







-
森林在陆地生态系统中发挥着重要的碳汇作用。森林生物量的空间精准量化对了解陆地碳储量、碳收支、碳平衡,以及它们与全球气候变化之间的关系具有重要意义[1-3]。森林地上生物量(AGB)为林木干、枝、叶的有机体干质量,不含林木根部、灌木、草本及林下枯枝落叶,是森林生物量的主体部分。近年来,采用遥感技术反演森林AGB展露出巨大的潜力,光学、激光雷达(LiDAR)、被动微波雷达和合成孔径雷达(SAR)等多种遥感数据用于森林AGB的估测,使得森林AGB估测的区域尺度性、经济性及精度等不断提升[3-7]。其中,SAR不受天时、天气的影响,且微波波长较长,在森林中具有较强的穿透能力,因此在获取森林密度、树高、AGB等森林垂直结构因子方面极具潜力[2, 8-10]。SAR估测森林AGB的能力有赖于其工作频率的高低,大量研究证实了低频波段后向散射对森林AGB变化的敏感性更强。在常用于森林监测的微波波段中,频率较低的P波段SAR后向散射系数对森林AGB变化最为敏感[11-13]。目前P波段的研究多基于机载数据展开,且研究区多位于国外,而国内相关研究则开展较少。CARTUS等[1]基于AfriSAR、BioSAR和TropiSAR机载飞行试验,使用P、C、L波段联合进行了森林AGB反演,结果表明P波段与森林结构因子的树干、树枝的相关性较高,P波段的加入有力地提高了反演的准确度和精度。LIAO等[14]使用TropiSAR机载P波段数据,采用层析方法(TomoSAR)将相干幅度、干涉相位和后向散射特征建模对法属圭亚那热带雨林AGB进行了反演,结果表明树高特征的引入可有效提高森林AGB反演精度,决定系数(R2)最高可达0.7。冯琦等[10]使用国产机载P波段数据结合坡度因子,在考虑当地入射角和坡度的情况下建立对数统计模型,对内蒙古根河市生态站的寒温带针叶林进行森林AGB的反演,最高反演精度R2为0.634、均方根误差(RMSE)为12.07 t·hm−2。
现有采用P波段SAR数据进行的森林AGB估测,多集中在采用P波段的后向散射信息、相位和相干性信息,而对于极化信息的利用则较少。由于SAR信息的差异,使得其用于森林AGB估测的方法也差异明显,如采用后向散射信息估测森林地上生物量,多采用线性回归参数模型;而采用相位和相干性信息利用层析技术进行森林AGB估测则多基于植被微波散射模型或电磁波信号模型进行反演。线性回归模型简单灵活,但通常无法表征P波段特征与森林AGB变化之间的复杂关系;植被散射模型或者电磁波信号模型能够体现森林与P波段电磁波之间的部分物理作用机制,但模型较复杂,应用推广困难。
近年来,随着极化SAR数据的丰富,可提取的极化SAR特征涌现,非参数模型被广泛应用于SAR极化特征农作物生长参数的定量反演,并表现出较强的反演能力和较好的应用推广性。鉴于P在森林监测中的潜力,其极化特征在森林AGB估测中并未深入探索,参数模型过于简单、植被微波散射模型理解过于困难,本研究以中国北方典型寒温带森林作为研究对象,使用机载P波段SAR数据,提取多种极化特征,在分析其P波段极化散射特征的基础上,探索采用参数和非参数模型进行森林AGB估测的可行性,旨在明确P波段森林的极化散射特征,探索采用P波段极化SAR数据进行森林AGB估测具体应用的有效方法。
-
研究区为位于内蒙古呼伦贝尔盟的根河市大兴安岭森林生态系统国家野外科学观测研究站(大兴安岭生态站),50°49′~50°51′N,121°30′~121°31′E。该研究站是中国纬度最高的森林生态研究站,面积为102 km2。研究区气候类型为典型的寒温带大陆性季风气候。研究区地势相对平缓,区域内80%的坡度小于15°,海拔高度分布在800~1 200 m,森林覆盖率大于75%,主要树种为兴安落叶松Larix gmelinii、白桦Betula platyphylla、樟子松Pinus sylvestris var. mongolica等。
-
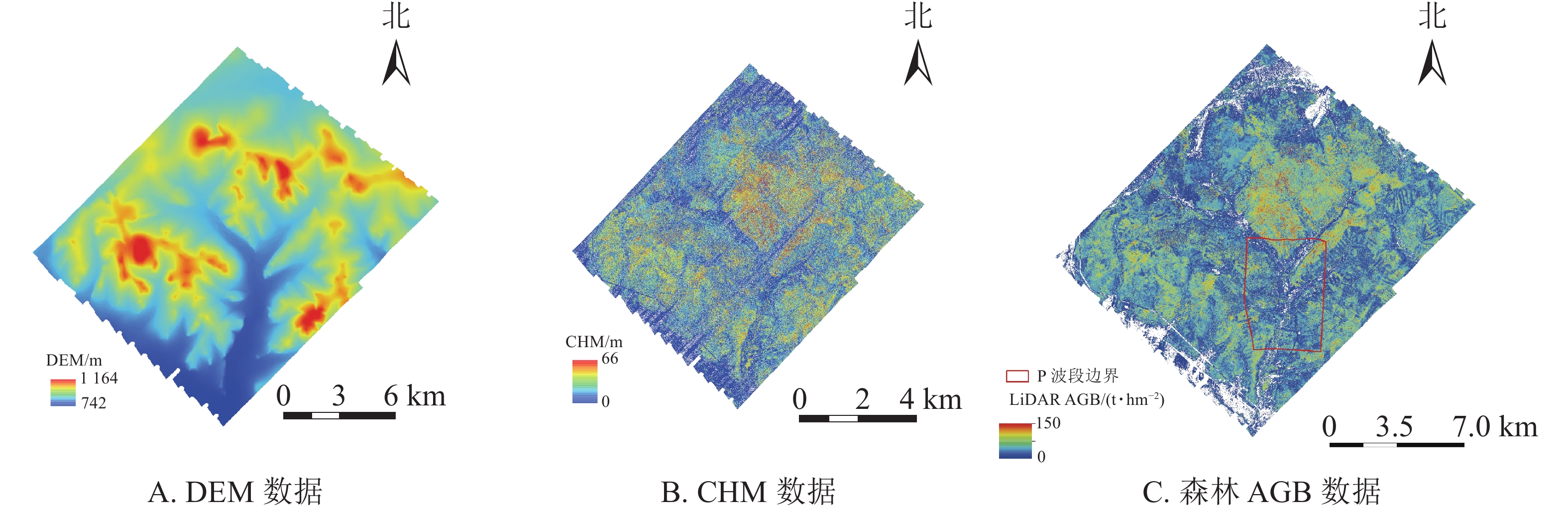
采用的P波段SAR数据由机载CAMSAR系统获得[15]。通过协议方式获取根河实验区机载P波段全极化(HH/HV/VV/VH) SLC数据。该数据是2013年9月以“奖状Ⅱ”飞机平台,以CASMSAR系统右视观测获取的全极化SAR数据。获取过程中飞行方向为自西向东,飞行高度为5 807 m,获取时间为2013年9月13—16日,极化方式为HH/HV/VH/VV,产品模式为SLC,幅宽为6 km × 7 km,中心入射角为55.058°,距离向分辨率为0.666 m,方位向分辨率为0.625 m。机载P波段SAR数据的预处理关键步骤包括多视处理、正射校正、入射角校正和极化方位角校正。多视视数在距离向和方位向均为3,数据的正射校正、入射校正参考文献[10];极化方位角校正过程和算法见文献[16]。P波段SAR数据预处理结果、森林AGB抽样点及林分概况见图1。
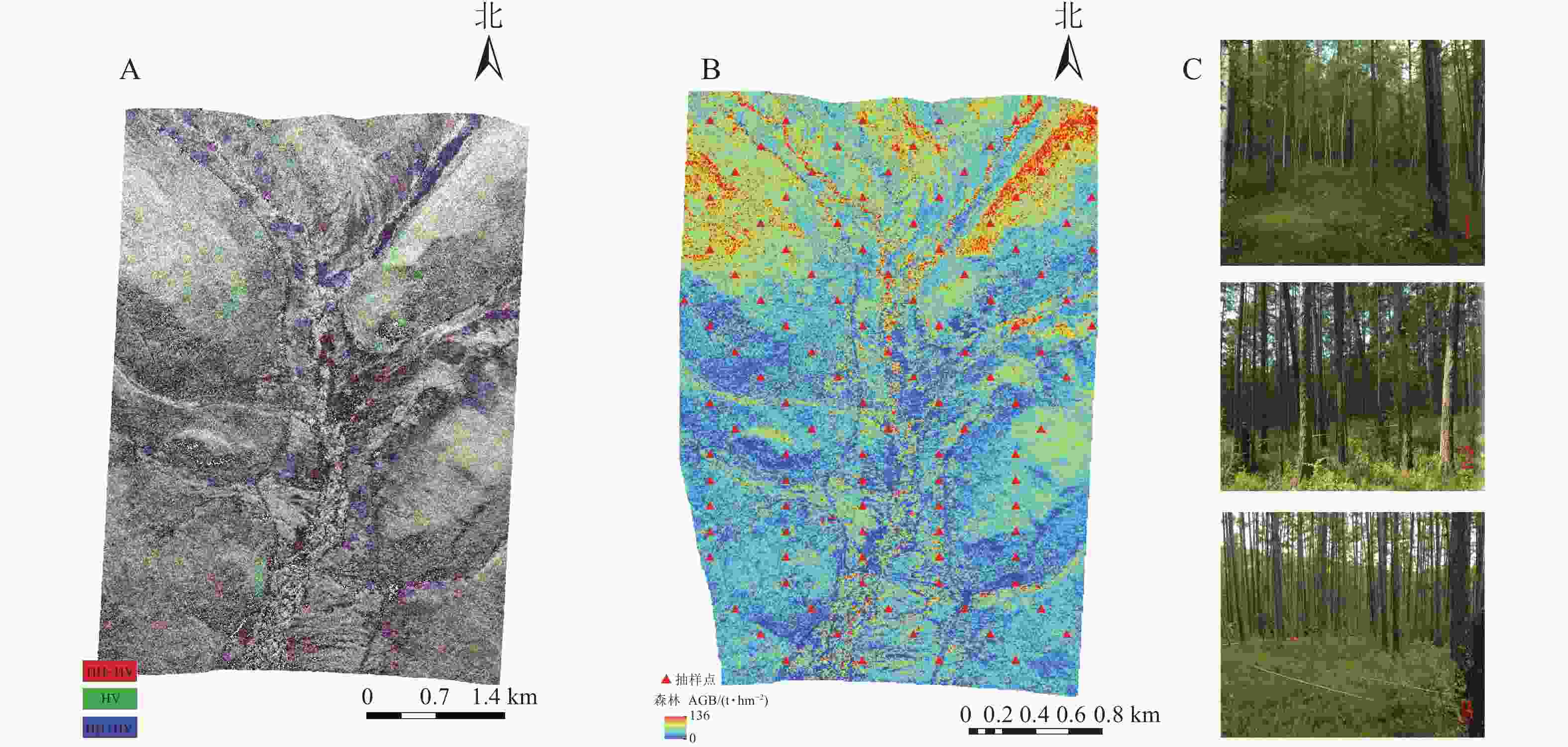
图 1 P波段SAR数据(A)及其覆盖区样点分布(B)、林分概况(C)
Figure 1. P band SAR data (A), the distribution of samples (B) and stand examples (C)
-
LiDAR获取的数字表面模型(DSM)、数字高程模型(DEM)用于P波段SAR数据的地理编码,由冠层高度模型(CHM)获取的LiDAR森林AGB数据用于反演模型的训练及验证。本研究获取的机载LiDAR数据是将Leica机载雷达系统荷载于“运-5”飞机平台上,于2012年8—9月在根河实验区开展飞行任务。该原始数据密度为5.6个·m−2的点云数据,激光中心波谱值为1550 nm,在初始点云数据的基础上,提取了研究区域高精度的DEM (图2A)、CHM (图2B)和森林AGB(图2C)等衍生产品。DSM、DEM、CHM数据的详细生成方法参考文献[17],高精度LiDAR森林AGB的详细提取过程与方法参考文献[10,18]。
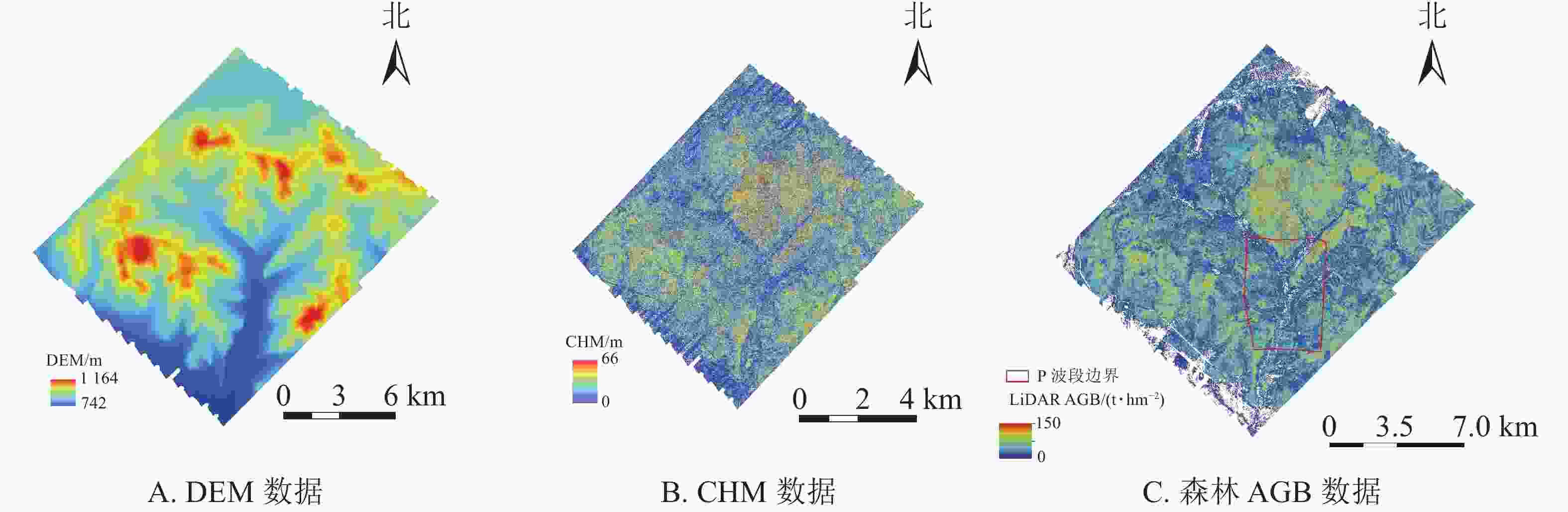
