





-
镉是一种有毒的金属元素。据《全国土壤污染状况调查公报》,镉的点位超标率达7.0%,是点位超标率最高的污染物之一。土壤中的镉可通过食物链进入人体,进而对人的身体健康造成严重的损害[1],因此亟待对镉污染土壤进行修复。
植物修复是指利用植物吸收土壤中的重金属,最终清除土壤中重金属的一种环境修复技术[2],具有经济、绿色、效果好等特点,适合大面积污染土地的修复。目前,多采用草本植物进行修复,例如鬼针草Bidens pilosa在镉质量分数为2.66 mg·kg−1的土壤生长60 d后,富集系数为4.16,对土壤中镉的去除率为4.3%~6.2%[3]。伴矿景天Sedum plumbizincicola在镉质量分数为0.55和1.85 mg·kg−1的土壤上生长至3 m时,修复效率分别为37.15%和21.82%[4]。但是草本植物存在生物量小、收割成本高、生物质后处理困难等不足,至今没有大规模的推广应用。研究表明:很多乔木可以积累重金属。张永超等[5]发现:白榆Ulmus pumila在镉质量分数为10.00 mg·kg−1的土壤中生长至7 m时,根、茎和叶的镉质量分数分别达11.2、2.42和4.74 mg·kg−1[5]。尽管乔木中重金属质量分数可能低于超积累植物,但是由于生物量大,其积累总量可能远高于超积累植物,而且重金属一旦进入乔木中,便可永久性的积累,不影响其工业用途。与超积累草本植物相比,木本植物尤其是乔木具有生物量大,不进入食物链,可以持续修复等优点,在重金属污染土壤修复中具有较大潜力[6]。然而,关于乔木吸收镉的研究大多针对器官水平,对其结构部位的研究鲜有报道。
本研究选择了在污染土壤栽植的泡桐Paulownia fortunei、楸树Catalpa bungei、悬铃木Platanus acerifolia、黑杨Populus nigra 和垂柳Salix babylonica等5种树种,分析了镉在其不同部位的分布特征,比较了不同部位的镉富集系数和镉的积累总量,旨在为合理植树造林奠定理论基础,同时也为中国镉污染土壤进行植物修复的树种选择提供依据。
-
研究区位于河南省某污染的工业用地。该地属暖温带季风气候,四季分明,气候温和,光、热、水资源丰富,年平均气温为15.5 ℃,年平均降水量为567.9 mm。拥有丰富的动植物资源及矿产资源。该地区分布有泡桐、楸树、悬铃木、黑杨和垂柳人工林纯林(于2013年前后栽植),树龄为10 a,株行距为4 m×5 m。本次采样时间为2023年7月21日。采样地泡桐、楸树、悬铃木、黑杨和垂柳土壤的镉质量分数分别为1.91、2.90、4.04、10.27和36.37 mg·kg−1。
