




-
松阿扁叶蜂Acantholyda posticalis是危害油松Pinus tabuliformis、赤松P. densiflora、华山松P. armandii等松类的主要食叶害虫[1]。该虫1 a发生1代,越冬幼虫4月下旬在地下化蛹,5月初成虫羽化,6月上、中旬幼虫在树上取食松针,7月下旬老熟幼虫在树冠下方吐丝下树进入土壤表层做土室越夏、越冬[2]。该幼虫危害严重时,针叶全无,只留下叶柄,受灾后的油松枝梢上布满残渣和粪屑,林分似火烧,松林成橘褐色,连续受害2~3 a,林木会干枯死亡[3]。昆虫作为森林生态系统的重要有机组成部分,其种群暴发受森林植被及环境的直接或间接影响[4−5],对系统内的物种组成、空间结构、生态功能产生极大的负面作用[6−7]。影响森林害虫种群进一步扩散暴发的因素既包括植物、动物、微生物等生物因子,也包括海拔、坡向、坡位等非生物因子[8−9]。
在森林生态系统中,林分因子和立地因子与森林虫害关系密切[10−11]。林分因子和立地因子与有害生物共同建立了相互依存和制约的关系,特别是林分因子中森林群落结构作用于食叶害虫来调控害虫的种群密度。如中华松针蚧Matsucoccus sinensis在低密度油松林中种群数量多,高密度林分低;枝下高较高的油松受中华松针蚧危害严重,枝下高较低的油松林受危害程度低[12]。立地因子通常是以影响寄主植物的质量来调控害虫种群密度。如WERNER等[13]认为:随坡度的减小,土壤厚度和养分储量会增加,有利于叶片中的营养物质升高,从而促进森林虫害的发生。苹果小吉丁虫Agrilus mali虫口密度与坡度、海拔呈显著负相关,与林木胸径呈显著正相关[14]。因此,加快森林虫害生态调控理论和可操作体系的系统研究显得尤为迫切,急需在生态系统的尺度上开展林分因子和立地因子与有害生物关系的研究。通过调整林分结构来实现对害虫的生态调控,是一项重要且可持续的森防措施。本研究以宁夏罗山国家级自然保护区林区的食叶害虫松阿扁叶蜂幼虫为研究对象,开展罗山森林生态系统林分因子和立地因子对松阿扁叶蜂幼虫种群数量影响的研究,旨在探讨该幼虫平均虫口密度与各因子间多重复杂的关系,揭示影响罗山区域松阿扁叶蜂幼虫平均虫口密度的关键因子,为控制松阿扁叶蜂提供依据。
-
宁夏罗山国家级自然保护区(37°11′~37°25′N,106°04′~106°24′E)位于宁夏回族自治区红寺堡区,地处贺兰山和六盘山之间的过渡带,是宁夏三大天然林区之一。保护区海拔为1 560.0~2 624.5 m,由呈南北走向的大、小罗山2个山体构成,南北长36 km,东西宽18 km。保护区属典型温带大陆性气候,温差大,降水少,蒸发量是降水量的9倍;全年主风向为东南风,其次西北风,四季分明,且冬春长、夏秋短。土壤类型单一,有普通山地灰褐土、淡灰钙土、风沙土和山地侵蚀灰褐土。森林覆盖率为9.6%,总蓄积量2.03×105 m3,乔木以油松和青海云杉Picea crassifolia为主,主要灌木有灰栒子Cotoneaster acutifolius、银露梅Potentilla glabra、紫丁香Lilium tenuifolium、榆树Ulmus pumila、水栒子Cotoneaster multiflorus、甘肃山楂Lonicera tangutica、黄刺玫Caragana frutex、绣线菊Spiraea salicifolia和虎榛子Ostryopsis davidiana;主要地被植物有蓬子菜Galium verum、小花草玉梅Anemone rivularis、大戟Euphorbia pekinensis、四叶葎Galium bungei、山丹Lilium pumilum等。
-
于2022和2023年6—9月,对研究区进行实地调查。根据林木分布海拔,随机设立有代表性林分作为样地,其中不同坡向(半阳坡、阴坡、半阴坡)、坡位(上坡、中坡和下坡)、坡度(16°≤坡度<26°为斜坡,26°≤坡度<36°为陡坡,36°≤坡度<46°为急坡)的样地至少重复3次,共21块。每块标准样地面积为20 m×20 m,同一坡向保留10 m以上保护行,同一坡位保留20 m以上隔离带。用全球定位系统(GPS)获得地理坐标。调查每块样地松阿扁叶蜂幼虫数量,计算虫口密度。另外,调查标准样地林分和立地因子。样地基本信息如表1所示。
表 1 调查样地基本信息
Table 1. Basic information of the sample site
样地号 海拔/m 坡度/(°) 坡向 坡位 优势树种 林分类型 样地号 海拔/m 坡度/(°) 坡向 坡位 优势树种 林分类型 1 2286.08 34.0 4 2 油松 纯林 12 2324.73 25.0 6 2 油松 纯林 2 2290.61 33.0 4 1 油松 纯林 13 2322.72 24.0 6 1 油松 纯林 3 2227.39 42.0 5 1 油松 纯林 14 2315.60 23.5 4 2 青海云杉+油松 混交林 4 2217.63 40.0 5 2 油松 纯林 15 2323.49 24.5 5 1 油松 纯林 5 2196.95 33.0 5 3 油松 纯林 16 2307.99 25.0 5 2 油松 纯林 6 2201.75 30.0 5 3 油松 纯林 17 2418.00 29.5 6 2 青海云杉+油松 混交林 7 2390.78 46.0 5 1 油松 纯林 18 2415.03 30.0 6 2 青海云杉+油松 混交林 8 2383.45 37.3 5 2 油松 纯林 19 2431.00 36.0 6 3 青海云杉 纯林 9 2345.23 36.5 5 3 油松 纯林 20 2432.89 37.0 6 3 青海云杉 纯林 10 2397.55 41.0 6 1 油松 纯林 21 2496.97 33.0 6 2 青海云杉 纯林 11 2335.56 39.0 6 1 油松 纯林 -
在松阿扁叶蜂幼虫危害时,按“五点法”在每块样地4个角及中心各取1株油松,共5株,进行虫口密度调查,按树冠东、西、南、北4个方位的上、中、下3层各选1条50 cm延长枝,共选12条,记录虫口数,以“头·株−1”为统计单位,计算平均虫口密度,并统计这5株松树的树高、枝下高、胸径、冠幅和林下草本盖度,计算各因子平均值。按“五点法”记录标准样地海拔、坡位、坡向、坡度和腐殖层厚度,坡位、坡向等定性因子参考邵方丽等[15]的方法将其赋值量化,即用1、2、3分别代表上坡、中坡和下坡,用4、5、6分别代表半阳坡、阴坡、半阴坡。郁闭度采用对角线法测定,在每块样地每条对角线上,隔1 m设1个测点,即每条对角线上分别设置28个测点,共设置56个测点,采用抬头法观测每个测点是否被树冠所遮盖,分别记录为覆盖或无覆盖,最后统计标准地内设置的总观测点数和有树冠覆盖的点数,据此计算郁闭度[16]。每个标准样地调查胸径≥5 cm的树种数和株数,计算每个样地林分密度;按“五点法”设置5个5 m×5 m的灌木样方,每个灌木样方设置1个1 m×1 m草本植物样方,进行林下灌木层种类、数量和草本层盖度的调查。