图 2 LiDAR衍生数据
Figure 2. Lidar derived data
为保证建模样本和验证样本能够代表整个研究区的森林AGB水平,以高精度LiDAR森林AGB图为基础,按照750 m的空间采样间隔,在ArcGIS中采用交互人工干预(去除道路及裸露地物)的方法选取113个样点(图1B)作为森林AGB反演模型的训练与验证。机载P波段SAR数据覆盖区森林林相不均匀,平均AGB较低,约46.7 t·hm−2,且大于100 t·hm−2的采样点仅有5个。113个样点AGB以10 t·hm−2间隔得分布情况(图1B):0~10 t·hm−2,3个;>10~20 t·hm−2,14个;>20~30 t·hm−2,17个;>30~40 t·hm−2,17个;>40~50 t·hm−2,18个;>50~60 t·hm−2,14个;>60~70 t·hm−2,10个;>70~80 t·hm−2,8个;>80~90 t·hm−2,7个;>90 t·hm−2,5个。
-
极化目标分解方法是从全极化数据中提取地物极化信息的有效方法,目前多种极化分解参数在森林类型识别中表现出巨大的潜力。本研究采用目前常用的极化后向散射系数、常用的3种极化分解方法提取极化SAR特征,并基于此分析森林的极化散射特征。提取P波段HH、HV和VV等3个极化的后向散射系数,基于3个后向散射系数的雷达植被指数(RVI)、极化辨别率参数(PDR);基于Freeman-Durden三分量分解的体散射分量(FVOL)、单次散射分量(FODD)、二次散射分量(FDBL)、体地散射比分量(FD1/FD2,1表示地散射分量是ODD和DBL的和,2表示地散射分量仅为ODD);基于Yamaguchi的体散射分量(YVOL)、单次散射分量(YODD)、二次散射分量(YDBL)、螺旋体散射分量(YHLX);基于H-A-ALPHA极化分解的极化散射熵(entropy)、反熵(anisotropy)、散射角(alpha)、目标方位向角(beta)、相位差角1(gamma)、相位差角2(delta)。3种极化分解方法参考文献[19-20]。由于本次飞行试验时并未布设角反射器用于定标,因此本研究使用的后向散射系数值仅有相对含义。
森林在P波段极化特征响应分析的目的是为了确定P波段对森林AGB动态变化敏感的极化特征参数,从而确定有效的极化特征参数进行森林AGB的估测。将研究区的森林AGB划分为A (表示生物量在0~30 t·hm−2变化时对应后向散射系数的箱线变化,均值约20 t·hm−2),B (31~50 t·hm−2,40 t·hm−2),C (51~70 t·hm−2,60 t·hm−2),D (71~90 t·hm−2,80 t·hm−2),E (>91 t·hm−2,100 t·hm−2)等5个变化等级,分别制作各等级相应P波段SAR提取参数值的箱线图,分析研究区各极化特征参数对森林AGB动态变化的响应,进而分析其极化散射特征。为了定量的分析各极化特征与森林AGB变化的关系,计算了它们之间的皮尔逊相关系数及显著性水平。
-
多元线性逐步回归模型(MLR)是森林AGB估测中最常用、最经典的方法之一。与常规的线性回归模型相比,MLR可同时完成模型输入参数的优选,进而提高森林AGB估测的效率和精度。MLR是将自变量逐个引入,每次判断自变量对因变量影响的显著性,并对模型中的自变量进行检验,逐个从模型中剔除不显著的变量,从而得到最优模型。即在保证显著性值在0.05以下的情况下,筛选相关性高的特征变量,进而得到因变量的最优估计。MLR的实现算法详见文献[21]。
-
KNN[22-25]、SVR[22, 24]和RF[24, 26]是森林AGB估测中常用的非参数模型,与参数模型相比,无固定的模型结构,通常通过数据驱动的方法来确定模型结构并用于森林AGB的估测,因此也称为机器学习方法。在森林AGB估测中,这3种方法各有优势,因此选取这3种方法来探究非参数方法在P波段森林AGB估测中的潜力。
-
反演结果精度的定量评价通过反演结果与真值之间的决定系数(R2)、均方根误差(RMSE)、平均绝对误差(MAE)、估测精度(Acc)来表征。这4个参数的计算公式参考文献[27]。
-
本研究获取的P波段中心波长为64 cm,该波段在森林中的穿透性较强。由图3可知:P波段20个SAR特征均表现出对研究区森林AGB变化的敏感性。P波段3种极化方式后向散射系数对森林AGB在0~110 t·hm−2内动态变化的响应与已有研究[28]相似,即后向散射系数的值均随着森林AGB的升高而缓慢增长。在各森林AGB水平中,HH、HV和VV散射能量强度相差并不明显,并且从箱线图(图3)的中值可以看出:HH的单调增长趋势较其他2个极化明显,且异常值较少。这可能是由于P波段波长较长,比较粗树干等圆柱体形状的散射体为森林中的散射体,使得去极化特征明显降低,因此同极化的HH和VV后向散射能量对AGB变化的敏感性明显高于HV[29]。此外,由后向散射系数计算的RVI也随着森林AGB增加而单调增加且未出现饱和现象,并且通过箱线图的宽度变化可以看出:各水平RVI箱宽变化不明显,说明该值对森林结构的变化不敏感,而对森林AGB的变化比较敏感。相比RVI,PDR非单调增长,在不同森林AGB水平,占主导散射的散射体会发生明显变化[30-31],该现象也可由Freeman-Durden中各个参数随森林AGB变化的响应加以说明。P波段Freeman-Durden分解相关特征中,除FODD和FDBL外,均随着森林AGB的增加而增加,并且2种体地散射比的值均远远大于1。Yamaguchi分解特征对森林AGB的响应趋势与Freeman-Durden分解相关特征的趋势基本一致。P波段H-A-ALPHA分解提取的参数中,散射熵(entropy)随着森林AGB增加而单调增加,而alpha则敏感性不明显。但beta、gamma和delta对森林AGB变化的敏感性则高于散射角alpha。P波段森林的极化散射特征随森林AGB的变化与已有研究中短波长的研究差异明显,说明现有的极化分解建模方法与该森林AGB水平P波段的散射特征有明显差异。
