




-
中国拥有世界总量一半以上的稻田,水稻Oryza sativa作为最主要的粮食作物,稻米质量达标是粮食安全的重要保障。在汞(Hg)污染严重的土壤上种植水稻将导致汞异常富集,危害人体健康[1]。如贵州省铜仁市由于长期汞矿开采及重金属冶炼等活动,周边农田土壤汞的平均质量分数达14.15 mg·kg−1 [2],贵州务川汞矿区种植的稻米中总汞质量分数达到51~550 ng·g−1 [3],远超国家食品安全标准(20 ng·g−1)。因此,减少水稻甲基汞的积累,是有效降低居民甲基汞暴露风险和确保粮食安全的关键。
当前土壤汞污染的修复方法主要包括土壤淋洗法[4]、低温热解法[5]等,这些方法大多受土壤要求限制,且需要消耗大量的人力、物力[6],难以广泛应用。因此,选择更高效经济的方法来降低土壤中重金属的生物有效性,减少水稻中汞的积累,是降低稻田汞污染风险的关键。研究表明:生物质炭由于比表面积大、含氧官能团丰富等特点,在施入土壤后可提高土壤pH及有机质质量分数,降低土壤重金属的有效态,从而降低土壤重金属的生物毒性,在吸附和去除土壤重金属污染物方面具有很高的潜力[7]。MAN等[8]发现生物质炭施入稻田后,降低了土壤有效态汞的质量分数,且3%添加量的生物质炭处理组中精米的总汞质量分数低于20 ng·g−1,达到了国家食品安全标准。通过改变生物质炭表面结构特征和理化性质,可以促进或抑制生物质炭对污染物吸附与降解能力,而生物质炭的粒径大小是影响其吸附特性的关键因素[9]。XIAO等[10]研究发现:生物质炭粒径减小后,生物质炭微孔面积和比表面积显著增大,对Cd2+吸附量达165.77 mg·g−1,相较于普通粒径提高了93.91%。XU等[11]研究发现:减小生物质炭粒径后,其比表面积从260~343 m2·g−1增加到300~452 m2·g−1,并发现氮气或真空条件更有利于减小生物质炭粒径、增加比表面积。
稻田中汞来源众多,生物地球化学过程复杂,生物质炭在降低污染土壤中汞生物有效性的同时,也会为土壤汞甲基化创造有利条件,使得重金属汞的毒性增加,危害人体健康[12]。因此,在稻田土壤中施入不同粒径的生物质炭需要综合考虑其对土壤中汞甲基化和去甲基化过程的影响,本研究以水稻秸秆为原料,在600 ℃厌氧条件下通过筛分法制备不同粒径生物质炭,分析其结构特征的变化,并通过盆栽试验研究不同粒径生物质炭的施用对稻田土壤和水稻作物中甲基汞质量分数的影响,以期为生物质炭修复汞污染稻田土壤提供科学依据。
-
供试生物质炭的热解原料为水稻秸秆,来自浙江省杭州市临安区於潜镇农产品加工厂。在氮气条件下,以10 ℃·min−1的速率升温至600 ℃,恒温2 h后冷却至室温。研磨过筛,获得粒径<0.5、0.5~1.0和1.0~2.0 mm的生物质炭[13],分别以RB1、RB2、RB3表示。将所有样品装入密封袋置于干燥器内备用。
-
供试土壤采自贵州万山汞矿区水稻田0~20 cm土层,风干后研磨过2 mm筛备用。试验土壤pH为8.44;有机质质量分数为53.14 g·kg−1;全氮、全硫质量分数分别为2.51和0.54 g·kg−1;总汞和甲基汞的质量分数为11.20 mg·kg−1和3.70 μg·kg−1。
-
表面超微结构分析:在扫描电子显微镜(ZEISS Sigma 300)上用2万倍的放大倍数观察不同粒径生物质炭形貌特征。表面官能团分析:将衰减全反射附件置于红外光谱仪的光路中,扫描空气背景,将生物质炭的待测表面紧贴于衰减全反射附件的红外透光晶体面上,使用傅里叶红外光谱仪(Nicolet iS20)测定生物质炭的红外光谱。元素组成分析:利用元素分析仪(Vario EL cube)测定生物质炭样品中的元素质量分数(碳、氢、氮、硫、氧)。比表面积和孔径分布:吸附质气体为氮气,采用全自动比表面积及孔隙度分析仪(ASAP 2460)测定3种粒径生物质炭比表面积的变化。
-
在浙江农林大学的温室大棚中进行水稻盆栽试验,试验于2022年6—11月进行,共设置4个处理组,分别为添加3种粒径各3%的生物质炭处理组(SRB、MRB、LRB)以及未添加生物质炭的对照组(ck),每个处理设置3个重复,共12盆。每盆称取4.20 kg土壤样品,与对应粒径的生物质炭混匀后装入盆中。参考《春季主要农作物科学施肥指导意见》和长江中下游地区水稻种植推荐的肥料施用量,在每盆中添加0.43 g·kg−1尿素、0.19 g·kg−1磷酸二氢钾以及0.21 g·kg−1氯化钾作为氮磷钾基肥,添加去离子水,形成约2 cm高的水层,淹水10 d后备用。
选择秧龄30 d以内的健康秧苗移栽,1穴1株,每盆种植4~5株。试验期间,所有盆栽在温室中随机摆放,并定期轮换,以确保均匀的光照条件。在水稻分蘖期、抽穗期、成熟期3个时期进行土壤采样,取水稻根部周围的土壤作为根际土,通过冻干机冷冻干燥后研磨过0.15 mm筛保存备用。水稻采样取根、茎、叶、籽粒4个部分,籽粒脱壳去皮得到精米,水稻样品通过冷冻干燥、球磨、过0.15 mm筛后,于−80 ℃条件下保存备用。
土壤甲基汞采用萃取-乙基结合GC-CVAFS法测定[14],总汞采用测汞仪(DMA80)测定,检出下限为0.20 μg·kg−1。土壤有效态汞采用氯化钙法提取,通过原子荧光光谱法(AFS)测定质量分数[15]。土壤可溶性有机碳使用TOC元素分析仪(Vario)测定。水稻样品中的甲基汞采用溶剂萃取-水相乙基化衍生GC-CVAFS联用法测定[16],使用手动汞形态测试系统(Brooks Rand),检出下限为0.60 pg·kg−1。
-
