




-
植物空间格局分析是研究植物群落特征、种内与种间关系及与环境关系的一种重要方法[1],被广泛用于群落动态变化研究,有利于揭示群落形成、演替和发展的内在机制[2]。空间分布格局类型主要有随机分布、均匀分布和聚集分布[1]。种群空间格局依赖于一定的空间尺度,可能在小尺度上表现为聚集分布,在大尺度上为随机分布或均匀分布[3-4]。空间关联性是植物群落特征的重要体现,有助于理解群落构建和演替动态过程[4]。空间关联性通常可分为空间正相关、负相关和无空间相关关系,可以反映群落内种间及种内相互作用关系[5]。植物空间格局的分析方法较多,传统的空间格局分析方法是基于单一尺度的若干个植物样方来研究植物种群的空间分布格局,只能认识到特定尺度上的空间分布格局[3-7]。RIPLEY[8]在1977年提出了使用K(t)函数分析种群的空间分布格局方法。随后,在K(t)函数的基础上又相继发展了O-ring函数、G(t)函数和Lest(t)函数[9-10],克服了单一尺度下空间分布格局方法的不足,现已被广泛用于研究不同空间尺度下群落空间分布格局以及种内与种间关系[11]。
川西云杉Picea likiangensis var. rubescens林主要分布在青藏高原东缘海拔3 000~4 100 m的区域,是中国西南地区亚高山针叶林的重要组成部分,在固碳、涵养水源和保护生物多样性等方面发挥着多种生态功能,是青藏高原生态安全屏障区[12-13]。20世纪40年代以来,川西云杉林遭受大面积砍伐和破坏,区域森林生态系统严重退化[14]。由于区域环境条件较为恶劣、自然恢复效果缓慢,森林生态恢复方式主要采用人工造林。从20世纪60年代开始,在青藏高原东缘亚高山区域陆续开展了人工造林工程,造林树种以川西云杉为主,形成了大面积的川西云杉人工林[15]。目前,关于川西云杉人工林的研究多集中在群落物种多样性、水源涵养和土壤质量等方面[16-18],对川西云杉人工林群落空间分布格局及种间关联性的研究鲜有报道。因此,本研究以川西云杉人工林为研究对象,比较分析不同林龄的人工林与天然林群落空间格局及种间关联性的差异,有助于认识人工林群落动态变化和演替规律,为青藏高原东缘亚高山采伐迹地的生态恢复及人工林结构的优化与改造提供科学依据。
-
研究区位于四川省甘孜藏族自治州道孚县木茹乡林场,30°32′~32°21′N,100°32′~101°44′E,海拔3 600~3 900 m,地处青藏高原东缘。该区域属于典型的寒温带大陆性高原季风气候,冬季寒冷干燥,夏季温暖湿润,昼夜温差较大。依据国家气象台站(道孚县)数据(1957—2019年),年平均气温为8.1 ℃,最冷月和最暖月分别为1月和7月,平均气温分别为−1.9和16.0 ℃;年降水量为614.0 mm,85%的降水量集中在5—9月。土壤类型以山地棕壤和山地褐土为主。区域亚高山针叶林以川西云杉林、鳞皮冷杉Abies squamata林和红杉Larix potaninii林为主,分布海拔为3 200~4 200 m。人工造林以川西云杉为主要树种,栽种时采用5~7年生实生苗,初始密度为3 750株·hm−2。
-
在川西云杉人工林和天然林集中分布区,分别选择林龄30、40和50 a的人工林以及天然林,各设置3个20 m×20 m的样地,共12个样地(表1)。采用全球定位系统(GPS)记录每个样地的经纬度和海拔,使用地质罗盘记录样地的坡向和坡度。对每个样地内所有胸径≥1 cm的乔木树种个体进行逐木调查,记录每个个体的空间位置坐标,以西南角为坐标原点,并测量其胸径、高度、枝下高和冠幅等。依据野外调查数据,林龄30、40和50 a的人工林样地中保留的川西云杉平均株数分别为(66±1)、(60±5)和(44±7) 株;天然林样地内乔木平均株数为(42±18)株。
表 1 川西云杉人工林与天然林样地基本信息
Table 1. General information of each plot for plantation and natural forests of P. likiangensis var. rubescens
样地号 类型 林龄/a 林分密度/
(株·hm−2)云杉种群密度/
(株·hm−2)海拔/m 坡向 坡度/(°) 郁闭度 树种 AR30-1 人工林 30 1 650 1 625 3 688 东北 23 0.70 PL、BA、BP AR30-2 人工林 30 2 125 2 100 3 702 东北 26 0.70 AR30-3 人工林 30 1 825 1 825 3 692 东北 25 0.60 AR40-1 人工林 40 2 500 1 800 3 760 东北 38 0.80 PL、BU、SV、PD、CC、BA、SW、AS AR40-2 人工林 40 2 625 1 625 3 756 东北 37 0.85 AR40-3 人工林 40 2 600 1 925 3 728 东北 37 0.85 AR50-1 人工林 50 1 025 825 3 810 东北 30 0.65 PL、BP、BU、SV、SW、RV AR50-2 人工林 50 1 425 1 175 3 790 东北 36 0.80 AR50-3 人工林 50 2 025 1 325 3 770 东北 38 0.80 NF-1 天然林 >150 475 50 3 840 东北 6 0.45 PL、BU、RV、AS NF-2 天然林 >150 1 100 25 3 842 东北 8 0.50 NF-3 天然林 >150 1 600 25 3 845 东北 10 0.40 说明:PL为川西云杉,BA为红桦Betula albosinensis,BP为白桦B. platyphylla,BU为糙皮桦B. utilis,SV为川滇花楸Sorbus vilmorinii,PD为山杨Populus davidiana,CC为微毛樱桃Cerasus clarofolia,SW为皂柳Salix wallichiana,RV为亮叶杜鹃 Rhododendron vernicosum,AS为鳞皮冷杉Abies squamata -