根据周润惠等[17]采用的方法计算灌木物种丰富度和灌木Shamnon多样性指数。
-
采用Excel 2007和SPSS 22.0对数据进行分析。对松阿扁叶蜂幼虫平均虫口密度与林分因子和立地因子进行逐步回归分析,筛选出影响松阿扁叶蜂幼虫平均虫口密度的关键影响因子。单因素方差分析(one-way ANOVA)、Duncan法多重比较、Pearson相关系数法探究松阿扁叶蜂平均虫口密度与林分因子和立地因子的关联性。用GraphPad Prism 10和Origin 2021作图。
-
为了同时表达林分因子和立地因子对松阿扁叶蜂幼虫危害情况,对数据进行多重回归分析,建立幼虫平均虫口密度与各因子的方程,设幼虫平均虫口密度(y)、树种(x1)、林分密度(x2)、郁闭度(x3)、树高(x4)、枝下高(x5)、冠幅(x6)、胸径(x7)、草本盖度(x8)、灌木丰富度(x9)、灌木多样性指数(x10)、海拔(x11)、腐殖层厚度(x12)、坡度(x13)、坡向(x14)、坡位(x15)为自变量,得到回归模型:y=−319.750−7.936x1+75.212x3−0.176x4−0.804x5+8.621x6+0.128x7−112.264x8−6.284x9+62.243x10+0.163x11−4.877x12−0.573x13−3.969x14−5.645x15,决定系数(R2)为0.941,调整后R2=0.907。从表2可以看出:F值为27.796,P<0.001,表明各因子多元线性模式的分析效果好。
表 2 林分因子和立地因子与松阿扁叶蜂幼虫平均虫口密度的方差分析
Table 2. Analysis of variance between stand and site factors and average population density of A. posticalis larvae
误差来源 平方和 自由度 均方 F P 回归 37 135.254 15 2 475.684 27.796 <0.001 残差 2 315.750 26 89.067 总计 39 451.004 41 -
如表3所示:通过逐步回归分析,筛选出了郁闭度(x3)、冠幅(x6)、草本盖度(x8)及坡位(x15)等4个因子是影响松阿扁叶蜂幼虫平均虫口密度的关键因子。建立关键因子影响平均虫口密度的多元线性预测模型为:y=22.832+54.377x3+14.279x6−195.339x8−6.531x15,R2=0.856,调整后R2=0.840。分析结果表明:郁闭度、冠幅、草本盖度及坡位与松阿扁叶蜂幼虫平均虫口密度间存在极显著的线性关系(P<0.01)。4个关键因子组成的线性决定系数(R2=0.840)与上述15个因子影响幼虫平均虫口密度的决定系数接近(R2=0.907),说明这4个因子能够较准确地预测松阿扁叶蜂幼虫平均虫口密度的变化。F=54.847,P<0.001,郁闭度、冠幅、草本盖度和坡位的膨胀因子(VIF)分别为1.377、1.480、1.483和1.610,均远小于10,说明各项因子间没有高度共线性,多元线性模式的分析效果好。
表 3 平均虫口密度与关键因子逐步回归分析
Table 3. Average insect population density and key factors were analyzed by stepwise regression
因子 系数a 系数bi F P R2 膨胀因子 郁闭度(x3) 22.832 54.377 54.847 <0.001 0.840 1.377 冠幅(x6) 14.279 1.480 草本盖度(x8) −195.339 1.483 坡位(x15) −6.531 1.610 -
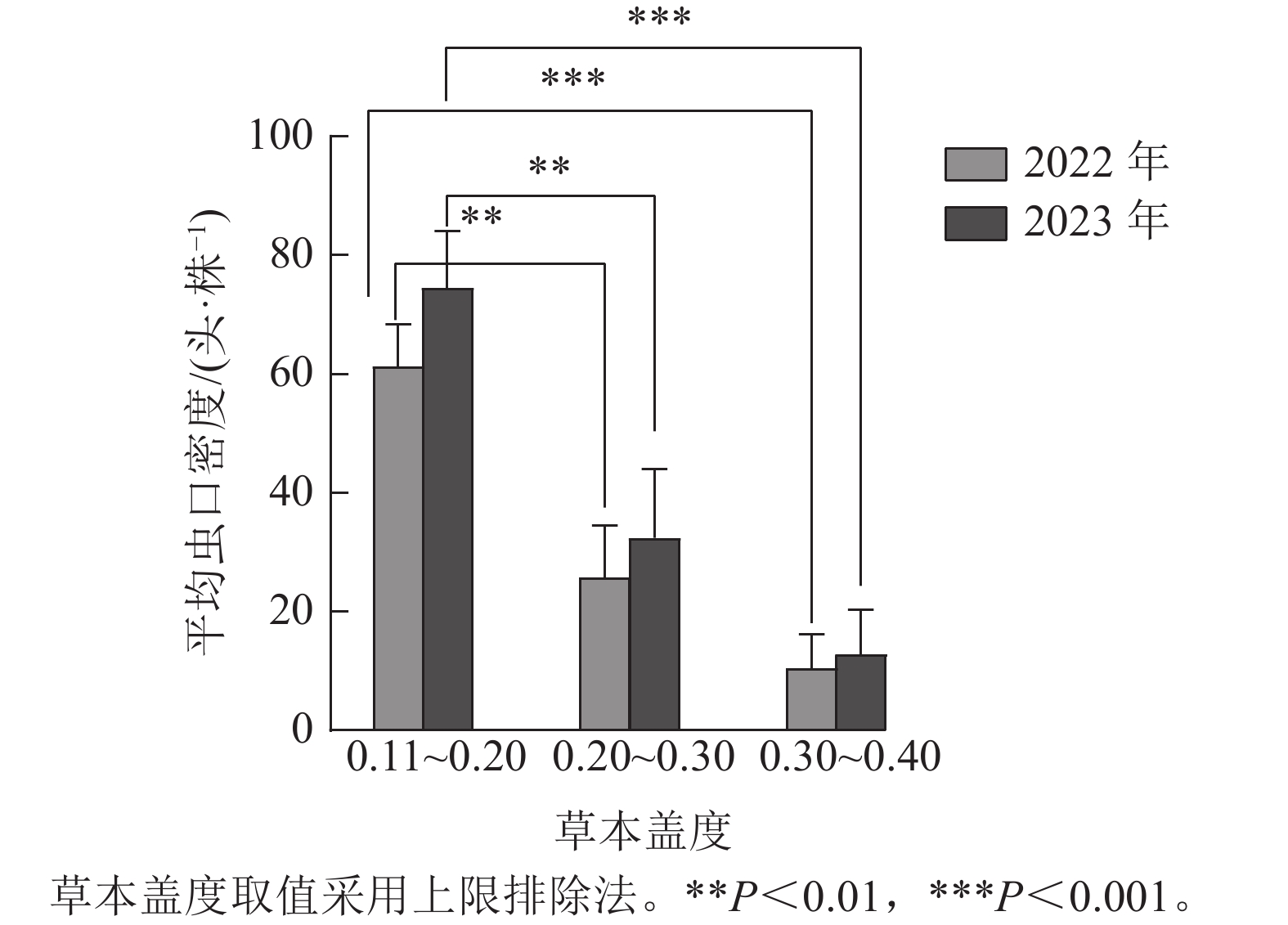
如图1所示:根据样地林下草本盖度的实际调查结果,将2022与2023年调查的草本盖度数据均分为3组,第1组:0.11~0.20,第2组:0.20~0.30,第3组:0.30~0.40(上限排除法)。方差分析结果表明:2022年(F=12.635,P<0.01)与2023年(F=11.599,P<0.01)不同草本盖度对松阿扁叶蜂幼虫平均虫口密度的影响达到极显著水平。Duncan多重比较表明:不同年份林下草本盖度为0.20~0.30(第2组)和0.30~0.40(第3组)时的松阿扁叶蜂幼虫平均虫口密度差异不显著,2022年幼虫平均虫口密度分别为25.82、10.56头·株−1,2023年幼虫平均虫口密度分别为32.43、12.89头·株−1,但第2组和第3组与第1组的幼虫平均虫口密度存在极显著差异(P<0.01),第1组2022与2023年幼虫平均虫口密度分别为61.30、74.50头·株−1。
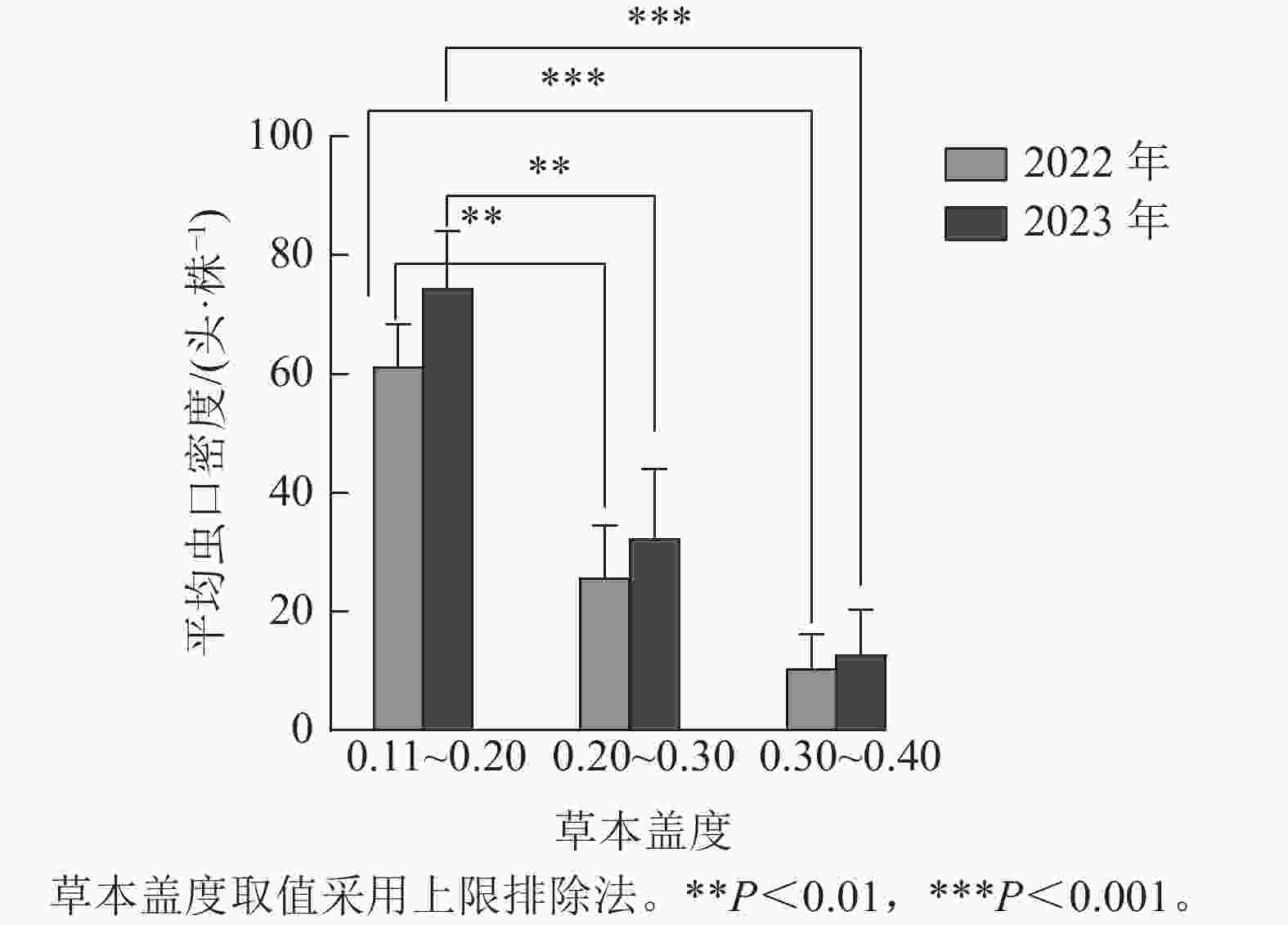
图 1 草本盖度对松阿扁叶蜂幼虫平均虫口密度的影响
Figure 1. Effect of herbaceous coverage on the average population density of A. posticalis
-
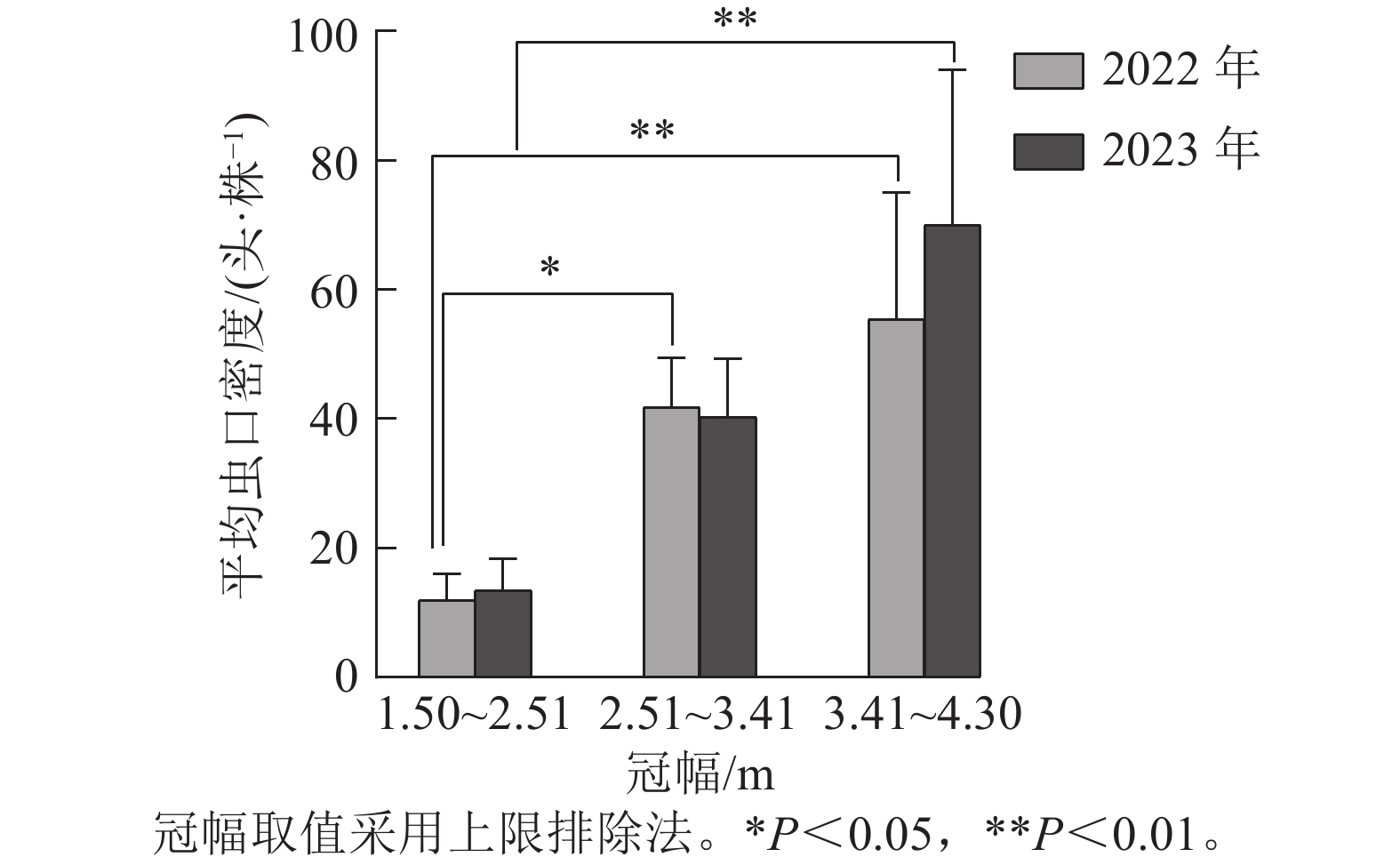
如图2所示:根据样地林木冠幅的实际调查结果,将2022与2023年调查的冠幅数据均分为3组,第1组:1.50~2.51 m,第2组:2.51~3.41 m,第3组:3.41~4.30 m(上限排除法)。方差分析结果表明:2022年不同冠幅对松阿扁叶蜂幼虫平均虫口密度的影响差异极显著(F=7.082,P<0.01),2023年影响差异显著(F=4.939,P<0.05)。经多重比较发现:2022与2023年冠幅第1组和第3组的松阿扁叶蜂幼虫平均虫口密度差异极显著(P<0.01),且2022年第3组的幼虫平均虫口密度(55.35头·株−1)显著高于第1组(11.93头·株−1),2023年第3组的幼虫平均虫口密度(40.30头·株−1)显著高于第1组(13.47头·株−1),2022年冠幅第2组幼虫平均虫口密度(41.76头·株−1)显著高于第1组(11.93头·株−1)。
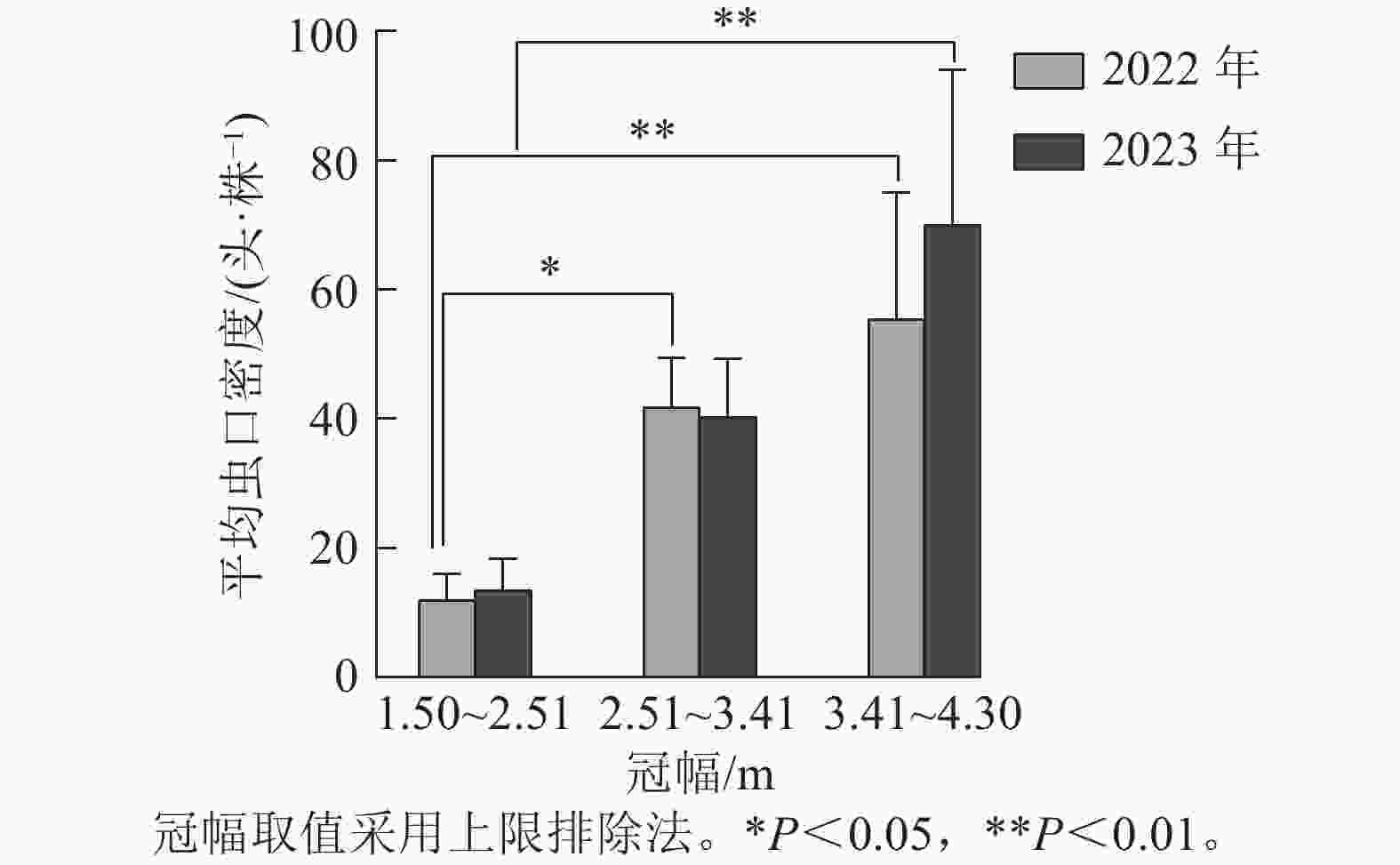
图 2 冠幅对松阿扁叶蜂幼虫平均虫口密度的影响
Figure 2. Effect of crown size on the average population density of A. posticalis
-
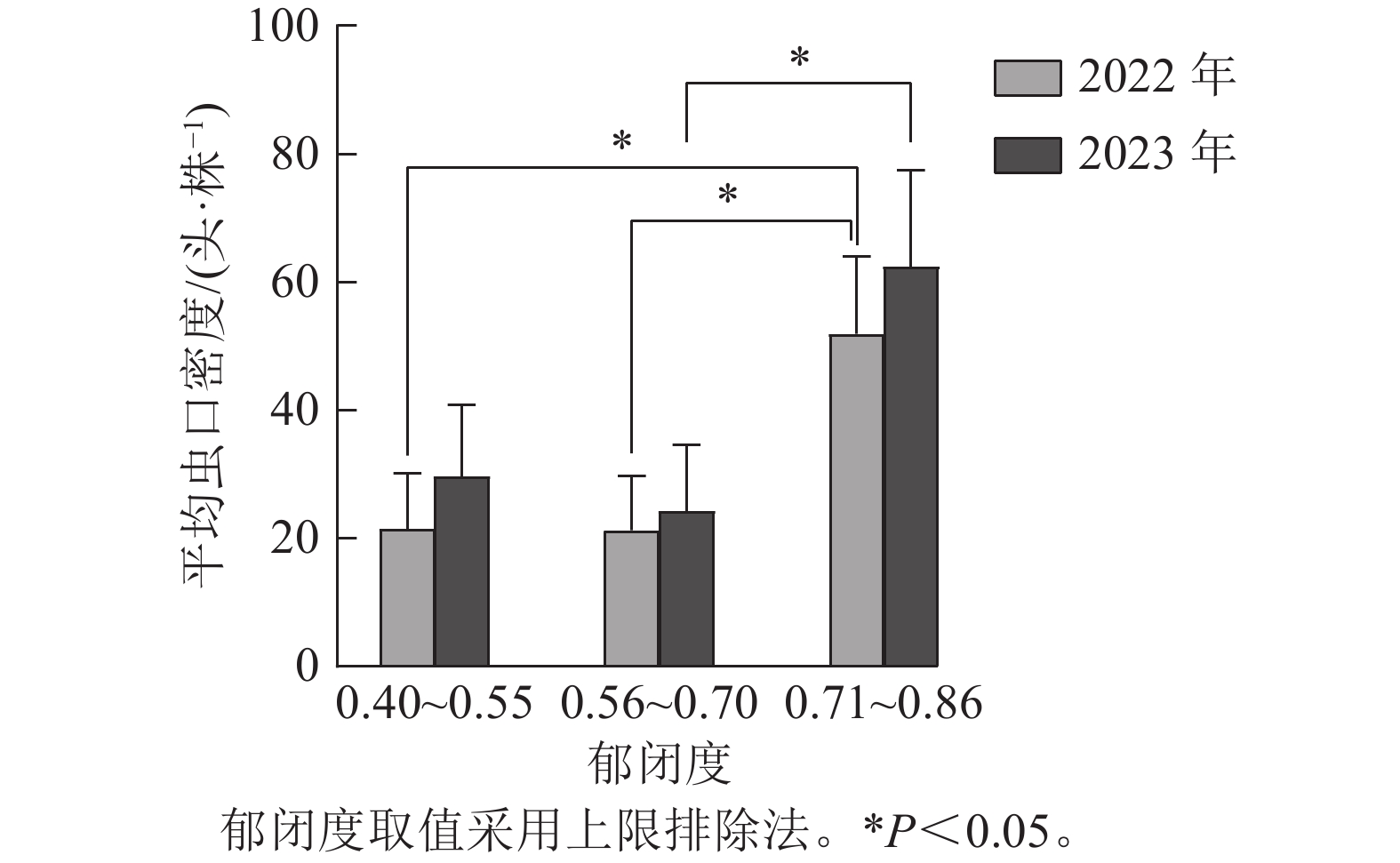
如图3所示:根据样地林分郁闭度的实际调查结果,将2022与2023年调查的林分郁闭度数据均分为3组,第1组:0.40~0.55,第2组:0.55~0.70,第3组:0.70~0.86(上限排除法)。方差分析结果表明:2 a的调查结果均显示在郁闭度小于0.70时松阿扁叶蜂幼虫平均虫口密度较小,Duncan多重比较表明:2022与2023年第2组和第3组郁闭度林分中的松阿扁叶蜂幼虫平均虫口密度存在显著差异(P<0.05),2022年幼虫平均虫口密度分别为21.32、51.92头·株−1,2023年幼虫平均虫口密度分别为24.32、62.42头·株−1。