使用Origin 2021和Excel 2010进行数据处理和绘图,使用SPSS 26计算各指标平均值及标准差,进行单因素ANOVA方差齐性检验和LSD-T2多重比较差异显著性分析(P<0.05)。
-
结合3种粒径生物质炭的基本理化性质和扫描电镜图分析(表1和图1),不同粒径生物质炭颗粒的表面形貌差异明显,RB1、RB2生物质炭表面形貌粗糙,RB1生物质炭表现为不规则的粗糙超细颗粒,RB3较RB1、RB2表面更光滑,孔隙结构减少。3种粒径生物质炭的比表面积表现为:RB1>RB2>RB3,RB1的比表面积和孔隙度显著高于其余2种粒径生物质炭(P<0.05),说明小粒径的秸秆生物质炭含有更丰富的孔隙结构。
表 1 生物质炭理化性质
Table 1. Physical and chemical properties of three particle sizes of biochar
编号 粒径/mm pH 总汞/
(mg·kg−1)元素组成/% 氢碳比 氧碳比 比表面积/
(m2·g−1)平均孔径/nm 碳 氢 氮 氧 硫 RB1 <0.5 10.71 − 55.38 1.67 1.12 10.26 0.28 0.36 0.14 134.36 0.04 RB2 0.5~1.0 10.57 − 60.75 1.72 0.99 9.65 0.24 0.34 0.12 107.85 0.03 RB3 1.0~2.0 10.65 − 62.50 1.70 0.95 9.58 0.27 0.33 0.12 62.80 0.02 说明: −为未检出。 
图 1 生物质炭扫描电镜图
Figure 1. Scanning electron microscopy of three particle sizes of biochar
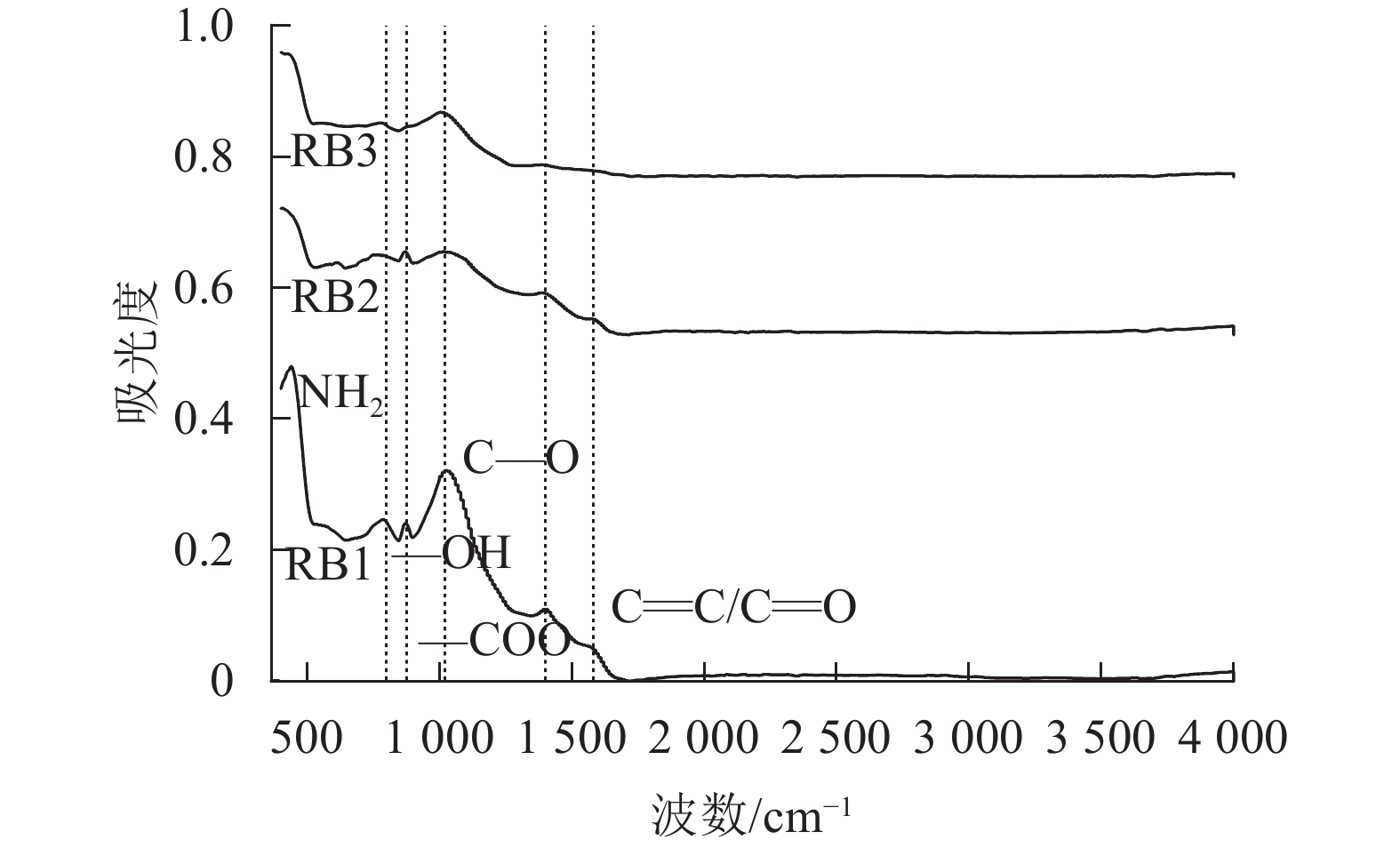
结合生物质炭的红外光谱图分析(图2),1 570和1 390 cm−1处的吸收峰分别对应C=C、C=O[17]和—COO的伸缩振动[18],824 cm−1对应羧酸中—OH的伸缩振动[19],这说明秸秆生物质炭的羧酸基团较丰富,而1 020 cm−1的振动带表明存在C—O的伸缩振动。随着粒径的减小,生物质炭C—O和—OH、—COO官能团强度增加,粒径越小的生物质炭反应活性更高,对土壤汞的络合作用更强,更有助于固定土壤中的重金属离子,降低土壤中汞的生物有效性。
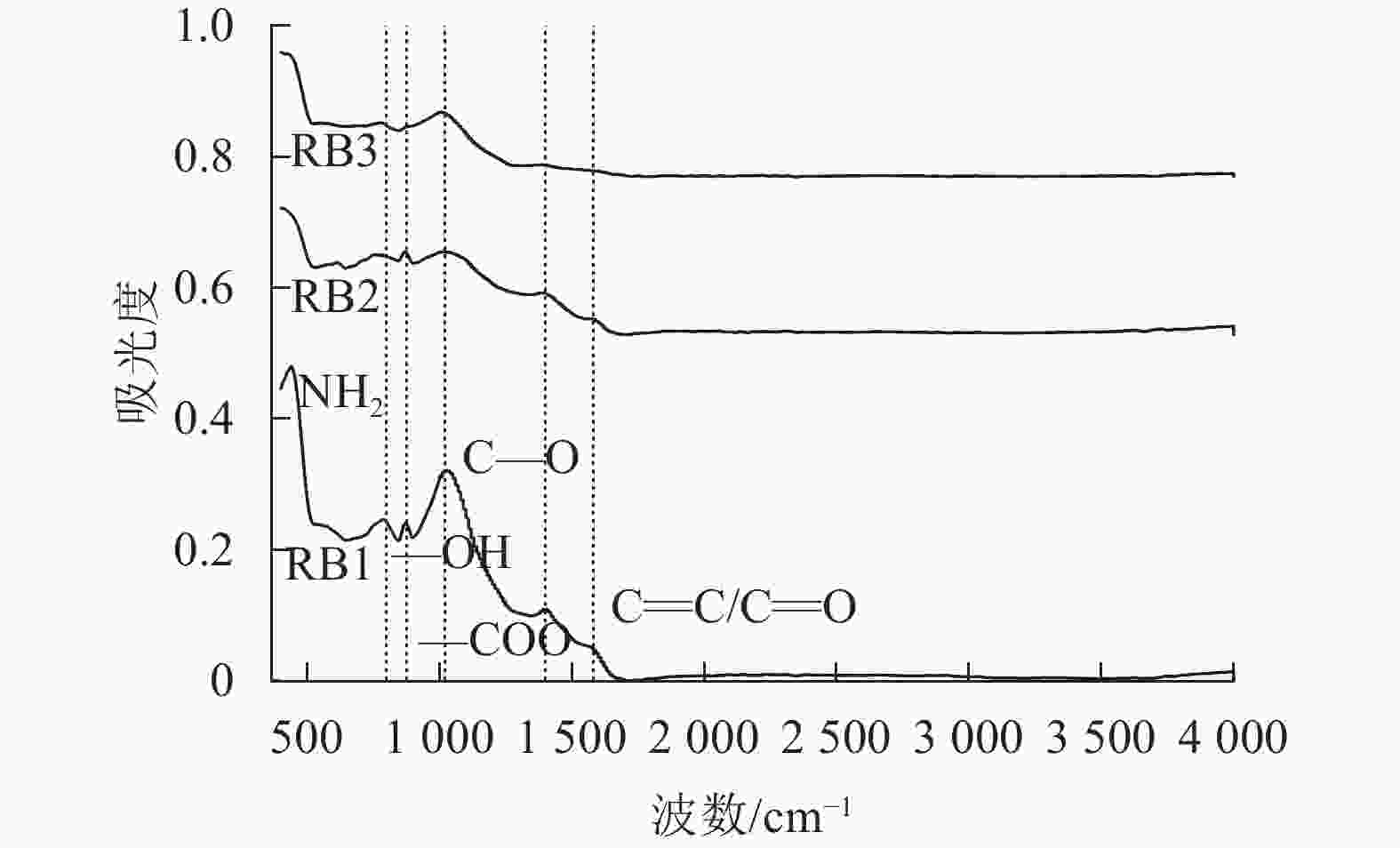
图 2 不同粒径生物质炭的红外光谱图
Figure 2. Infrared spectra of biochar with different particle sizes
-
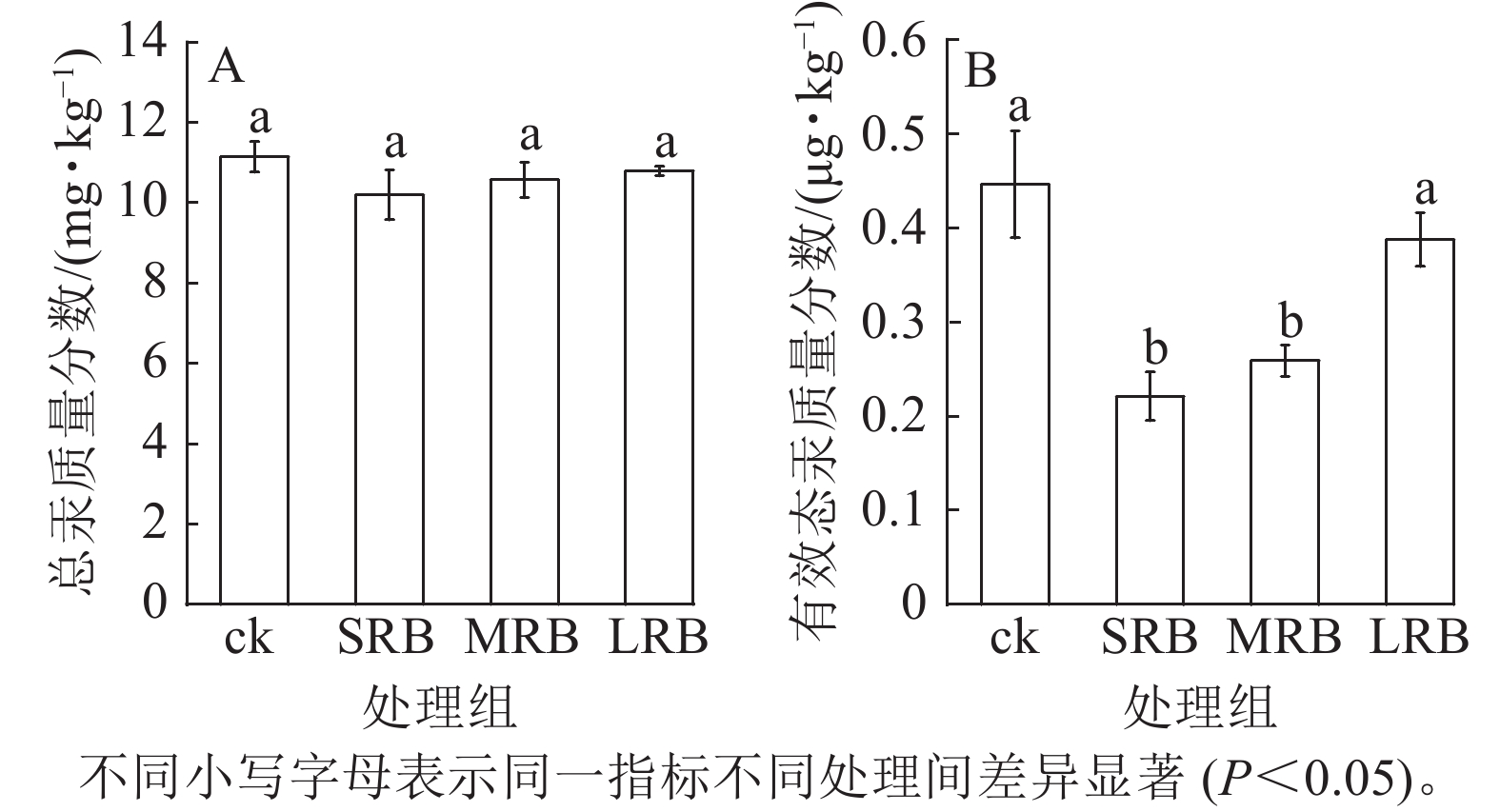
由图3A可见:不同处理组中土壤总汞质量分数没有显著差异。各处理组中土壤总汞质量分数为10.21~10.83 mg·kg−1,与ck相比降低了3.23%~8.44%,变化幅度较小。相比ck,SRB和MRB处理组有效态汞的质量分数显著降低(P<0.05,图3B),分别降低了51.15%和42.19%,土壤中汞的迁移性或者生物可利用性降低。对比不同处理组,发现随着生物质炭粒径的减小,土壤中有效态汞质量分数进一步降低,生物质炭基本理化性质以及红外光谱图表明供试生物质炭中含有丰富的含氧官能团,能够通过形成含氧吸附、络合官能团对土壤中的汞进行固定,降低土壤中汞的生物有效性。
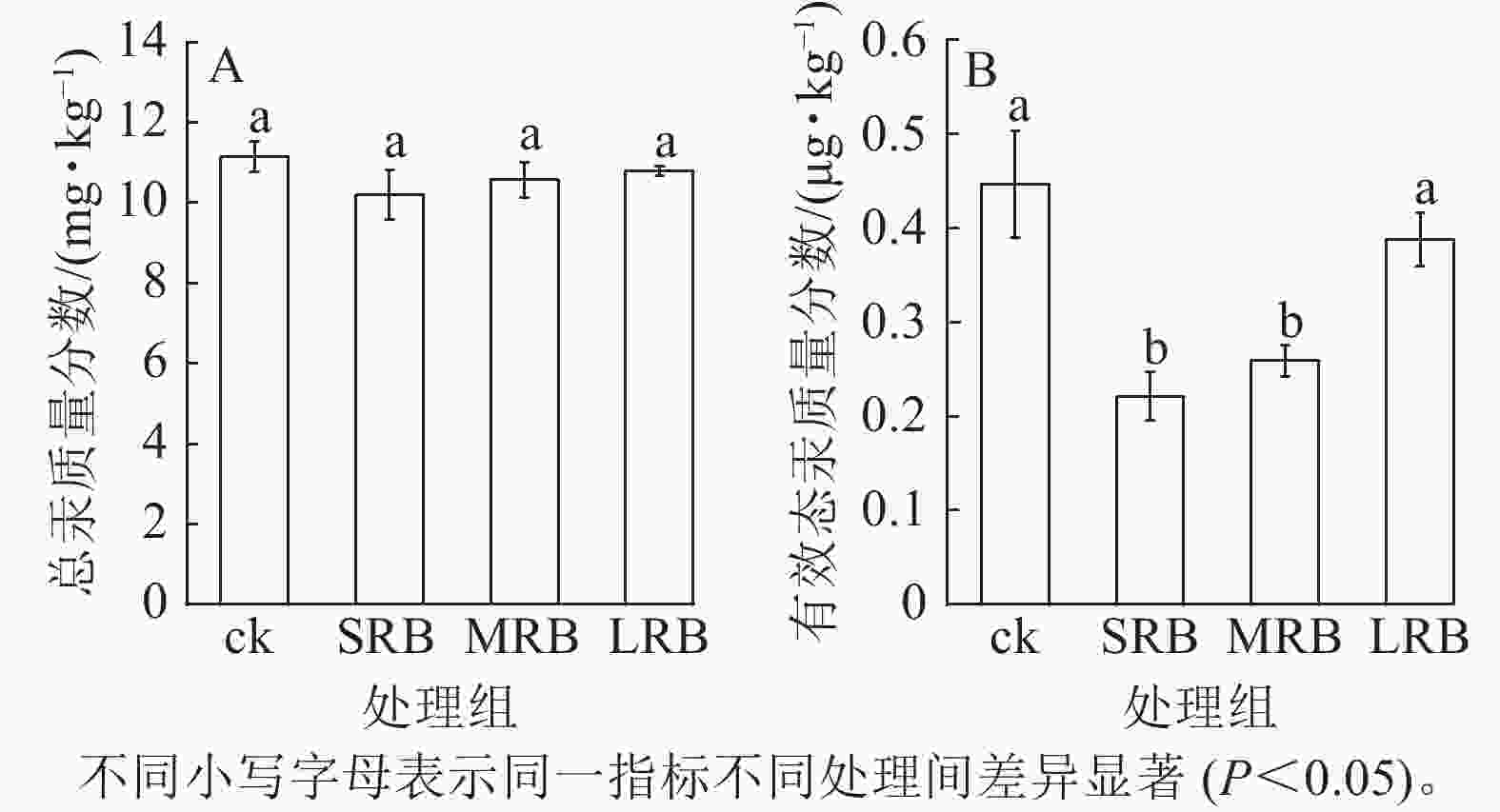
图 3 土壤总汞和有效态汞质量分数的变化
Figure 3. Changes in mass fraction of total Hg and available Hg in soil
-
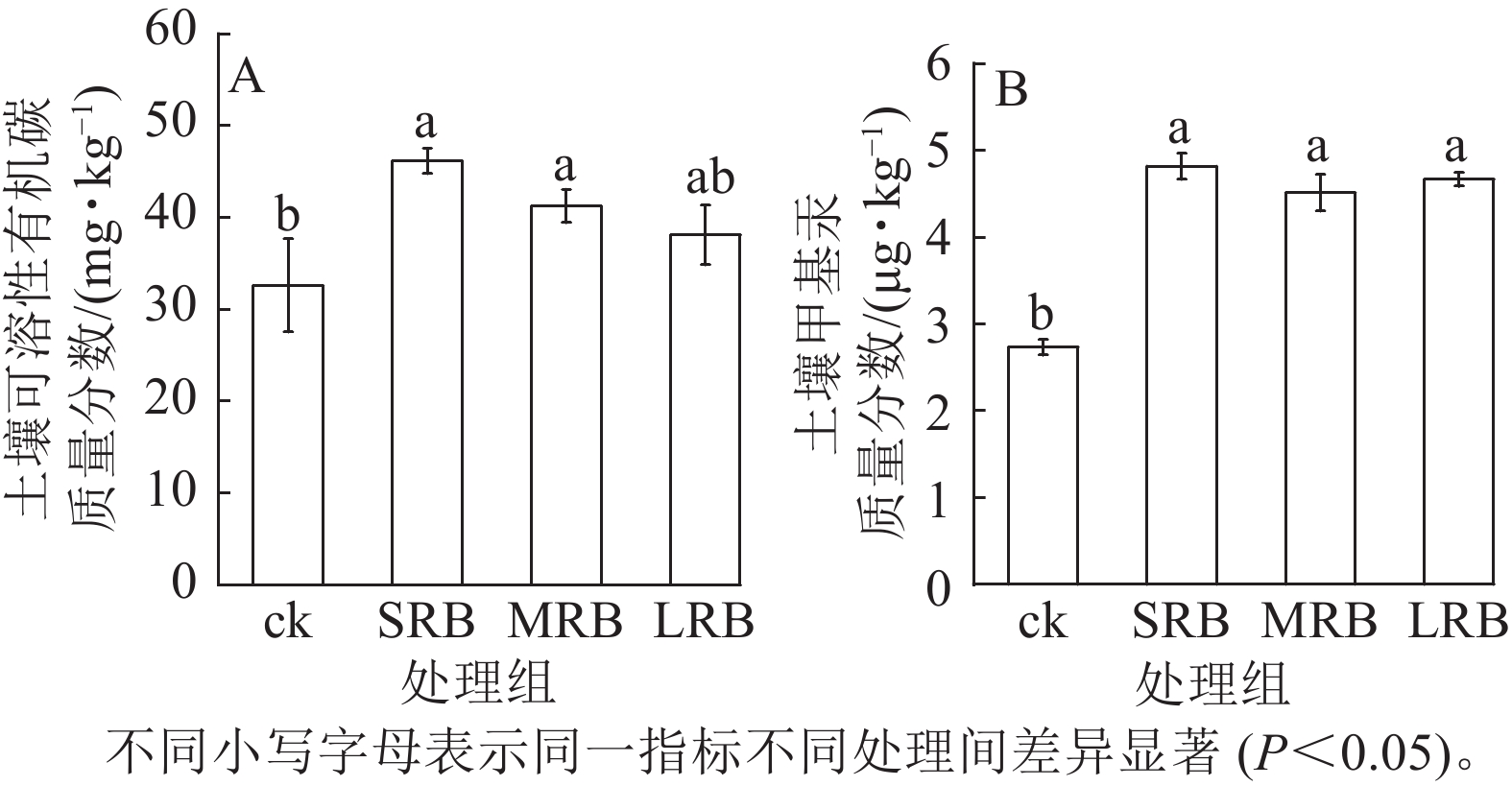
由图4A可见:生物质炭处理组土壤中可溶性有机碳质量分数显著高于ck,且可溶性有机碳质量分数随着生物质炭粒径的减小而增大,生物质炭的施入以及水稻根系分泌的大量醋酸、草酸直接向土壤中提供了大量的可溶性有机碳,土壤中可溶性有机物质量分数增多。与ck相比,SRB、MRB和LRB 处理组甲基汞质量分数差异显著(P<0.05,图4B),土壤甲基汞质量分数分别增加了76.56%、69.54%和70.69%。SRB处理组土壤甲基汞质量分数最高,为4.91 μg·kg−1,试验中施加的生物质炭含有较高的硫(0.24%~0.28%),外源硫的添加活化了土壤中的汞,促进了土壤中汞的甲基化。
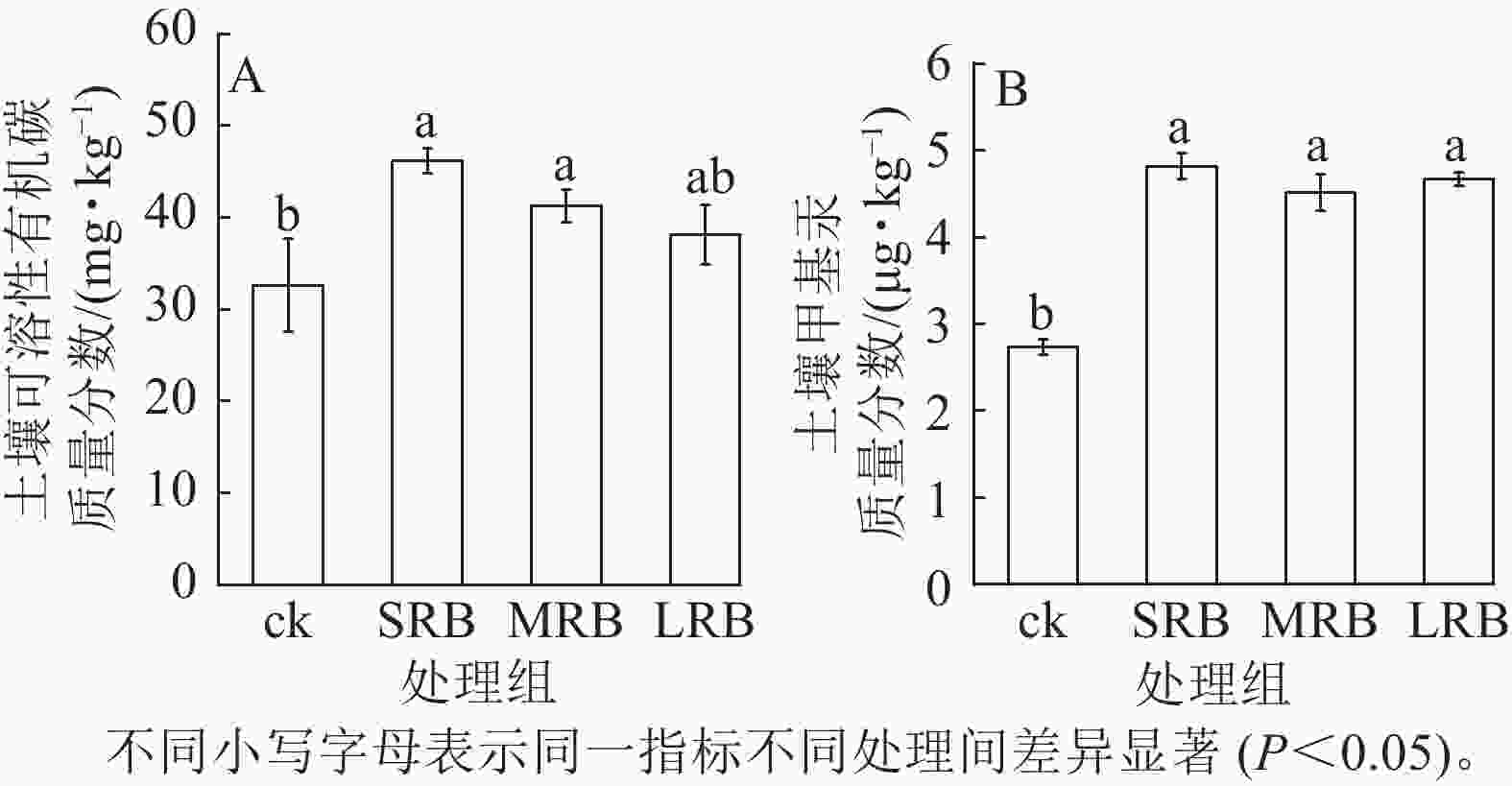
图 4 土壤可溶性有机碳和甲基汞质量分数的变化
Figure 4. Changes in mass fraction of DOC and MeHg in soil
-
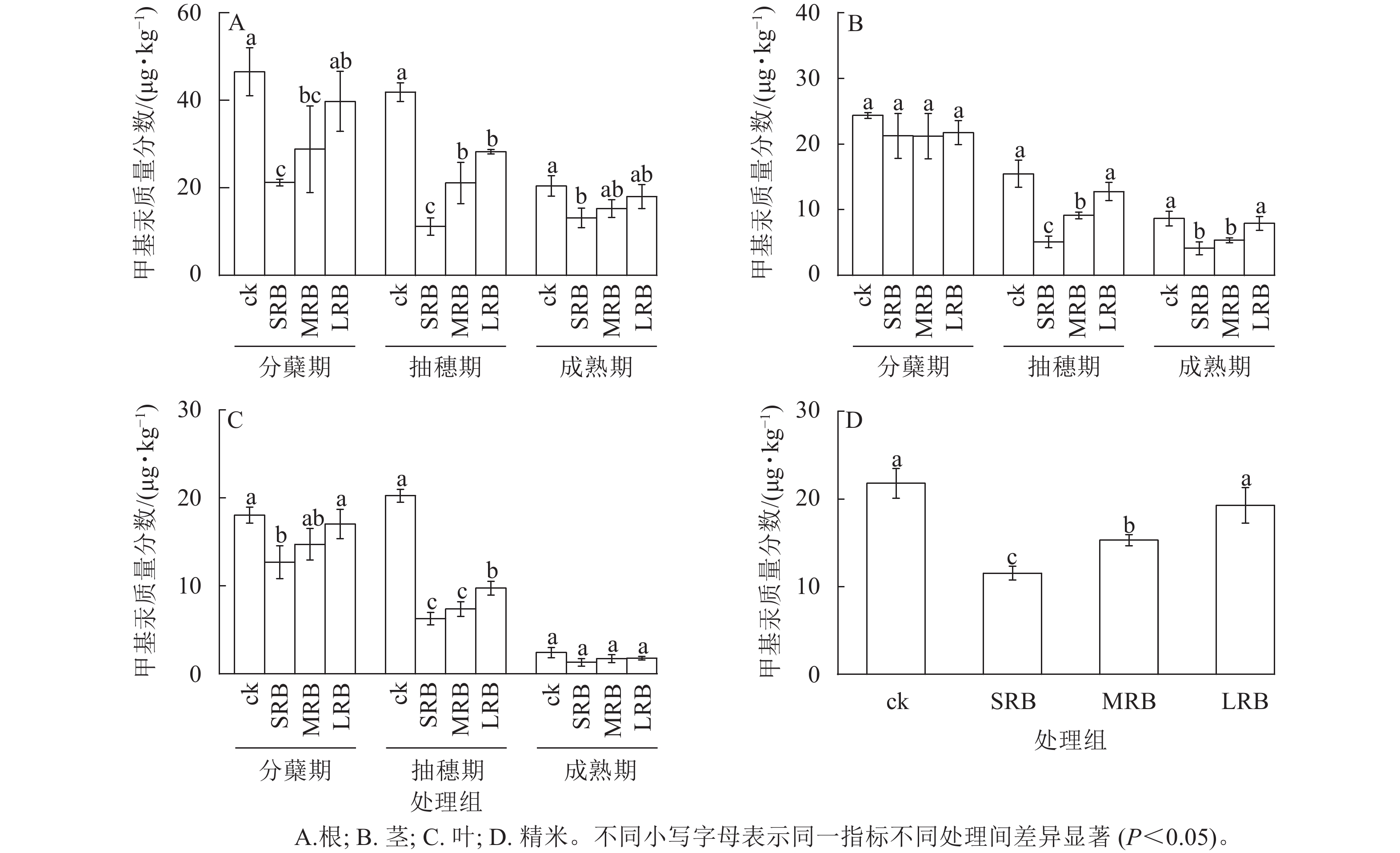
水稻生长过程中根、茎、叶甲基汞质量分数存在较大差异,成熟期根部甲基汞质量分数最高,叶甲基汞质量分数最低 (图5)。由图5A可见:水稻从分蘖期到成熟期,根部甲基汞质量分数逐渐降低,3个处理组水稻根部甲基汞质量分数在不同时期较ck均有所降低。成熟期时,与ck相比,LRB、MRB处理组根部甲基汞质量分数的变化不明显,SRB处理组根部甲基汞质量分数降低了22.10%~50.90%。
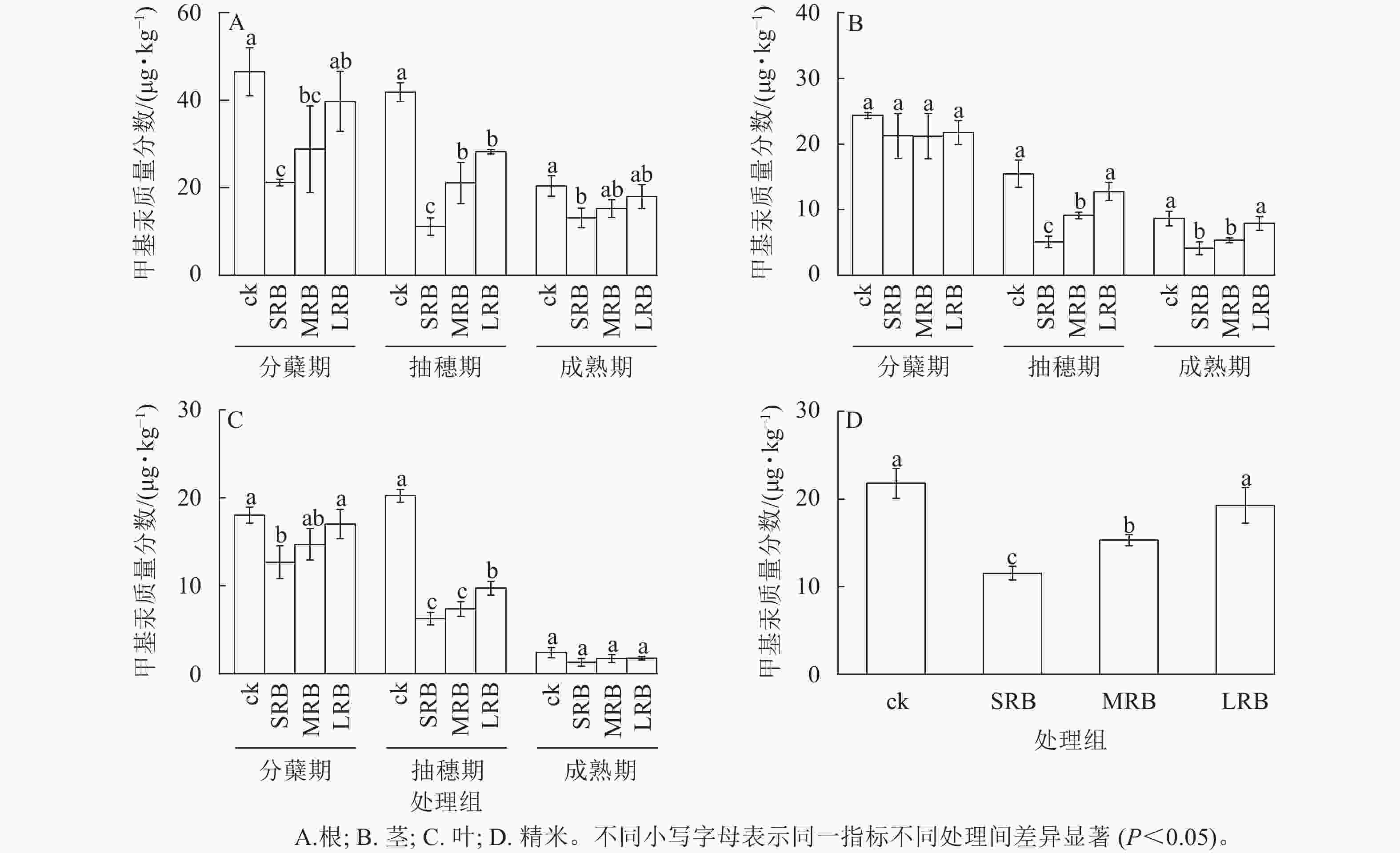
图 5 水稻各组织中甲基汞质量分数的变化
