




-
土壤与植被是紧密相连的整体,土壤养分直接影响着植物的生长状况[1]。相关研究表明:土壤养分质量分数的分布存在空间异质性[2−4]。目前,已有众多学者对国内不同气候带、不同地形区的土壤养分空间异质性进行了研究。例如,广西热带地区北部森林土壤养分质量分数高于南部[5],浙江中亚热带安吉县毛竹Phyllostachys edulis林土壤有机质和全氮质量分数呈现中等程度的空间变异(变异系数为10%~90%)[6],温带针阔混交林的土壤有机质、全磷等养分空间特征表现出由植被类型引起的条带状和斑块状分布[7]。也有学者研究了亚热带土壤养分空间异质性,如FU等[8]和董佳琦等[9]探讨了浙江亚热带的山核桃Carya cathayensis林和香榧Torreya grandis‘Merrillii’ 林地土壤养分空间分布规律,发现果实养分和土壤养分的空间分布格局具有相似性,大面积山核桃林具有较低的土壤pH和土壤磷质量分数,香榧主产区土壤养分失衡现象较为严重,应调整并改善区域施肥及管理模式;牛文鹏等[10]对珠江三角洲耕地、园地土壤养分空间变异特征的研究发现:土壤全氮、全钾空间异质性较高,土壤养分分布受控于气候、地形等自然因素,这对因地制宜选择作物种类具有指导意义;李超等[11]对亚热带高原耕地土壤主要养分空间变异规律的研究阐明了有机质与海拔的相关关系,为合理改良和利用耕地资源提供了依据。研究土壤养分空间分布的变异及其影响因素,可为探明区域土壤生产力,管理植被资源提供基础数据和科学理论。目前,关于亚热带土壤养分空间异质性的研究多集中于经济林和耕地等,缺乏亚热带天然生态系统的土壤养分空间异质性研究。
清凉峰是中国的典型亚热带国家级自然保护区,地处浙江省杭州市临安区境内。区内植被资源丰富,育有多种国家重点保护的珍稀特色植物种群。当前,对清凉峰的研究集中在探究保护区内植物多样性及地理分布等方面。此外,已有学者分析了该区内华南梅花鹿Cervus pseudaxis主要栖息地千顷塘的土壤肥力指标的空间变异规律[12],但对于土壤养分特征缺乏更全面、系统的调查研究。本研究的目的是:揭示亚热带地区天然生态系统土壤养分的质量分数特征;探明亚热带地区天然生态系统土壤养分的空间变异规律;解译亚热带地区天然生态系统土壤养分空间变异的影响因素,以期为保护中国亚热带丘陵地区的珍稀植被和天然生态系统提供科学依据。
-
清凉峰国家级自然保护区地处浙江省杭州市临安区西北部(30°00′42″~30°19′33″N,118°50′57″~119°13′23″E),属于亚热带季风气候区,总面积为11252 hm2,最高海拔为1787.4 m,年平均气温为12.5 ℃,年降水量为1862.2~2331.9 mm。该区地层以侏罗系分布最广,主要包括中酸性火山岩、紫红色层状粉砂岩、粉晶灰岩、硅质页岩以及岩浆岩等,土壤结构由低海拔到高海拔呈现红壤带、山地黄壤带、山地棕黄壤带及草甸土带的垂直带谱。清凉峰由龙塘山、顺溪坞、千顷塘3块区域组成。本研究区域为龙塘山和顺溪坞(图1),境内海拔高差悬殊,由西北向东南方向倾斜,地势呈现“高—低—高”特征。植被垂直分布明显,蕴藏着丰富的植物资源,包括华榛Corylus chinensis、银缕梅Parrotia subaequalis、鹅掌楸Liriodendron chinense等。因保护区内存在轻微的人为土地开垦活动,本研究将该部分区域(农用地)纳入研究,与其他自然区域进行对比分析。
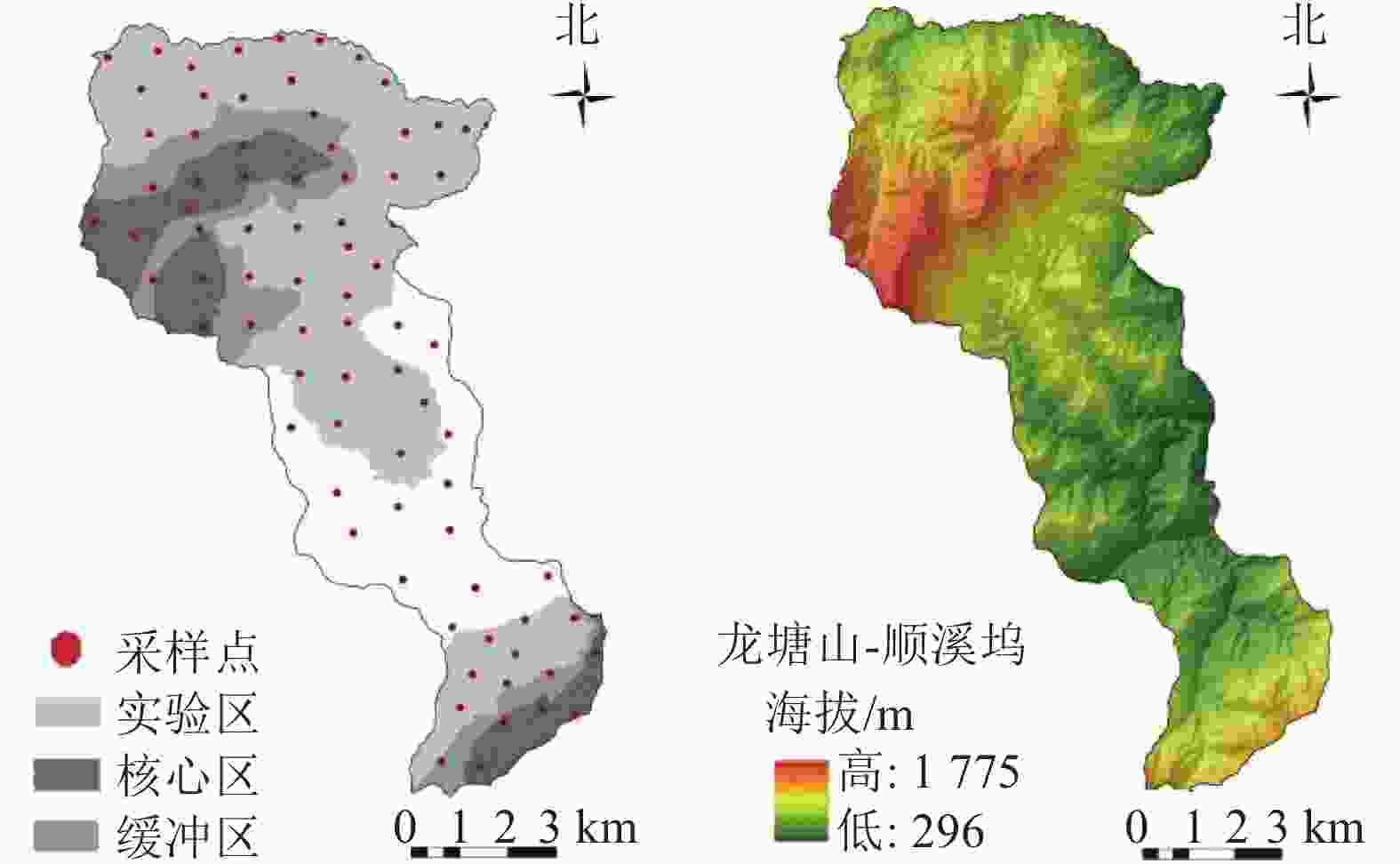
图 1 研究区样点分布及海拔信息示意图
Figure 1. Spatial distribution and elevation information of soil sampling sites
-
基于样点分布的均质性和科学性原则,结合地形图和植被类型分布特征,在研究区内按等距离网格法布设1.0 km×1.0 km的样方。于2019年11月至2020年4月,在研究区每个样方中采集0~20 cm表层土壤样品3个,并均匀混合后得到1个土壤样品,共采集土壤样品56个。由于网格样方较大,为增加研究的精确性,采样过程中结合实际地形地势,在6个海拔范围(200~500、500~800、800~1 000、1 000~1 200、1 200~1 400、1 400~1 600 m)补充采集24个土壤样品,最终得到80个样品。采样时使用全球定位系统(GPS)记录样点地理信息和植被类型。土壤样品经风干,去除植物残体及石块等杂物,研磨过筛备用。
土壤样品养分测定[13]:pH用pH酸度计测定;土壤有机质采用重铬酸钾氧化还原滴定法测定;全氮采用半微量凯氏法测定;全磷采用氢氧化钠碱熔-钼锑抗比色法测定;全钾采用氢氧化钠熔融-火焰分光光度法测定;容重采用环刀法测定。
-
研究区采用美国国家航空航天局(NASA)官方网站(
https://search.asf.alaska.edu/#/ )的12.5 m×12.5 m数字高程模型(DEM)。在采样时用GPS测定野外采样点实地坐标、高程数据。利用ArcGIS 10.2空间分析工具直接提取基本地形数据,结合空间分析工具、水文分析模块以及栅格计算器经复合计算获取坡度数值。 -
半方差函数是地统计学中描述变量的定量参数,在土壤元素空间变异性研究中应用广泛,用于揭示区域化变量的随机性和结构性特征,具体计算公式及参数见文献[14]。
半方差分析的参数中,块金值(C0)表示由采样和检测分析误差引起的随机变异,基台值(C0+C)代表总空间变异程度,块基比[C0/(C0+C)]表示随机变异的占比情况,用于衡量空间相关程度[15],变程(a)表示空间自相关的作用范围。
-
土壤属性在地理空间上与相邻区域的观测值存在的相互依赖性,即空间自相关,可用全局Moran’s I指数(IN)反映指标的空间自相关性大小,具体计算公式及参数见文献[16]。IN指数取值范围是−1~1,小于0表示负相关,等于0表示不相关,大于0表示正相关。IN的显著性水平采用下式检验:
$$Z=\left[I_{{\rm{N}}}-{E}\left(I_{{\rm{N}}}\right)\right] / \sqrt{\operatorname{var}\left(I_{{\rm{N}}}\right)} 。 $$ 其中:Z为检验IN指数的显著性统计量;E(IN)为期望值;var(IN)为方差。
$ \left| Z \right| $ ≥1.96和$ \left| Z \right| $ ≥2.58分别是空间自相关显著和极显著的分界值。 -
本研究利用3倍标准差法(阈值法)剔除统计数据的异常值[17]。采用SPSS 20.0软件进行土壤养分数据的描述性统计分析、数据正态性检验[Kolmogorov-Smirnov (K-S)法]、不同植被下土壤养分的显著性检验、地形因子和土壤养分之间的相关性分析;采用GS+7.0软件进行半变异函数分析和相关模型参数优化;利用Geoda进行空间自相关计算;基于ArcGIS 10.7软件进行地形因子的计算和插值图的绘制。
-
龙塘山-顺溪坞土壤养分元素质量分数描述性统计结果如表1所示。土壤pH为3.82~6.70,均值为5.10,属于酸性土壤;研究区土壤有机质、全磷、全氮、全钾平均质量分数分别为65.04、0.67、1.63、18.45 g·kg−1。根据全国第2次土壤普查属性分级标准[18],有机质质量分数处于极丰富水平(一级),全氮处于丰富水平(二级),全钾、全磷质量分数处于较丰富水平(三级)。土壤各养分的变异系数为12%~50%,根据王政权等[19]划分标准,所有养分均属于中等程度变异。K-S法正态性检验结果显示:pH、全氮符合正态分布,有机质、全磷、全钾经对数转换后符合正态分布。
表 1 龙塘山-顺溪坞土壤养分描述性统计特征
Table 1. Descriptive statistical characteristics of soil fertility properties in Longtangshan-Shunxiwu