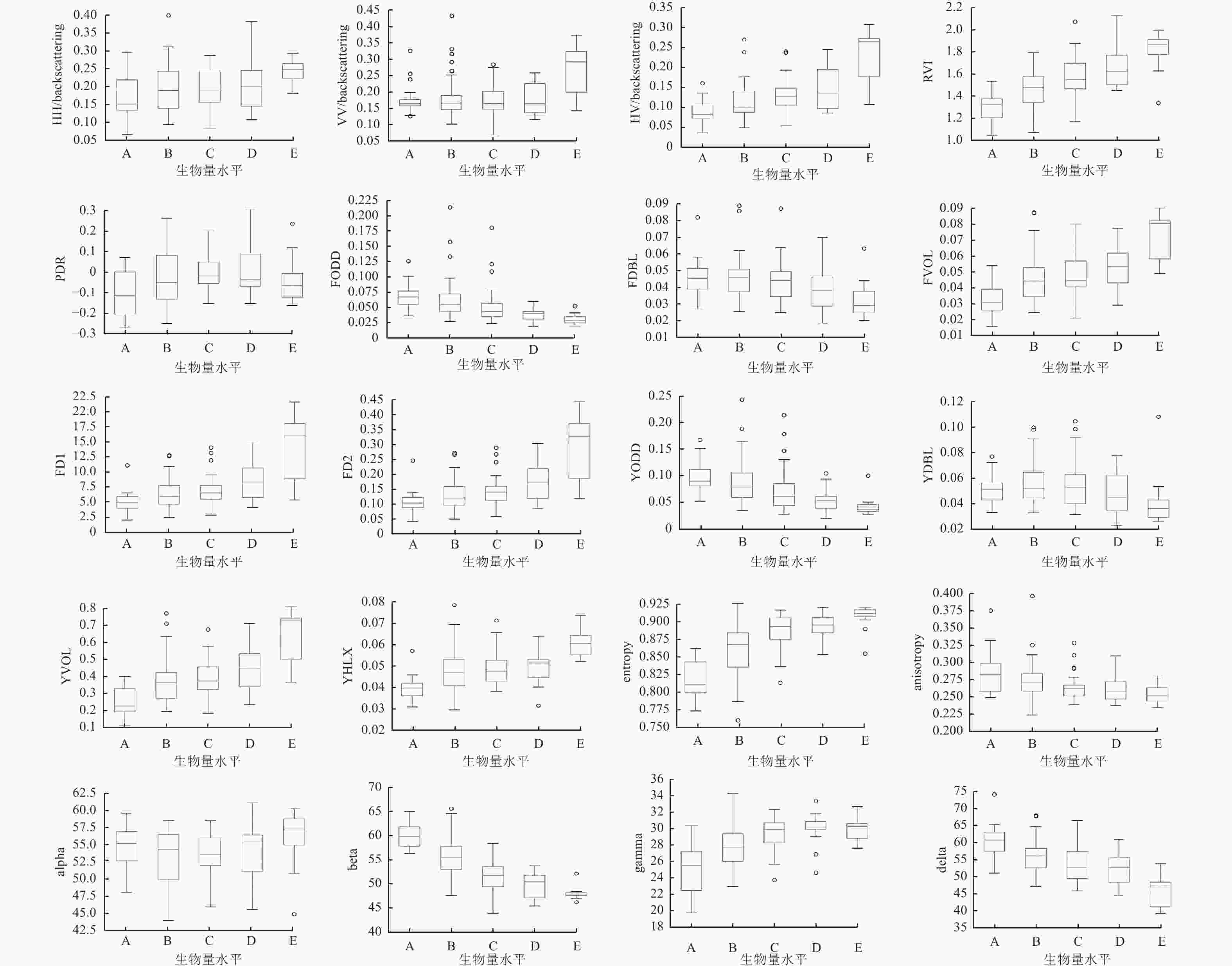
图 3 P波段森林散射机制分析
Figure 3. Forest scattering mechanism analysis at P band
由表1可知:极化分解中表征体散射分量的极化特征与森林AGB变化的显著性较低或不存在显著性,而二次散射特征分量则与森林AGB变化具有强相关性且显著性水平最高。这说明在本研究区,树干与地表形成的二次散射对森林AGB的变化最为敏感,且目前极化分解中的体散模型并不适合P波段研究区森林AGB水平下森林的散射机制。表1的定量分析结果与图4的定性分析结果对比可知:由于各极化参数的值域范围差异明显,图3对森林AGB敏感性较明显的参数在定量分析时相关性并不一定最高,因此图4的分析仅可作为初步参考,对于森林AGB敏感的极化特征参数的分析,仍需要采用皮尔逊系数进行相关性定量分析。
表 1 P波段SAR极化特征与森林AGB相关性分析
Table 1. Correlation coefficients of polarization feature and forest AGB
特征 R P 特征 R P 特征 R P HH 0.401** 0 F_VOL −0.129 0.173 alpha −0.734** 0 HV −0.249** 0.008 Y_DBL 0.719** 0 beta 0.037 0.696 VV −0.322** 0 Y_ODD 0.323** 0 delta 0.221* 0.018 FD1 −0.453** 0 Y_VOL −0.213* 0.024 gamma 0.314** 0.001 FD2 −0.448** 0 Y_HLX 0.187* 0.047 PDR 0.629** 0 F_DBL 0.650** 0 entropy −0.136 0.151 RVI −0.374** 0 F_ODD 0.303** 0.001 anisotropy 0.436** 0 说明:R代表Pearson相关系数;P代表显著性水平,**代表0.01水平上显著;*代表0.05水平上显著 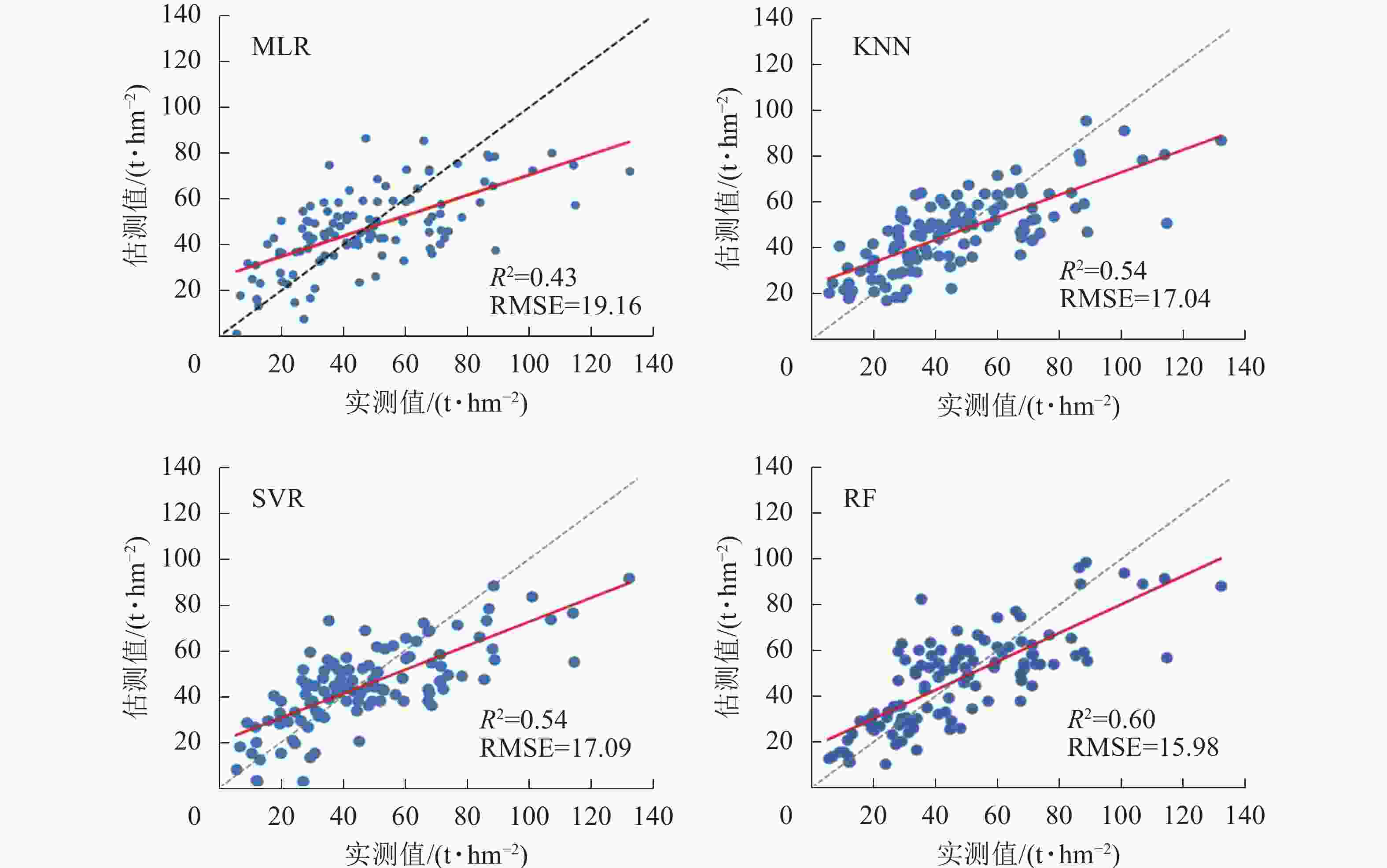
图 4 4种模型森林AGB估测与LiDAR抽样点散点图
Figure 4. Scatter plots of estimated and LiDAR forest AGB of four models
-
从表2可知:4种模型中,MLR模型估测结果精度最低,RF精度最高;而KNN和SVR模型的估测精度则相近,R2相同,RMSE、MAE和Acc也基本相同,比RF估测结果的最高值仅低约2%。
表 2 基于4种模型的P波段SAR 森林AGB估测结果
Table 2. P band SAR forest AGB inversion using four models
波段 反演模型 模型参数 R2 RMSE Acc/% MAE P MLR 显著性为0.0008 0.43 19.16 63.55 16.99 KNN K值为11,欧氏距离 0.54 17.04 71.18 13.44 SVR 惩罚系数7.0 0.54 17.09 71.15 13.45 RF 决策树数为100,树深度为11 0.60 15.98 72.97 12.60 从图4可看出:在整个森林AGB分布中,MLR估测结果分布较其他3种非参数方法分散,在森林AGB大于80 t·hm−2时出现了明显的饱和低估现象。其他3种方法尽管也有低估的现象,但是饱和现象并不明显。此外,本研究中4种方法估测结果的相对误差约30%,而以往区域性森林AGB的估测误差为37%~67%[32],采用P波段HV后向散射系数的估测结果中,同质性森林地区的相对误差约13%,而异质性地区相对误差则约60%[28]。在采用L-波段极化分解参数进行森林AGB反演研究中,在森林AGB水平低于120 t·hm−2时,后向散射的特征优于3种极化分解的特征,这与本研究中P波段的研究结果基本一致[33]。4种方法的估测结果中均出现了低值高估和高值低估的现象,这与以往采用不同遥感数据源进行森林AGB估测的结果一致[32]。图5中,RF的残差分布最接近高斯分布,尽管峰值出现在10 t·hm−2左右,由于其值分布较窄,因此具有较高的估测精度。KNN和SVR的残差分布图均较为连续,并且分布区间明显高于MLR,但由于正值占比较大,因此总体上呈现高估现象。MLR估测结果的残差分布图在10和20 t·hm−2出现了2个明显的峰值,且所有残差值的分布范围较宽,解释了其估测精度低于其他3种方法的原因。在ENGHART等[34]基于C和L-波段参数和非参数方法森林AGB估测的研究中,也发现MLR的估测结果要略低于非参数模型。然而参数和非参数模型的适用性还受到训练样本大小的影响,因此在训练样本较大时可选择非参数模型进行估测,当样本较小时则可优先选择MLR方法进行森林AGB估测[32]。
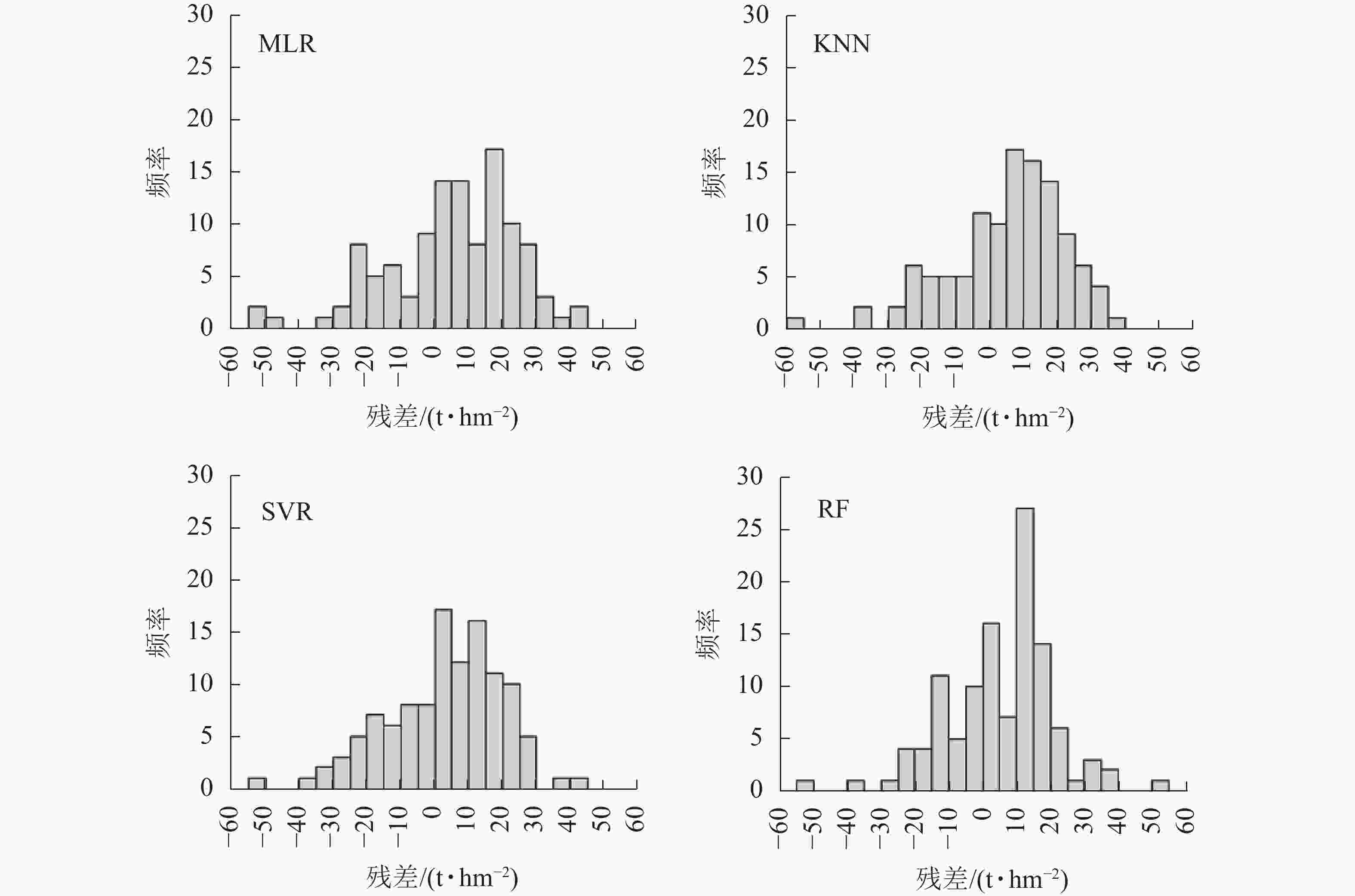
图 5 森林AGB估测结果与LiDAR抽样点差值直
Figure 5. Histograms of difference between forest AGB estimation results and LiDAR sampling points
为了进一步分析P波段极化特征对森林AGB估测的潜力,本研究将研究区森林AGB划分为不同的等级,然后采用4种估测模型中估测结果最优的RF模型进行估测,估测结果见表3。在本研究中将森林AGB划分为3组,其中第1组分别以30、60 t·hm−2为边界划分为3个子组;而第2组和第3组分别以40和50 t·hm−2为界划分为2个子组。由表3可知:分组界限不同,估测精度有明显的差异,分组划分越详细,估测精度(Acc)越高;此外,3种分组情况的估测结果均表明:在森林AGB平均值约45 t·hm−2,最高值不超过120 t·hm−2时,P波段在森林AGB水平较高的分组估测精度较高,如在第1组中,森林AGB大于30 t·hm−2分组的估测精度比小于30 t·hm−2分组的估测精约高5%;而在以50 t·hm−2为分组界限的2组中,森林高AGB组的估测精度比低AGB组的估测精度约高出6%。表3的结果分析表明:待估森林的AGB水平对P波段极化特征进行森林AGB估测的估测精度有明显影响,且P波段极化信息更适合森林AGB较高区域森林AGB的估测。