Figure 5. Changes in MeHg mass fraction in various tissues of rice
由图5B可见:整个水稻生长周期中,水稻茎部与根部的甲基汞质量分数变化规律均呈降低的趋势,与ck相比,生物质炭处理组水稻茎部甲基汞质量分数在分蘖期没有显著变化。在抽穗期,生物质炭处理组水稻茎部甲基汞质量分数与ck相比差异显著,甲基汞质量分数随着生物质炭粒径的减小而减少(P<0.05)。在成熟期,与ck相比,SRB和MRB处理组水稻茎部甲基汞质量分数分别显著降低了52.20%和38.10% (P<0.05)。
由图5C可见:水稻根、茎、叶3个部位甲基汞质量分数均呈逐渐下降的趋势,这是因为随着水稻生长发育,水稻各部位的甲基汞逐步向籽粒中转移,根、茎、叶部位中甲基汞质量分数减少,籽粒中的甲基汞明显增多。在抽穗期,ck与生物质炭处理组叶片甲基汞质量分数差异显著(P<0.05),其中SRB处理组叶片甲基汞质量分数最低。在水稻成熟期,相较于其他部位来说,叶片甲基汞质量分数最低,各处理组叶片甲基汞质量分数无明显差异。
由图5D可见:精米中甲基汞质量分数的分布情况为:ck>LRB>MRB>SRB。与ck相比,SRB、MRB、LRB处理组精米中甲基汞质量分数分别降低了47.01%、29.78%、11.52%,其中SRB处理组对水稻甲基汞富集的抑制效果最为明显。
-
从水稻不同部位甲基汞质量分数变化规律来看,根部甲基汞质量分数大于茎、叶、籽粒。在生长后期水稻根、茎、叶的甲基汞质量分数均呈下降趋势,其中叶的甲基汞质量分数最小。水稻叶片中的汞主要来源于大气,而种植水稻的温室大气汞质量分数较低,对叶片中的汞质量分数影响较小[20]。根部甲基汞质量分数最大,精米的甲基汞质量分数仅次于根部。其主要原因在于水稻根部能直接接触土壤,在接触土壤后吸收的甲基汞大量富集于根部[21]。随着水稻新陈代谢和生长发育,甲基汞从根部迁往茎部、叶部,最终在成熟期时积累在籽粒中[22]。
在水稻整个生长周期中,分蘖期水稻各部位甲基汞质量分数显著高于抽穗期和成熟期,成熟期甲基汞质量分数最低。这是由于水稻中甲基汞主要来源于种植初期根系对土壤甲基汞的吸收,该时期水稻植株矮小,生物量相对较低,所以水稻各部位甲基汞质量分数均较高。而随着水稻的生长,水稻生物量增大,在生物量稀释和生物质炭吸附固定作用下,水稻各部位甲基汞质量分数逐渐降低[23]。
3种粒径的生物质炭均能有效降低水稻中甲基汞的质量分数,与大粒径生物质炭相比,小粒径生物质炭的降低效果更好。其主要原因在于生物质炭粒径的减小能够增加其比表面积和官能团强度,扩大生物质炭的孔隙并提高生物质炭的反应活性。相关研究表明:生物质炭在施入土壤后对甲基汞的去除作用主要通过其表面官能团与甲基汞的稳定络合实现[24],粒径减小后—OH、—COO、C—O官能团强度的增加使生物质炭与汞离子络合作用增强,土壤中的甲基汞被进一步固定,甲基汞的生物有效性降低,从而抑制了水稻对甲基汞的吸收,减少了水稻各组织中甲基汞的质量分数[25−26]。另一方面,可能是小粒径生物质炭比表面积和孔隙度的增大使其能够更好地释放速效养分,增强土壤保水、保肥能力,促进水稻生长,在生物量稀释作用下有效降低水稻的甲基汞质量分数[27]。
与水稻中甲基汞质量分数变化趋势不同的是,3种粒径生物质炭的施入均提高了土壤中甲基汞的质量分数,且这种正向的促进作用随着生物质炭粒径的减小而更为明显。这可能与施加生物质炭后,水稻生长过程中有机物质分解增强了铁细菌活性有关[28]。已有研究报道:生物质炭的添加提高了土壤可溶性有机物的质量分数[29],加快了微生物介导下Fe3+的还原[30−31],提高了汞甲基化微生物的活性,且随着施入生物质炭粒径的减小,更大的比表面积和更丰富的孔隙结构显著提升了土壤微生物的附着空间,进一步促进了土壤中汞的甲基化[32]。此外,土壤中的硫与土壤重金属有着密切的联系,生物质炭中质量分数较高的硫提高了土壤中硫酸根的质量分数,促进了水稻土壤中硫酸盐还原菌的微生物甲基化过程,增大了土壤中甲基汞的质量分数[33−34]。
-
生物质炭的施入能够有效降低稻田土壤中甲基汞的生物有效性,通过含氧吸附、络合官能团为甲基汞在土壤中的固定创造有利条件,减少水稻对土壤中甲基汞的富集,且随着生物质炭粒径的减小,生物质炭对水稻组织甲基汞富集的抑制作用增强,水稻中甲基汞质量分数进一步降低。同时,施用生物炭增加了水稻根际土壤中甲基汞的质量分数,且这种正向促进作用随着粒径的减小而更为明显。
Effect of biochar with different particle sizes on the enrichment of methylmercury in rice
-
摘要:
目的 研究生物质炭粒径的改变对其理化性质以及汞污染稻田土壤中修复能力的影响,以期为减少汞污染地区水稻Oryza sativa甲基汞富集量、保障粮食安全提供科学依据。 方法 以水稻秸秆为制备生物质炭的原料,通过筛分法制备不同粒径(<0.5、0.5~1.0、1.0~2.0 mm)的水稻秸秆生物质炭,利用元素分析仪、比表面分析仪、傅里叶红外光谱仪和扫描电镜仪分析不同生物质炭的结构特征变化,并结合盆栽试验研究施用不同粒径生物质炭对水稻甲基汞富集的影响。 结果 ①随着粒径的减小,生物质炭的表面形貌变化明显,生物质炭比表面积增大,孔隙增多, C—O、—OH和—COO含氧官能团强度增加。②与对照相比,3种粒径生物质炭的施入均降低了土壤中甲基汞的生物有效性,水稻各组织中甲基汞富集量减少,其中小粒径生物质炭的降低效果最佳,添加粒径<0.5 mm的生物质炭组精米中的甲基汞质量分数降低了47.01%。