指标 pH 有机质/(g·kg−1) 全磷/(g·kg−1) 全氮/(g·kg−1) 全钾/(g·kg−1) 最小值 3.82 10.24 0.24 0.35 5.96 最大值 6.70 143.43 1.53 3.06 41.54 均值 5.10 65.04 0.67 1.63 18.45 标准差 0.59 32.61 0.27 0.60 8.73 变异系数/% 12 50 40 37 47 偏度 0.56 −0.27 0.14 0.37 −0.09 峰度 0.07 0.62 −0.30 −0.31 −0.66 K-S 0.20 0.20 0.20 0.20 0.20 分布类型 正态分布 对数正态分布 对数正态分布 正态分布 对数正态分布 -
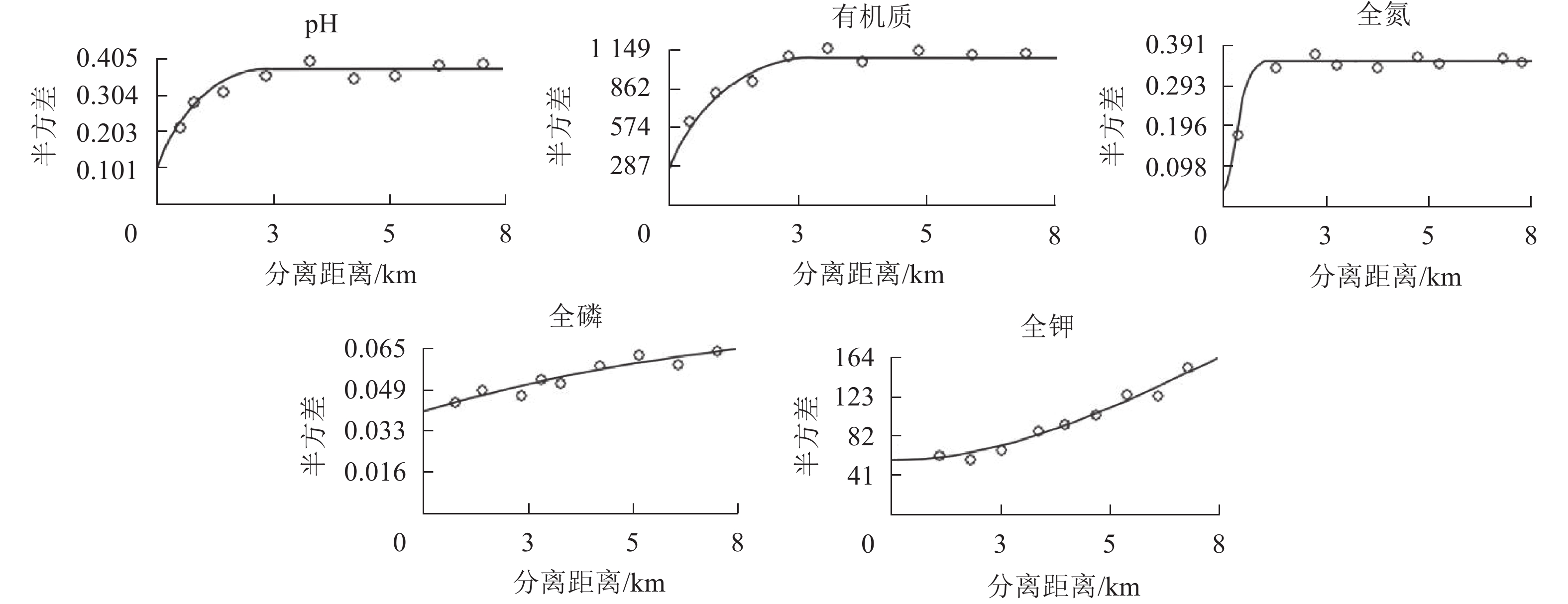
在一定的分离距离内,土壤pH和各养分的半方差均随分离距离的增加而增大(图2),最终近似趋于基台值或是围绕基台值上下波动,说明各养分在采样尺度上有一定的空间依赖性[20]。
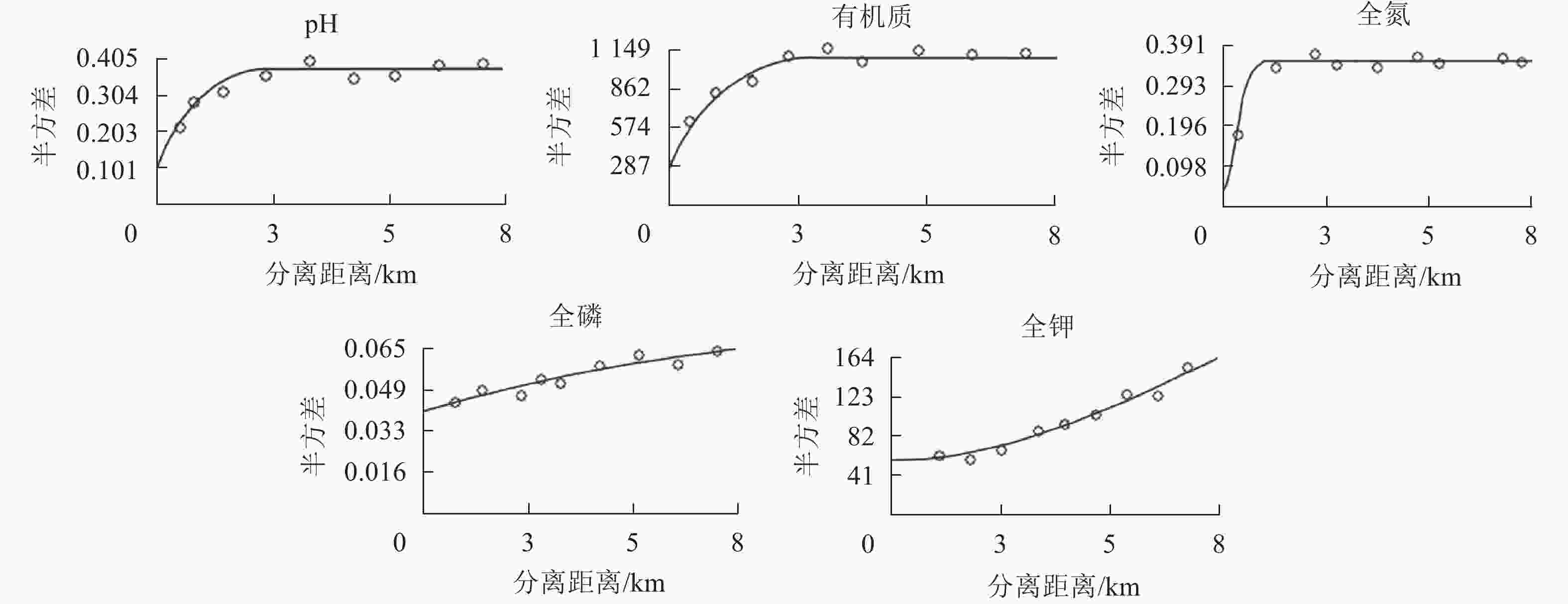
图 2 龙塘山-顺溪坞土壤养分半变异函数分析
Figure 2. Semivariograms of soil fertility properties in Longtangshan-Shunxiwu
利用地统计学半方差特征参数函数分析各养分的理论拟合模型(表2)。土壤pH、有机质、全磷质量分数分布符合指数模型,土壤全氮、全钾质量分数分布符合高斯模型。拟合模型能较准确反映龙塘山-顺溪坞土壤养分的空间变异结构特征(决定系数R2>0.5)。土壤有机质、全氮、全钾的块基比分别为0.23、0.11、0.18,表现为强烈空间自相关,说明其变异主要受结构性因素影响。土壤pH、全磷的块基比分别为0.26、0.50,处于中等强度的空间相关性,表明随机因素可能存在一定的影响。IN指数分析结果显示:土壤有机质具有极显著的空间自相关性(P<0.01,Z>2.56),表明有机质分布较聚集;而土壤全氮、全钾的空间自相关性不显著,在空间上呈离散分布。土壤pH的变程很小,说明土壤pH的主控因素的空间连续性尺度较小,分布趋向于随机化,这与IN显著性检验结果较为一致。
表 2 土壤养分半变异模型参数及IN指数
Table 2. Semivariogram model parameters of soil fertility properties and global IN index
指标 模型 块金值C0 基台值(C0+C) 块基比[C0/(C0+C)] IN Z 变程/km 决定系数(R2) pH 指数模型 0.10 0.38 0.26 0.08 1.35 2.00 0.76 有机质 指数模型 250.00 1084.00 0.23 0.24** 3.29 5.00 0.72 全磷 指数模型 0.04 0.08 0.50 0.16* 2.31 26.00 0.90 全氮 高斯模型 0.04 0.36 0.11 0.04 0.66 796.74 0.82 全钾 高斯模型 57.90 326.70 0.18 <0.05 −0.52 354.20 0.93 说明:*表示在0.05水平上显著相关(|Z|>1.96);**表示在0.01水平上显著相关(|Z|>2.58) -
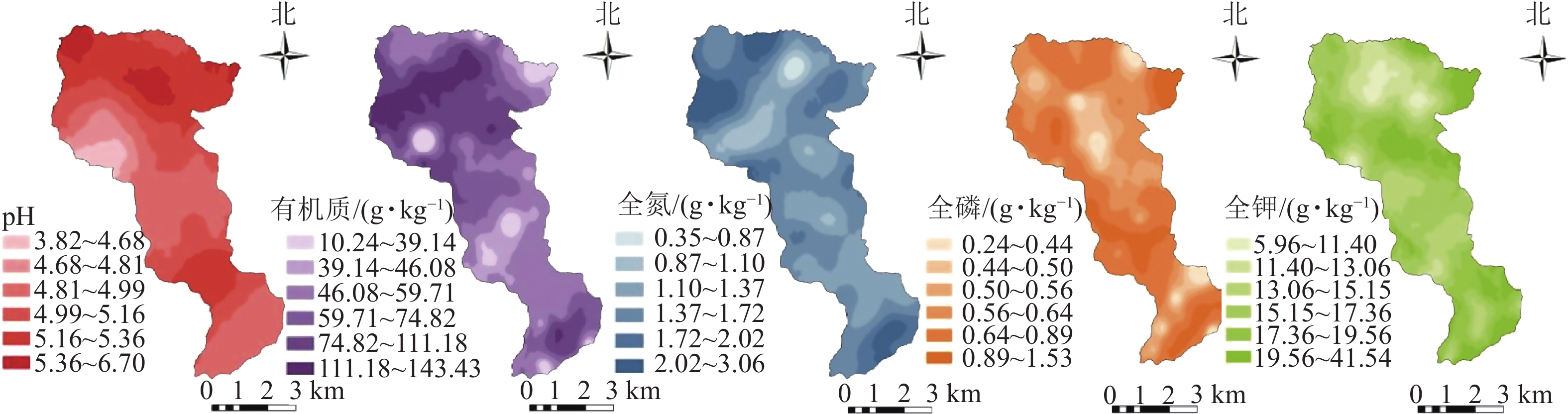
龙塘山-顺溪坞土壤养分质量分数的空间分布如图3所示。整体上,各土壤养分的空间分布不均,斑块特征显著,表现较为明显的空间异质性。土壤pH处于5.0~5.5的范围最大,整体呈酸性;有机质质量分数大于40 g·kg−1的点位分布广泛,高值区集中分布在中偏西北部,少量分布在东南部,低值区以斑状散布在整个研究区;全氮的高值区以斑状散布在研究区边缘且主要分布于西北、东南两端,低值区则以斑块状散布于研究区内部;全磷、全钾高低值区分布较为分散,在海拔较高处分布着明显的低值区。在南部(海拔为340~816 m)出现土壤pH、全磷、全钾质量分数较高而土壤有机质和土壤全氮质量分数较低的情况。
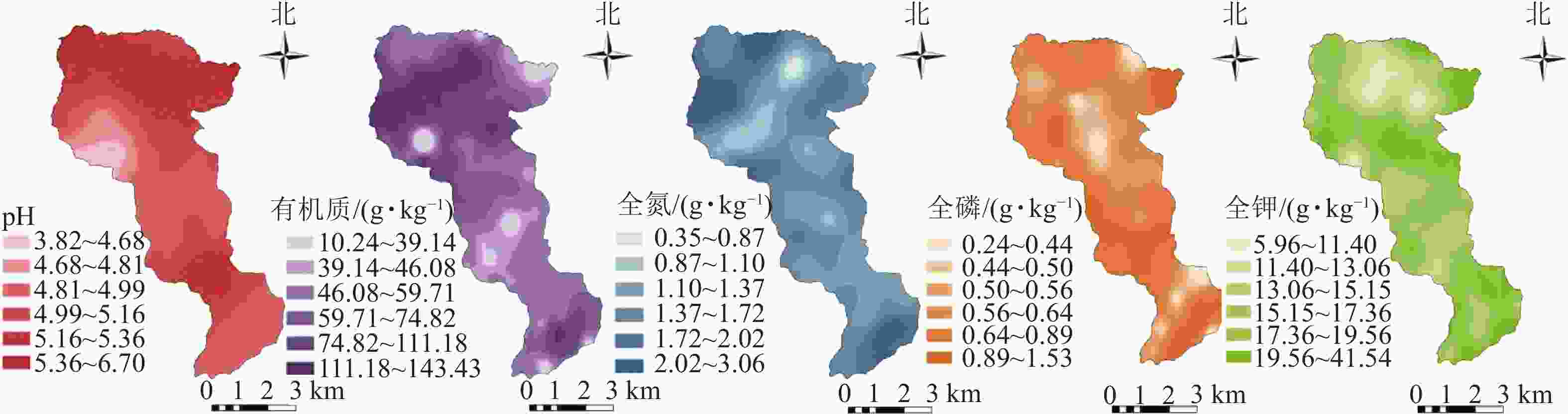
图 3 龙塘山-顺溪坞土壤养分空间分布示意图
Figure 3. Spatial distribution map of soil fertility properties in Longtangshan-Shunxiwu
-
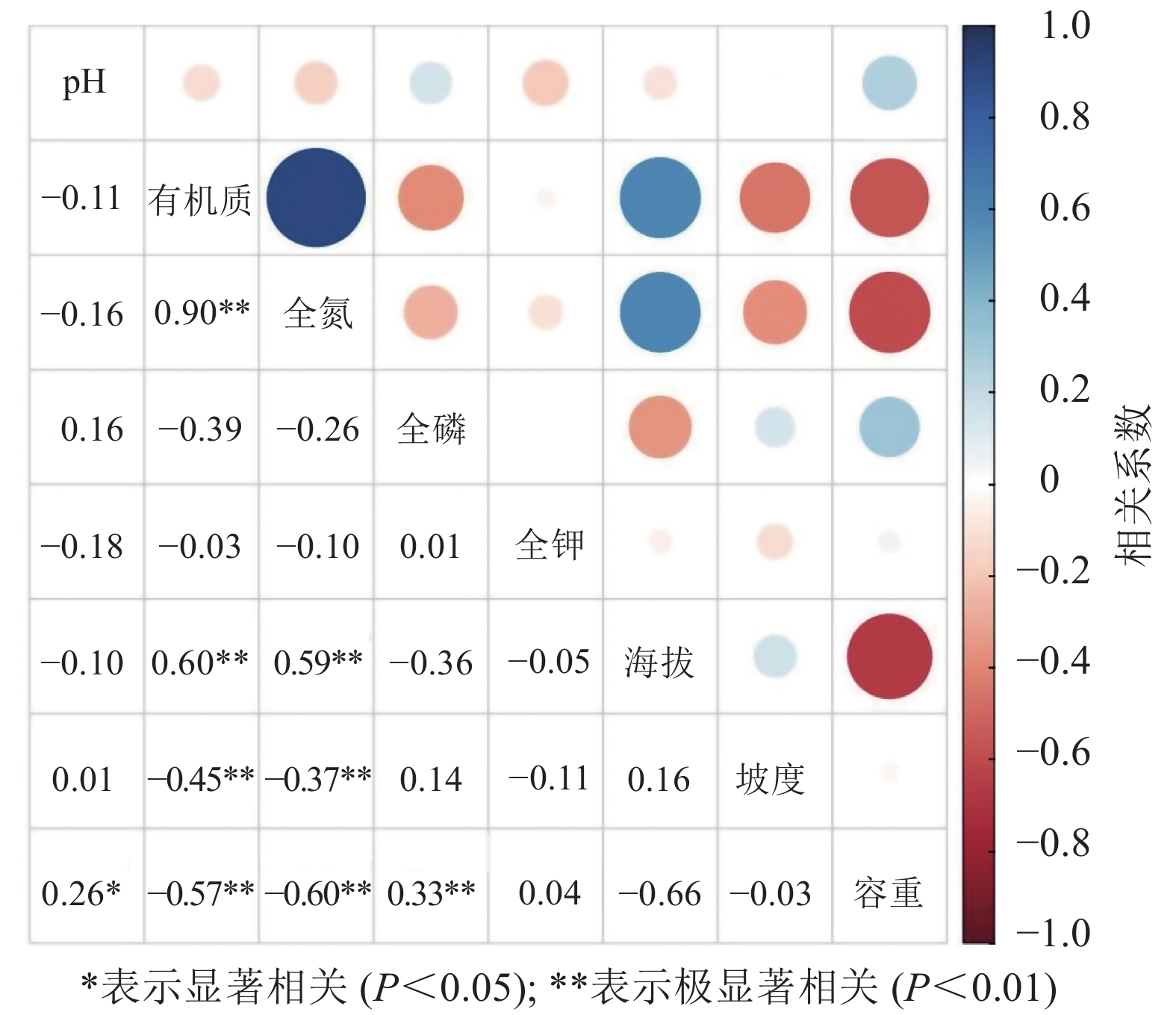
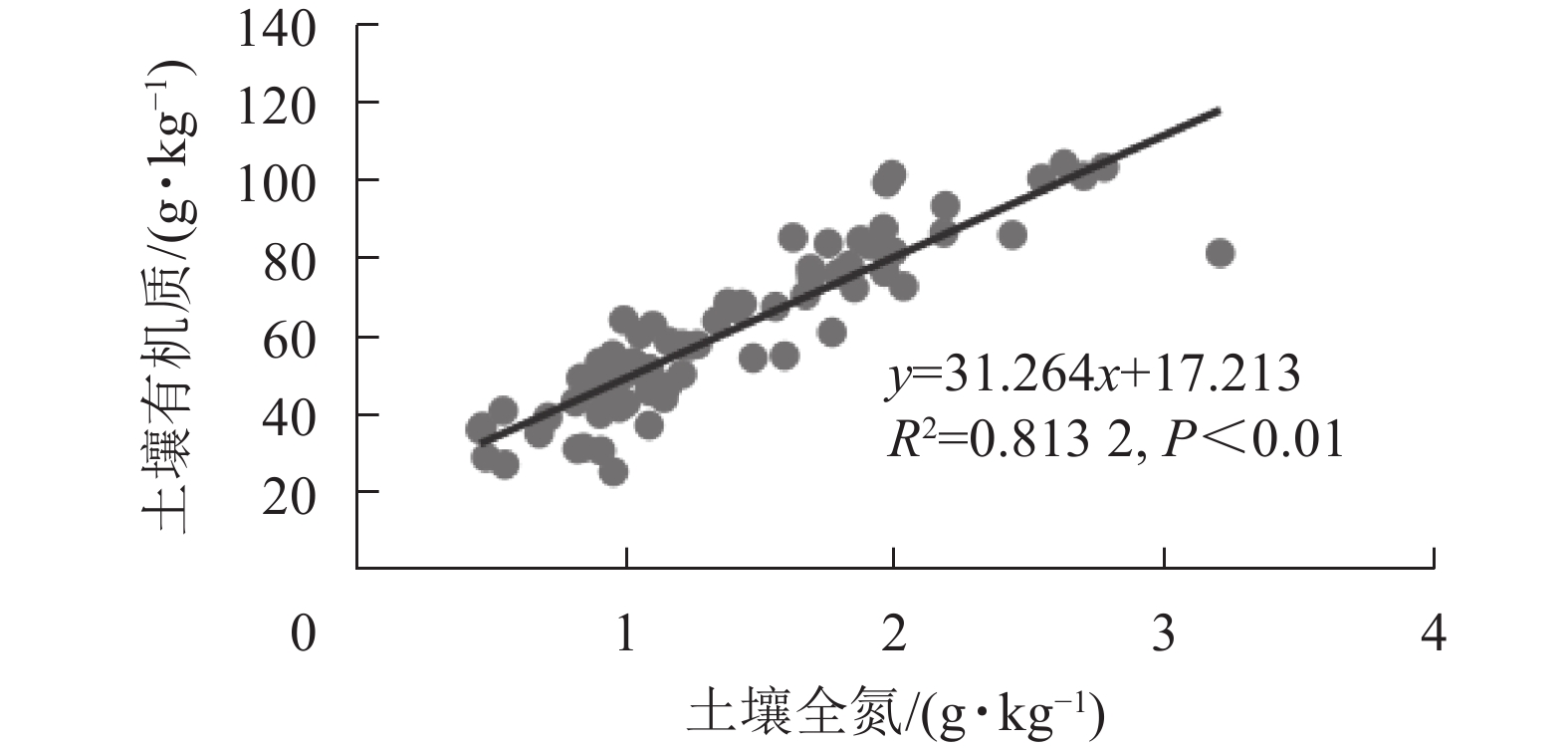
龙塘山-顺溪坞土壤养分与环境因子的相关分析结果显示(图4):有机质和全氮与地形因子具有较为一致的相关性。线性拟合结果表明:土壤有机质和全氮之间呈极显著的正相关性(P<0.01)(图5)。有机质和全氮均与海拔呈极显著正相关(P<0.01);全磷、全钾质量分数呈现出随海拔升高而下降的趋势;有机质和全氮与坡度、土壤容重均呈极显著负相关(P<0.01);土壤pH与土壤容重呈正相关性(P<0.05),土壤全磷与土壤容重呈极显著正相关(P<0.01)。
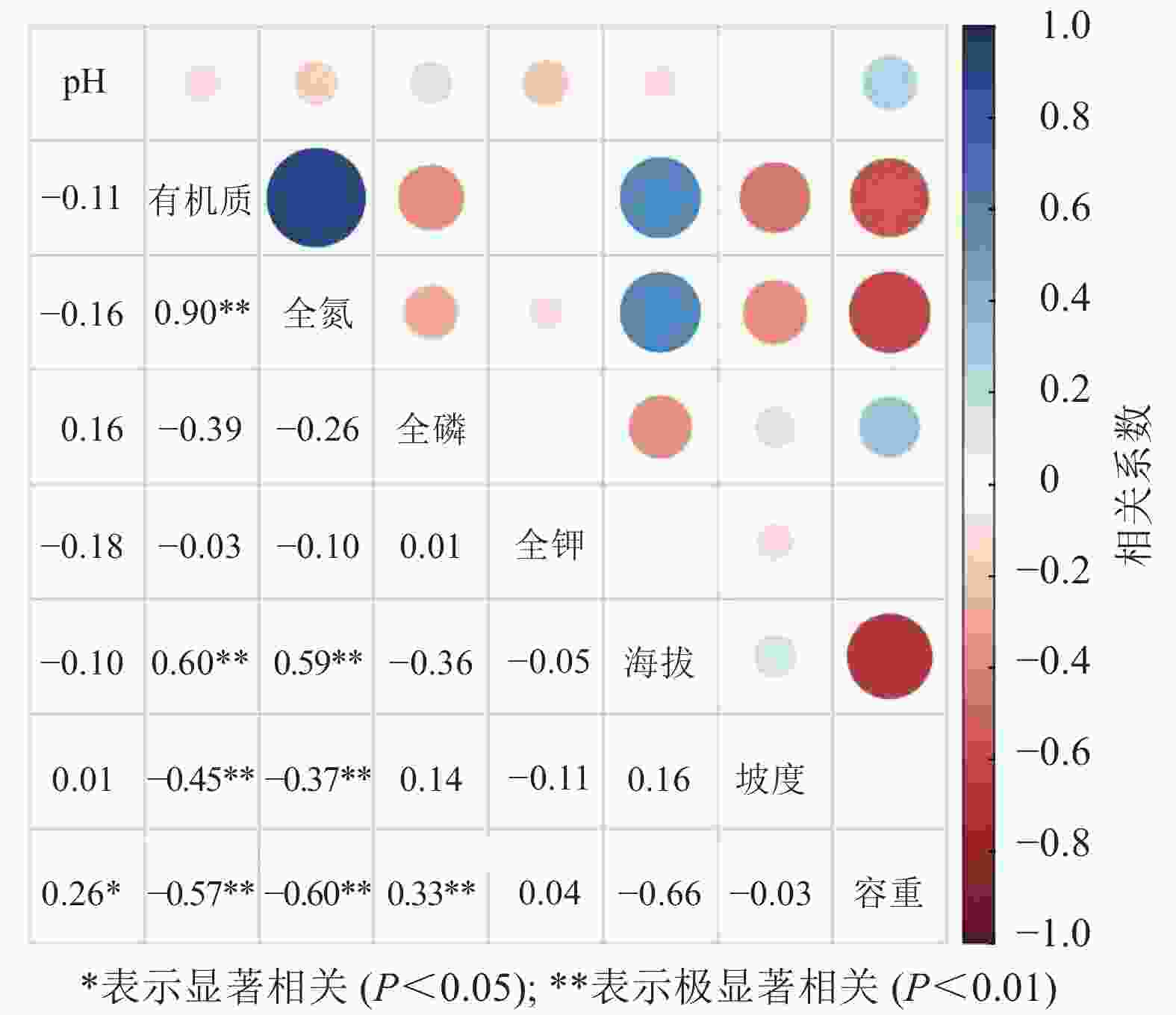
图 4 龙塘山-顺溪坞土壤养分与环境因子的相关分析
Figure 4. Correlation analysis between soil nutrients and environmental factors in Longtangshan-Shunxiwu
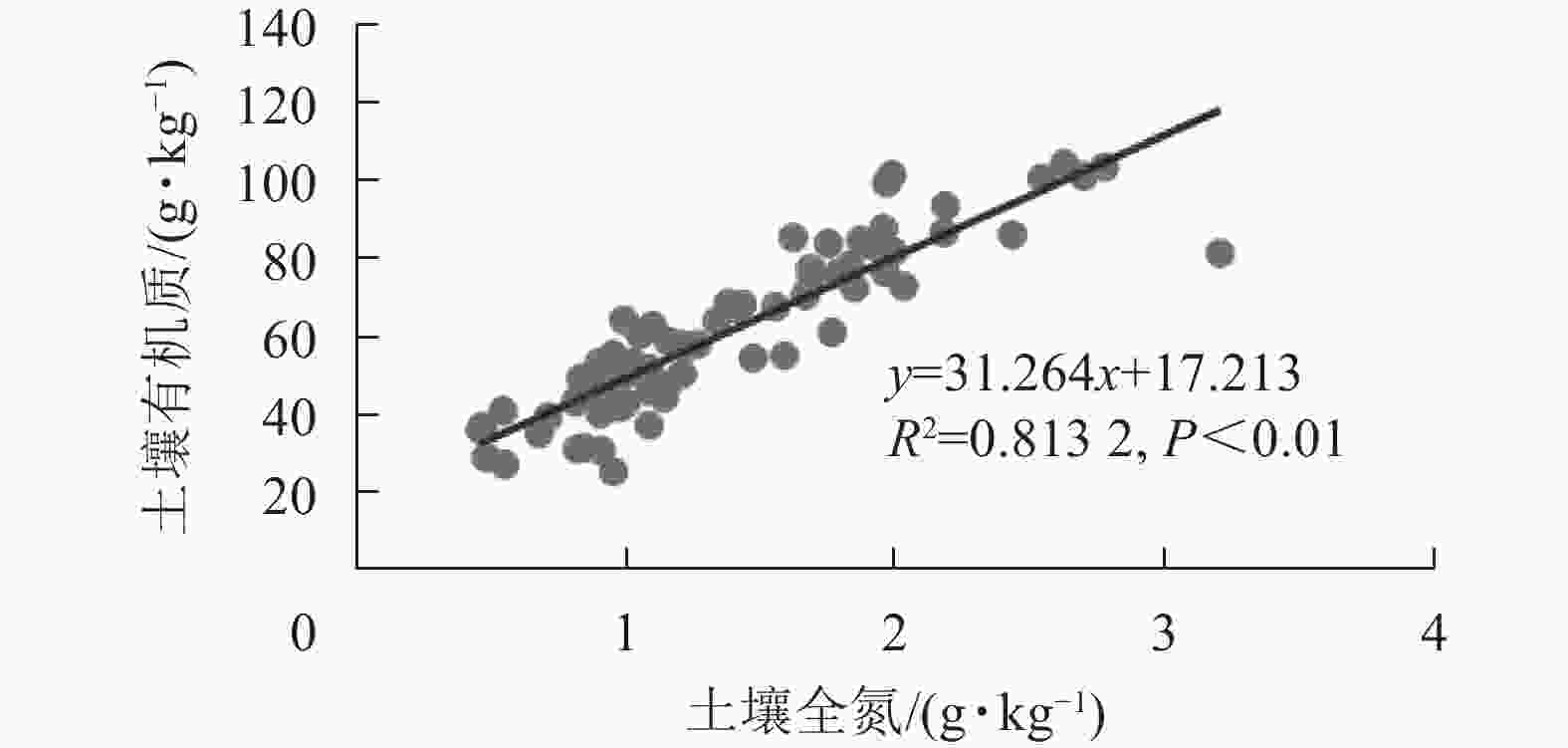
图 5 土壤有机质与土壤全氮的关系
Figure 5. Relationship between soil organic matter and total nitrogen
-
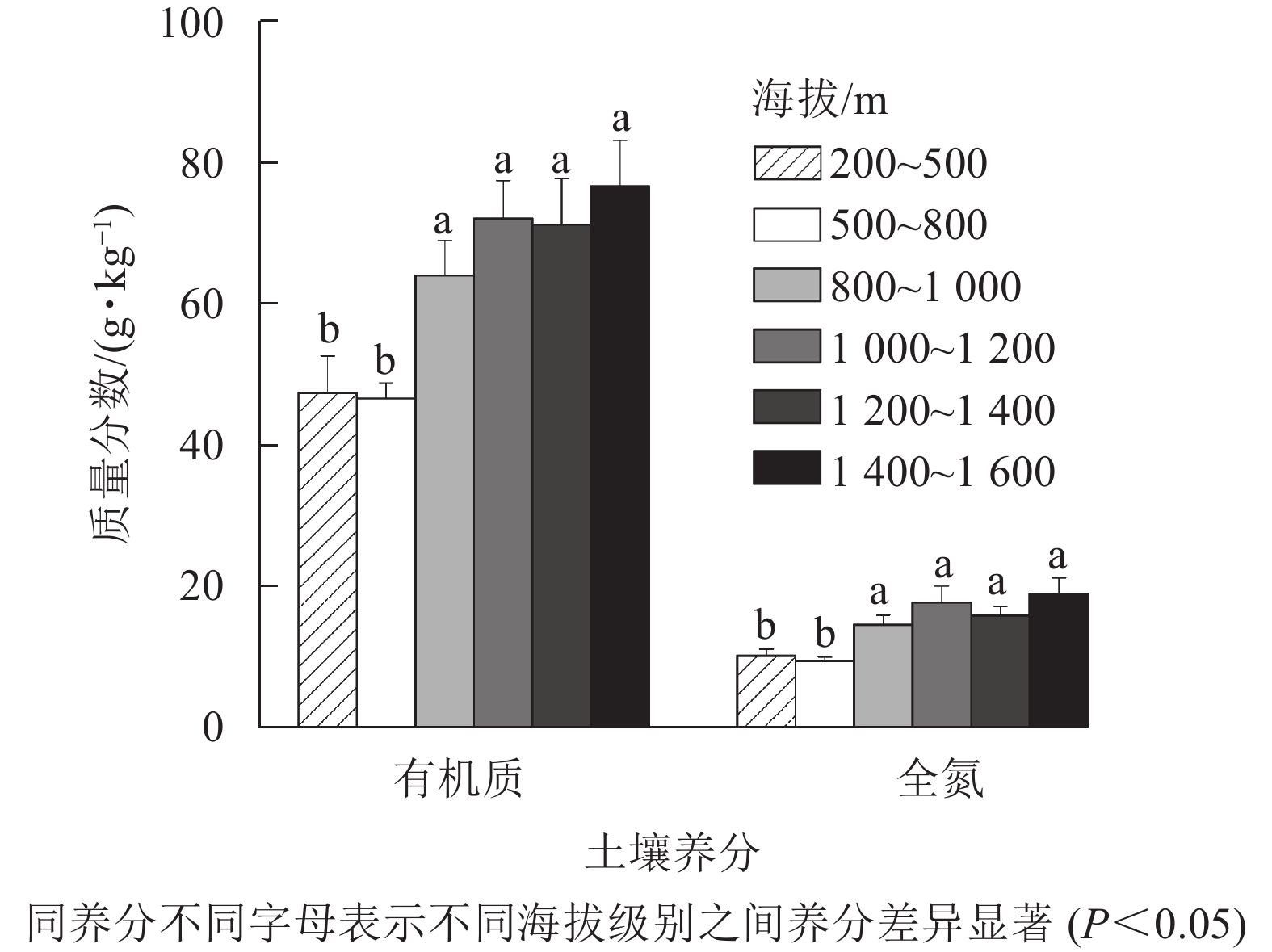
为了进一步研究龙塘山-顺溪坞土壤有机质和全氮随海拔变化的规律,将海拔划分为200~500、500~800、800~1 000、1 000~1 200、1 200~1 400、1 400~1 600 m等6个等级(下文分别以1、2、3、4、5、6级海拔代替) (图6)。由图6可知:土壤有机质和土壤全氮随海拔升高而增加,变化规律一致。在1、2级海拔,土壤有机质和土壤全氮与3、4、5、6级海拔间均存在显著差异(P<0.05)。
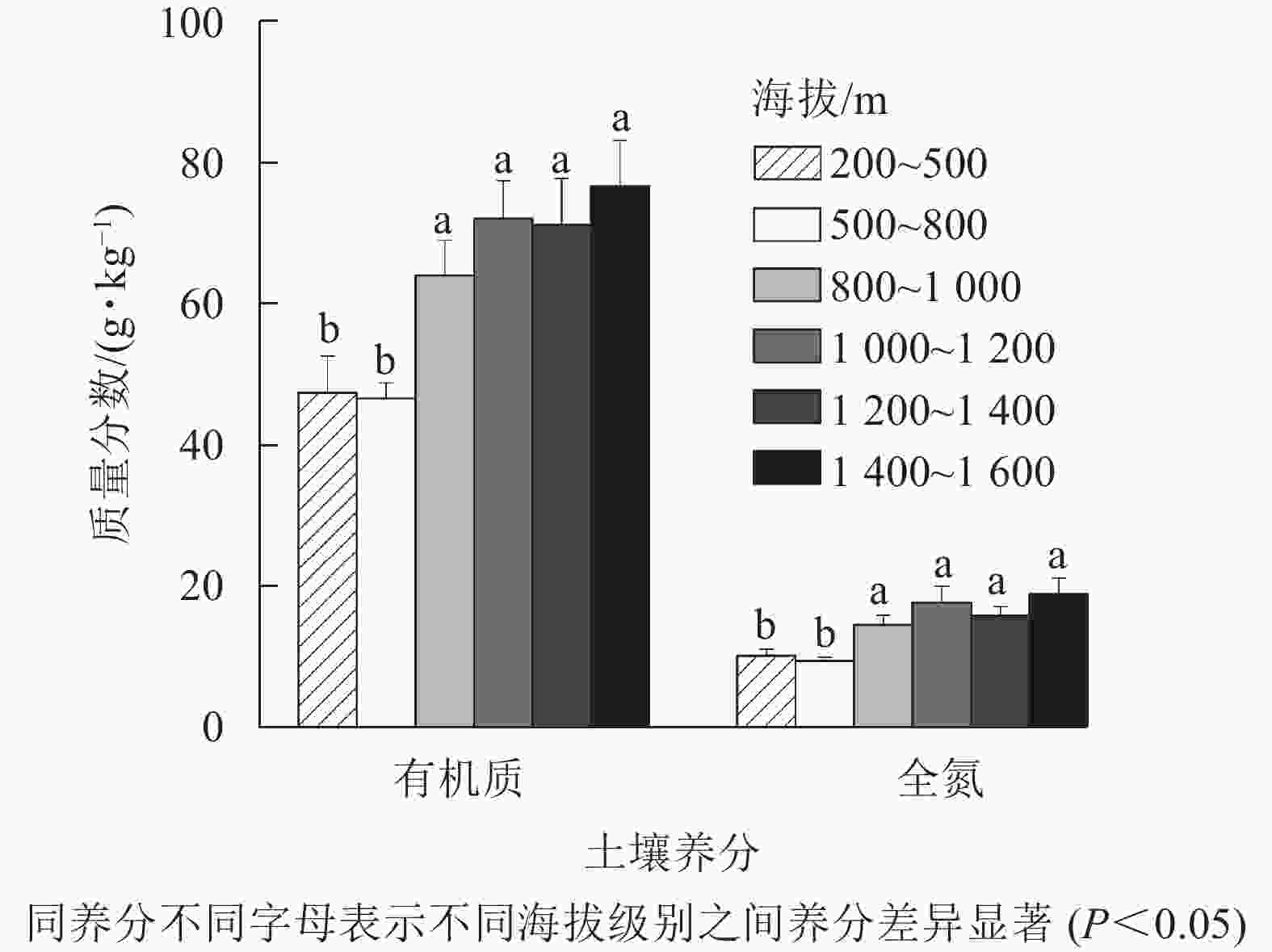
图 6 龙塘山-顺溪坞不同海拔土壤养分特征
Figure 6. Characteristics of soil fertility properties under different elevations in Longtangshan-Shunxiwu
-
有机质和全氮质量分数随着容重增加而减少(表3),有机质质量分数下降较快,全氮质量分数下降较慢。土壤pH和全磷随着容重增加而增加,表明pH和全磷与土壤容重呈正向相关性,且两者上升趋势较小,总体变化平缓。
表 3 龙塘山-顺溪坞土壤化学性质随土壤容重的变化
Table 3. Changes of soil chemical properties with soil bulk density in Longtangshan-Shunxiwu
