st}}}(-K+T_{{\rm{s}}-{\rm{a}}})/T_{{\rm{as}}}Z_{{\rm{ax}}}Z_{{\rm{tg}}}L_{{\rm{sw}}}。 $$ 用HFD法测量液流密度,非零液流下,利用线性外推法能准确测得零液流密度,相比其他热技术方法优势显著。同时HFD法结合了对称与非对称温差测量,利用对称温差测量低液流密度较为有效,而测得的高液流密度与实际蒸腾量线性关系不显著,因此高液流密度准确性不够[30]。而利用非对称温差测量是中高液流密度准确性较高。因此,HFD法对于低液流量和高液流量都可以准确测定。
HFD法广泛应用于液流指数(the sap flow index,SFI) 的测定。SFI是植物水分状况的敏感指标,用来决定植物是否需要灌溉。SFI值通过在加热器周围轴向等距安装差动热电偶测得,是液流速率测定的原始数据之一[31]。此外,HFD法可以直接监测木质部的水分运动[32],通过沿着木质部半径的不同深度,用围绕普通线性加热器的传感器进行液流测定,具有快速响应和高度敏感的特性。NADEZHDINA[33]在对枫树Acer spp. 水运输路径的研究中,利用HFD法测定枫树木质部液流,证实了枫树的维管结构具有完整拓扑结构。
-
外热比法(External Heat-Ratio,EHR)是在热比率法的基础上提出的,用外部加热元件代替插入式加热元件,其基本原理与激光脉冲法(laser heat-pulse gauge,LHPG)类似。不同点是后者用近红外激光源代替插入式加热元件,并通过红外摄像机从外部监控热量传播,热脉冲速度由温度数据确定,并与液流速率相关。HELFTER等[34]利用激光脉冲法对小茎木本植物的液流速率进行了测定,发现小茎木本植物韧皮部与木质部液流速率几乎一致。CLEARWATER等[35]首次提出了外热比法(图1F),将1个微型外部加热器(电子芯片电阻)和温度传感器(精密热电偶)粘在软木块上,并压在茎干表面。释放热脉冲后,根据2个热电偶的增温比来计算液流密度。利用外热比法可以测定灌木液流速率[36],研究植物水动力学,对直径较小的茎干具有良好的适用性。外热比法最小可测直径为5 mm,可测液流密度为0.36~50.00 cm3·cm−2·h−1,较少应用于直径较大的茎干。因此,下一步可改进EHR技术,用于测定较大茎段植物的液流密度。
-
探针的使用会对树干边材造成一定的破坏,使得探针处树干边材的热均匀性改变,从而降低测量结果的准确性。GREEN等[37]用二维的“热-液流”模型确定不同伤口大小的校正因子,给出了补偿脉冲法和T-max法的校正因子表,并通过比较美洲黑杨Populus deltoides与白柳Salix alba的液流通量值与实际蒸腾速率值的关系证明了校正因子的有效性。TESTI等[38]在补偿热脉冲法的基础上提出了校准平均梯度法(calibrated average gradient,CAG),有效测定了低速液流,使用也较为简易。
LANGENSIEPEN等[39] 发现:为更好地适应小麦Triticum aestivum茎的解剖结构和热物理特性,在应用茎热平衡法测量小麦液流速率时,通过引入降噪方程可有效提高液流计的测量精度。TRCALA等[40]利用热场变形法的温度场理论,通过改变传感器的几何形状(从垂直到水平)来改善热平衡法的传感性能,实现了零液流和反向液流的测定。这种方法也被称为线性热平衡法[41],是从基础传导—对流传热方程解析得出的精确方程,不仅提高了液流测定的精度,而且基于热导率信息实现了水含量的估算。NAKANO等[42]发现:对金柑Fortunella crassifocia进行环剥处理后,可利用热平衡法测定其韧皮部和木质部的液流速率。
TDM法测定液流速率需要估算线性回归关系,确定零液流状态下的温差,而这个过程会产生一定误差[43],许多情况下准确性受到质疑[44]。因此用热扩散法确定树木的蒸腾量时,有必要对测量树种液流量估计方程进行校准[45-47]。
外热比法也存在一些不足。首先,大多数加热传感器从加热芯片的中心到两侧感温元件有一定的窄间距。随着热量沿横截面向内传播和沿茎轴上下传播,热量到达木质部导管时变得非常分散,来自液流的热比率信号会减弱。其次,加热器和温度传感器被安置在1个矩形的不导电硅酮/软木块中,无法有效隔绝环境温度对检测温度的影响,增加了液流检测结果的误差。再次,矩形加热芯片横压在圆柱形树干上,载荷不均匀,加热器元件使用窄的矩形芯片电阻,比圆形芯片更容易断开。为此,王胜[48]开发了1种新设计的EHR加热传感器,增加了加热元件至温度传感器的间距,使之更适应直径较大的茎干。改良后的装置茎干直径检测范围扩大,可用于胸径较小的树木测量。
-