③生物质炭的施入增大了水稻根际土壤甲基汞质量分数,且这种正向促进作用随着生物质炭粒径的减小而更加明显。 结论 生物质炭粒径的减小能够增强其在汞污染稻田土壤中的修复能力,减少水稻对甲基汞的积累,降低甲基汞暴露风险。图5表1参34 Abstract:Objective The impact of different particle sizes of biochar on the physicochemical properties and its remediation efficacy in mercury-contaminated rice soil was investigated, aiming to establish a scientific foundation for mitigating methylmercury accumulation in rice within mercury-polluted regions and ensure food safety. Method Rice straw was utilized as the feedstock for biochar production. The rice straw biochar with different particle sizes (< 0.5, 0.5 − 1.0, 1.0 − 2.0 mm) were obtained through sieving, and the structural characteristics of various biochar were analyzed using element analyzer, specific surface analyzer, FTIR spectrometer and scanning electron microscope. The impact of different biochar particle sizes on the accumulation of methylmercury in rice was investigated through a rice pot experiment. Result (1) With the reduction of particle size, the surface morphology of biochar had significant changes. There was an increase in the specific surface area and pore volume of biochar, as well as an enhancement in the strength of C—O, —OH and —COO oxygen-containing functional groups. (2) Compared with the control group, the application of biochar with different particle sizes reduced the bioavailability of methylmercury in soil, and the enrichment of methylmercury in rice tissues decreased. The application of biochar with the smallest particle size demonstrated the most significant effect, the addition of biochar with particles <0.5 mm reduced the mass fraction of methylmercury in polished rice grain by 47.01% compared to the control. (3) The application of biochar increased the mass fraction of methylmercury in the rhizosphere soil of rice, and this positive promoting effect became more obvious with the decrease of the particle size of biochar. Conclusion The reduction of biochar particle size can enhance the remediation capacity of biochar in mercury-contaminated rice soil, decrease the accumulation of methylmercury in rice, and mitigate the exposure risk of methylmercury. [Ch, 5 fig. 1 tab. 34 ref.] -
Key words:
- biochar /
- particle size /
- rice /
- methylmercury
-
图 1 生物质炭扫描电镜图
Figure 1 Scanning electron microscopy of three particle sizes of biochar
图 3 土壤总汞和有效态汞质量分数的变化