空间点格局分析最初由RIPLEY[18]提出,后经DIGGLE等[19]发展,现已广泛应用于植物群落空间分布以及种内与种间空间关联性的研究中。Ripley’s K(t)函数公式如下:
$$ {K}{\left(t\right)}=\frac{A}{{n}^{2}}\sum\limits_{i = 1}^n \sum\limits_{j = 1}^n \frac{{I}_{t}\left({u}_{ij}\right)}{{w}_{ij}}\left(i\ne j\right) 。 $$ 其中:A是样地面积,n为样地所有个体总数,t为空间尺度,可以是>0 的任何值,
$ {u}_{ij} $ 是第i株与j株树木之间的距离,$ {I}_{t}\left({u}_{ij}\right) $ 是指示函数。当$ {u}_{ij} $ ≤t时,$ {I}_{t}\left({u}_{ij}\right) $ =1,当$ {u}_{ij} $ >t时,$ {I}_{t}\left({u}_{ij}\right) $ =0;$ {w}_{ij} $ 为权重值,用于边缘校正。Ripley’s K(t)函数是一个累积分布函数,在应用过程中常因为累积效应的存在影响结果的准确性[11],后在Ripley’s K(t)函数的基础上发展形成了L(t)函数。空间分布格局分析采用单变量L(t)函数,其公式为:
$$ {L}{\left(t\right)}=\sqrt{\frac{{K}{\left(t\right)}}{\text{π} }-t} 。 $$ 本研究空间尺度为0~5 m,利用R语言“spatial”程序包中Monte Carlo拟合检验,假设个体分布符合泊松分布,样方检验的零假设是完全空间随机零假设(complete spatial randomness, CSR),通过模拟泊松分布和99次独立模拟得到的点过程,计算估计L函数,获得模拟曲线的逐点上下包络。L(t)值位于上包迹线以上为聚集分布,在上下包迹线之间为随机分布,在下包迹线以下为均匀分布。
-
空间关联性运用双变量点格局分析方法,即L12(t)函数,其公式如下:
$$ {K}_{12}\left(t\right)=\frac{A}{{n}_{1}{n}_{2}}\sum\limits_{i = 1}^n \sum\limits_{j = 1}^n\frac{{I}_{t}\left({u}_{ij}\right)}{{w}_{ij}}\left(i\ne j\right) \text{;} $$ $$ {L}_{12}\left(t\right)=\sqrt{\frac{{K}_{12}\left(t\right)}{{\text{π}} }-t} 。 $$ 其中:n1、n2分别为样地内川西云杉和其他乔木植株的总数。 对于双变量点格局分析来说,
$ {L}_{12}{\left(t\right)} $ 值若位于上下包迹线内,则种间关系为不相关;位于上包迹线之上,为正相关,位于下包迹线以下,为负相关。 -
空间格局分析较为复杂,往往需要针对不同的问题假设选择不同的零模型[20]。为了认识人工林与天然林群落中活立木的空间分布状况以及样地内种间关联性,本研究选择最常用的零模型——完全空间随机模型(CSR),该模型主要是假设研究样地中各个体间不受其他生物或非生物的影响,相互独立,即研究个体在研究区域内的密度是恒定的,且个体分布符合泊松分布[21-24]。
-
所有数据处理分析、图形制作等均通过R语言实现,点格局分析采用“Spatstat”程序包完成。
-
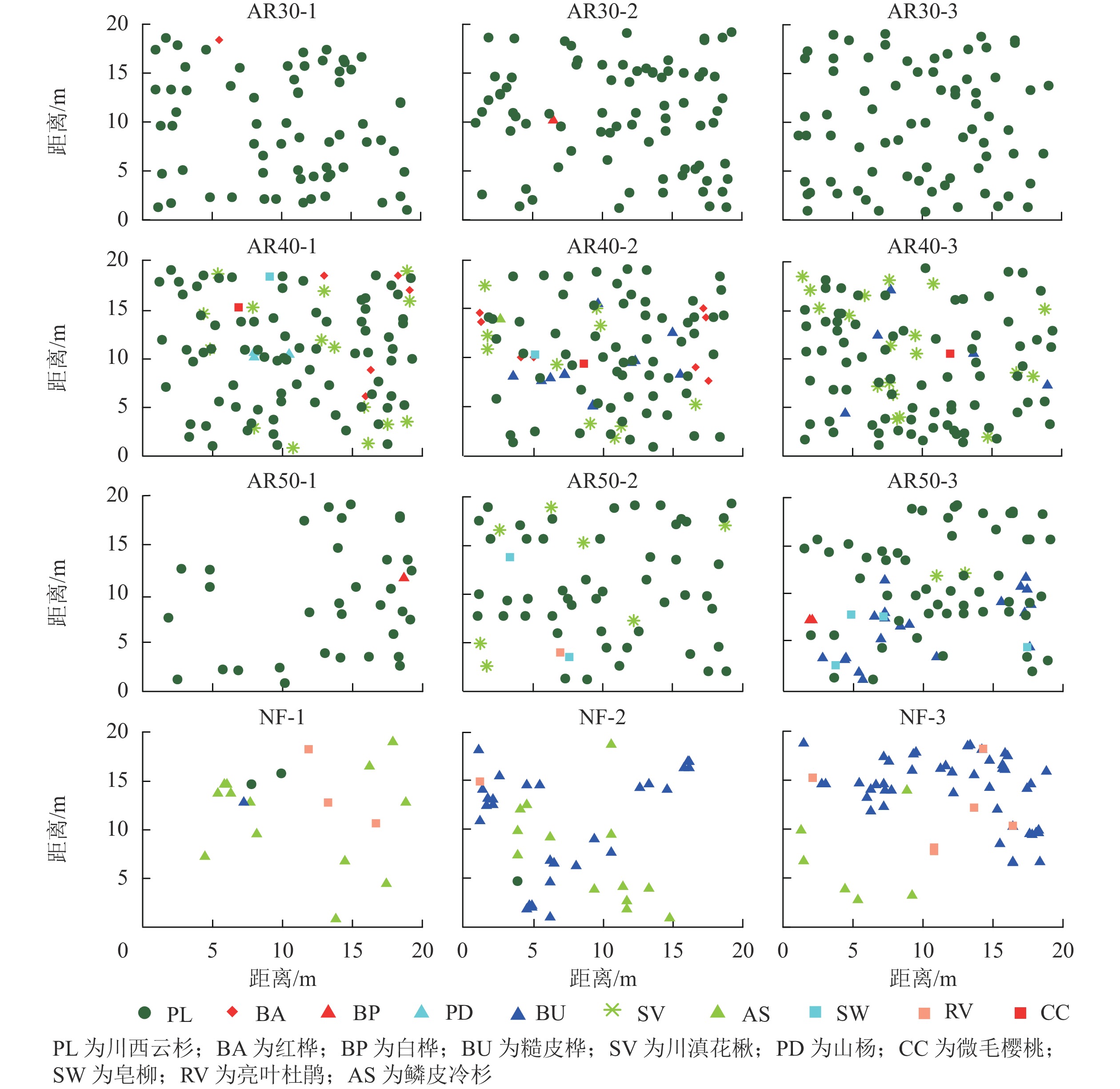
由表1可见:人工林群落主要有川西云杉、红桦、白桦、糙皮桦、川滇花楸、山杨、微毛樱桃和皂柳等树种组成,天然林群落主要有鳞皮冷杉、川西云杉、糙皮桦和亮叶杜鹃等树种组成,可见人工林与天然林群落树种组成存在差异。在人工林中林龄为30 a样地内所含树种最少,仅有3种,而林龄为40 a样地内所含树种最多,达8种。总体上,随着林龄的增加,人工林样地内树种数量呈先增加后减少的趋势,物种组成整体上逐渐丰富,川西云杉种群密度呈下降趋势。
分析人工林和天然林群落内优势树种的空间分布发现(图1):人工林与天然林群落树种组成和结构存在明显差异,天然林除了鳞皮冷杉和川西云杉外,其他阔叶树种的个体数量较多,人工林川西云杉种群密度明显高于天然林;在人工林样地中,川西云杉林分密度差异较大,林龄为30 a的人工林样地内川西云杉植株密度最大,林龄为50 a的人工林样地内川西云杉植株密度最小,表明随着林龄增加,开始出现林木自疏现象;同时,在人工林样地中,林龄为30 a的人工林群落内几乎没有其他树种分布,而林龄为40和50 a人工林群落内有大量其他阔叶树种分布,表明随着林龄增加,人工林内的树种多样性呈现增加趋势。
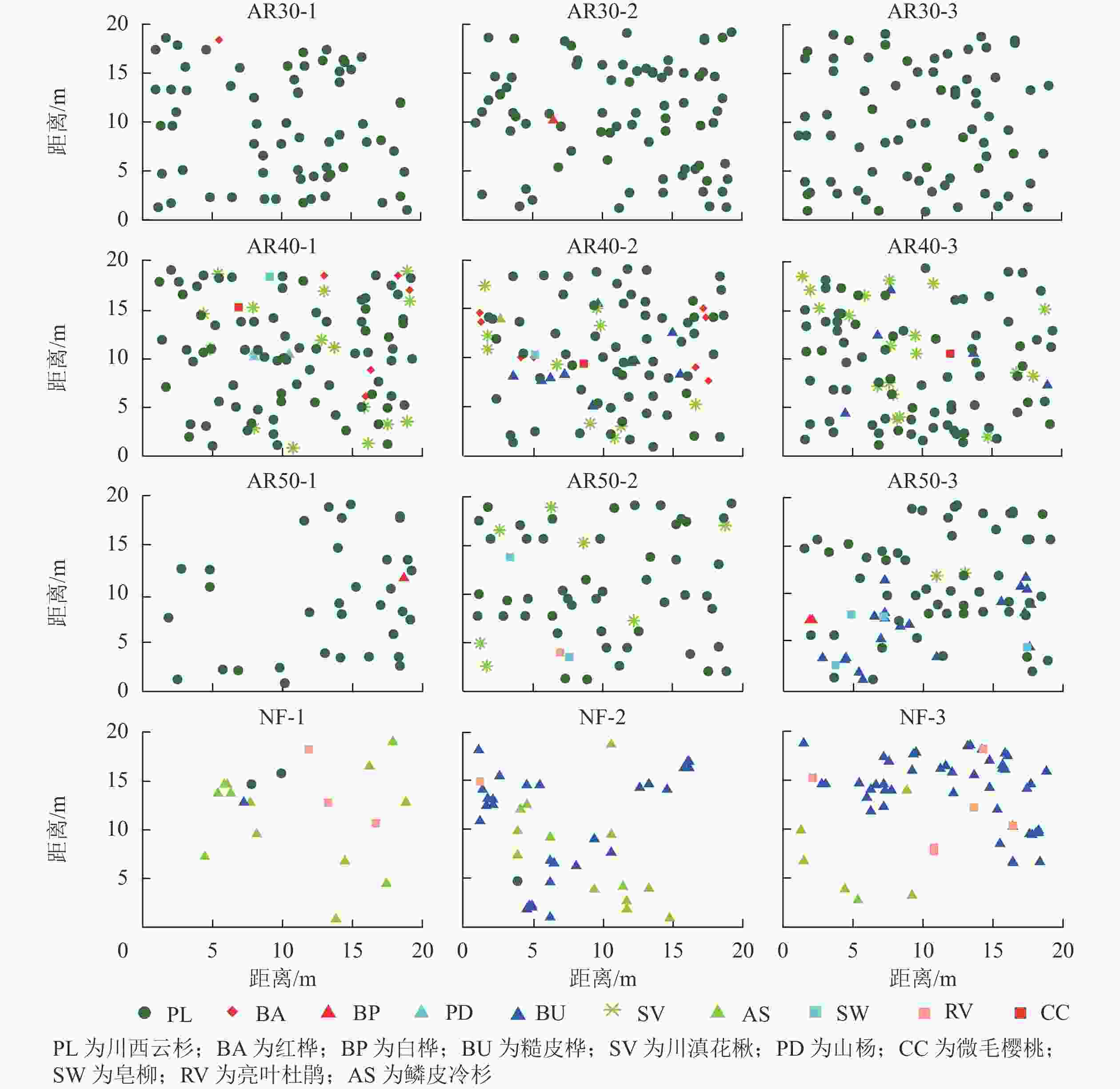
图 1 川西云杉人工林与天然林样地树种空间分布
Figure 1. Spatial distribution of tree species in each plot for plantations and natural forests P. likiangensis var. rubescens
-
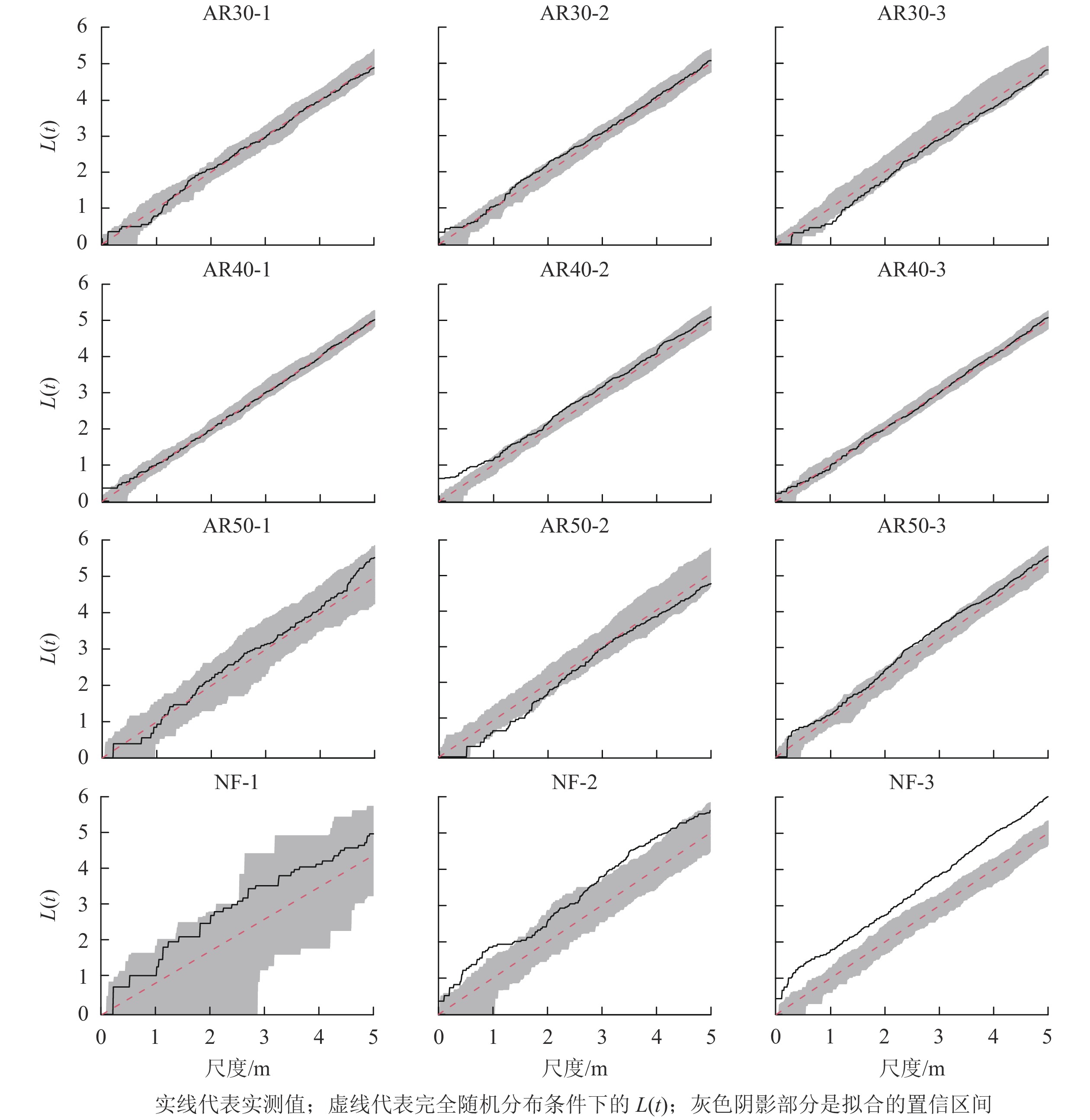
如图2所示:不同空间尺度下,川西云杉人工林各样地之间个体分布格局差异相对较小,林龄为30 a的人工林中活立木在所有空间尺度上均表现为随机分布格局,随机分布程度随着空间尺度增大而增强;林龄40 a的人工林中,AR40-1、AR40-3样地活立木在所有尺度上均为随机分布,AR40-2样地中活立木在0~1.0 m空间尺度内呈聚集分布,在大于1.0 m的空间范围内呈随机分布;林龄40 a的人工林中活立木在所有尺度上为随机分布。在天然林中,NF-1样地活立木在所有尺度上为随机分布,NF-2样地活立木在1.3~2.6 m范围内呈随机分布,在0~1.3 m和2.6~5.0 m空间尺度下为聚集分布,NF-3样地在研究范围内均为聚集分布。