容重/(g·cm−3) pH 有机质/(g·kg−1) 全磷/(g·kg−1) 全氮/(g·kg−1) 容重/(g·cm−3) pH 有机质/(g·kg−1) 全磷/(g·kg−1) 全氮/(g·kg−1) 0.8~0.9 4.61±0.68 79.41±20.73 0.11±0.04 2.29±0.90 1.4~1.5 5.37±0.58 59.48±19.48 0.12±0.04 1.26±0.59 0.9~1.0 5.29±1.13 80.81±5.40 0.07±0.03 1.81±0.11 1.5~1.6 5.13±0.75 52.82±14.42 0.17±0.10 1.04±0.32 1.0~1.1 4.87±0.39 74.86±18.27 0.11±0.06 1.89±0.47 1.6~1.7 5.17±0.68 41.08±10.31 0.14±0.08 0.91±0.13 1.1~1.2 5.03±0.76 71.68±18.62 0.10±0.03 1.72±0.67 1.7~1.8 5.80±0.45 42.50±13.67 0.39±0.43 0.80±0.29 1.2~1.3 4.96±0.31 65.44±20.67 0.10±0.05 1.42±0.55 F 5.13 36.69 11.01 41.04 1.3~1.4 5.17±0.57 46.44±11.63 0.15±0.09 1.08±0.33 P 0.03 <0.001 0.001 <0.001 说明:数值为平均值±标准差 -
对龙塘山-顺溪坞不同植被类型下土壤养分质量分数进行统计(表4)可知:不同植被类型对土壤养分均有影响,且对土壤有机质和全氮的影响达显著水平(P<0.05)。与其他植被类型土壤相比,农田土壤有机质和全氮质量分数偏低,而土壤pH、全磷较高;草甸中土壤有机质质量分数最高;针叶林土壤全钾质量分数最高。
表 4 龙塘山-顺溪坞不同植被类型下土壤养分特征
Table 4. Characteristics of soil fertility properties under different vegetation types in Longtangshan-Shunxiwu
植被类型 pH 养分/(g·kg−1) 有机质 全磷 全氮 全钾 落叶阔叶林 5.30±0.13 a 62.06±6.19 ab 0.20±0.05 a 1.51±0.18 ab 17.25±2.44 a 针阔叶混交林 4.93±0.19 a 67.48±8.71 ab 0.08±0.01 a 1.70±0.31 ab 14.86±2.11 a 针叶林 4.98±0.46 a 71.54±6.35 ab 0.07±0.02 a 1.72±0.06 ab 26.74±6.35 a 竹林 5.05±0.21 a 65.13±10.47 ab 0.11±0.04 a 1.20±0.25 b 17.61±5.32 a 草甸 4.57±0.11 a 82.89±7.54 a 0.07±0.01 a 2.08±0.28 a 20.41±4.97 a 农作物 5.23±0.35 a 46.63±7.78 b 0.18±0.03 a 1.09±0.19 b 18.11±2.31 a 说明:同养分不同字母表示不同植被类型之间差异显著(P<0.05);数值为平均值±标准差 -
本研究结果显示:龙塘山-顺溪坞区土壤中所有养分均属于中等程度变异,并且有机质变异系数最高,表明其易受结构因素和随机因素影响。空间变异主要分为结构因素变异和随机因素变异[7]。本研究中,土壤pH、土壤全磷的块基比分别为0.26、0.50,呈中等强度的空间自相关性,说明自然和人为因素共同对目标变量存在影响。土壤有机质、全氮、全钾的块基比分别为0.23、0.11、0.18,表现为强烈空间自相关,说明其变异主要受土壤母质、植被、地形等非人为因素影响。成土母质及其风化程度也是影响土壤养分分布状况的主要因素[21],但也有研究表明:母质对土壤有机质、全氮、有效磷质量分数的影响并无显著差异[22]。本研究区土壤母质在景观尺度上的变异较为平稳,主要是酸性岩石矿物,因此对母质因子不作重点分析。地形因子导致的水热分布差异是造成土壤属性空间异质性的重要原因[23−24]。森林土壤有机质空间分布受地形地貌等自然因素以及受其制约的土地利用的影响较大,受人类活动、耕作措施等随机因素影响很小[25]。由龙塘山-顺溪坞的DEM图可知:该研究区的高海拔区主要分布在西北和东南部,与土壤有机质和全氮高值区的空间分布特点具有较高的一致性。总体上,龙塘山-顺溪坞土壤养分质量分数较高,随地势的空间连续性尺度较小,分布随机化而且各土壤养分的空间变异较大,说明土壤养分的空间异质性可能受地形地势等自然因素的影响。
总体来看,该研究区的土壤养分质量分数均较高,但全钾、全磷质量分数相对较低,多数全磷、全钾低值区分布于海拔较高处,可能与该区受地形因子控制的降水量有关[26]。海拔升高到一定高度时,降水量的增加加强了磷、钾流失。可见高海拔处的植被生产力可能易受磷、钾养分限制。在该研究区南部海拔340~816 m处存在着大面积土壤pH、全磷、全钾质量分数较高而土壤有机质和全氮质量分数较低区域。这可能与该区域的新增农田有关。有研究表明:施肥会增加土壤酸性以及磷、钾质量分数[27]。与耕地相比,自然土地的人为干扰少且凋落物养分归还多,土壤有机质和全氮质量分数较高[28]。
-
土壤养分的空间异质性受地形地势等自然因素的影响[23],不同海拔带间的水热分布、温湿度差异等会影响土壤养分的迁移、转化与累积,海拔因素是影响森林土壤有机碳积累的主导因子之一[29]。本研究表明:土壤有机质和全氮均与海拔呈极显著正相关,有机质在1、2级海拔和3、4、5、6级海拔间均存在显著差异。张亚茹等[30]在鼎湖山研究表明:土壤有机质具有明显的空间变异性且随海拔升高而增加。两者的研究结果一致。在高海拔处的低温环境中,微生物活性降低而凋落物增加,一方面增加了土壤有机碳的归还与积累,另一方面减慢了有机质分解转化速率。本研究结果显示:有机质和全氮呈显著正相关,这与相关森林土壤研究的结果一致[24],并且有机质和全氮与海拔因子具有较为一致的相关性。总体上,在低海拔处,水、气、热等条件有利于加速土壤中物质循环,从而提高土壤养分的利用率。随着海拔升高,气温降低,湿度变大,植物蒸腾作用减弱,微生物对凋落物及动植物残体等的分解活动减弱,生物积累作用增强,造成有机质、全氮质量分数显著高于低海拔地区[11]。
本研究区中土壤全磷、全钾质量分数呈现出随海拔升高而下降的趋势,这可能与磷、钾的元素特性有关。在低山丘陵区,海拔较高的地方受水流的侵蚀作用较强,导致土壤强淋失,降低土壤中钾质量分数[31]。在自然生态系统中,与土壤有机质和全氮不同,土壤磷主要来源于基岩的风化[32],并通过植物的聚集作用积累在表层土壤。凋落物中的磷是表层土壤磷的主要来源[33]。随着海拔升高,气温和热量下降,凋落物的分解速率降低,阻碍了植物的聚集作用,减少了土壤中磷的输入。坡度是影响土壤养分分布的重要因素之一,它能够调节地表径流,改变土壤养分的空间分布[11]。本研究中土壤有机质和全氮与坡度呈极显著负相关,坡度增大,地表径流和侵蚀变强,地表的扰动作用减少了土壤有机质和全氮的质量分数。
本研究发现:土壤有机质和全氮质量分数随着容重增加而减少,这可能与受容重因子控制的土壤储水量有关。土壤储水量会随容重的减小而增加,因为随土壤容重的减小,土壤团粒结构和土壤有效孔隙增多,土壤结构得到改善,导致入渗能力、持水能力增大[28],有利于植物细根的穿插和发育,提升土壤养分循环,从而进一步提高土壤有机质回归能力和氮矿化速率[34]。本研究中,pH和全磷与土壤容重呈正相关。pH变化较慢,这可能是由于自然保护区的原始生态保存较为完好,研究区土壤酸性主要来自于空间连续性尺度较小的酸性土壤母岩。磷通过凋落物和根系分泌物等途径归还土壤。随土壤容重增大,土壤持水能力降低,凋落物和根系分泌物的生物化学反应加剧,可促进磷元素的溶解与周转[33−35]。研究发现:土壤pH和全磷与土壤有机质呈负相关[27]。本研究区土壤整体呈酸性,土壤有机质随着容重减少而增加,同时土壤pH和全磷相应降低。总体上,随着土壤容重的变化,有机质变化较大,而全氮、全磷的变化则相反,这与有机质高值区分布较为集中而低值区分散,全氮和全磷分布较离散的研究结果一致。