表 3 基于4种模型的P波段SAR 森林AGB反演情况
Table 3. P band SAR forest AGB inversion based on four models
序号 AGB分段/
(t·hm−2)R2 RMSE/
(t·hm−2)Acc/% 1 0~30 0.55 4.92 76.12 30~60 0.13 7.95 81.85 >60 0.25 14.89 81.57 2 0~50 0.47 8.46 72.15 >50 0.30 15.80 78.19 3 0~40 0.48 6.52 74.36 >40 0.36 16.21 74.82 -
针对P波段极化SAR数据在森林AGB估测中的潜力,提取了20个P波段极化SAR参数,探索了森林AGB估测中常用的MLR、KNN、SVR和RF方法在使用P波段极化SAR数据进行森林AGB估测的潜力。结果表明:P波段极化SAR信息在森林AGB估测中具潜力,但估测精度受到待估区域森林AGB水平高低的影响;4种估测方法中,非参数方法的估测结果明显优于MLR估测。P波段森林AGB估测结果中,同样存在AGB低值高估和高值低估的现象,其原因仍需要进一步探索。此外由于本研究区的森林AGB均值为45 t·hm−2,最高值低于120 t·hm−2,所以P波段极化信息在AGB高于120 t·hm−2的森林覆盖区中对森林AGB的估测能力仍有待进一步研究。
Forest above ground biomass estimation using airborne P band polarimetric SAR data
-
摘要:
目的 森林生物量的空间精准量化对了解陆地碳储量、碳收支、碳平衡,以及揭示森林碳储量与全球气候变化的影响过程具有重要意义。P波段波长较长,在森林中具有更高的穿透能力,研究机载P波段SAR数据提高森林地上生物量(AGB)估测精度的可行性。 方法 以机载P波段全极化合成孔径雷达(SAR)数据和高精度激光雷达(LiDAR)数据估测的森林AGB抽样点为基础,提取20个极化SAR特征,并分别与森林AGB变化作敏感性响应情况分析。采用多元线性回归模型(MLR)、K近邻方法(KNN)、支持向量回归(SVR)和随机森林(RF)4种估测方法,探究机载P波段SAR数据的森林AGB估测精度。 结果 在较低森林AGB(均值约45 t·hm−2)的森林覆盖区中,P波段的同极化后向散射系数、Freeman-Durden和Yamaguchi分解中的表面和二次散射分量对森林AGB变化敏感;此外H-A-ALPHA极化分解的散射角(alpha)、拓展极化参数极化辨别率参数(PDR)也对森林AGB变化敏感。4种方法估测的森林AGB相对误差均约30%,其中MLR估测结果精度最低,估测精度为63.55%,均方根误差(RMSE)为19.16 t·hm−2;RF估测结果精度最高,估测精度为72.97%,RMSE为15.98 t·hm−2;KNN和SVR估计结果差别不明显,RMSE分别为17.04和17.09 t·hm−2。 结论 P波段SAR数据对估测森林AGB具有一定潜力,3种非参数方法的估测结果明显优于MLR参数方法。此外,P波段的森林AGB估测精度受到待估森林AGB水平高低的影响明显,在森林AGB水平较高的分组中估测精度较高。在森林AGB均值为45 t·hm−2,最大值为120 t·hm−2的森林覆盖区,以50 t·hm−2将森林AGB样点分为2组时,高森林AGB组的估测精度高出低AGB组约6%。图5表3参34 -
关键词:
- P波段 /
- 森林地上生物量 /
- 合成孔径雷达(SAR) /
- 极化
Abstract:Objective Forests play an important role in carbon sequestration in terrestrial ecosystems. The spatial accurate quantification of forest biomass is of great significance to understand terrestrial carbon reserves, carbon budget, carbon balance and the resulting global climate change. Taking the advantage of the longer wavelength of P-band and higher penetration ability in the forest, the feasibility of improving the accuracy of forest above ground biomass (AGB) estimation using airborne P-band SAR data need to be studied . Method Based on the domestic airborne P band full polarimetric SAR data, 20 polarimetric SAR features are extracted, and were analyzed their sensitivity to change of forest AGB. Multiple linear regression model (MLR), k-nearest neighbor method (KNN), support vector regression (SVR) and random forest (RF), which were more popular forest AGB estimation models in previous studies, were used