目前,基于热技术的树干液流测定方法日趋完善,不同方法具有相对应的优势和劣势。由表1可知:不同热技术方法液流测定装置均包括为加热器提供能量的能量供应单元和用于收集检测数据的数据记录仪。具体来看,热脉冲法不受环境条件以及树冠结构及根系特性的影响,装置简洁,但存在一定的灵敏度和精度问题。热平衡法无需标定,测量精度有所提高,但仅适于测定高液流密度。热扩散法是目前研究蒸腾耗水特性应用最广泛的方法,测定结果较准确,仪器成本较低,安装简单,有较成熟的商业化产品,但测定结果容易被低估。热场变形法操作复杂,应用较少,但该方法能够准确测定零液流以及逆向液流,测定精度与范围也有很大的提升。外热比法与激光脉冲法均可实现精确的零破坏检测,但仅适用于胸径较小的树干,另外,激光脉冲法装置成本昂贵,未能普及。
表 1 树干液流的测定方法对比
Table 1. Comparison of methods for sap flow measurement
方法 装置 优点 缺点 热脉冲法 加热器,2个热电偶 不受环境条件,树冠结构及根系特性的影响,简洁准确, 经济可行[49] 存在测定精度问题[37−38] 热平衡法 探针,加热元件 无需标定,进一步提高了测量精度[22−23] 不适用于液流速率较高的 植物[50] 热扩散法 加热探针,参照探针 测定结果较准确,仪器成本较低,安装简单,有较成熟的 商品化产品[51] 液流可能被低估[43] 激光热脉冲法 近红外激光源,红外摄像机 无须将热源插入植物茎干内,避免对茎干内组织破坏而造 成误差[34] 成本较高 热场变形法 加热器,3根探针 能够准确地测定零液流量以及逆向液流[52] 测定过程较复杂[29] 外热比法 微型外部加热传感器 精确、无损地测定胸径较小的树干中的双向液流[53] 微型外部量规的配置尚存 在问题[35−36] -
利用热技术方法测定液流速率,可以精确估算树木蒸腾耗水量[54],但不同热技术方法的测量精度、测定范围以及适用性不尽相同,实际应用时应根据不同实验条件选择不同的热技术方法。选择热技术方法测定树干液流通常需要考虑树木胸径大小、热技术方法的误差范围、热技术方法的测定精度、热技术方法的可行性等因素。
首先,不同胸径活立木应选用不同的热技术方法,外热比法和茎热平衡法适合测定胸径较小的树木,树干热平衡法适合测定胸径较大的树木。其次,不同热技术方法可测得的液流速率范围不同,补偿热脉冲法、T-max法以及热扩散法测定低速液流的误差较大,茎热平衡法测定高液流速率时误差较大,热比率法和热场变形法测定液流范围较广。热场变形法和外热比法可以测定逆向液流,热场变形法还可以准确测定零液流。热脉冲法、热平衡法和热扩散法较成熟[55],应用较广泛,可行性较高。另外,将不同测定范围的热技术方法组合使用,可以有效提高测定精度。不同植物的液流速率不同。向日葵Helianthus annuus和玉米Zea mays等植物的液流速率相对较低,在利用T-max方法测定时,测量值总是略高于实际值[56];换成热比率法测定也不够精确,而采用T-max法与热比率法组合测量则较为准确。
-
利用热技术方法测定液流速率约80多年的研究,方法不断得到改进与创新,在校准度、测定范围和测定精度上均有所提高,同时,实验操作不断简化,数据实现自动化采集和存储,并逐步实现连续时间以及多层空间的同步测定[57]。其中,热脉冲法、热平衡法、热扩散法经一系列的发展与完善,极大程度上减小了测量误差[58]。热扩散法还形成了成熟的商业化产品,并得到了广泛的应用。虽然利用外热比法和热场变形法测定活立木液流速率的研究有限,但外热比法实现了精确的零破坏检测,热场变形法液流速率测定范围广,并可准确的测定零液流和逆向液流。在利用外热比法测定液流速率时,需要针对不同样本以及实验条件设计不同的量规,这是外热比法的不足之处。因此,外热比法和热场变形法亟待更为深入研究。
目前,热技术方法成为液流测量的首要选择。在未来,应用热技术测定树干液流仍需关注以下热点:在完善研究活立木蒸腾耗水特点的同时,结合土壤生物因子和气象因子与树干液流的关系,进一步深入研究活立木生理作用;从微观和宏观方面监控水分运动,研究水分利用与树木生长的关系;在生产实际方面,进一步完善活立木单株和森林林区的数据监控,为实现高效的林区治理提供有力依据。
Overview of the methods for sap flow measurement of standing tree based on thermal technology
-