Figure 3 Changes in mass fraction of total Hg and available Hg in soil
图 5 水稻各组织中甲基汞质量分数的变化
Figure 5 Changes in MeHg mass fraction in various tissues of rice
表 1 生物质炭理化性质
Table 1. Physical and chemical properties of three particle sizes of biochar
编号 粒径/mm pH 总汞/
(mg·kg−1)元素组成/% 氢碳比 氧碳比 比表面积/
(m2·g−1)平均孔径/nm 碳 氢 氮 氧 硫 RB1 <0.5 10.71 − 55.38 1.67 1.12 10.26 0.28 0.36 0.14 134.36 0.04 RB2 0.5~1.0 10.57 − 60.75 1.72 0.99 9.65 0.24 0.34 0.12 107.85 0.03 RB3 1.0~2.0 10.65 − 62.50 1.70 0.95 9.58 0.27 0.33 0.12 62.80 0.02 说明: −为未检出。  下载: 导出CSV
下载: 导出CSV
-
[1] 张超, 仇广乐, 冯新斌. 汞矿山环境汞污染研究进展[J]. 生态学杂志, 2011, 30(5): 865 − 873. ZHANG Chao, QIU Guangle, FENG Xinbin. Environmental mercury pollution in mercury mining areas: a review [J]. Chinese Journal of Ecology, 2011, 30(5): 865 − 873. [2] 胡国成, 张丽娟, 齐剑英, 等. 贵州万山汞矿周边土壤重金属污染特征及风险评价[J]. 生态环境学报, 2015, 24(5): 879 − 885. HU Guocheng, ZHANG Lijuan, QI Jianying, et al. Contaminant characteristics and risk assessment of heavy metals in soils from Wanshan mercury mine area, Guizhou Province [J]. Ecology and Environmental Sciences, 2015, 24(5): 879 − 885. [3] QIU Guangle, FENG Xinbin, WANG Shaofeng, et al. Environmental contamination of mercury from Hg-mining areas in Wuchuan, northeastern Guizhou, China [J]. Environmental Pollution, 2006, 142(3): 549 − 558. [4] 许蕾. 化学淋洗法修复重金属Hg污染土壤的应用实例研究[J]. 皮革制作与环保科技, 2022, 3(11): 88 − 90. XU Lei. Case study of remediation of contaminated soil with heavy metal Hg by chemical washing technology [J]. Leather Manufacture and Environmental Technology, 2022, 3(11): 88 − 90. [5] 余志, 赵婷, 黄代宽, 等. 低温热解技术在中试条件下修复汞污染土壤的研究[J]. 环保科技, 2017, 23(4): 1 − 4. YU Zhi, ZHAO Ting, HUANG Daikuan, et al. A study on a pilot-scale remediation employing low-temperature pyrolysis technology for mercury contaminated soil [J]. Environmental Protection and Technology, 2017, 23(4): 1 − 4. [6] 马跃峰, 武晓燕, 薛向明. 汞污染土壤修复技术的发展现状与筛选流程研究[J]. 环境科学与管理, 2015, 40(12): 107 − 111. MA Yuefeng, WU Xiaoyan, XUE Xiangming. Present situation and screening strategies of remediation technology for mercury-contaminated soil [J]. Environmental Science and Management, 2015, 40(12): 107 − 111. [7] 张建云, 高才慧, 朱晖, 等. 生物质炭对土壤中重金属形态和迁移性的影响及作用机制[J]. 