植物体养分通过凋落物的归还和植物根系的输送直接影响土壤养分质量分数及分布。本研究结果显示:不同植被类型对土壤有机质和全氮的影响达显著水平,草甸土壤中有机质和全氮质量分数最高。相关研究发现:草甸土中有机质质量分数高于针叶林中[36],浙江省乌岩岭不同植被类型土壤中有机质质量分数从大到小依次为针阔混交林、竹林、农地[37]。本研究结果与相关学者的研究结果较为一致。海拔可通过改变植被覆盖度、草地生产力等来实现对表层土壤有机质的影响。水、气、热条件随海拔变化,而这些自然条件制约着植被类型,影响着植被的养分消耗与归还,进而影响植物根系对土壤有机碳的输送量。同时,温、湿度等环境条件影响微生物对表层土壤凋落物的分解和转化。本研究中,草甸大面积分布在高海拔区域,随着海拔高度的增加,气温下降,使得有机质分解转化速率减慢,而草地覆盖度及有机碳输送能力增加,有机质易于积累。刘跃建等[38]研究表明:随海拔升高,草甸土中全氮质量分数明显高于其他森林植被类型土壤中的全氮,且全氮与有机质对环境因子的响应具有一致性。植物体养分循环是土壤全钾质量分数的主要决定过程之一。相对于其他植被类型,针叶林土壤全钾质量分数较高,可能是因为针叶林凋落物对土壤全钾有更明显的富集作用。
-
清凉峰国家级自然保护区内龙塘山-顺溪坞土壤养分全磷、全钾质量分数相对较低,各土壤养分均表现为中等程度变异,具有较为明显的空间异质性。养分低值区普遍以斑状散布在研究区中,土壤有机质、全氮的高值区集中分布于西北及东南部分区域,而全磷、全钾的高值区分布较为分散。总体上,土壤养分空间分布较为破碎化,斑块特征显著。受结构性因素影响,土壤有机质和全氮与海拔、坡度、容重、植被类型的相关性达显著水平,全磷仅与容重的相关性达显著水平。研究区内土壤酸性较强,同时受地形地势的影响,研究区内土壤存在水土流失及土壤养分淋失风险,建议注重高海拔区水土保持。
Spatial heterogeneity of soil nutrients and the influencing factors in Zhejiang subtropical nature reserve
-
摘要:
目的 揭示亚热带国家自然保护区的土壤养分空间变异特征及其影响因子 , 为中国亚热带丘陵地区的珍稀植被和天然生态系统的保护提供理论依据。 方法 以浙江清凉峰国家级自然保护区内龙塘山-顺溪坞的土壤养分为研究对象,采用经典统计学和地统计学方法对龙塘山-顺溪坞土壤的pH、有机质、全氮、全磷、全钾质量分数进行了系统分析,并探究了其主要影响因子。 结果 龙塘山-顺溪坞保护区内土壤整体呈酸性,有机质、全磷、全氮、全钾质量分数平均值分别为65.04、0.67、1.63 、18.45 g·kg−1,均属于中等程度变异。半方差分析结果表明:土壤有机质、全氮、全钾表现为强烈空间自相关性,土壤pH、全磷表现为中等空间自相关性,说明研究区土壤养分主要受结构性因素影响。全局Moran’s I指数显示:土壤有机质的空间自相关达到显著水平,其空间分布较为聚集,而土壤pH分布趋于随机性。Kriging插值结果显示:多种养分的空间分布较为碎片化,斑块特征显著。相关性分析显示:土壤有机质和全氮与海拔、容重、坡度呈显著相关(P<0.05),并且有机质和全氮之间呈极显著正相关(P<0.01)。 结论 4种土壤养分均表现出较为明显的空间异质性,海拔和容重分别与研究区土壤有机质和全氮空间变异的相关性较强。图6表4参38 Abstract:Objective This study aims to reveal the spatial variability of soil nutrients and the influencing factors in subtropical national nature reserves of China, so as to provide a theoretical basis for protecting rare vegetation and natural ecosystems in China’s subtropical hilly regions. Method Soil samples were collected from Longtangshan-Shunxiwu in Qingliangfeng Nature Reserve of Zhejiang Province. Classical statistics and geostatistical method were used to systematically analyze the soil variables such as pH, soil organic matter (SOM), total nitrogen (TN), total phosphorus (TP) and total potassium (TK), and the main influencing factors were explored. Result The soil in the study area was acidic as a whole. The average contents of SOM, TP, TN and TK were 65.04, 0.67, 1.63 and 18.45 g·kg−1, respectively, which belonged to moderate variation. Semi variance analysis showed that SOM, TN and TK displayed strong spatial autocorrelation while pH and TP displayed moderate spatial autocorrelation, indicating that the soil nutrients were mainly affected by structural factors. The global Moran’s I index revealed that the spatial autocorrelation of SOM reached a significant level and its spatial distribution was aggregated, while the distribution of soil pH tended to be random. Kriging interpolation results showed that the spatial distribution of various nutrients was fragmented and the patch characteristics were significant. Correlation analysis showed that SOM and TN were significantly correlated with altitude, bulk density, and slope (P<0.05), and there was a significant positive correlation between SOM and TN (P<0.01). Conclusion All the soil nutrients show an obvious spatial heterogeneity. Altitude and bulk density are strongly correlated with the spatial variation of SOM and TN in the study area. [Ch, 6 fig. 4 tab. 38 ref.] -