摘要: 比较不同研究目标和实验条件下适用的基于热技术的树干液流测量方法,展望不同热技术方法在液流测定中的应用前景,为活立木液流测量的方法选择提供参考。目前常用的基于热技术的液流检测方法包括热脉冲法、热平衡法、热扩散法、热场变形法和外热比法等5种。其中,热脉冲法通过测定热脉冲到达热电偶处所需的时间来计算液流速率,热平衡法通过加热元件输入的热量、热传导以及液流携带热量之间的能量平衡关系计算液流。而热扩散法、热场变形法和外热比法则是利用温度差值来表征树液流动产生的热场变化,进而确定液流速率和液流密度。近年来,结合相关热技术原理对已有热技术方法进行了改进,热平衡法、热场变形法和外热比法的测定精度有所提高。但热场变形法的测定过程较为复杂,外热比法的外部量规配置尚待探讨,应用并不广泛。热扩散法和热脉冲法虽然已经具备较成熟的商品化产品,但测定精度仍存在问题。液流速率的检测精度直接影响研究结果的准确性。在测定液流速率时,需依据不同条件选择适合的方法,从而有效提高液流速率的测定精度。图1表1参58Abstract: With the comparion of the five thermal methods commonly used for trunk sap flow measurement with the focus on their respective advantages and disadvantages under different research objectives and experimental circumstance, this study is aimed to provide reference for practices in liquid flow measurements of standing trees. Of the five thermal methods commonly employed, Heat Pluse Velocity Method (HPVM) has mainly been used to calculate the flow rate by measuring the time required for the heat pulse to reach the thermocouple. Heat Balance Method (HBM) is used to calculate the sap flow by means of the energy balance between the heat input via the heating element, the heat conduction and the heat carried by the sap flow. Whereas Thermal Dissipation Method (TDM), Heat Field Deformation (HFD), and External Heat-Ratio (EHR) all use the temperature difference to characterize the thermal field changes caused by the sap flow before the sap flow rate and density are determined. In recent years, with the employment of relevant thermal technology, researches aimed to improve the existing thermal technology methods have been rewarded with higner sap flow measurement accuracy of HBM, HFD and EHR. Neither HBM nor EHR has been widely employed due to the complex measurement process and uncertainly in the configuration of the micro external gauge respectively. TDM and HPVM still suffer from low accuracy of determination despite the relatively mature commercial products of them. In conclusion, improperly selected determination methods will lead to great deviation in research rusults, therefore, is it proposed that suitable methods should be chosen in accordance with different experimental conditions so as to better promote the measurement accuracy of tree sap flow. [Ch, 1 fig. 1 tab. 58 ref.]