浙江农林大学学报, 2017, 34(3): 543 − 551. ZHANG Jianyun, GAO Caihui, ZHU Hui, et al. Mechanism and effects of biochar application on morphology and migration of heavy metals in contaminated soil [J]. Journal of Zhejiang A&F University, 2017, 34(3): 543 − 551. [8] MAN Yi, WANG Bo, WANG Jianxu, et al. Use of biochar to reduce mercury accumulation in Oryza sativa L: a trial for sustainable management of historically polluted farmlands [J/OL]. Environment International, 2021, 153: 106527[2023-12-06]. doi:10.1016/j.envint.2021.106527. [9] 迟杰, 邢海文, 张海彤, 等. 不同粒径生物炭和微塑料共存对菲吸附的影响[J]. 农业环境科学学报, 2022, 41(3): 616 − 621. CHI Jie, XING Haiwen, ZHANG Haitong, et al. Effects of the coexistence of biochar and microplastic in different particle sizes on phenanthrene sorption [J]. Journal of Agro-Environment Science, 2022, 41(3): 616 − 621. [10] XIAO Jiang, HU Rui, CHEN Gangcai. Micro-nano-engineered nitrogenous bone biochar developed with a ball-milling technique for high-efficiency removal of aquatic Cd(Ⅱ), Cu(Ⅱ) and Pb(Ⅱ) [J/OL]. Journal of Hazardous Materials, 2019, 387: 121980[2023-12-06]. doi:10.1016/j.jhazmat.2019.121980. [11] XU Xiaoyun, ZHENG Yulin, GAO Bin, et al. N-doped biochar synthesized by a facile ball-milling method for enhanced sorption of CO2 and reactive red [J]. Chemical Engineering Journal, 2019, 368: 564 − 572. [12] HU Hualing, GAO Yiman, TAN Wenbing, et al. Effects of dissolved organic matter on mercury speciation in rice rhizosphere amended with sulfur-rich biochar [J/OL]. Soil & Environmental Health, 2023, 1(2): 100022[2023-12-06]. doi: 10.1016/j.seh.2023.100022. [13] 王兴栋, 张斌, 余广炜, 等. 不同粒径污泥热解制备生物炭及其特性分析[J]. 化工学报, 2016, 67(11): 4808 − 4816. WANG Xingdong, ZHANG Bin, YU Guangwei, et al. Preparation of biochar with different particle sized sewage sludge and its characteristics [J]. CIESC Journal, 2016, 67(11): 4808 − 4816. [14] 何天容, 冯新斌, 戴前进, 等. 萃取-乙基化结合GC-CVAFS法测定沉积物及土壤中的甲基汞[J]. 地球与环境, 2004, 32(2): 83 − 86. HE Tianrong, FENG Xinbin, DAI Qianjin, et al. Determination of methylmercury in sediments and soils by GC-CVAFS after aqueous phase ethylation [J]. Earth and Environment, 2004, 32(2): 83 − 86. [15] 赵首萍, 张棋, 肖文丹, 等. 不同提取剂对土壤有效态Hg提取的效果浅析[J]. 浙江农业科学, 2020, 61(10): 2176 − 2181. ZHAO Shouping, ZHANG Qi, XIAO Wendan, et al. Comparison of extraction efficiency of bioavailable Hg from soil by different extraction agents [J]. Journal of Zhejiang Agricultural Sciences, 2020, 61(10): 2176 − 2181. [16] 仇广乐, 冯新斌, 梁琏, 等. 溶剂萃取-水相乙基化衍生GC-CVAFS联用测定苔藓样品中的甲基汞[J]. 