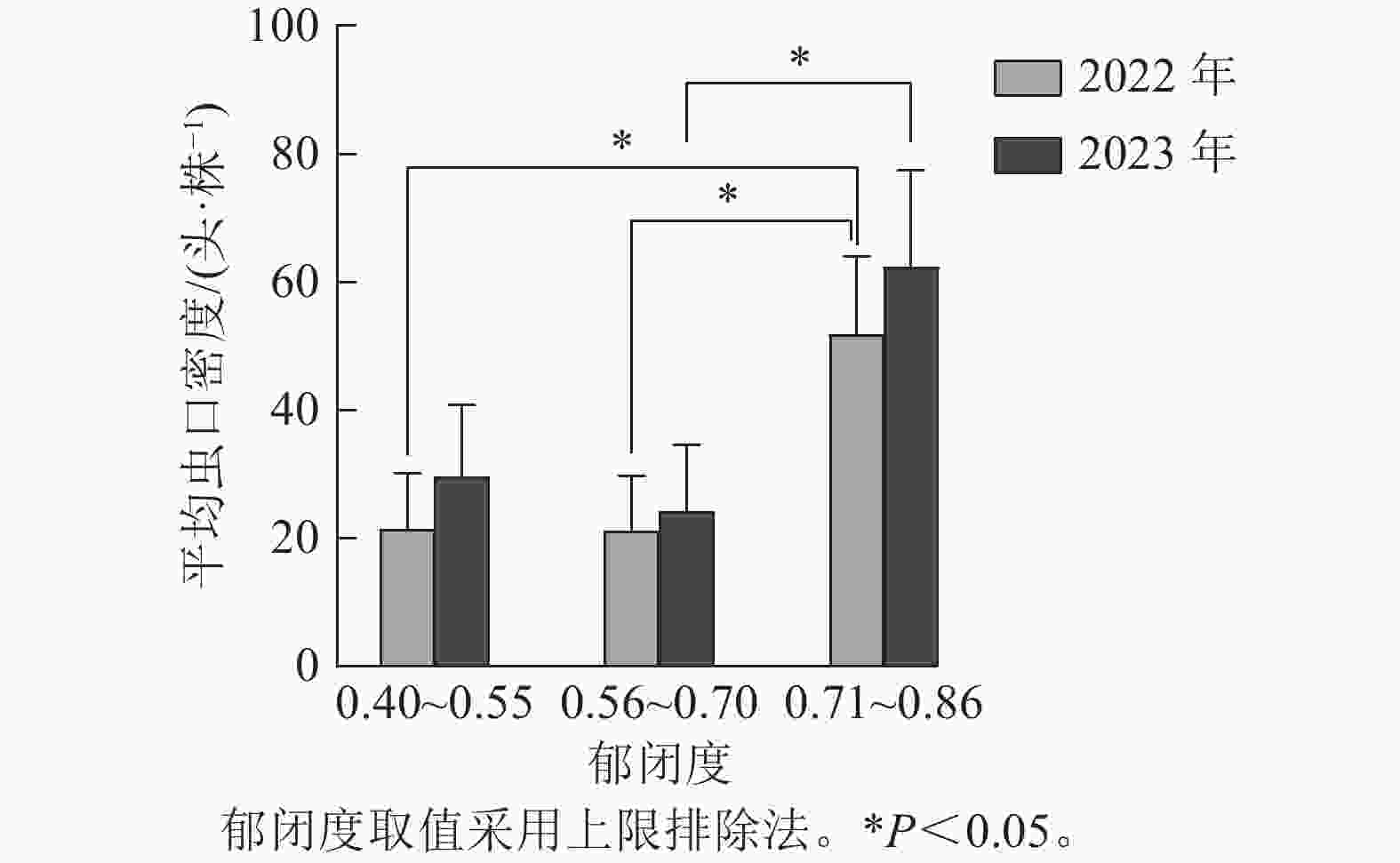
图 3 郁闭度对松阿扁叶蜂幼虫平均虫口密度的影响
Figure 3. Effect of canopy cover on the average population density of A. posticalis
-
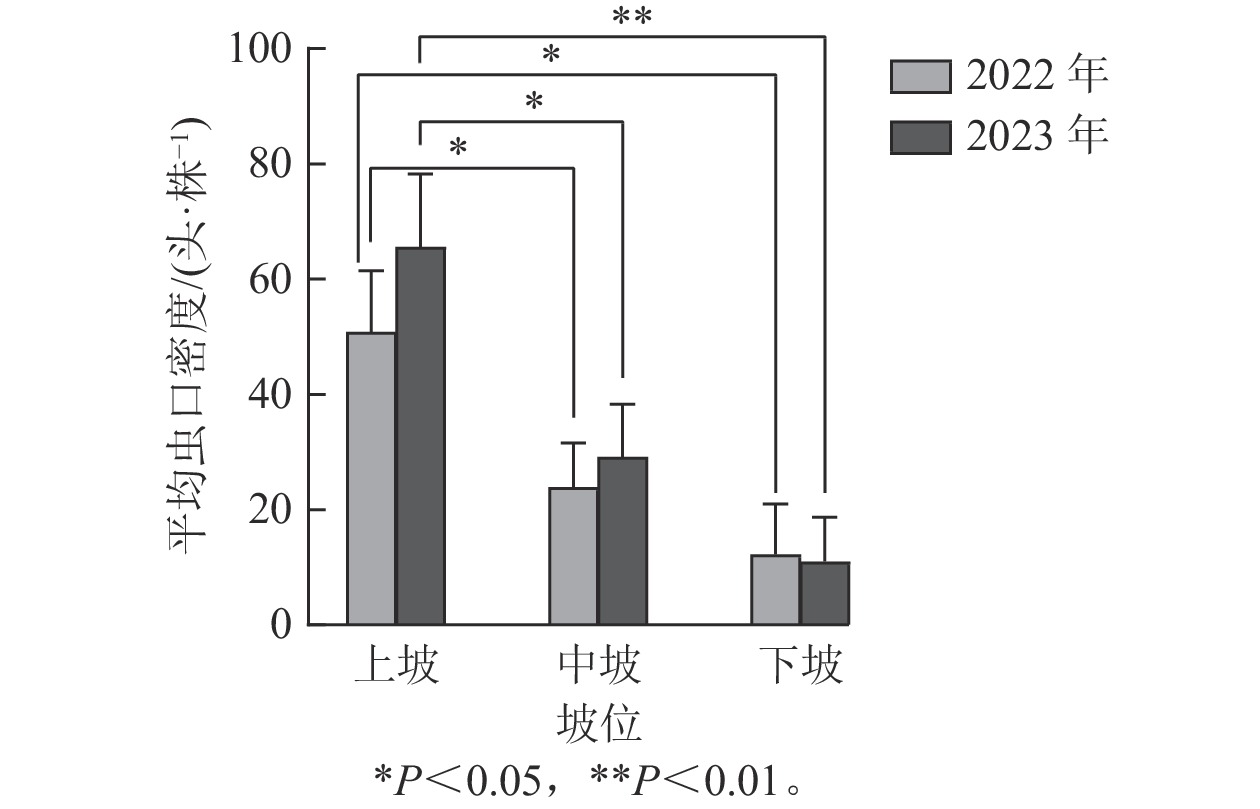
如图4所示:2022年坡位对松阿扁叶蜂幼虫平均虫口密度的影响达到显著水平(F=4.249,P<0.05),2023年坡位对松阿扁叶蜂幼虫平均虫口密度的影响达到极显著水平(F=6.270,P<0.01)。Duncan多重比较表明:2022与2023年中坡位和下坡位的松阿扁叶蜂幼虫平均虫口密度差异不显著,但两者与上坡位比较差异显著。2022与2023年上坡位的幼虫平均虫口密度平均值分别是50.90、65.62头·株−1,显著高于中坡位(23.91、29.16头·株−1)和下坡位(12.31、11.06头·株−1)。
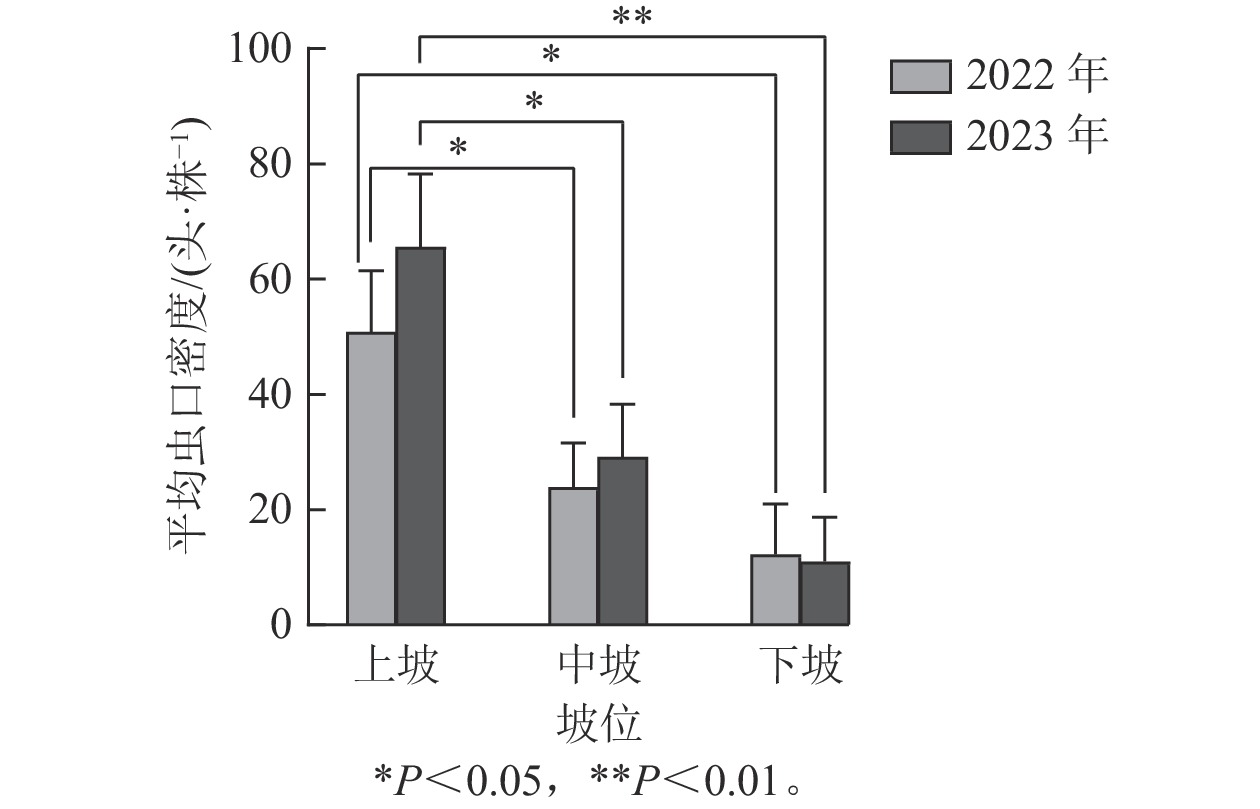
图 4 坡位对松阿扁叶蜂幼虫平均虫口密度的影响
Figure 4. Effect of slope position on the average population density of A. posticalis
-
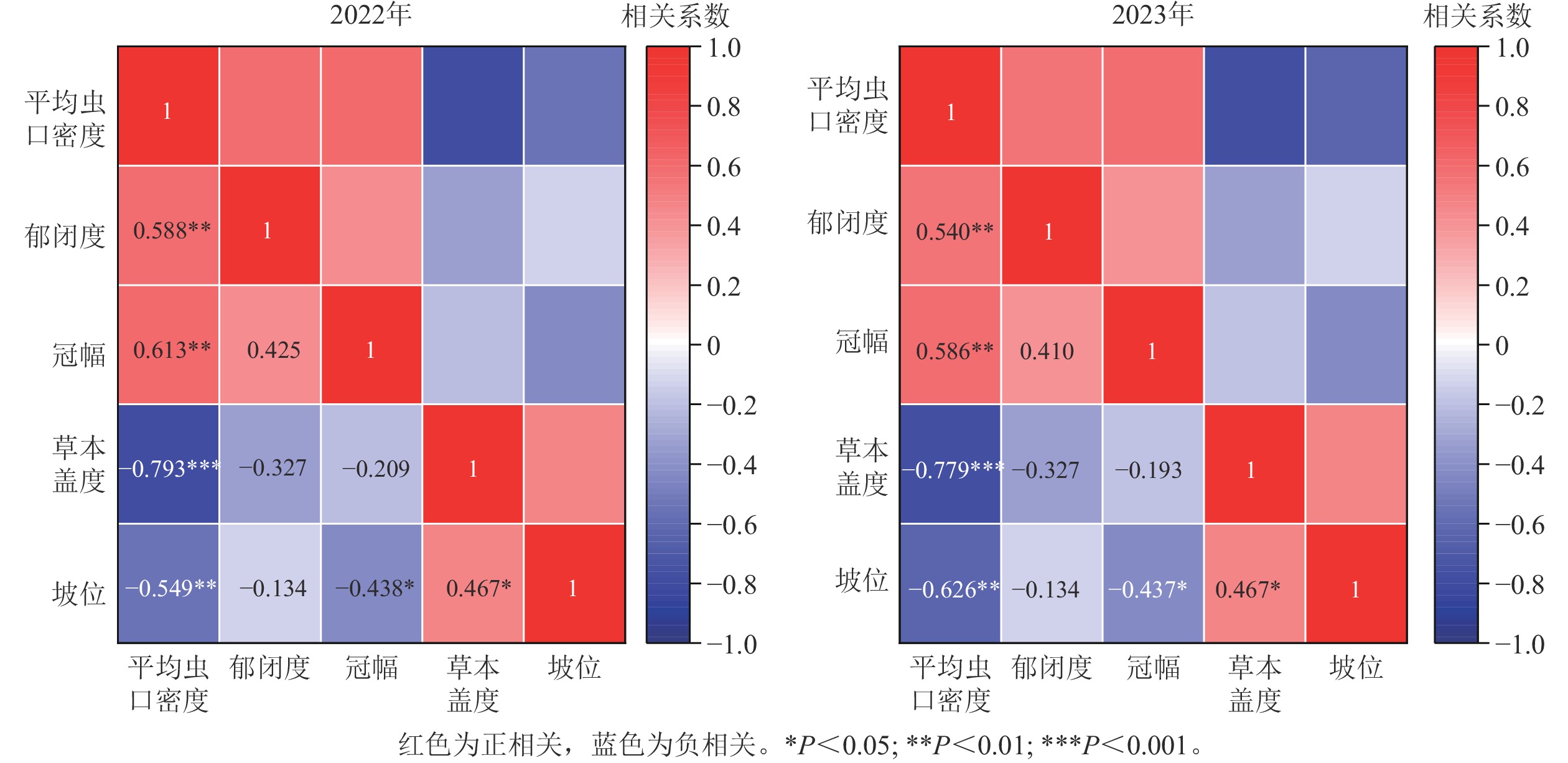
如图5所示:2022年(r=−0.793,P<0.01)、2023年(r=−0.779,P<0.01)林下草本盖度与松阿扁叶蜂幼虫平均虫口密度均呈极显著负相关,即松阿扁叶蜂幼虫平均虫口密度大小随草本盖度的增大而减小;2022年(r=0.613,P<0.01)、2023年(r=0.586,P<0.01)松阿扁叶蜂幼虫平均虫口密度随冠幅的增大呈极显著上升的趋势;2022年(r=0.588,P<0.01)、2023年(r=0.540,P<0.01)松阿扁叶蜂幼虫平均虫口密度均随林分郁闭度的增大极显著增大;2022年(r=−0.549,P<0.01)、2023年(r=−0.626,P<0.01)林木分布坡位与松阿扁叶蜂幼虫平均虫口密度均呈极显著负相关,即松阿扁叶蜂幼虫平均虫口密度在上坡位最大,中坡位次之,下坡位最小。
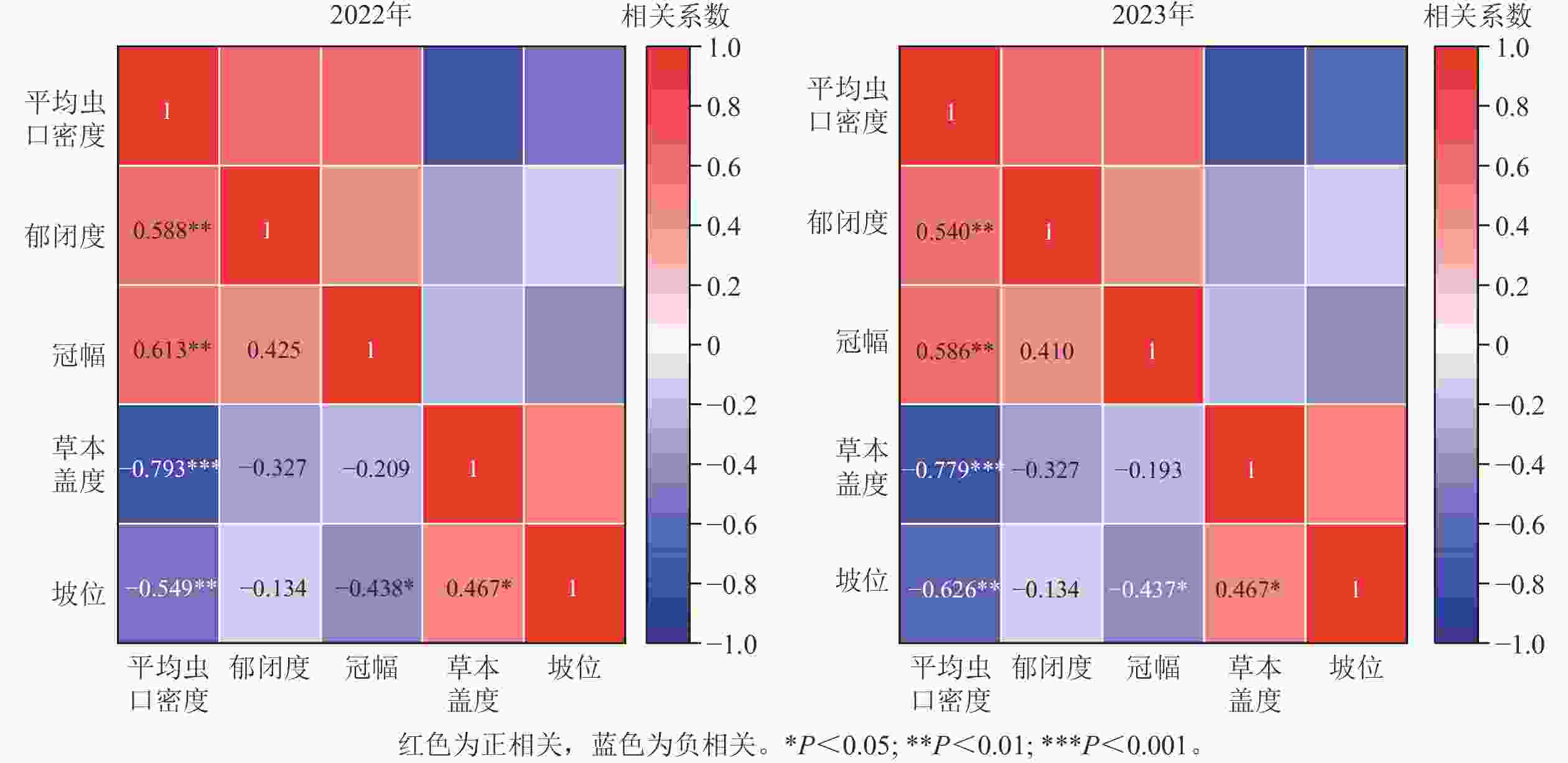
图 5 松阿扁叶蜂幼虫平均虫口密度与关键因子相关性及显著性检验
Figure 5. Correlation and significant test coefficients between key factors and A. posticalis larvae population density
-
松阿扁叶蜂是一种暴发性强,危害程度高的食叶害虫,作为森林生态系统的组成部分,与生态系统中的林分因子和立地因子共同建立了相互依存和制约的关系,对林木的危害过程就是林分因子和立地因子与松阿扁叶蜂幼虫对抗的过程[18]。因此,从森林生态系统出发,找出林分因子和立地因子与松阿扁叶蜂发生危害的关系,充分利用这种关系实现森林有害生物生态控制就显得尤为重要。
在自然条件下,影响害虫发生的影响因子一般不能独立起作用,即任意2个及多个因子间相互作用会影响害虫的发生。因此,需要将本研究调查的林分因子和立地因子通过逐步回归筛选出最显著、最能解释因变量的因子,即筛选关键影响因子[19−21]。胡瑞瑞等[22]研究发现:影响昆嵛山腮扁叶蜂Cephalcia kunyushanica发生的关键因子是昆嵛山赤松林林木的冠幅、枝下高和林分密度。唐艳龙等[23]研究发现:影响松褐天牛深沟茧蜂Iphiaulax monochamusi寄生率的关键环境因子是寄主树木的生长坡位和其他天敌寄生率,其中,坡位与松褐天牛深沟茧蜂寄生率呈显著正相关,其他天敌寄生率与松褐天牛深沟茧蜂寄生率呈显著负相关。韩大校等[24]研究发现:影响落叶松毛虫Dendrolimus superans发生的关键因子为坡向和草本盖度。本研究通过逐步回归法筛选出影响松阿扁叶蜂幼虫平均虫口密度的关键因子为郁闭度、冠幅、草本盖度和坡位。对4项因子进行多重共线诊断,方差膨胀因子远小于10,说明各关键影响因子之间互相独立[25]。为进一步明确某一自变量与因变量的相关性,所得系数能真实反映2个变量间的相关程度与性质,对本研究中的关键因子与松阿扁叶蜂幼虫平均虫口密度进行相关分析,4项因子中草本盖度、坡位均对松阿扁叶蜂幼虫平均虫口密度有极显著负影响,冠幅、郁闭度均有极显著正影响。可见,对松阿扁叶蜂幼虫平均虫口密度的影响程度从大到小依次为草本盖度、冠幅、郁闭度、坡位。
植被作为植食性昆虫的食物来源和庇护场所,其结构是影响害虫发生的因子之一[26]。林下草本植物是森林群落重要组成部分,在森林生态系统中扮演着重要角色。本研究中松阿扁叶蜂幼虫平均虫口密度和草本盖度的关系,与庞圣江等[27]发现西南桦Betula alnoides人工林林下植被盖度越大,拟木蠹蛾Arbela spp.危害率越低的结论一致。究其原因,可能与林下草本盖度的增加,提高了植物群落多样性,进而为松阿扁叶蜂幼虫的天敌提供了良好的生活栖息环境有关[28],符合天敌假说,即植物多样性的增加提高了天敌昆虫多样性和庇护所复杂性,最终提高了天敌昆虫的生物防治作用,对植食性昆虫产生上调控制作用[29];也可能是松阿扁叶蜂老熟幼虫在土层中越冬,草本的增加,其根系阻碍了越冬幼虫下潜入土,或者下潜浅,易被天敌发现,从而减轻了林地害虫的发生[2]。韩大校等[24]发现:落叶松毛虫随着草本盖度的增加,发生概率增加,这可能是研究空间尺度或者区域地理环境不同引起的差异,如所在温度带及优势树种的不同。冠幅反映了林木长期生长过程中的竞争水平[30],冠幅的大小影响林分空间结构[31],进而导致微气候发生变化,对虫害的发生产生一定影响。本研究中松阿扁叶蜂幼虫平均虫口密度随林木冠幅的增大而增大,可能是因为冠幅大的林分树冠衔接程度更高,提供给害虫更充分的营养,扩大了其生存空间,造成更大范围的扩散[32]。郁闭度是森林生态系统中重要的林分因子,是林木拥挤程度和林冠衔接程度的重要体现,直接影响林木生长发育、林内温湿度和光照条件等,间接影响林下植物组成和分布[33],也对虫害的发生造成影响。周泓杨等[34]研究林分郁闭度对土壤动物群落的影响中,发现郁闭度大的凋落物层捕食性土壤动物个体和类群数少,在罗山也可能存在林分郁闭度大时捕食性土壤动物少的情况,故而造成松阿扁叶蜂越冬幼虫数量增多。