-
表 1 树干液流的测定方法对比
Table 1. Comparison of methods for sap flow measurement
方法 装置 优点 缺点 热脉冲法 加热器,2个热电偶 不受环境条件,树冠结构及根系特性的影响,简洁准确, 经济可行[49] 存在测定精度问题[37−38] 热平衡法 探针,加热元件 无需标定,进一步提高了测量精度[22−23] 不适用于液流速率较高的 植物[50] 热扩散法 加热探针,参照探针 测定结果较准确,仪器成本较低,安装简单,有较成熟的 商品化产品[51] 液流可能被低估[43] 激光热脉冲法 近红外激光源,红外摄像机 无须将热源插入植物茎干内,避免对茎干内组织破坏而造 成误差[34] 成本较高 热场变形法 加热器,3根探针 能够准确地测定零液流量以及逆向液流[52] 测定过程较复杂[29] 外热比法 微型外部加热传感器 精确、无损地测定胸径较小的树干中的双向液流[53] 微型外部量规的配置尚存 在问题[35−36]  下载: 导出CSV
下载: 导出CSV
-
[1] 徐利岗, 杜历, 李金泽, 等. 基于SPAC系统的干旱区枸杞蒸腾耗水模拟与分析[J]. 节水灌溉, 2017(7): 1 − 5, 10. XU Ligang, DU Li, LI Jinze, et al. Simulation and analysis of transpiration water consumption of Lycium barbarum in arid area based on SPAC system [J]. Water Sav Irrig, 2017(7): 1 − 5, 10. [2] 郭孟霞, 毕华兴, 刘鑫, 等. 树木蒸腾耗水研究进展[J]. 中国水土保持科学, 2006, 4(4): 114 − 120. GUO Mengxia, BI Huaxing, LIU Xin, et al. Review on the water consumption of tree transpiration [J]. Sci Soil Water Conserv, 2006, 4(4): 114 − 120. [3] 巨关升, 刘奉觉, 郑世锴, 等. 稳态气孔计与其它3种方法蒸腾测值的比较研究[J]. 林业科学研究, 2000, 13(4): 360 − 365. JU Guansheng, LIU Fengjue, ZHENG Shikai, et al. A comparison on the transpiration values measured with steady state porometer and other three methods [J]. For Res, 2000, 13(4): 360 − 365. [4] LAGERGREN F, LINDROTH A. Transpiration response to soil moisturein pine and spruce trees in Sweden [J]. Agric For Meteorol, 2002, 112(2): 67 − 85. [5] 刘子奇, 王渝淞, 张欢, 等. 基于稳定同位素和热比率技术的侧柏水分逆向运移特征与过程[J]. 应用生态学报, 2020, 31(6): 1817 − 1826. LIU Ziqi, WANG Yusong, ZHANG Huan, et al. Characteristics and processes of reverse sap flow of Platycladus orientalis based on stable isotope technique and heat ratio method [J]. Chin J Appl Ecol, 2020, 31(6): 1817 − 1826. [6] 孙守家, 孟平, 张劲松, 等. 基于氘示踪剂和热扩散技术的栓皮栎水分运输速率与效率研究[J]. 生态学报, 2012, 32(12): 3892 − 3901. SUN Shoujia, MENG Ping, ZHANG Jinsong, et al. Water transport velocity and efficiency in Quercus variabilis detected with deuterium tracer and thermal dissipation technique [J]. Acta Ecol Sin, 2012, 32(12): 3892 − 3901. [7] 孙慧珍, 周晓峰, 康绍忠. 应用热技术研究树干液流进展[J]. 应用生态学报, 2004, 15(6): 1074 − 1078. SUN Huizhen, ZHOU Xiaofeng, KANG Shaozhong. Research advance in application of heat technique in studying stem sap flow [J]. Chin J Appl Ecol, 2004, 15(6): 1074 − 1078. [8] 赵平, 邹绿柳, 饶兴权, 等. 成熟马占相思林的蒸腾耗水及年际变化[J]. 生态学报, 2011, 31(20): 6038 − 6048. ZHAO Ping, ZHOU Lüliu, RAO Xingquan, et al. Water consumption and annual variation of transpiration in mature Acacia mangium plantation [J]. Acta Ecol Sin, 2011, 31(20): 6038 − 6048. [9] 蔡锡安, 赵平, 曾小平, 等. 两种木兰科植物的树干液流特征及其与环境因子的关系[J]. 生态学杂志, 2012, 31(9): 2163 − 2169. CAI Xi’an, ZHAO Ping, ZENG Xiaoping, et al. Dynamics of xylem sap flow of two Magnoliaceae tree species in relation to environmental factors [J]. Chin J Ecol, 2012, 31(9): 2163 − 2169. [10] 赵春彦, 司建华, 冯起, 等. 