分析测试学报, 2005, 24(1): 29 − 32. QIU Guangle, FENG Xinbin, LIANG Lian, et al. Determination of methylmercury in moss by ethylation-gas chromatography-cold vapor atomic fluorescence spectrometry with solvent extraction [J]. Journal of Instrumental Analysis, 2005, 24(1): 29 − 32. [17] WANG Zhenyu, LIU Guocheng, ZHENG Hao, et al. Investigating the mechanisms of biochar's removal of lead from solution [J]. Bioresurce Technology, 2015, 177: 308 − 317. [18] ZHOU Qiwen, LIAO Bohan, LIN Li’na, et al. Adsorption of Cu(Ⅱ) and Cd(Ⅱ) from aqueous solutions by ferromanganese binary oxide-biochar composites [J]. Science of the Total Environment, 2018, 615: 115 − 122. [19] 陈明. 生物炭纳米颗粒协同土壤中典型污染物的迁移行为[D]. 上海: 上海交通大学, 2019. CHEN Ming. Co-Transport Behaviors of Biochar Nanoparticles and Contaminants in Soils [D]. Shanghai: Shanghai Jiaotong University, 2019. [20] 陈剑, 王章玮, 张晓山, 等. 开顶式气室原位研究水稻汞富集对大气汞浓度升高的响应[J]. 环境科学, 2015, 36(8): 2997 − 3003. CHEN Jian, WANG Zhangwei, ZHANG Xiaoshan, et al. Open-top chamber for in situ research on response of mercury enrichment in rice to the rising gaseous elemental mercury in the atmosphere [J]. Environmental Science, 2015, 36(8): 2997 − 3003. [21] MENDOZA-CÓZATL D G, JOBE T O, HAUSER F, et al. Long-distance transport, vacuolar sequestration, tolerance, and transcriptional responses induced by cadmium and arsenic [J]. Current Opinion in Plant Biology, 2011, 14(5): 554 − 562. [22] DAGO A, GONZÁLEZ I, CRISTINA A, et al. Evaluation of mercury stress in plants from the Almadén mining district by analysis of phytochelatins and their Hg complexes [J]. Environmental Science &Technology, 2014, 48(11): 6256 − 6263. [23] 张志渊, 胡文俊, 卢畅, 等. 不同生育期水稻对甲基汞的蓄积[J]. 福建农业学报, 2021, 36(9): 1087 − 1091. ZHANG Zhiyuan, HU Wenjun, LU Chang, et al. Methylmercury accumulation in rice at different growth stages [J]. Fujian Journal of Agricultural Sciences, 2021, 36(9): 1087 − 1091. [24] LIU Zhen’gang, ZHANG Fushen. Removal of lead from water using biochars prepared from hydrothermal liquefaction of biomass [J]. Journal of Hazardous Materials, 2009, 167(1/3): 933 − 939. [25] NAGHDI M, TAHERAN M, BRAR S K, et al. A green method for production of nanobiochar by ball milling- optimization and characterization [J]. Journal of Cleaner Production, 2017, 164: 1394 − 1405. [26] 刘晶晶, 杨兴, 陆扣萍, 等. 生物质炭对土壤重金属形态转化及其有效性的影响[J]. 