Key words:
- subtropical hilly region /
- nature reserve /
- soil nutrients /
- spatial variation /
- influencing factors
-
图 1 研究区样点分布及海拔信息示意图
Figure 1 Spatial distribution and elevation information of soil sampling sites
图 2 龙塘山-顺溪坞土壤养分半变异函数分析
Figure 2 Semivariograms of soil fertility properties in Longtangshan-Shunxiwu
图 3 龙塘山-顺溪坞土壤养分空间分布示意图
Figure 3 Spatial distribution map of soil fertility properties in Longtangshan-Shunxiwu
图 4 龙塘山-顺溪坞土壤养分与环境因子的相关分析
Figure 4 Correlation analysis between soil nutrients and environmental factors in Longtangshan-Shunxiwu
图 5 土壤有机质与土壤全氮的关系
Figure 5 Relationship between soil organic matter and total nitrogen
图 6 龙塘山-顺溪坞不同海拔土壤养分特征
Figure 6 Characteristics of soil fertility properties under different elevations in Longtangshan-Shunxiwu
表 1 龙塘山-顺溪坞土壤养分描述性统计特征
Table 1. Descriptive statistical characteristics of soil fertility properties in Longtangshan-Shunxiwu
指标 pH 有机质/(g·kg−1) 全磷/(g·kg−1) 全氮/(g·kg−1) 全钾/(g·kg−1) 最小值 3.82 10.24 0.24 0.35 5.96 最大值 6.70 143.43 1.53 3.06 41.54 均值 5.10 65.04 0.67 1.63 18.45 标准差 0.59 32.61 0.27 0.60 8.73 变异系数/% 12 50 40 37 47 偏度 0.56 −0.27 0.14 0.37 −0.09 峰度 0.07 0.62 −0.30 −0.31 −0.66 K-S 0.20 0.20 0.20 0.20 0.20 分布类型 正态分布 对数正态分布 对数正态分布 正态分布 对数正态分布  下载: 导出CSV
下载: 导出CSV
表 2 土壤养分半变异模型参数及IN指数
Table 2. Semivariogram model parameters of soil fertility properties and global IN index
指标 模型 块金值C0 基台值(C0+C) 块基比[C0/(C0+C)] IN Z 变程/km 决定系数(R2) pH 指数模型 0.10 0.38 0.26 0.08 1.35 2.00 0.76 有机质 指数模型 250.00 1084.00 0.23 0.24** 3.29 5.00 0.72 全磷 指数模型 0.04 0.08 0.50 0.16* 2.31 26.00 0.90 全氮 高斯模型 0.04 0.36 0.11 0.04 0.66 796.74 0.82 全钾 高斯模型 57.90 326.70 0.18 <0.05 −0.52 354.20 0.93 说明:*表示在0.05水平上显著相关(|Z|>1.96);**表示在0.01水平上显著相关(|Z|>2.58)
下载: 导出CSV
表 3 龙塘山-顺溪坞土壤化学性质随土壤容重的变化
Table 3. Changes of soil chemical properties with soil bulk density in Longtangshan-Shunxiwu
容重/(g·cm−3) pH 有机质/(g·kg−1) 全磷/(g·kg−1) 全氮/(g·kg−1) 容重/(g·cm−3) pH 有机质/(g·kg−1) 全磷/(g·kg−1) 全氮/(g·kg−1) 0.8~0.9 4.61±0.68 79.41±20.73 0.11±0.04 2.29±0.90 1.4~1.5 5.37±0.58 59.48±19.48 0.12±0.04 1.26±0.59 0.9~1.0 5.29±1.13 80.81±5.40 0.07±0.03 1.81±0.11 1.5~1.6 5.13±0.75 52.82±14.42 0.17±0.10 1.04±0.32 1.0~1.1 4.87±0.39 74.86±18.27 0.11±0.06 1.89±0.47 1.6~1.7 5.17±0.68 41.08±10.31 0.14±0.08 0.91±0.13 1.1~1.2 5.03±0.76 71.68±18.62 0.10±0.03 1.72±0.67 1.7~1.8 5.80±0.45 42.50±13.67 0.39±0.43 0.80±0.29 1.2~1.3 4.96±0.31 65.44±20.67 0.10±0.05 1.42±0.55 F 5.13 36.69 11.01 41.04 1.3~1.4 5.17±0.57 46.44±11.63 0.15±0.09 1.08±0.33 P 0.03 <0.001 0.001 <0.001 说明:数值为平均值±标准差
下载: 导出CSV
表 4 龙塘山-顺溪坞不同植被类型下土壤养分特征
Table 4. Characteristics of soil fertility properties under different vegetation types in Longtangshan-Shunxiwu
植被类型 pH 养分/(g·kg−1) 有机质 全磷 全氮 全钾 落叶阔叶林 5.30±0.13 a 62.06±6.19 ab 0.20±0.05 a 1.51±0.18 ab 17.25±2.44 a 针阔叶混交林 4.93±0.19 a 67.48±8.71 ab 0.08±0.01 a 1.70±0.31 ab 14.86±2.11 a 针叶林 4.98±0.46 a 71.54±6.35 ab 0.07±0.02 a 1.72±0.06 ab 26.74±6.35 a 竹林 5.05±0.21 a 65.13±10.47 ab 0.11±0.04 a 1.20±0.25 b 17.61±5.32 a 草甸 4.57±0.11 a 82.89±7.54 a 0.07±0.01 a 2.08±0.28 a 20.41±4.97 a 农作物 5.23±0.35 a 46.63±7.78 b 0.18±0.03 a 1.09±0.19 b 18.11±2.31 a 说明:同养分不同字母表示不同植被类型之间差异显著(P<0.05);数值为平均值±标准差
下载: 导出CSV
-