树干液流研究进展与展望[J]. 西北林学院学报, 2015, 30(5): 98 − 105. ZHAO Chunyan, SI Jianhua, FENG Qi, et al. Stem sap flow research: progress and prospect [J]. J Northwest For Univ, 2015, 30(5): 98 − 105. [11] 王檬檬, 党宏忠, 李钢铁, 等. 晋西黄土区苹果树液流特征及其与环境因子的关系[J]. 中国农业科技导报, 2020, 22(7): 140 − 147. WANG Mengmeng, DANG Hongzhong, LI Gangtie, et al. Sap flux density characteristics of apple orchards and their relationship with environmental factors in gully region of loess plateau [J]. J Agric Sci Technol, 2020, 22(7): 140 − 147. [12] 温淑红, 韩新生, 蔡进军, 等. 宁南黄土丘陵区山桃树干液流速率及其与气象因子的关系[J]. 西南农业学报, 2020, 33(6): 1301 − 1308. WEN Shuhong, HAN Xinsheng, CAI Jinjun, et al. Relationships between sap flow velocity in tree trunks of Amygdalus davidiana and meteorological factors in loess hilly regions of southern Ningxia [J]. Southwest China J Agric Sci, 2020, 33(6): 1301 − 1308. [13] 杨洁, 吕金林, 何秋月, 等. 黄土丘陵区辽东栎和刺槐树干液流时滞效应与蒸腾特征的关联性[J]. 应用生态学报, 2019, 30(8): 2607 − 2613. YANG Jie, LÜ Jinlin, HE Qiuyue, et al. Time lag of stem sap flow and its relationships with transpiration characteristics in Quercus liaotungensis and Robina pseudoacacia in the loess hilly region, China [J]. Chin J Appl Ecol, 2019, 30(8): 2607 − 2613. [14] 孙慧珍, 康绍忠, 胡笑涛. 梨树干木质部液流速度径向分布特征[J]. 园艺学报, 2008, 35(7): 937 − 944. SUN Huizhen, KANG Shaozhong, HU Xiaotao. The radial distribution of xylem sap flow velocity in trunks of pear trees [J]. Acta Hortic Sin, 2008, 35(7): 937 − 944. [15] 靳新红, 王百田, 郭红艳, 等. 黄土半干旱区枣、榆水分利用效率的比较研究[J]. 中国生态农业学报, 2009, 17(1): 90 − 93. JIN Xinhong, WANG Baitian, GUO Hongyan, et al. Comparison of water use efficiency of Zizyphus jujube and Ulmus pumila in semi-arid zones of the Loess Plateau [J]. Chin J Eco-Agric, 2009, 17(1): 90 − 93. [16] HUBER B, SCHMIDT E. Eine kompensationsmethode zur thermoelektrischen messung langsamer saftströme [J]. Ber Deutsch Bot Ges, 1937, 59(9): 514 − 529. [17] MARSHALL D C. Measurement of sap flow in conifers by heat transport [J]. Plant Physiol, 1958, 33(6): 385 − 396. [18] SWANSON R, WHITFIELD D. A numerical analysis of heat pulse velocity theory and practice [J]. J Exp Bot, 1981, 32(1): 221 − 239. [19] COHEN Y, FUCHS M, GREEN G C. Improvement of the heat pulse method for determining sap flow in trees [J]. Plant Cell Environ, 1981, 4(5): 391 − 397. [20] BURGESS S S O, ADAMS M A, TURNER N C, et al. An improved heat pluse method to measure low and reverse rates of sap flow in woody plants [J]. Tree Physiol, 2001, 21: 589 − 598. [21] 岳广阳, 张铜会, 刘新平, 等. 热技术方法测算树木茎流的发展及应用[J]. 