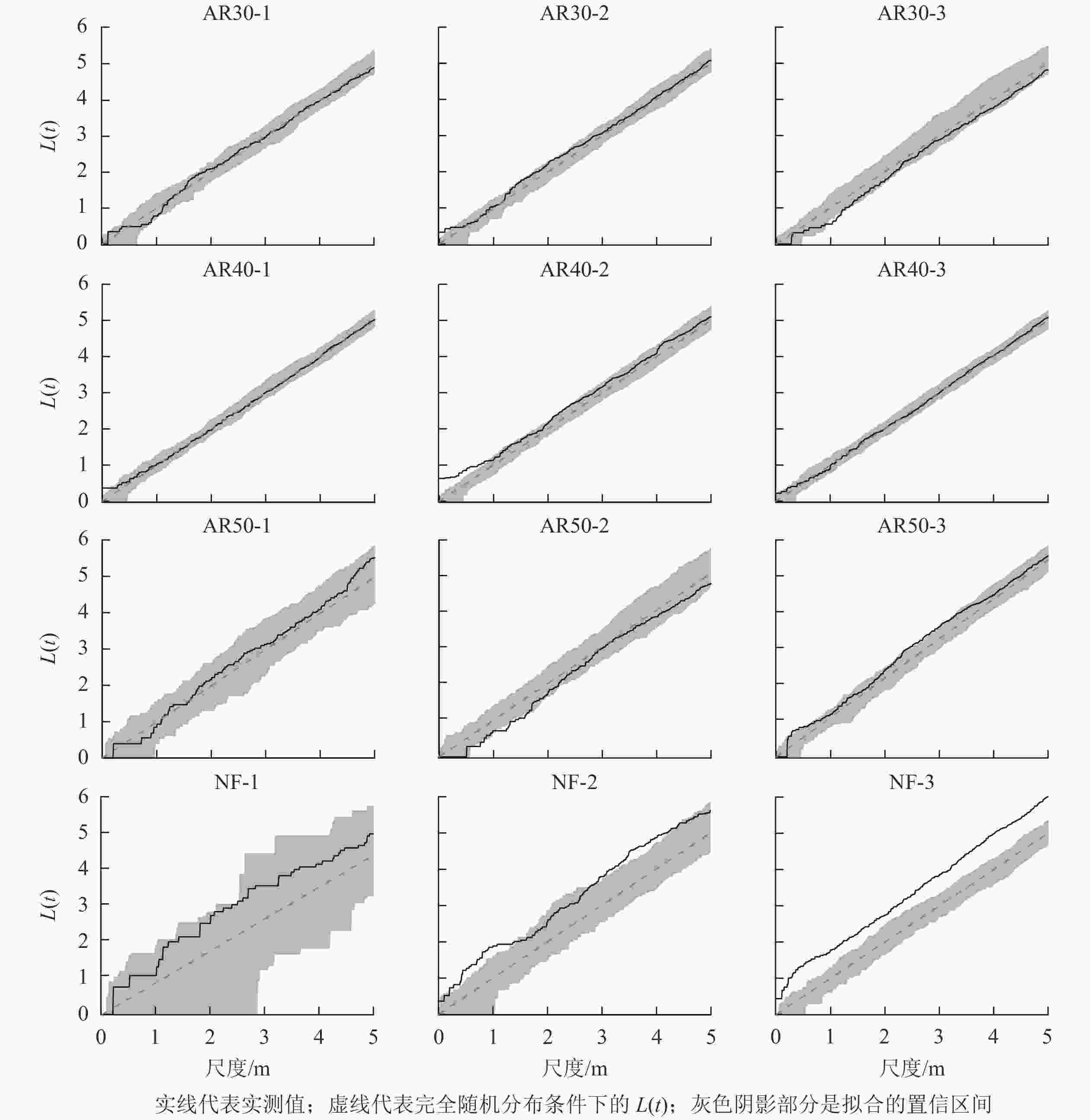
图 2 川西云杉人工林与天然林样地活立木的空间点格局分析
Figure 2. Point pattern analysis of standing trees in each plot for plantations and natural forests of P. likiangensis var. rubescens
-
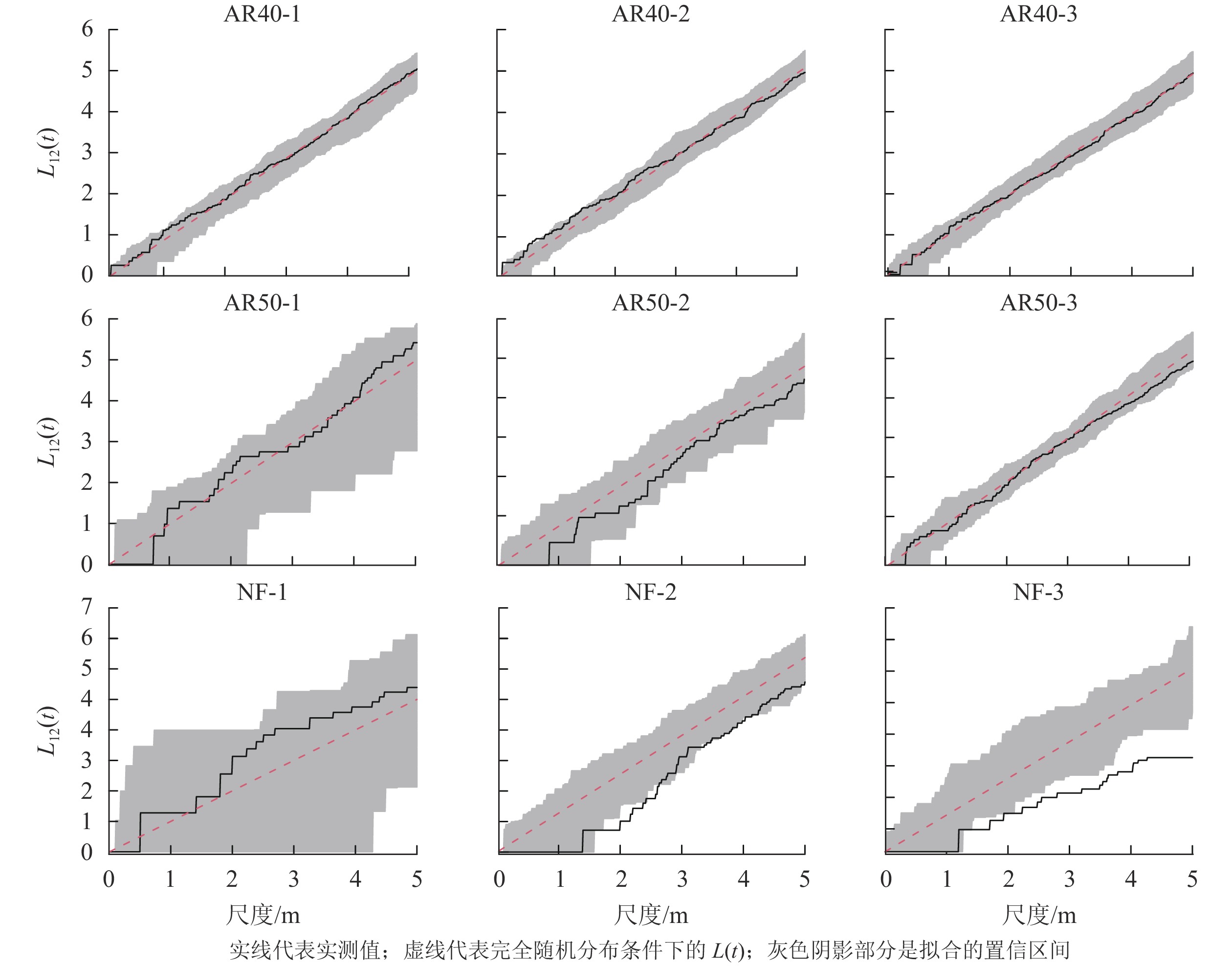
由于林龄为30 a的人工林以川西云杉为主,其他树种仅有2株,所以本研究中不分析AR30-1、AR30-2、AR30-3共3块样地物种间的空间关联性。对林龄为40和50 a的人工林以及天然林样地物种间的空间关联性进行分析(图3),结果表明:人工林和天然林群落树种空间关联性存在明显差异,所有人工林群落中川西云杉与其他树种间总体上表现为无空间关联性;而天然林样地中鳞皮冷杉与其他树种间的空间关联性变化较大,NF-1样地表现为无空间关联性,NF-2样地在1.8~2.6 m空间尺度上为负相关,其余尺度上为无空间关联性,NF-3样地除在0~2.1 m空间尺度上无空间关联外,其余尺度上均为显著的负相关,并随着空间尺度的增大,负相关程度逐渐增强。
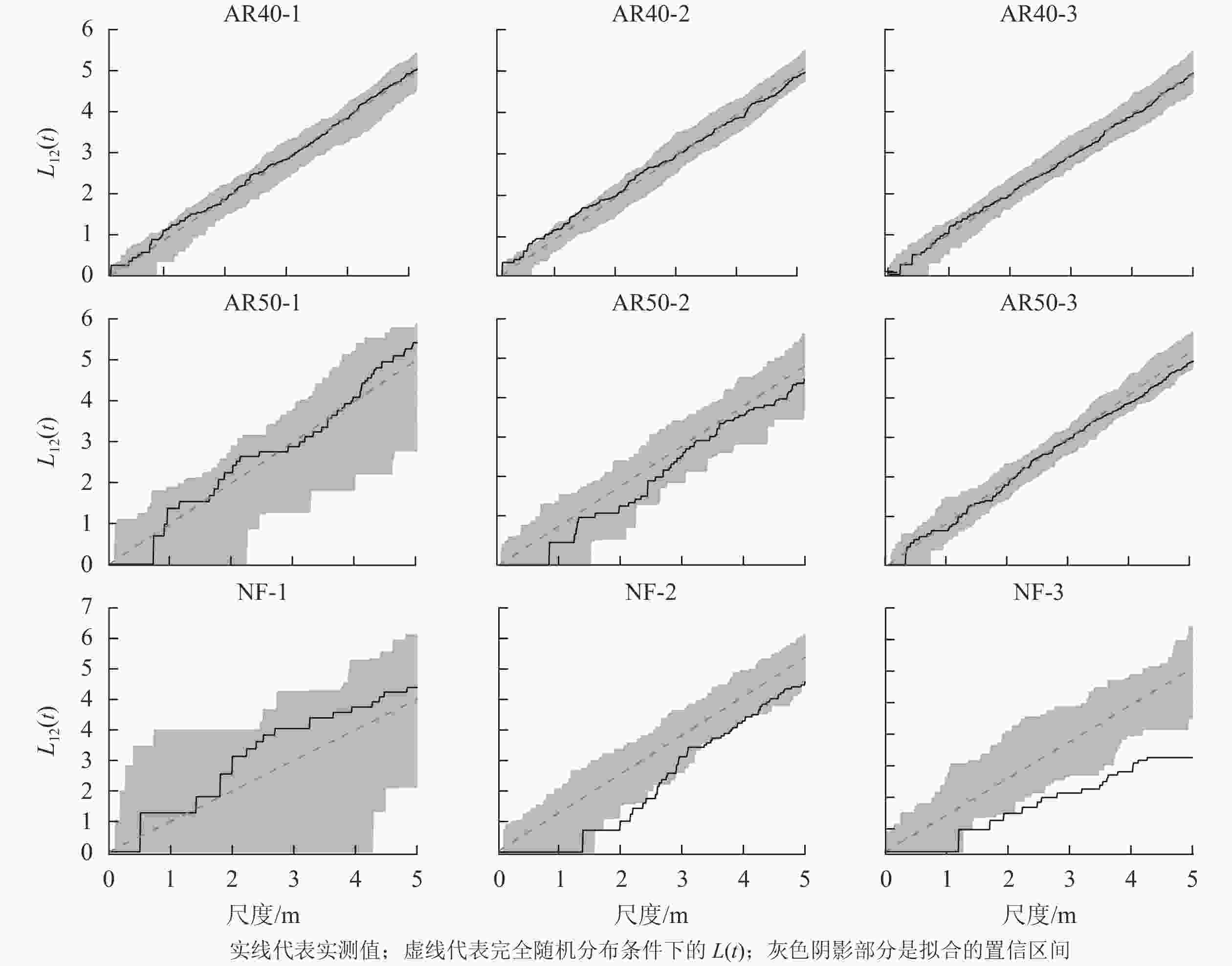
图 3 川西云杉人工林与天然林样地活立木的空间关联性
Figure 3. Spatial association of standing trees in each plot for plantations and natural forests of PL
-
植物种群的空间分布格局是自身的生物学特性、环境、植物种内和种间相互作用以及人为干扰的共同结果[25-27]。有研究[27-28]表明:种群空间格局的变化表现出种群适应新环境的生存策略及应对机制,利于种群更有效地获取资源。本研究分析了川西云杉人工林与天然林群落中活立木空间分布格局,除AR40-2样地在小尺度上呈聚集分布,大尺度上为随机分布外,其他人工林样地在所有研究尺度上均呈随机分布,而天然林群落空间格局总体上呈聚集分布。刘宪钊等[29]对华北落叶松Larix principis-rupprechtii人工林研究得出,群落空间点格局分布总体表现为小尺度上的聚集分布和中大尺度上的随机分布。闫海冰等[30]对华北山地典型次生林群落进行研究也得出,华北落叶松、云杉(白杄Picea meyeri和青杄Picea wilsonii)、桦木(白桦和红桦)、山杨天然次生林在小尺度上聚集分布,随尺度增加空间格局逐渐表现为随机分布。人工林与天然林群落空间格局的差异可能是因为林分起源不同,人工种植导致人工林具有较大导向性,林分密度和郁闭度均较大,林下更新相对较差,群落以均匀分布为主;而天然林以自然更新为主,受种子传播扩散限制,天然更新的幼树一般聚集生长于母树附近,故种子萌发不均匀可能导致活立木空间格局为聚集分布[26]。同时,树木呈聚集分布,在一定程度上能够相互庇护并以集群形式获取有利资源,提高其生存与竞争能力,有利于维持种群自身稳定[25]。本研究中川西云杉人工林随着恢复年份的增加,栽种的5年生幼苗在生长过程中不断天然更新与自然稀疏,一定程度上削弱了人为导向,同时树木个体对生存空间和养分的需求不断增长,越来越激烈的竞争使得弱势个体不断死亡、淘汰,导致个体越来越倾向于服从随机分布[27]。
种间关联性体现种群在群落中的相互关系,反映其争夺有利资源的能力[31]。在本研究中,人工林样地川西云杉与其他植物间以不相关为主,物种间不存在竞争资源的关系。这可能是人工林中川西云杉种群密度大,其他树种通过种子扩散进入人工林中,多分布在林间空地,因而种间竞争在群落动态中的作用微弱。该区域在前期森林砍伐时,保留的大量腐木、枝丫、叶和树根等腐烂分解后进入土壤,补充大量的营养元素,使人工林保持了可持续利用的资源,种间竞争关系相对弱。天然林样地鳞皮冷杉与其他植物在小尺度上不相关,大尺度上为负相关,这表明天然林树种间在资源及生存空间的争夺中存在较为激烈的竞争关系,这可能由于天然林中物种呈聚集分布,对资源利用的程度较高,树种间竞争关系强烈。
自从实施退耕还林工程和天然林资源保护工程以来,川西高原人工林面积不断增加,已经成为该区森林资源的重要组成部分,在改善生态环境、调节气候和保护生物多样性等方面扮演着重要角色[15]。然而,目前人工林面临着质量差、林分结构不合理和树种单一等生态问题[32-33]。本研究发现:随着林龄的增加,人工林群落内川西云杉种群密度呈下降趋势,周围天然林中的阔叶树种不断扩散到人工林中,物种组成逐渐丰富,川西云杉与其他阔叶树种无空间关联性,表明了人工林正逐渐向天然林群落演替,但川西云杉人工林与天然林仍然存在一定的差异。有研究[34-35]显示:间伐能够促进云杉人工林的生长、林下更新,提高幼树和幼苗的成活率。针对人工林中活立木种群随机分布和种间无关联性的特性,可以适当间伐一些生长较差的川西云杉植株,为其他阔叶树种进入人工林群落内提供生存空间。同时根据刘世荣等[31]提出的近自然改造的方式,适当种植天然林存在的主要物种,将人工针叶纯林转化为以乡土树种为主的混交林,并抚育人工林中已存在的阔叶树种,丰富物种组成,改善人工林群落结构,从而加快人工林向天然林群落演替进程。
Spatial distribution pattern and interspecific correlation between plantation and natural forest of Picea likiangensis var. rubescens in western Sichuan, China
-
摘要:
目的 随着国家退耕还林工程的实施,川西云杉Picea likiangensis var. rubescens人工林面积和蓄积量不断增长,已成为川西地区森林资源的重要组成部分。本研究分析道孚县川西云杉人工林与天然林活立木空间分布格局特征及种间关联性差异,以揭示人工林群落动态变化及演替规律。 方法 采用典型样方法设置12块固定样地,测量每株活立木中心位置坐标、胸径、树高和冠幅等,运用点格局分析中的单变量Ripley’s L(t)函数和双变量Ripley’s L12(t)函数分别分析人工林与天然林群落的空间分布格局和群落中树种间的空间关联性。 结果 ①随着恢复年限的增加,人工林群落内川西云杉种群密度呈下降趋势,而群落物种数逐渐增加;②人工林群落中活立木空间分布格局总体上表现为随机分布,而天然林内活立木空间分布以聚集分布为主;③人工林中川西云杉与其他阔叶树种无空间关联性,而天然林群落内主要树种间关联性在小尺度范围上呈不相关,在大尺度上表现为负相关。 结论 经过50 a人工恢复后,川西云杉人工林与天然林在群落空间分布格局和种间关联性方面仍然存在一定的差异,生态恢复进程较慢。建议采取适当间伐与种植天然林优势树种相结合的方式,优化改造人工林结构,从而加快人工林向天然林群落演替进程。图3表1参35 Abstract:Objective As an important component of forest resources in western Sichuan, the area and standing volume of Picea likiangensis var. rubescens (PL) plantations are increasing with the implementation of the national project Converting Farmland to Forest. This study aims to analyze the spatial distribution pattern and interspecific correlation between natural forest and plantation in Daofu County, so as to reveal community dynamic change and succession law of plantation community. Method The typical sampling method used to set up 12 fixed plots to measure the central position coordinates, diameter at breast height (DBH), height and crown width of each living tree. The univariate Ripley’s L(t) function and bivariate Ripley’s L12(t) function in point pattern analysis were used to analyze the spatial distribution pattern of plantation and natural forest colony, and the spatial correlation between tree species in the community. Result (1)With the increase of restoration years, the population density of PL gradually decreased, while the number of community species gradually increased. (2)The spatial distribution pattern of living trees was generally random in plantations but was aggregated in natural forests. (3)The spatial correlation between PL and other broad-leaved tree species was not significant in plantations, while the main tree species in the natural forest community were not correlated on a small scale but negatively correlated on a large scale. Conclusion After 50 years of artificial restoration, there are still distinct differences between PL plantations and natural forests in community spatial distribution pattern and interspecific correlation, and the process of ecological restoration is slow. It is suggested that the structure of plantations could be optimized and improved by combining thinning measures and planting dominant tree species of natural forests, so as to accelerate the succession process from plantations to natural forests. [Ch, 3 fig. 1 tab. 35 ref.] -
图 1 川西云杉人工林与天然林样地树种空间分布
Figure 1 Spatial distribution of tree species in each plot for plantations and natural forests P. likiangensis var. rubescens
图 2 川西云杉人工林与天然林样地活立木的空间点格局分析