地形作为植被格局和资源利用的代表,可以通过重新分配土壤水分和太阳辐射影响植物的生长势,从而影响森林害虫对资源的选择及分布[35]。坡度、坡向、坡位等局部地形因子与虫害的发生存在非直接关系。坡度影响山坡上树冠之间的距离,控制水分的运动,从而影响森林害虫的传播;坡向和坡位影响日照时数和光照强度,并引起温度、水分和土壤条件的变化,最终会影响森林虫害的蔓延[36]。本研究中上坡位松阿扁叶蜂幼虫平均虫口密度最大,中坡次之,下坡最少。可能与害虫种类有关,不同害虫对生存环境的要求不同,造成不同坡位害虫分布不均匀。相关研究发现:在中坡位寄主植物占比高的混交林黄褐天幕毛虫Malacosoma neustria testacea危害严重[37];在下坡位舞毒蛾Lymantria dispar的平均虫口密度最大[38]。肖玖金等[39]研究发现:土壤动物密度随着坡位的上升而减少,从而增加了天敌昆虫数量,也可能是造成害虫数量变化的原因。
-
在宁夏罗山国家级自然保护区林区,影响松阿扁叶蜂幼虫种群数量变化的关键因子是林下草本盖度(x8)、冠幅(x6)、郁闭度(x3)和坡位(x15)。根据关键因子建立了幼虫平均虫口密度的线性预测方程y=22.832+54.377x3+14.279x6−195.339x8−6.531x15,R2=0.856,调整后R2=0.840。松阿扁叶蜂幼虫平均虫口密度与草本盖度、坡位均呈极显著负相关,与冠幅、郁闭度均呈极显著正相关,即松阿扁叶蜂幼虫在林下草本盖度低、林分郁闭度和冠幅大的上坡位易发生,因此,防治重点区域可以通过调控林下草本盖度、林分郁闭度、林木冠幅等的大小,将虫害控制在低水平。
Key factors affecting the occurrence of Acantholyda posticalis larvae in Luoshan Mountains, Ningxia
-
摘要:
目的 研究松阿扁叶蜂Acantholyda posticalis幼虫的发生与林分因子和立地因子的关系,筛选影响松阿扁叶蜂幼虫发生的关键因子。 方法 2022—2023年,在宁夏罗山国家级自然保护区松阿扁叶蜂主要发生区域设立21块样地,调查松阿扁叶蜂幼虫虫口密度和林分因子及立地因子。应用逐步回归分析法筛选出影响松阿扁叶蜂幼虫平均虫口密度的关键因子;采用方差分析和相关性分析法得出松阿扁叶蜂幼虫平均虫口密度与关键因子之间的关系。 结果 逐步回归法筛选出草本盖度、冠幅、郁闭度和坡位是影响松阿扁叶蜂幼虫发生的关键因子。根据关键因子建立了幼虫平均虫口密度的线性预测方程。筛选出的4个关键因子中,草本盖度和坡位对松阿扁叶蜂幼虫的发生有抑制作用,冠幅和郁闭度对松阿扁叶蜂幼虫的发生有促进作用。 结论 松阿扁叶蜂幼虫在林下草本盖度低、林分郁闭度和冠幅大的上坡位林地易发生。建议将这些林地作为防治重点区域,调整位于上坡位的林木草本盖度至0.3以上,冠幅至2.5 m以下,郁闭度至0.7以下,实现对宁夏罗山松阿扁叶蜂幼虫种群数量的生态控制。图5表3参39 Abstract:Objective The objective is to study the relationship between the occurrence of Acantholyda posticalis larvae and stand and site factors, and screen out the key factors affecting the occurrence of A. posticalis larvae. Method 21 sample plots were set up from 2022 to 2023 in the main occurrence areas of the larvae in Luoshan National Nature Reserve of Ningxia to investigate the larval population density of A. posticalis, stand factors and site factors. Using stepwise regression analysis, the key factors affecting the average population density of A. posticalis larvae were screened out. The relationship between mean population density and key factors was obtained by variance analysis and correlation analysis. Result By stepwise regression analysis, it was found that herb coverage, crown width, canopy density, and slope position were the key factors affecting A. posticalis larval occurrence. A linear prediction equation for the average larval population density was established based on the key factors. Among the 4 key factors selected, herb coverage and slope position inhibited the larval occurrence, while crown width and canopy density promoted the larval occurrence. Conclusion Larvae of A. posticalis are easy to occur in uphill forest areas with low herb coverage, high canopy density and crown width in the understory. It is suggested that these forest areas be taken as the key areas for prevention and control, and herb coverage be adjusted to above 0.3, crown width below 2.5 m and canopy density below 0.7, so as to realize the ecological control of the larval population of A. posticalis in this area. [Ch, 5 fig. 3 tab. 39 ref.] -
Key words:
- Acantholyda posticalis /
- average population density /
- herb coverage /
- crown width /
- canopy density /
- slope position
-