and compared in forest AGB estimation in this study. Result The results showed that polarimetric features including co-polarimetric backscatter coefficients, odd and double bounce scattering components extracted from Freeman-Durden and Yamaguchi decomposition methods, alpha from H-A-ALPHA decomposition method and polarization discrimination ratio (PDR), the extended polarimetric feature were sensitive to the change of forest AGB. The relative errors of estimated AGB using the four estimation methods were all about 30%, among which the accuracy of MLR estimation result was the lowest, with accurancy of 63.55% and root mean square error (RMSE) of 19.16 t·hm−2; The accuracy of RF estimation result was the highest, with Acc of 72.97% and RMSE of 15.98 t·hm−2; There is no significant difference between the accuracies between the estimated results of KNN and SVR, and the values of RMSE for them were 17.04 and 17.09 t·hm−2, respectively. Conclusion P-band SAR data has certain potential for estimating forest AGB. The estimation results of nonparametric method are significantly better than those of MLR. The AGB estimation accuracy of P-band is obviously affected by the level of forest AGB to be estimated, and the estimation accuracy is higher in the group with higher forest AGB level. In the study area, with an average forest AGB around 45 t·hm−2 and maximum value around 120 t·hm−2, the accurancy value for the estimated forest AGB at group with all AGB values lower than 50 t·hm−2 was lower around 6% than the value at group with all forest AGB values higher than 50 t·hm−2. [Ch, 5 fig. 3 tab. 34 ref.] -
Key words:
- P band /
- forest above ground biomass /
- SAR /
- polarimetric
-
图 1 P波段SAR数据(A)及其覆盖区样点分布(B)、林分概况(C)
Figure 1 P band SAR data (A), the distribution of samples (B) and stand examples (C)
图 4 4种模型森林AGB估测与LiDAR抽样点散点图
Figure 4 Scatter plots of estimated and LiDAR forest AGB of four models
图 5 森林AGB估测结果与LiDAR抽样点差值直
Figure 5 Histograms of difference between forest AGB estimation results and LiDAR sampling points
表 1 P波段SAR极化特征与森林AGB相关性分析
Table 1. Correlation coefficients of polarization feature and forest AGB
特征 R P 特征 R P 特征 R P HH 0.401** 0 F_VOL −0.129 0.173 alpha −0.734** 0 HV −0.249** 0.008 Y_DBL 0.719** 0 beta 0.037 0.696 VV −0.322** 0 Y_ODD 0.323** 0 delta 0.221* 0.018 FD1 −0.453** 0 Y_VOL −0.213* 0.024 gamma 0.314** 0.001 FD2 −0.448** 0 Y_HLX 0.187* 0.047 PDR 0.629** 0 F_DBL 0.650** 0 entropy −0.136 0.151 RVI −0.374** 0 F_ODD 0.303** 0.001 anisotropy 0.436** 0 说明:R代表Pearson相关系数;P代表显著性水平,**代表0.01水平上显著;*代表0.05水平上显著  下载: 导出CSV
下载: 导出CSV
表 2 基于4种模型的P波段SAR 森林AGB估测结果