环境科学学报, 2015, 35(11): 3679 − 3687. LIU Jingjing, YANG Xing, LU Kouping, et al. Effect of bamboo and rice straw biochars on the transformation and bioavailability of heavy metals in soil [J]. Acta Scientiae Circumstantiae, 2015, 35(11): 3679 − 3687. [27] 梁传斌, 李建国, 沈枫, 等. 移栽密度和施用生物炭对水稻产量的影响[J]. 中国土壤与肥料, 2021(2): 240 − 247. LIANG Chuanbin, LI Jianguo, SHEN Feng, et al. Effect of transplanting density and biochar application on the yield of rice [J]. Soil and Fertilizer Sciences in China, 2021(2): 240 − 247. [28] MARVIN M, WINDHAMS L, AGEE J L, et al. Methylmercury production in sediment from agricultural and non-agricultural wetlands in the Yolo Bypass, California, USA [J]. Science of the Total Environment, 2014, 484: 288 − 299. [29] AULAKH M S, WASSMANN R, BUENO C, et al. Characterization of root exudates at different growth stages of ten rice (Oryza sativa L. ) cultivars [J]. Plant Biology, 2001, 3(2): 139 − 148. [30] CHEN Zheng, WANG Yuanpeng, XIA Dong, et al. Enhanced bioreduction of iron and arsenic in sediment by biochar amendment influencing microbial community composition and dissolved organic matter content and composition [J]. Journal of Hazardous Materials, 2016, 311: 20 − 29. [31] QIAO Jiangtao, LI Xiaomin, HU Min, et al. Transcriptional activity of arsenic-reducing bacteria and genes regulated by lactate and biochar during arsenic transformation in flooded paddy soil [J]. Environmental Science &Technology, 2018, 52(1): 61 − 70. [32] HUANG Limin, YU Guangwei, ZOU Fuzhen, et al. Shift of soil bacterial community and decrease of metals bioavailability after immobilization of a multi-metal contaminated acidic soil by inorganic-organic mixed amendments: a field study [J]. Applied Soil Ecology, 2018, 130: 104 − 119. [33] LI Yunyun, ZHAO Jiating, ZHONG Huan, et al. Understanding enhanced microbial MeHg production in mining-contaminated paddy soils under sulfate amendment: changes in Hg mobility or microbial methylators [J]. Environmental Science &Technology, 2019, 53: 1844 − 1852. [34] HU Hualing, XI Beidou, TAN Wenbing. Effects of sulfur-rich biochar amendment on microbial methylation of mercury in rhizosphere paddy soil and methylmercury accumulation in rice [J/OL]. Environmental Pollution, 2021, 286: 117290[2023-12-06]. doi:10.1016/j.envpol.2021.117290. -

-
链接本文:
https://zlxb.zafu.edu.cn/article/doi/10.11833/j.issn.2095-0756.20230588
 点击查看大图
点击查看大图

计量
- 文章访问数: 1303
- HTML全文浏览量: 206
- PDF下载量: 435
- 被引次数: 0