图 1 草本盖度对松阿扁叶蜂幼虫平均虫口密度的影响
Figure 1 Effect of herbaceous coverage on the average population density of A. posticalis
图 2 冠幅对松阿扁叶蜂幼虫平均虫口密度的影响
Figure 2 Effect of crown size on the average population density of A. posticalis
图 3 郁闭度对松阿扁叶蜂幼虫平均虫口密度的影响
Figure 3 Effect of canopy cover on the average population density of A. posticalis
图 4 坡位对松阿扁叶蜂幼虫平均虫口密度的影响
Figure 4 Effect of slope position on the average population density of A. posticalis
图 5 松阿扁叶蜂幼虫平均虫口密度与关键因子相关性及显著性检验
Figure 5 Correlation and significant test coefficients between key factors and A. posticalis larvae population density
表 1 调查样地基本信息
Table 1. Basic information of the sample site
样地号 海拔/m 坡度/(°) 坡向 坡位 优势树种 林分类型 样地号 海拔/m 坡度/(°) 坡向 坡位 优势树种 林分类型 1 2286.08 34.0 4 2 油松 纯林 12 2324.73 25.0 6 2 油松 纯林 2 2290.61 33.0 4 1 油松 纯林 13 2322.72 24.0 6 1 油松 纯林 3 2227.39 42.0 5 1 油松 纯林 14 2315.60 23.5 4 2 青海云杉+油松 混交林 4 2217.63 40.0 5 2 油松 纯林 15 2323.49 24.5 5 1 油松 纯林 5 2196.95 33.0 5 3 油松 纯林 16 2307.99 25.0 5 2 油松 纯林 6 2201.75 30.0 5 3 油松 纯林 17 2418.00 29.5 6 2 青海云杉+油松 混交林 7 2390.78 46.0 5 1 油松 纯林 18 2415.03 30.0 6 2 青海云杉+油松 混交林 8 2383.45 37.3 5 2 油松 纯林 19 2431.00 36.0 6 3 青海云杉 纯林 9 2345.23 36.5 5 3 油松 纯林 20 2432.89 37.0 6 3 青海云杉 纯林 10 2397.55 41.0 6 1 油松 纯林 21 2496.97 33.0 6 2 青海云杉 纯林 11 2335.56 39.0 6 1 油松 纯林  下载: 导出CSV
下载: 导出CSV
表 2 林分因子和立地因子与松阿扁叶蜂幼虫平均虫口密度的方差分析
Table 2. Analysis of variance between stand and site factors and average population density of A. posticalis larvae
误差来源 平方和 自由度 均方 F P 回归 37 135.254 15 2 475.684 27.796 <0.001 残差 2 315.750 26 89.067 总计 39 451.004 41
下载: 导出CSV
表 3 平均虫口密度与关键因子逐步回归分析
Table 3. Average insect population density and key factors were analyzed by stepwise regression
因子 系数a 系数bi F P R2 膨胀因子 郁闭度(x3) 22.832 54.377 54.847 <0.001 0.840 1.377 冠幅(x6) 14.279 1.480 草本盖度(x8) −195.339 1.483 坡位(x15) −6.531 1.610
下载: 导出CSV
-
[1] 张同心, 孙绪艮, 崔为正, 等. 松阿扁叶蜂对不同树种的选择行为[J]. 林业科学, 2006, 42(6): 66 − 70. ZHANG Tongxin, SUN Xugen, CUI Weizheng, et al. Selective behaviors of Acantholyda posticalis to different trees [J]. Scientia Silvae Sinicae, 2006, 42(6): 66 − 70. [2] 贾彦霞, 田会刚, 姜灵, 等. 松阿扁叶蜂越冬幼虫空间分布型研究[J]. 中国森林病虫, 2017, 36(3): 5 − 7, 12. JIA Yanxia, TIAN Huigang, JIANG Ling, et al. Study on spatial distribution patterns of overwintering larvae of Acantholyda posticalis [J]. Forest Pest and Disease, 2017, 36(3): 5 − 7, 12. [3] 梁中贵, 李建军, 刘建强, 等. 松阿扁叶蜂研究进展[J]. 中国植保导刊, 2007, 27(5): 14 − 17. LIANG Zhonggui, LI Jianjun, LIU Jianqiang, et al. Research progress on Acantholyda posticalis [J]. China Plant Protection, 2007, 27(5): 14 − 17. [4] 赵紫华. 从害虫“综合治理”到“生态调控”[J]. 科学通报, 2016, 61(18): 2027 − 2034. ZHAO Zihua. From “integrated pest management” to “ecologically based pest management” [J]. Chinese Science Bulletin, 2016, 61(18): 2027 − 2034. [5] 赵紫华, 马建华, 高峰, 等. 害虫种群区域性生态调控的系统策略[J]. 