Figure 2 Point pattern analysis of standing trees in each plot for plantations and natural forests of P. likiangensis var. rubescens
图 3 川西云杉人工林与天然林样地活立木的空间关联性
Figure 3 Spatial association of standing trees in each plot for plantations and natural forests of PL
表 1 川西云杉人工林与天然林样地基本信息
Table 1. General information of each plot for plantation and natural forests of P. likiangensis var. rubescens
样地号 类型 林龄/a 林分密度/
(株·hm−2)云杉种群密度/
(株·hm−2)海拔/m 坡向 坡度/(°) 郁闭度 树种 AR30-1 人工林 30 1 650 1 625 3 688 东北 23 0.70 PL、BA、BP AR30-2 人工林 30 2 125 2 100 3 702 东北 26 0.70 AR30-3 人工林 30 1 825 1 825 3 692 东北 25 0.60 AR40-1 人工林 40 2 500 1 800 3 760 东北 38 0.80 PL、BU、SV、PD、CC、BA、SW、AS AR40-2 人工林 40 2 625 1 625 3 756 东北 37 0.85 AR40-3 人工林 40 2 600 1 925 3 728 东北 37 0.85 AR50-1 人工林 50 1 025 825 3 810 东北 30 0.65 PL、BP、BU、SV、SW、RV AR50-2 人工林 50 1 425 1 175 3 790 东北 36 0.80 AR50-3 人工林 50 2 025 1 325 3 770 东北 38 0.80 NF-1 天然林 >150 475 50 3 840 东北 6 0.45 PL、BU、RV、AS NF-2 天然林 >150 1 100 25 3 842 东北 8 0.50 NF-3 天然林 >150 1 600 25 3 845 东北 10 0.40 说明:PL为川西云杉,BA为红桦Betula albosinensis,BP为白桦B. platyphylla,BU为糙皮桦B. utilis,SV为川滇花楸Sorbus vilmorinii,PD为山杨Populus davidiana,CC为微毛樱桃Cerasus clarofolia,SW为皂柳Salix wallichiana,RV为亮叶杜鹃 Rhododendron vernicosum,AS为鳞皮冷杉Abies squamata  下载: 导出CSV
下载: 导出CSV
-
[1] 张金屯, 孟东平. 芦芽山华北落叶松林不同龄级立木的点格局分析[J]. 生态学报, 2004, 24(1): 35 − 40. ZHANG Jintun, MENG Dongping. Spatial pattern analysis of individuals in different age-classes of Larix principis-rupprechtii in Luya mountain reserve, Shanxi, China [J]. Acta Ecol Sin, 2004, 24(1): 35 − 40. [2] 许爱云, 许冬梅, 刘金龙, 等. 基于零模型的宁夏荒漠草原优势种群点格局分析[J]. 生态学报, 2020, 40(12): 4180 − 4187. XU Aiyun, XU Dongmei, LIU Jinlong, et al. Point pattern analysis of dominant populations based on null models in desert steppe in Ningxia [J]. Acta Ecol Sin, 2020, 40(12): 4180 − 4187. [3] 孙伟中, 赵士洞. 长白山北坡椴树阔叶红松林群落主要树种分布格局的研究[J]. 应用生态学报, 1997, 8(2): 119 − 122. SUN Weizhong, ZHAO Shidong. Distribution patterns of main tree species in Tilia broadleaf Korean pine forest on northern slope of Changbai Mountain [J]. Chin J Appl Ecol, 1997, 8(2): 119 − 122. [4] 池森, 王从军, 黎庆菊, 等. 喀斯特次生林幼树更新空间分布格局及种间关联性[J]. 应用生态学报, 2020, 31(12): 3989 − 3996. CHI Sen, WANG Congjun, LI Qingju, et al. Spatial distribution and interspecific associations of regenerating saplings in karst secondary forests [J]. Chin J Appl Ecol, 2020, 31(12): 3989 − 3996. [5] 刘珏宏, 高慧, 张丽红, 等. 内蒙古锡林郭勒草原大针茅-克氏针茅群落的种间关联特征分析[J]. 植物生态学报, 2010, 34(9): 1016 − 1024. LIU Juehong, GAO Hui, ZHANG Lihong, et al. Comparative analysis of interspecific association within the Stipa grandis-S. krylovii community in typical steppe of Inner Mongolia, China [J]. Chin J Plant Ecol, 2010, 34(9): 1016 − 1024. [6] 刘淑燕, 岳永杰, 余新晓, 等. 