Table 2. P band SAR forest AGB inversion using four models
波段 反演模型 模型参数 R2 RMSE Acc/% MAE P MLR 显著性为0.0008 0.43 19.16 63.55 16.99 KNN K值为11,欧氏距离 0.54 17.04 71.18 13.44 SVR 惩罚系数7.0 0.54 17.09 71.15 13.45 RF 决策树数为100,树深度为11 0.60 15.98 72.97 12.60
下载: 导出CSV
表 3 基于4种模型的P波段SAR 森林AGB反演情况
Table 3. P band SAR forest AGB inversion based on four models
序号 AGB分段/
(t·hm−2)R2 RMSE/
(t·hm−2)Acc/% 1 0~30 0.55 4.92 76.12 30~60 0.13 7.95 81.85 >60 0.25 14.89 81.57 2 0~50 0.47 8.46 72.15 >50 0.30 15.80 78.19 3 0~40 0.48 6.52 74.36 >40 0.36 16.21 74.82
下载: 导出CSV
-
[1] CARTUS O, SANTORO M, WEGMÜLLER U, et al. Benchmarking the retrieval of biomass in boreal forests using P band SAR backscatter with multi-temporal C- and L-band observations[J/OL]. Remote Sensing, 2019, 11(14): 1695 [2022-01-01]. doi: 10.3390/rs11141695. [2] SANTI E, PALOSCIA S, PETTINATO S, et al. Machine-learning applications for the retrieval of forest biomass from airborne P band SAR data[J/OL]. Remote Sensing, 2020, 12(5): 804 [2022-01-05]. doi: 10.3390/rs12050804. [3] LIAO Zhanmang, HE Binbin, QUAN Xingwen. Potential of texture from SAR tomographic images for forest aboveground biomass estimation[J/OL]. Int J Appl Earth Obs Geoinf, 2020, 88: 102049 [2022-01-05]. doi: 10.1016/j.jag.2020.102049. [4] QUAN Xingwen, HE Binbin, YEBRA M, et al. A radiative transfer model-based method for the estimation of grassland aboveground biomass [J]. Int J Appl Earth Obs Geoinf, 2017, 54: 159 − 168. [5] JI Yongjie, XU Kunpeng, ZENG Peng, et al. GA-SVR algorithm for improving forest above ground biomass estimation using SAR data [J]. IEEE J Selected Top Appl Earth Obs Remote Sensing, 2021, 14: 6585 − 6595. [6] LIU Y Y, DIJK A, RAMD J, et al. Recent reversal in loss of global terrestrial biomass [J]. Nat Clim Change, 2015, 5(5): 470 − 474. [7] BOUDREAU J, NELSON R F, MARGOLIS H A, et al. Regional aboveground forest biomass using airborne and spaceborne LiDAR in Québec [J]. Remote Sensing Environ, 2008, 112(10): 3876 − 3890. [8] JI Yongjie, HUANG Jimao, JU Yilin, et al. Forest structure dependency analysis of L-band SAR backscatter[J/OL]. PeerJ, 2020, 8: e10055 [2022-01-05]. doi: 10.7717/PEERJ.10055. [9] 李增元, 赵磊, 李堃, 等. 合成孔径雷达森林资源监测技术研究综述[J]. 南京信息工程大学学报(自然科学版), 2020, 12(2): 150 − 158. LI Zengyuan, ZHAO Lei, LI Kun, et al. A survey of developments on forest resources monitoring technology of synthetic aperture radar [J]. J Nanjing Univ Inf Sci Technol Nat Sci Ed, 2020, 12(2): 150 − 158. [10] 冯琦, 陈尔学, 李增元, 等. 基于机载P波段全极化SAR数据的复杂地形森林地上生物量估测方法[J]. 