中国生物防治学报, 2021, 37(5): 855 − 862. ZHAO Zihua, MA Jianhua, GAO Feng, et al. Systematic strategy of ecologically based pest management of insect pest population at landscape scales [J]. Chinese Journal of Biological Control, 2021, 37(5): 855 − 862. [6] SÁNCHEZ-MARTINEZ G, WAGNER M R. Bark beetle community structure under four Ponderosa pine forest stand conditions in northern Arizona [J]. Forest Ecology and Management, 2002, 170(1/3): 145 − 160. [7] GRAY D R. The relationship between climate and outbreak characteristics of the spruce budworm in eastern Canada [J]. Climatic Change, 2008, 89: 447 − 449. [8] JACTEL H, BROCKEROFF E G. Tree diversity reduces herbivory by forest insects [J]. Ecology Letters, 2007, 10(9): 835 − 848. [9] PRATHER R M, CASTILLIONI K, WELTI E A R, et al. Abiotic factors and plant biomass, not plant diversity, strongly shape grassland arthropods under drought conditions [J/OL]. Ecology, 2020, 101(6): e03033[2023-11-01]. doi: 10.1002/ecy.3033. [10] JOHN P C, LAURA J N, WILLIAM R M, et al. Influence of landscape factors and abiotic conditions on dispersal behavior and overwintering site selection by Halyomorpha halys (Hemiptera: Pentatomidae) [J]. Journal of Economic Entomology, 2020, 113(4): 2016 − 2021. [11] FORISTER M L, NOVOTNY V, PANORSKA A K, et al. The global distribution of diet breadth in insect herbivores [J]. Proceedings of the National Academy of Sciences, 2014, 112(2): 442 − 447. [12] 李丹春, 付作霖, 徐红霞, 等. 白龙江林区油松林分因子对中华松针蚧危害的影响[J]. 林业科学研究, 2021, 34(3): 180 − 186. LI Danchun, FU Zuolin, XU Hongxia, et al. Effects of stand factors of Pinus tabulaeformis stand on damage of Matsucoccus sinensis in Bailongjiang forest area [J]. Forest Research, 2021, 34(3): 180 − 186. [13] WERNER F A, HOMEIER J. Is tropical montane forest heterogeneity promoted by a resource-driven feedback cycle? Evidence from nutrient relations, herbivory and litter decomposition along a topographical gradient [J]. Functional Ecology, 2015, 29(3): 430 − 440. [14] 孔婷婷, 刘爱华, 张静文. 天山野苹果林苹果小吉丁虫虫口密度与林分因子的关系[J]. 中国植保导刊, 2019, 39(11): 42 − 46. KONG Tingting, LIU Aihua, ZHANG Jingwen. Relationship between the stand factors and population density of Agrilus mali Matsumura in wild apple trees in Tianshan Mountain [J]. China Plant Protection, 2019, 39(11): 42 − 46. [15] 邵方丽, 余新晓, 郑江坤, 等. 北京山区防护林优势树种分布与环境的关系[J]. 生态学报, 2012, 32(19): 6092 − 6099. SHAO Fangli, YU Xinxiao, ZHEN Jiangkun, et al. Relationships between dominant arbor species distribution and environmental factors of shelter forests in the Beijing mountain area [J]. Acta Ecologica Sinica, 2012, 32(19): 6092 − 6099. [16] 任德智, 葛立雯, 罗大庆. 小尺度森林郁闭度测定方法及应用[J]. 林业科技开发, 2014, 28(1): 94 − 97. REN Dezhi, GE Liwen, LUO Daqing. A method for estimating small-scale forest crown closure and its application [J]. China Forestry Science and Technology, 2014, 28(1): 94 − 97. [17] 周润惠, 唐永彬, 王敏, 等. 威远不同年龄桉树人工林林下物种多样性和土壤理化性质[J]. 应用与环境生物学报, 2021, 27(3): 742 − 748. ZHOU Runhui, TANG Yongbin, WANG Min, et al. Species diversity and soil physicochemical properties at Eucalyptus robusta plantations of different ages in Weiyuan [J]. Chinese Journal of Applied and Environmental Biology, 2021, 27(3): 742 − 748. [18] 曾凡勇, 孙志强. 森林生态系统中植食性昆虫与寄主的互作机制、假说与证据[J]. 