[1] 高郯, 李江荣, 卢杰, 等. 色季拉山急尖长苞冷杉林不同坡向土壤养分及肥力研究[J]. 生态学报, 2020, 40(4): 1331 − 1341. GAO Tan, LI Jiangrong, LU Jie, et al. Soil nutrient and fertility of different slope directions in the Abies georgei var. smithii forest in Sejila Mountain [J]. Acta Ecologica Sinica, 2020, 40(4): 1331 − 1341. [2] QIN Zihan. Research on the spatial variability of soil nutrients prediction based on GIS [J]. Advanced Materials Research, 2013, 791/793: 1681 − 1685. [3] FU Weihun, TUNNEY H, ZHANG Chaosheng. Spatial variation of soil nutrients in a dairy farm and its implications for site-specific fertilizer application [J]. Soil &Tillage Research, 2010, 106(2): 185 − 193. [4] FU Weijun, ZHANG Keli, ZHANG Chaosheng, et al. Using Moran’ s I and geostatistics to identify spatial patterns of soil nutrients in two different long-term phosphorus-application plots [J]. Journal of Plant Nutrition and Soil Science, 2011, 174(5): 785 − 798. [5] 王淑彬, 徐慧芳, 宋同清, 等. 广西森林土壤主要养分的空间异质性[J]. 生态学报, 2014, 34(18): 5292 − 5299. WANG Shubin, XU Huifang, SONG Tongqing, et al. Spatial heterogeneity of the main nutrients in Guangxi forest soils [J]. Acta Ecologica Sinica, 2014, 34(18): 5292 − 5299. [6] 柳丽娜, 金爱武. 集约经营毛竹林土壤养分空间变异特征初探[J]. 浙江农林大学学报, 2011, 28(5): 828 − 832. LIU Lina, JIN Aiwu. Spatial variability of soil nutrients for an intensively managed Phyllostachys pubescens forest [J]. Journal of Zhejiang A&F University, 2011, 28(5): 828 − 832. [7] 邹佳何, 王海燕, 张美娜, 等. 温带云冷杉针阔混交林土壤养分的空间分布特征及影响因素[J]. 应用与环境生物学报, 2021, 27(6): 1554 − 1562. ZOU Jiahe, WANG Haiyan, ZHANG Meina, et al. Spatial distribution characteristics and influence factors of soil nutrients in temperate mixed spruce-fir coniferous and broadleaf forests [J]. Chinese Journal of Applied and Environmental Biology, 2021, 27(6): 1554 − 1562. [8] FU Weijun, DONG Jiaqi, DING Lizhong, et al. Spatial correlation of nutrients in a typical soil-hickory system of southeastern China and its implication for site-specific fertilizer application[J/OL]. Soil Tillage Research, 2022, 217: 105265[2022-02-09]. doi: 10.1016/j.still.2021.105265. [9] 董佳琦, 张勇, 傅伟军, 等. 香榧主产区林地土壤养分空间异质性及其肥力评价[J]. 生态学报, 2021, 41(6): 2292 − 2304. DONG Jiaqi, ZHANG Yong, FU Weijun, et al. Spatial variation of soil nutrients and evaluation of integrated soil fertility in Torreya grandis cv. Merrillii region [J]. Acta Ecologica Sinica, 2021, 41(6): 2292 − 2304. [10] 牛文鹏, 李青圃, 李铖, 等. 珠江三角洲土壤养分多尺度空间分异及环境驱动力[J]. 生态环境学报, 2021, 30(4): 743 − 755. NIU Wenpeng, LI Qingpu, LI Cheng, et al. Multi-scale spatial variability and environmental drivers of soil nutrient distributions in the Pearl River Delta, South China [J]. Ecology and Environmental Sciences, 2021, 30(4): 743 − 755. [11] 李超, 李文峰. 高原耕地土壤养分空间分布与影响因子相关性研究[J]. 土壤通报, 2014, 45(5): 1113 − 1118. LI Chao, LI Wenfeng. Study on the relations between the spatial distribution of plateau cultivated soil nutrients and impact factors [J]. Chinese Journal of Soil Science, 2014, 45(5): 1113 − 1118. [12] 姜霓雯, 童根平, 叶正钱, 等. 浙江清凉峰自然保护区土壤肥力指标空间变异及其影响因素[J]. 生态学报, 2022, 42(6): 1 − 12. JIANG Niwen, TONG Genping, YE Zhengqian, et al. Spatial variability of soil fertility properties and its affecting factors of Qingliangfeng Nature Reserve, Zhejiang [J]. Acta Ecologica Sinica, 2022, 42(6): 1 − 12. [13] 鲍士旦. 土壤农化分析[M]. 3版. 北京: 中国农业出版社, 2000. BAO Shidan. Soil Agrochemical Analysis[M]. 3rd ed. Beijing: China Agriculture Press, 2000. [14] FU Weijun, ZHAO Keli, JIANG Peikun, et al. Field-scale variability of soil test phosphorus and other nutrients in grasslands under long-term agricultural managements [J]. Soil Research, 2013, 51(6): 503 − 512. [15] 李龙, 姚云峰, 秦富仓, 等. 