北京山区刺槐林种群的空间点格局[J]. 东北林业大学学报, 2010, 38(4): 33 − 36. LIU Shuyan, YUE Yongjie, YU Xinxiao, et al. Spatial pattern of Robinia pseudoacacia plantation population in mountainous area of Beijing [J]. J Northeast For Univ, 2010, 38(4): 33 − 36. [7] 张金屯. 植物种群空间分布的点格局分析[J]. 植物生态学报, 1998, 22(4): 57 − 62. ZHANG Jintun. Analysis of spatial point pattern for plant species [J]. Chin J Plant Ecol, 1998, 22(4): 57 − 62. [8] RIPLEY B D. Spatial Statistics[M]. New York: Wiley, 1981. [9] LOOSMORE N B, FORD E D. Statistical inference using the G or K point pattern spatial statistics [J]. Ecology, 2006, 87(8): 1925 − 1931. [10] CHAKRABORTY A, GELF A E, WILSON A M, et al. Point pattern model ling forde graded presence-only dataover large regions [J]. J Royal Stat Soc Ser C, 2011, 60(5): 757 − 776. [11] 樊登星, 余新晓. 北京山区栓皮栎林优势种群点格局分析[J]. 生态学报, 2016, 36(2): 318 − 325. FAN Dengxing, YU Xinxiao. Spatial point pattern analysis of Quercus variabilis and Pinus tabulaeformis populations in a mountainous area of Beijing [J]. Acta Ecol Sin, 2016, 36(2): 318 − 325. [12] 申静霞, 袁秀锦, 李迈和, 等. 土壤温度和水分变化对川西云杉幼苗氮和磷含量的影响[J]. 林业科学, 2019, 55(4): 31 − 41. SHEN Jingxia, YUAN Xiujin, LI Maihe, et al. Effects of soil temperature and moisture on nitrogen and phosphorus contents in Picea balfouriana seedlings [J]. Sci Silv Sin, 2019, 55(4): 31 − 41. [13] 刘庆, 吴彦, 何海. 中国西南亚高山针叶林的生态学问题[J]. 世界科技研究与发展, 2001, 23(2): 63 − 69. LIU Qing, WU Yan, HE Hai. Ecological problems of subalpine coniferous forest in the southwest of China [J]. World Sci-Tech R&D, 2001, 23(2): 63 − 69. [14] 刘兴良, 宿以明, 刘世荣, 等. 川西高山林区人工林生态学的研究——人工林分区与分类[J]. 四川林业科技, 2004(1): 1 − 9. LIU Xingliang, SU Yiming, LIU Shirong, et al. Studies of ecology of plantations in alpine forest areas of western Sichuan: division and classification of plantations [J]. J Sichuan For Sci Technol, 2004(1): 1 − 9. [15] 马姜明, 刘世荣, 史作明, 等. 川西亚高山暗针叶林恢复过程中群落物种组成和多样性的变化[J]. 林业科学, 2007, 47(5): 20 − 26. MA Jiangming, LIU Shirong, SHI Zuoming, et al. Changes of species composition and diversity in the restoration process of sub-alpine dark brown coniferous forests in Western Sichuan, China [J]. Sci Silv Sin, 2007, 47(5): 20 − 26. [16] 冯秋红, 王毅, 刘兴良, 等. 川西亚高山不同林龄云杉人工林林地水源涵养能力比较研究[J]. 四川林业科技, 2004, 41(1): 5 − 10. FENG Qiuhong, WANG Yi, LIU Xingliang, et al. Comparison on ground cover and soil hydrological function of Picea asperata Mast plantation at different ages in the subalpine region of western Sichuan [J]. J Sichuan For Sci Technol, 2004, 41(1): 5 − 10. [17] 庞学勇, 刘庆, 刘世全, 等. 川西亚高山云杉人工林土壤质量性状演变[J]. 