林业科学, 2006, 42(8): 102 − 108. YUE Guangyang, ZHANG Tonghui, LIU Xinping, et al. Development and application of thermal methods in measuring stem sap flow [J]. Sci Silv Sin, 2006, 42(8): 102 − 108. [22] SAKURATANI T. A heat balance method for measuring water flux in the stem of intact plants [J]. J Agric Meteorol, 1981, 37(1): 9 − 17. [23] ČERMÁK J, DEML M, PENKA M. A new method of sap flow rate determination in trees [J]. Biol Plantrum, 1973, 15(3): 171 − 178. [24] GRANIER A. A new method of sap flow measurement in tree stems [J]. Ann Des Sci For, 1985, 42(2): 193 − 200. [25] 万艳芳, 于澎涛, 刘贤德, 等. 祁连山青海云杉树干液流密度的优势度差异[J]. 生态学报, 2017, 37(9): 3106 − 3114. WAN Yanfang, YU Pengtao, LIU Xiande, et al. Variation in sap flow density among levels of tree dominance in Picea crassifolia in the Qilian Mountains [J]. Acta Ecol Sin, 2017, 37(9): 3106 − 3114. [26] 朱敏捷, 赵从举, 徐文娴, 等. 尾叶桉树干液流方位差异及其对环境因子的响应[J]. 海南师范大学学报(自然科学版), 2017, 30(2): 177 − 184. ZHU Minjie, ZHAO Congju, XU Wenxian, et al. The differences of Eucalyptus urophylla stem sap flow velocities in different directions and relations between velocities and environmental factors [J]. J Hainan Norm Univ Nat Sci, 2017, 30(2): 177 − 184. [27] 姚增旺, 褚建民, 吴利禄, 等. 民勤绿洲荒漠过渡带梭梭树干液流的时滞特征[J]. 应用生态学报, 2018, 29(7): 2339 − 2346. YAO Zengwang, CHU Jianmin, WU Lilu, et al. Time lag characteristics of the stem sap flow of Haloxylon ammodendron in the Minqin oasis desert ectone, China [J]. Chin J Appl Ecol, 2018, 29(7): 2339 − 2346. [28] 王志超, 许宇星, 竹万宽, 等. 雷州半岛尾叶桉人工林夜间耗水特征及驱动因素[J]. 浙江农林大学学报, 2020, 37(4): 646 − 653. WANG Zhichao, XU Yuxing, ZHU Wankuan, et al. Characteristics and driving factors of nocturnal water consumption of Eucalypt usurophylla plantations in Leizhou Peninsula [J]. J Zhejiang A&F Univ, 2020, 37(4): 646 − 653. [29] NADEZHDINA N, VANDEGEHUCHTE M W, STEPPE K. Sap flux density measurements based on the heat field deformation method [J]. Trees, 2012, 26(5): 1439 − 1448. [30] TRIBUTSCH H, NADEZHDINA N, CERMÁK J. Infrared images of heat fields around a linear heater in tree trunks: what can be learned about sap flow measurement? [J]. Ann For Sci, 2006, 63: 653 − 660. [31] NADEZHDINA N, ČERMAK J, DOWNEY A, et al. Sap flow index as an indicator of water storage use [J]. J Hydrol Hydromechan, 2015, 63(2): 124 − 133. [32] NADEZHDINA N, DAVID J S, PINTO C A, et al. Root sap flow as a tool to establish hydro logical thresholds for plant growth and survival [J/OL]. Agric Water Manage, 2020, 241: 106388[2021-02-12]. doi: 10.1016/j.agwat.2020.106388. [33] NADEZHDINA N. Integration of water transport pathways in a maple tree: responses of sap flow to branch severing [J/OL]. Ann For Sci, 2010, 67: 107[2021-03-01]. doi: 10.1051/forest/2009092 . [34] HELFTER C, SHEPHARD J D, MARTÍNEZ-VILALTA J, et al. A