生态学报, 2014, 34(5): 1061 − 1071. ZENG Fanyong, SUN Zhiqiang. Mechanism, hypothesis and evidence of herbivorous insect-host interactions in forest ecosystem [J]. Acta Ecologica Sinica, 2014, 34(5): 1061 − 1071. [19] LANTSCHNER M V, CORLEY J C. Spatiotemporal outbreak dynamics of bark and wood-boring insects [J/OL]. Current Opinion in Insect Science, 2023, 55: 101003 [2023-11-01]. doi: 10.1016/j.cois.2022.101003. [20] ZHAN Zhongyi, YU Linfeng, REN Lili, et al. The association between stand and landscape level factors and red turpentine beetle damage in different infestation stages [J/OL]. Forest Ecology and Management, 2023, 531: 120790 [2023-11-01]. doi: 10.1016/j.foreco.2023.120790. [21] 辛未冬, 杜一丹, 刘华煜, 等. 地表节肢动物多样性对煤矸石山不同植被恢复方式的响应及生物指示作用[J]. 生态环境学报, 2022, 31(10): 2079 − 2088. XIN Weidong, DU Yidan, LIU Huayu, et al. Responses and biological indications of ground-dwelling arthropods diversity to different vegetation restoration patterns in coal gangue [J]. Ecology and Environmental Sciences, 2022, 31(10): 2079 − 2088. [22] 胡瑞瑞, 梁军, 谢宪, 等. 昆嵛山腮扁叶蜂发生与赤松纯林林分因子的关系[J]. 生态学杂志, 2019, 38(5): 1285 − 1291. HU Ruirui, LIANG Jun, XIE Xian, et al. Relationship between Cephalcia kunyushanica occurrence and stand factors of Pinus densiflora pure forests [J]. Chinese Journal of Ecology, 2019, 38(5): 1285 − 1291. [23] 唐艳龙, 杨忠岐, 高尚坤, 等. 寄主树木、寄主和环境因子对松褐天牛深沟茧蜂寄生率的影响研究[J]. 林业科学研究, 2018, 31(1): 72 − 77. TANG Yanlong, YANG Zhongqi, GAO Shangkun, et al. A study on host tree, host and environmental factors affecting the parasitism of Iphiaulax monochamusi Yang (Hymenoptera: Braconidae), a larval parasitoid of Monochamus alternatus (Coleoptera: Cerambycidae) [J]. Forest Research, 2018, 31(1): 72 − 77. [24] 韩大校, 王千雪, 王烁, 等. 地形和森林植被因子对落叶松毛虫越冬代发生及数量的影响[J]. 林业科学研究, 2023, 36(2): 144 − 152. HAN Daxiao, WANG Qianxue, WANG Shuo, et al. Effects of topography and forest vegetation factors on occurrence and population size of overwintering Dendrolimus superans [J]. Forest Research, 2023, 36(2): 144 − 152. [25] 曾康华. 计量经济学[M]. 北京: 清华大学出版社, 2016. ZENG Kanghua. Econometrics [M]. Beijing: Tsinghua University Press, 2016. [26] 刘强, 吴志伟, 林世滔, 等. 松材线虫病发生点格局及影响因素[J]. 应用生态学报, 2022, 33(9): 2530 − 2538. LIU Qiang, WU Zhiwei, LIN Shitao, et al. Spatial point pattern analysis of pine wilt disease occurrence and its influence factors [J]. Chinese Journal of Applied Ecology, 2022, 33(9): 2530 − 2538. [27] 庞圣江, 唐诚, 张培, 等. 广西大青山西南桦人工林拟木蠹蛾为害的影响因子[J]. 东北林业大学学报, 2016, 44(11): 85 − 88. PANG Shengjiang, TANG Cheng, ZHANG Pei, et al. Attack factors of Arbela spp. in Betula alnoides plantations at mountain Daqingshan, Guangxi [J]. Journal of Northeast Forestry University, 2016, 44(11): 85 − 88. [28] 李巧, 陈又清, 周兴银, 等. 元谋干热河谷桉树林昆虫群落初步研究[J]. 浙江林学院学报, 2008, 25(4): 502 − 506. LI Qiao, CHEN Youqing, ZHOU Xingyin, et al. Insect communities in Eucalyptus plantations in Yuanmou Arid-hot Valley [J]. Journal of Zhejiang Forestry College, 2008, 25(4): 502 − 506. [29] BELLONE D, KLAPWIJK M J, BJORKMAN C. Habitat heterogeneity affects predation of European pine sawfly cocoons [J]. Ecology and Evolution, 2017, 7(24): 11011 − 11020. [30] 惠刚盈, 张连金, 胡艳波, 等. 