赤峰市黄花甸子流域土壤有机碳含量的空间变异特征研究[J]. 环境科学学报, 2014, 34(3): 742 − 748. LI Long, YAO Yunfeng, QIN Fucang, et al. Spatial variations of organic carbon of Huanghuadianzi watershed in Chifeng [J]. Acta Scientiae Circumstantiae, 2014, 34(3): 742 − 748. [16] 宋轩, 李立东, 寇长林, 等. 黄水河小流域土壤养分分布及其与地形的关系[J]. 应用生态学报, 2011, 22(12): 3163 − 3168. SONG Xuan, LI Lidong, KOU Changlin, et al. Soil nutrient distribution and its relations with topography in Huangshui River drainage basin [J]. Chinese Journal of Applied Ecology, 2011, 22(12): 3163 − 3168. [17] 陈涛, 常庆瑞, 刘京, 等. 黄土高原南麓县域耕地土壤速效养分时空变异[J]. 生态学报, 2013, 33(2): 554 − 564. CHEN Tao, CHANG Qingrui, LIU Jing, et al. Temporal and spatial variability of soil available nutrients in arable lands of Heyang County in South Loess Plateau [J]. Acta Ecologica Sinica, 2013, 33(2): 554 − 564. [18] 张福锁. 测土配方施肥技术要览[M]. 北京: 中国农业大学出版社, 2006. ZHANG Fusuo. Technical Overview of Soil testing and Formula Fertilization[M]. Beijing: China Agricultural University Press, 2006. [19] 王政权, 王庆成. 森林土壤物理性质的空间异质性研究[J]. 生态学报, 2000, 20(6): 945 − 950. WANG Zhengquan, WANG Qingcheng. The spatial heterogeneity of soil physical properties in forests [J]. Acta Ecologica Sinica, 2000, 20(6): 945 − 950. [20] TEFERI T, ROBERT L H. Intensive tillage effects on spatial variability of soil test, plant growth, and nutrient uptake measurements [J]. Soil Science, 1998, 163(2): 155 − 165. [21] 郭旭东, 傅伯杰, 陈利顶, 等. 低山丘陵区土地利用方式对土壤质量的影响——以河北省遵化市为例[J]. 地理学报, 2001, 56(4): 447 − 455. GUO Xudong, FU Bojie, CHEN Liding, et al. Effects of land use patterns on soil quality in low hilly region: a case study of Zunhua City, Hebei Province [J]. Acta Geographica Sinica, 2001, 56(4): 447 − 455. [22] 杜可, 王乐, 张淑香, 等. 黑土区县域土壤养分空间分布特征及其影响因子[J]. 植物营养与肥料学报, 2018, 24(6): 1465 − 1474. DU Ke, WANG Le, ZHANG Shuxiang, et al. Spatial distribution characteristics and influencing factors of soil nutrients in black soil region counties [J]. Journal of Plant Nutrition and Fertilizers, 2018, 24(6): 1465 − 1474. [23] SUMFLETH K, DUTTMANN R. Prediction of soil property distribution in paddy soil landscapes using terrain data and satellite information as indicators [J]. Ecological Indicators, 2008, 8(5): 485 − 501. [24] 赵青, 刘爽, 陈凯, 等. 武夷山自然保护区不同海拔甜槠天然林土壤有机碳变化特征及影响因素[J]. 生态学报, 2021, 41(13): 5328 − 5339. ZHAO Qing, LIU Shuang, CHEN Kai, et al. Change characteristics and influencing factors of soil organic carbon in Castanopsis eyrei natural forests at different altitudes in Wuyishan Nature Reserve [J]. Acta Ecologica Sinica, 2021, 41(13): 5328 − 5339. [25] 苗娟, 周传艳, 李世杰, 等. 不同林龄云南松林土壤有机碳和全氮积累特征[J]. 应用生态学报, 2014, 25(3): 625 − 631. MIAO Juan, ZHOU Chuanyan, LI Shijie, et al. Accumulation of soil organic carbon and total nitrogen in Pinus yunnanensis forests at different age stages [J]. Chinese Journal of Applied Ecology, 2014, 25(3): 625 − 631. [26] XU Zhonglin, CHANG Yapeng, LI Lu, et al. Climatic and topographic variables control soil nitrogen, phosphorus, and nitrogen: phosphorus ratios in a Picea schrenkiana forest of the Tianshan Mountains [J]. PLoS One, 2018, 13(11): 204 − 130. [27] 武小钢, 郭晋平, 杨秀云, 等. 芦芽山典型植被土壤有机碳剖面分布特征及碳储量[J]. 生态学报, 2011, 31(11): 3009 − 3019. WU Xiaogang, GUO Jinping, YANG Xiuyun, et al. Soil organic carbon storage and profile inventory in the different vegetation types of Luya Mountain [J]. Acta Ecologica Sinica, 2011, 31(11): 3009 − 3019. [28] 李青华, 张静, 王力, 等. 