noninvasive optical system for the measurement of xylem and phloem sap flow in woody plants of small stem size [J]. Tree Physiol, 2007, 27(2): 169 − 179. [35] CLEARWATER M J, LUO Zhiwei, MAZZEO M, et al. An external heat pulse method for measurement of sap flow through fruit pedicels, leaf petioles and other small-diameter stems [J]. Plant Cell Environ, 2009, 32(12): 1652 − 1663. [36] SKELTON R P, WEST A G, DAWSON T E, et al. External heat-pulse method allows comparative sap flow measurements in diverse functional types in a Mediterranean-type shrubland in South Africa [J]. Func Plant Biol, 2013, 40(10): 1076 − 1087. [37] GREEN S, CLOTHIER B, JARDINE B. Theory and practical application of heat pulse to measure sap flow [J]. Agron J, 2003, 95(6): 1371 − 1379. [38] TESTI L, VILLALOBOS F J. New approach for measuring low sap velocities in trees [J]. Agric For Meteorol, 2009, 149(3): 730 − 734. [39] LANGENSIEPEN M, KUPISCH M, GRAF A, et al. Improving the stem heat balance method for determining sap-flow in wheat [J]. Agric For Meteorol, 2014, 186: 34 − 42. [40] TRCALA M, ČERMÁK J. Improvement of the trunk heat balance method including measurement of zero and reverse sap flows [J]. Agric For Meteorol, 2012, 166/167: 120 − 126. [41] TRCALA M, ČERMÁK J. A new heat balance equation for sap flow calculation during continuous linear heating in tree sapwood [J]. Appl Therm Eng, 2016, 102: 532 − 538. [42] NAKANO Y, IWASAKI N. Determination of phloem sap flow rate using a combination of the heat balance method and girdling in citrus [J/OL]. Agric For Meteorol, 2019, 278: 107669[2021-01-20]. doi: 10.1016/j.agrformet.2019.107669. [43] LU Ping, URBAN L, ZHAO Ping. Granier’s thermal dissipation probe (TDP) method for measuring sap flow in trees: theory and practice [J]. Acta Bot Sin, 2004, 46(6): 631 − 646. [44] 马玉洁, 李春友, 武鹏飞, 等. 基于实测白榆蒸腾速率校正计算液流速率的Granier原始公式[J]. 林业科学, 2020, 56(6): 179 − 185. MA Yujie, LI Chunyou, WU Pengfei, et al. Correction of Granier’s original formula coefficient for calculating sap flow based on the measured transpiration rate of Ulmus pumila [J]. Sci Silv Sin, 2020, 56(6): 179 − 185. [45] SANTOS I M S, VELLAME L M, ARAÚJO J F, et al. Calibration of the thermal dissipation probe for atemoya (Annona squamosa × A. cherimola) [J]. Engenharia Agrícola, 2020, 40(4): 545 − 554. [46] PASQUALOTTO G, CARRARO V, MENARDI R, et al. Calibration of Granier-Type (TDP) sap flow probes by a high precision electronic potometer [J/OL]. Sensors, 2019, 19: 2419[2021-02-10]. doi: 10.3390/s19102419. [47] LINK R M, FUCHS S, AGUILAR D A, et al. Tree height predicts the shape of radial sap flow profiles of Costa-Rican tropical dry forest tree species [J]. Agric For Meteorol, 2020, 287:107913[2021-03-13]. doi: 10.1016/s.agrformet.2020.107913. [48] 王胜. 