林业科学, 2016, 52(3): 10 − 22. FENG Qi, CHEN Erxue, LI Zengyuan, et al. Forest above-ground biomass estimation method for rugged terrain based on airborne P-band PolSAR data [J]. Sci Silv Sin, 2016, 52(3): 10 − 22. [11] DOBSON M C, ULABY F T, LETOAN T, et al. Dependence of radar backscatter on coniferous forest biomass [J]. IEEE Trans Geosci Remote Sensing, 1992, 30(2): 412 − 415. [12] TOAN T L, BEAUDOIN A, RIOM J, et al. Relating forest biomass to SAR data [J]. IEEE Trans Geosci Remote Sensing, 1992, 30(2): 403 − 411. [13] TOAN T L, QUEGAN S, WOODWARD I, et al. Relating radar remote sensing of biomass to modelling of forest carbon budgets [J]. Clim Change, 2004, 67(2): 379 − 402. [14] LIAO Zhanmang, HE Binbin, QUAN Xingwen, et al. Biomass estimation in dense tropical forest using multiple information from single-baseline P band PolInSAR data [J]. Remote Sensing Environ, 2019, 221: 489 − 507. [15] 黄国满. 机载多波段多极化干涉SAR测图系统——CASMSAR[J]. 测绘科学, 2014, 39(8): 111 − 115. HUANG Guoman. An airborne interferometric SAR mapping system with multi-band and multi-polarization: CASMSAR [J]. Sci Surv Mapp, 2014, 39(8): 111 − 115. [16] ZHAO Lei, CHEN Erxue, LI Zengyuan, et al. Three-step semi-empirical radiometric terrain correction approach for PolSAR data applied to forested areas[J/OL]. Remote Sensing, 2017, 9(3): 269 [2022-01-05]. doi: 10.3390/rs9030269. [17] 穆喜云. 森林地上生物量遥感估测方法研究[D]. 呼和浩特: 内蒙古农业大学, 2015. MU Xiyun. A Study on the Estimating Method of Forest Above Ground Biomass Based on Remote Sensing Data[D]. Huhhot: Inner Mongolia Agricultural University, 2015. [18] 陈传国. 东北主要林木生物量手册[M]. 北京: 中国林业出版社, 1989. CHEN Chuanguo. Woody Biomass Manual of Typical Species in the Northeast of China[M]. Beijing: China Forestry Publishing House, 1989. [19] CLOUDE S R, POTTIER E. A review of target decomposition theorems in radar polarimetry [J]. IEEE Trans Geosci Remote Sensing, 1996, 34(2): 498 − 518. [20] 张王菲, 姬永杰. 极化与干涉SAR植被参数反演[M]. 北京: 中国林业出版社, 2019. ZHANG Wangfei, JI Yongjie. Retrieval of Vegetation Parameters from Polarimetric and Interferometric SAR[M]. Beijing: China Forestry Publishing House, 2019. [21] 姬永杰. 多频极化SAR技术森林地上生物量反演研究[D]. 昆明: 西南林业大学, 2021. JI Yongjie. Retrieval of Forest above Ground Biomass Using Multi-frequency Polarimetric SAR[D]. Kunming: Southwest Forestry University, 2021. [22] 郭颖. 森林地上生物量的非参数化遥感估测方法优化[D]. 北京: 中国林业科学研究院, 2011. GUO Ying. Optimum Non-Parametric Method for Forest Above Ground Biomass Estimation Based on Remote Sensing Data[D]. Beijing: Chinese Academy of Forestry, 2011. [23] TIAN Xin, SU Zhongbo, CHEN Erxue, et al. Estimation of forest above-ground biomass using multi-parameter remote sensing data over a cold and arid area [J]. Int J Appl Earth Obs Geoinf, 2011, 17: 102 − 110. [24] LU Dengsheng, CHEN Qi, WANG Guangxing, et al. A survey of remote sensing-based aboveground biomass estimation methods in forest ecosystems [J]. Int J Digital Earth, 2016, 9(1): 63 − 105. [25] 韩宗涛, 江洪, 王威, 等. 