黄土丘陵沟壑区山地苹果林土壤干化及养分变异特征[J]. 土壤学报, 2018, 55(2): 503 − 514. LI Qinghua, ZHANG Jing, WANG Li, et al. Desiccation and nutrient status of the soil in apple orchards in hilly-gully region of the Loess Plateau [J]. Acta Pedologica Sinica, 2018, 55(2): 503 − 514. [29] NIE Xiaodong, GUO Wang, HUANG Bin, et al. Effects of soil properties, topography and landform on the understory biomass of a pine forest in a subtropical hilly region [J]. Catena, 2019, 176: 104 − 111. [30] 张亚茹, 欧阳旭, 褚国伟, 等. 鼎湖山季风常绿阔叶林土壤有机碳和全氮的空间分布[J]. 应用生态学报, 2014, 25(1): 19 − 23. ZHANG Yaru, OUYANG Xu, CHU Guowei, et al. Spatial heterogeneity of soil organic carbon and total nitrogen in a monsoon evergreen broadleaf forest in Dinghushan, Guangdong, China [J]. Chinese Journal of Applied Ecology, 2014, 25(1): 19 − 23. [31] 杨家慧, 谭伟, 冯艳. 马尾松人工林土壤养分空间分布特征及其与地形因子的相关性分析[J]. 西南林业大学学报(自然科学), 2020, 40(4): 23 − 29. YANG Jiahui, TAN Wei, FENG Yan. Spatial distribution characteristics of soil nutrients and the correlation with topographic factors in Pinus massoniana [J]. Journal of Southwest Forestry University (Natural Sciences) , 2020, 40(4): 23 − 29. [32] 张伟, 刘淑娟, 叶莹莹, 等. 典型喀斯特林地土壤养分空间变异的影响因素[J]. 农业工程学报, 2013, 29(1): 93 − 101. ZHANG Wei, LIU Shujuan, YE Yingying, et al. Spatial variability of soil nutrients and influencing factors in typical karst virgin forest [J]. Transactions of the Chinese Society of Agricultural Engineering, 2013, 29(1): 93 − 101. [33] 刘丛强. 生物地球化学过程与地表物质循环——西南喀斯特土壤-植被系统生源要素循环[M]. 北京: 科学出版社, 2009. LIU Congqiang. Biogeochemical Processes and Surface Material Cycling: Biogenic Elements Cycling of Soil-vegetation System in Karst Region of Southwest China [M]. Beijing: Science Press, 2009. [34] 刘潘伟, 高鹏, 刘晓华, 等. 大岗山流域土壤碳氮要素空间分布特征及影响因素[J]. 中国水土保持科学, 2018, 16(2): 73 − 79. LIU Panwei, GAO Peng, LIU Xiaohua, et al. Spatial distribution and influential factors of soil carbon and nitrogen in Dagangshan Watershed [J]. Science of Soil and Water Conservation, 2018, 16(2): 73 − 79. [35] 张岩松, 雷泽勇, 于东伟, 等. 沙质草地营造樟子松林后土壤容重的变化及其影响因子[J]. 生态学报, 2019, 39(19): 7144 − 7152. ZHANG Yansong, LEI Zeyong, YU Dongwei, et al. Changes in soil bulk density and its influencing factors after sandy grassland afforestation with Pinus sylvestris var. mongolica [J]. Acta Ecologica Sinica, 2019, 39(19): 7144 − 7152. [36] 杨玉海, 陈亚宁, 李卫红, 等. 伊犁河谷不同植被带下土壤有机碳分布[J]. 地理学报, 2010, 65(5): 605 − 612. YANG Yuhai, CHEN Yaning, LI Weihong, et al. Soil organic carbon distribution under different vegetation belts in Ili River Valley [J]. Acta Geographica Sinica, 2010, 65(5): 605 − 612. [37] 王阳, 章明奎. 不同类型林地土壤颗粒态有机碳和黑碳的分布特征[J]. 浙江大学学报(农业与生命科学版), 2011, 37(2): 193 − 202. WANG Yang, ZHANG Mingkui. Distribution characters of particulate organic carbon and black carbon in soils under different forestry vegetations [J]. Journal of Zhejiang University (Agriculture and Life Sciences) , 2011, 37(2): 193 − 202. [38] 刘跃建, 李强, 马明东. 四川西北部主要森林植被类型土壤养分库比较研究[J]. 水土保持学报, 2010, 24(5): 146 − 152. LIU Yuejian, LI Qiang, MA Mingdong. Comparison of soil nutrient storage of forest vegetation type in northwest of Sichuan Province [J]. Journal of Soil and Water Conservation, 2010, 24(5): 146 − 152. -

-
链接本文:
https://zlxb.zafu.edu.cn/article/doi/10.11833/j.issn.2095-0756.20220213
 点击查看大图
点击查看大图

计量
- 文章访问数: 1874
- HTML全文浏览量: 330
- PDF下载量: 99
- 被引次数: 0