水蚀风蚀交错区典型乔灌树种蒸腾耗水特征研究[D]. 北京: 中国科学院, 2019. WANG Sheng. Transpiration Characteristics of Typical Shrub and Tree Species in the Water-Wind Erosion Crisscross Region [D]. Beijing: Chinese Academy of Sciences, 2019. [49] 杜梦鸽, 王胜, 樊军. 五针热脉冲探头在测定树干液流中的应用[J]. 应用生态学报, 2017, 28(8): 2438 − 2444. DU Mengge, WANG Sheng, FAN Jun. Application of the penta-needle heat pulse probes to determine the stem sap flow [J]. Chin J Appl Ecol, 2017, 28(8): 2438 − 2444. [50] 田清远. 基于热平衡方式的植物茎流检测系统研究[D]. 昆明: 昆明理工大学, 2013. TIAN Qingyuan. A Study of Sap Flow Detection System based on Heat Balance [D]. Kunming: Kunming University of Science and Technology, 2013. [51] 刘庆新. 热扩散法液流误差校正及其在林分水量平衡中的应用[D]. 北京: 中国林业科学研究院, 2012. LIU Qingxin. Error Correction of Thermal Dissipation Method for Sap Flow and Its Application in the Research of Stand Water Balance [D]. Beijing: Chinese Academy of Forestry, 2012. [52] NADEZHDINA N, ČERMÁK J, CEULEMANS R. Radial patterns of sap flow in woody stems of dominant and understory species: scaling errors associated with positioning of sensors [J]. Tree Physiol, 2002, 22(13): 907 − 918. [53] CLEARWATER M J, MEINZER F C, ANDRADE J L, et al. Potential errors in measurement of nonuniform sap flow using heat dissipation probes [J]. Tree Physiol, 1999, 19(10): 681 − 687. [54] 郑怀舟, 朱锦懋, 魏霞, 等. 5种热动力学方法在树干液流研究中的应用评述[J]. 福建师范大学学报(自然科学版), 2007, 23(4): 18 − 23. ZHENG Huaizhou, ZHU Jinmao, WEI Xia, et al. Review on the five thermodynamic technology applied in the research of sap flow [J]. J Fujian Norm Univ Nat Sci Ed, 2007, 23(4): 18 − 23. [55] 孙慧珍, 孙龙, 王传宽, 等. 东北东部山区主要树种树干液流研究[J]. 林业科学, 2005, 41(3): 36 − 42. SUN Huizhen, SUN Long, WANG Chuankuan, et al. Sap flow of the major tree species in the eastern mountainous region in northeast China [J]. Sci Silv Sin, 2005, 41(3): 36 − 42. [56] MINER G L, HAM J M, KLUITENBERG G J. A heat-pulse method for measuring sap flow in corn and sun- flower using 3D-printed sensor bodies and low-cost electronics [J]. Agric For Meteorol, 2017, 246: 86 − 79. [57] SIQUEIRA J M, PAÇO T A, da SILVA J M, et al. Biot-Granier sensor: a novel strategy to measuring sap flow in trees [J/OL]. Sensors, 2020, 20: 3538[2021-01-28]. doi: 10.3390/s20123538. [58] 龙秋波, 贾绍凤. 茎流计发展及应用综述[J]. 水资源与水工程学报, 2012, 23(4): 18 − 23. LONG Qiubo, JIA Shaofeng. Review on the development and application of sap flow gauge [J]. J Water Resour Water Eng, 2012, 23(4): 18 − 23. -

-
链接本文:
https://zlxb.zafu.edu.cn/article/doi/10.11833/j.issn.2095-0756.20210286
 点击查看大图
点击查看大图
图(1) / 表(1)
计量
- 文章访问数: 2440
- HTML全文浏览量: 687
- PDF下载量: 82
- 被引次数: 0