林分拥挤度及其应用[J]. 北京林业大学学报, 2016, 38(10): 1 − 6. HUI Gangying, ZHANG Lianjin, HU Yanbo, et al. Stand crowding degree and its application [J]. Journal of Beijing Forestry University, 2016, 38(10): 1 − 6. [31] 张晔珵, 张怀清, 陈永富, 等. 基于树冠因子的林木竞争指数研究[J]. 林业科学研究, 2016, 29(1): 80 − 84. ZHANG Yecheng, ZHANG Huaiqing, CHEN Yongfu, et al. Study of tree competition index based on crown feature [J]. Forest Research, 2016, 29(1): 80 − 84. [32] O’ROURKE M E, PETERSEN M J. Extending the ‘resource concentration hypothesis’ to the landscape-scale by considering dispersal mortality and fitness costs [J]. Agriculture,Ecosystems &Environment, 2017, 249: 1 − 3. [33] 董威, 刘泰瑞, 覃志杰, 等. 不同林分密度油松天然林土壤理化性质及微生物量碳氮特征研究[J]. 生态环境学报, 2019, 28(1): 65 − 72. DONG Wei, LIU Tairui, QIN Zhijie, et al. Research on the characteristics of soil physicochemical properties and microbial biomass carbon and nitrogen in natural Pinus tabulaeformis forests with different stand densities [J]. Ecology and Environmental Sciences, 2019, 28(1): 65 − 72. [34] 周泓杨, 张丹桔, 张捷, 等. 马尾松人工林郁闭度对大型土壤动物功能群的影响[J]. 应用生态学报, 2017, 28(6): 1860 − 1868. ZHOU Hongyang, ZHANG Danju, ZHANG Jie, et al. Effects of canopy density on the functional group of soil macro fauna in Pinus massoniana plantations [J]. Chinese Journal of Applied Ecology, 2017, 28(6): 1860 − 1868. [35] 孙志强, 张星耀, 林琳, 等. 赤松纯林林分特征对昆嵛山鳃扁叶蜂发生量的影响[J]. 生态学报, 2010, 30(4): 857 − 866. ZHANG Zhiqiang, ZHANG Xingyao, LIN Lin, et al. Impact of the stand characteristics of Pinus densiflora pure forests on the infestations of Cephalcia kunyushanica in Kunyushan region [J]. Acta Ecologica Sinica, 2010, 30(4): 857 − 866. [36] 罗先轶, 张永光. 基于多源遥感数据的森林虫害监测及驱动力分析[J]. 航天返回与遥感, 2022, 43(6): 129 − 140. LUO Xianyi, ZHANG Yongguang. Monitoring forest pests and its driving factors based on multi-source remote sensing data [J]. Spacecraft Recovery &Remote Sensing, 2022, 43(6): 129 − 140. [37] 程立超, 迟德富. 立地因子和林分因子对黄褐天幕毛虫的影响[J]. 湖南农业大学学报 (自然科学版), 2016, 42(2): 177 − 181. CHENG Lichao, CHI Defu. Effect of site factors and stand factors on Malacosoma neustria testacea Motschulsky [J]. Journal of Hunan Agricultural University (Natural Sciences), 2016, 42(2): 177 − 181. [38] 程立超, 迟德富. 立地和林分因子对舞毒蛾虫口密度的影响[J]. 西北林学院学报, 2017, 32(6): 224 − 227. CHENG Lichao, CHI Defu. Effects of site and stand factors on the average population density of Lymantria dispar [J]. Journal of Northwest Forestry University, 2017, 32(6): 224 − 227. [39] 肖玖金, 林宏贵, 周鑫, 等. 不同坡位柳杉人工林夏季土壤动物群落特征[J]. 浙江农林大学学报, 2016, 33(2): 257 − 264. XIAO Jiujin, LIN Honggui, ZHOU Xin, et al. Soil fauna community structure in Cryptomeria fortunei artificial stands at different slope elevations in summer [J]. Journal of Zhejiang A&F University, 2016, 33(2): 257 − 264. -

-
链接本文:
https://zlxb.zafu.edu.cn/article/doi/10.11833/j.issn.2095-0756.20230581
 点击查看大图
点击查看大图
计量
- 文章访问数: 1397
- HTML全文浏览量: 493
- PDF下载量: 26
- 被引次数: 0