生态学报, 2004, 24(2): 261 − 267. PANG Xueyong, LIU Qing, LIU Shirong, et al. Changes of soil fertility quality properties under subalpine spruce plantation in Western Sichuan [J]. Acta Ecol Sin, 2004, 24(2): 261 − 267. [18] RIPLEY B D. Modelling spatial patterns [J]. J Royal Stat Soc Ser B Methodol, 1977, 39(2): 172 − 212. [19] DIGGLE P J. Statistical Analysis of Spatial Point Patterns[M]. London: Academic Press, 1983: 140−15. [20] 王鑫厅, 侯亚丽, 刘芳, 等. 羊草+大针茅草原退化群落优势种群空间点格局分析[J]. 植物生态学报, 2011, 35(12): 1281 − 1289. WABG Xinting, HOU Yali, LIU Fang, et al. Point pattern analysis of dominant populations in a degraded community in Leymus chinensis﹢Stipa grandis steppe in Inner Mongolia, China [J]. Chin J Plant Ecol, 2011, 35(12): 1281 − 1289. [21] 梁爽, 许涵, 林家怡, 等. 尖峰岭热带山地雨林优势树种白颜树空间分布格局[J]. 植物生态学报, 2014, 38(12): 1273 − 1282. LIANG Shuang, XU Han, LIN Jiayi, et al. Spatial distribution pattern of the dominant species Gironniera subaequalis in tropical montane rainforest of Jianfengling, Hainan Ialand, China [J]. Chin J Plant Ecol, 2014, 38(12): 1273 − 1282. [22] MOLONEY K A, WIEGAND T, et al. Handbook of Spatial Point-pattern Analysis in Ecology[M]. Boca Ration: CRC Press, 2013: 715﹣715. [23] 高福元, 石福习. 基于不同零模型的三江平原沼泽湿地主要物种小尺度点格局分析[J]. 生态学报, 2015, 35(7): 2029 − 2037. GAO Fuyuan, SHI Fuxi. Small-scale point pattern analysis based on different null models for detecting spatial patterns of dominant spacies in Sanjiang Plain, China [J]. Acta Ecol Sin, 2015, 35(7): 2029 − 2037. [24] 沈志强, 华敏, 丹曲, 等. 藏东南川滇高山栎种群不同生长阶段的空间格局与关联性[J]. 应用生态学报, 2016, 27(2): 387 − 394. SHEN Zhiqiang, HUA Min, DAN Qu, et al. Spatial pattern analysis and associations of Quercus aquifolioides population at different growth stages in southeast Tibet, China [J]. Chin J Appl Ecol, 2016, 27(2): 387 − 394. [25] 刘旻霞. 亚高寒草甸不同坡向金露梅种群的空间分布格局及空间关联[J]. 应用生态学报, 2017, 28(6): 1817 − 1823. LIU Minxia. Spatial distribution and spatial association of Potentilla fruticosa populations on different slope aspects in subalpine meadow [J]. Chin J Appl Ecol, 2017, 28(6): 1817 − 1823. [26] 李宪, 廖良宁, 杨海鹏, 等. 杉木与火力楠人工混交林群落空间格局研究[J]. 西南林业大学学报(自然科学), 2020, 40(5): 1 − 9. LI Xian, LIAO Liangning, YANG Haipeng, et al. Spatial pattern of artificial mixed forest of Cunninghamia lanceolata and Michelia macclurei [J]. J Westsouth For Univ Nat Sci Ed, 2020, 40(5): 1 − 9. [27] 杨君珑, 王辉, 王彬, 等. 子午岭油松林灌木层主要树种的空间分布格局和种间关联性研究[J]. 西北植物学报, 2007, 27(4): 4791 − 4796. YANG Junlong, WANG Hui, WANG Bing, et al. Spatial distribution pattern and interspecific association of main tree spacies in Pinus tabuliformis forest in Ziwuling Mountains [J]. Acta Bot Boreal-Occident Sin, 2007, 27(4): 4791 − 4796. [28] 胡文强, 黄世能, 李家湘, 等. 