基于多源遥感的森林地上生物量KNN-FIFS估测[J]. 林业科学, 2018, 54(9): 70 − 79. HAN Zongtao, JIANG Hong, WANG Wei, et al. Forest above-ground biomass estimation using KNN-FIFS method based on multi-source remote sensing data [J]. Sci Silv Sin, 2018, 54(9): 70 − 79. [26] 方匡南, 吴见彬, 朱建平, 等. 随机森林方法研究综述[J]. 统计与信息论坛, 2011, 26(3): 32 − 38. FANG Kuangnan, WU Jianbin, ZHU Jianping, et al. A review of technologies on random forests [J]. Stat Inf Forum, 2011, 26(3): 32 − 38. [27] 魏晶昱, 范文义, 于颖, 等. GF-3全极化SAR数据极化分解估算人工林冠层生物量[J]. 林业科学, 2020, 56(9): 174 − 183. WEI Jingyu, FAN Wenyi, YU Ying, et al. Polarimetric decomposition parameters for artificial forest canopy biomass estimation using GF-3 fully polarimetric SAR data [J]. Sci Silv Sin, 2020, 56(9): 174 − 183. [28] SAATCHI S, MARLIER M, CHAZDON R L, et al. Impact of spatial variability of tropical forest structure on radar estimation of aboveground biomass [J]. Remote Sensing Environ, 2011, 115(11): 2836 − 2849. [29] TOAN T L, QUEGAN S, DAVIDSON M W J, et al. The BIOMASS mission: mapping global forest biomass to better understand the terrestrial carbon cycle [J]. Remote Sensing Environ, 2011, 115(11): 2850 − 2860. [30] NEUMANN M, SAATCHI S S, ULANDER L M H, et al. Assessing performance of L and P band polarimetric interferometric SAR data in estimating boreal forest above-ground biomass [J]. IEEE Trans Geosci Remote Sensing, 2012, 50(3): 714 − 726. [31] KASISCHKE E S, CHRISTENSEN N L, BOURGEAU-CHAVEZ L L. Correlating radar backscatter with components of biomass in loblolly pine forests [J]. IEEE Trans Geosci Remote Sensing, 1995, 33(3): 643 − 659. [32] RODRÍGUEZ-VEIGA P, QUEGAN S, CARREIRAS J, et al. Forest biomass retrieval approaches from earth observation in different biomes [J]. Int J Appl Earth Obs Geoinf, 2019, 77: 53 − 68. [33] GOLSHANI P, MAGHSOUDI Y, SOHRABI H. Relating ALOS-2 PALSAR-2 parameters to biomass and structure of temperate broadleaf Hyrcanian forests [J]. J Ind Soc Remote Sensing, 2019, 47(5): 749 − 761. [34] ENGLHART S, KEUCK V, SIEGERT F. Modeling aboveground biomass in tropical forests using multi-frequency SAR data: a comparison of methods [J]. IEEE J Selected Top Appl Earth Obs Remote Sensing, 2012, 5(1): 298 − 306. -

-
链接本文:
https://zlxb.zafu.edu.cn/article/doi/10.11833/j.issn.2095-0756.20220111
 点击查看大图
点击查看大图

计量
- 文章访问数: 2120
- HTML全文浏览量: 351
- PDF下载量: 115
- 被引次数: 0