南岭石坑崆山顶矮林乔木优势种群的种间关联性[J]. 生态学杂志, 2013, 32(10): 2665 − 2671. HU Wenqiang, HUANG Shineng, LI Jiaxiang, et al. Interspecific associations among dominant tree populations in an elfin forest community in Shikengkong of Nanling Mountains, China [J]. Chin J Ecol, 2013, 32(10): 2665 − 2671. [29] 刘宪钊, 李卫珍, 王金龙, 等. 2种不同起源华北落叶松林空间点格局及植物多样性[J]. 南京林业大学学报(自然科学版), 2017, 41(6): 102 − 108. LIU Xianzhao, LI Weizhen, WANG Jinlong, et al. Spatial distributions and species diversity of two types of Larix principis-rupprechtii [J]. J Nanjing For Univ Nat Sci Ed, 2017, 41(6): 102 − 108. [30] 闫海冰, 韩有志, 杨秀清, 等. 华北山地典型天然次生林群落的树种空间分布格局及其关联性[J]. 生态学报, 2010, 30(9): 2311 − 2321. YAN Haibing, HAN Youzhi, YANG Xiuqing, et al. Spatial distributions patterns and associations of tree species in typical natural secondary mountain forest communities of Northern China [J]. Acta Ecol Sin, 2010, 30(9): 2311 − 2321. [31] 刘世荣, 杨予静, 王晖. 中国人工林经营发展战略与对策: 从追求木材产量的单一目标经营转向提升生态系统服务质量和效益的多目标经营[J]. 生态学报, 2018, 38(1): 1 − 10. LIU Shirong, YANG Yujing, WANG Hui. Development strategy and management countermeasures of planted forests in China: transforming from timber-centered single objective management towards multi-purpose management for enhancing quality and benefits of ecosystem services [J]. Acta Ecol Sin, 2018, 38(1): 1 − 10. [32] YANG Jida, ZHANG Zhiming, ZHAXI Dawa, et al. Spatial distribution patterns and intra-specific competition of pine (Pinus yunnanensis) in abandoned farmland under the Sloping Land Conservation Program [J]. Ecol Eng, 2019, 135: 17 − 27. [33] 秦燕燕, 蒋斌, 曹秀文, 等. 间伐强度对白龙江林区云杉人工林植物多样性及其更新的影响[J]. 草业科学, 2014, 31(4): 599 − 606. QIN Yanyan, JIANG Bin, CAO Xiuwen, et al. Effects of thinning intensity on diversity of undergrowth vegetation and regeneration in artificial spruce forest [J]. Pratacult Sci, 2014, 31(4): 599 − 606. [34] 幸福, 包维楷, 庞学勇, 等. 云杉人工纯林中树木个体径向生长过程及林窗疏伐后的释放效应[J]. 应用与环境生物学报, 2013, 19(2): 262 − 271. XING Fu, BAO Weikai, PANG Xueyong, et al. Radial growth process of Picea asperata and its response to gap model thinning within the single spruce plantation at the western Sichuan, China [J]. Chin J Appl Environ Biol, 2013, 19(2): 262 − 271. [35] QI Lin, YANG Jian, YU Dapao, et al. Responses of regeneration and species coexistence to single-tree selective logging for a temperate mixed forest in eastern Eurasia [J]. Ann For Sci, 2016, 73: 449 − 460. -

-
链接本文:
https://zlxb.zafu.edu.cn/article/doi/10.11833/j.issn.2095-0756.20210433
 点击查看大图
点击查看大图
计量
- 文章访问数: 2009
- HTML全文浏览量: 727
- PDF下载量: 113
- 被引次数: 0