-
随机选择污染土壤上栽植的泡桐、楸树、悬铃木、黑杨和垂柳各5株,用围尺测得胸径。采集的植物样品分为叶片、叶柄、枝皮、枝材、茎皮、茎材、根皮和根材8个部分,分别装入塑封袋。茎皮和茎材的采样高度与胸径高度一致,茎皮取样大小为4 cm×4 cm。枝皮、枝材和叶的采样高度为距地面3~5 m处的树冠。从树木东、南、西、北各个方向采集0~20 cm土壤样品,混合后装入样品袋。
-
用自来水冲洗掉附着于植物样品表面的泥土及其他污染物,然后用去离子水冲洗3遍,晾干。将样品放入烘箱105 ℃杀青30 min后,75 ℃ 下烘干至恒量。植物样品剪碎后,先用样品粉碎机初步粉碎,再用球磨机进一步磨碎,过0.149 mm筛,用来测定植物样品中镉质量分数(由于垂柳叶柄较小,难以获取,因此不作测定);土壤样品自然风干,拣出砖头、碎石、杂草等,用木棒碾碎过0.149 mm 筛,用于土壤镉质量分数测定。
土壤样品采用盐酸-硝酸-氢氟酸-高氯酸全消解法进行消解,使土壤样品中的镉全部进入试液,用石墨炉(WFX-200)测定土壤中的镉质量分数。植物样品用硝酸-过氧化氢(m硝酸∶m过氧化氢 = 10∶1)消解,经消解后植物样品用电感耦合等离子体发射光谱仪(Avio 200) 测定镉质量分数。
-
植物对镉的富集系数(F)计算公式如下:F = Ctarget/Csoil。其中:Ctarget为目标部位镉质量分数;Csoil为土壤镉质量分数。
-
植物对镉的分配计算公式如下:R = mpart/mtoatal。其中:R为相对含量;mpart为植物某部位的镉积累量; mtotal为植物中镉的积累总量。
-
采用异速生长方程估算各树种不同部位及整体生物量[7−10]。叶部、枝部、茎部和根部由于分成2个部分检测其镉质量分数,因此对两者取平均算得叶部、枝部、茎部和根部的平均镉质量分数,再估算出各树种不同部位及整体镉积累量。
-
采用SPSS 23进行数据处理及单因素方差分析,采用Origin 2022作图。
-
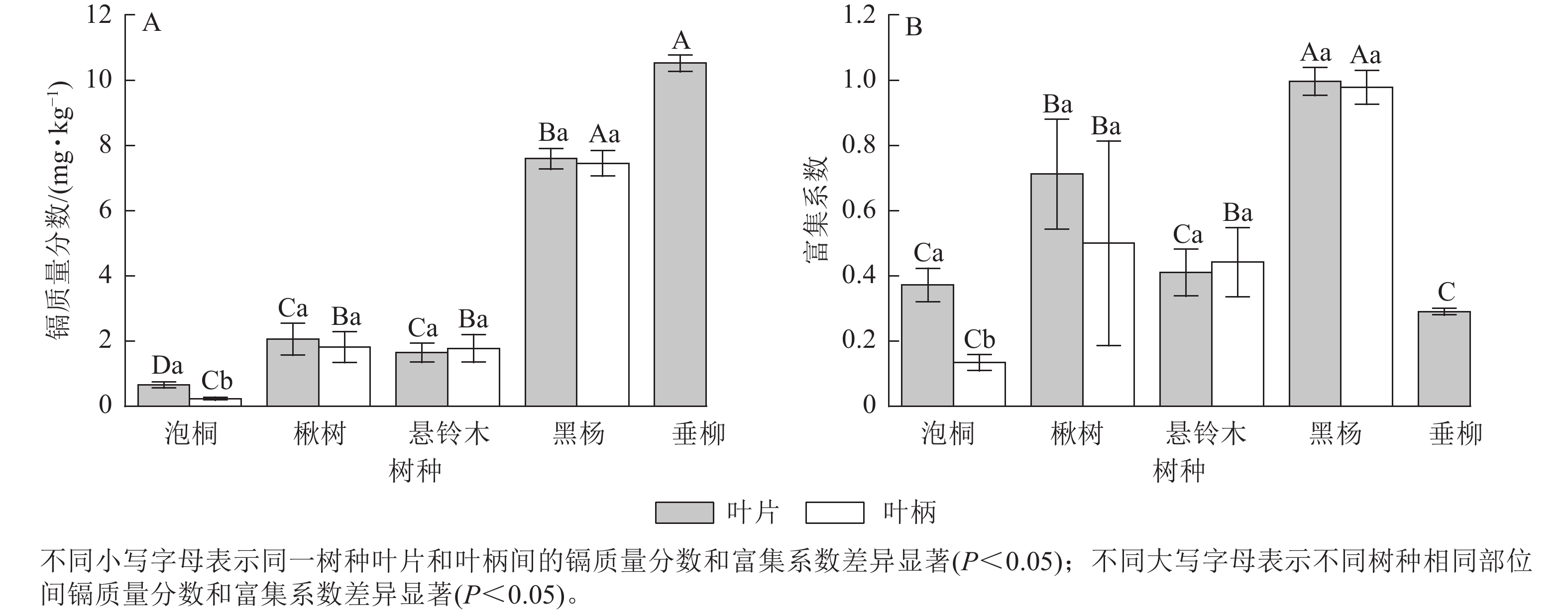
比较不同树种叶部的镉质量分数发现(图1A):垂柳叶片的镉质量分数最高,达10.52 mg·kg−1;其次是黑杨,达7.59 mg·kg−1;泡桐叶片的镉质量分数最低,为0.66 mg·kg−1。黑杨叶柄的镉最高,达7.45 mg·kg−1;其次是楸树,达2.04 mg·kg−1;泡桐叶柄的镉质量分数最低,为0.23 mg·kg−1。对比叶片和叶柄的镉质量分数发现:泡桐叶片镉质量分数显著大于叶柄(P<0.05),其他树木叶片和叶柄之间无显著差异。比较不同树木叶部的富集系数发现(图1B):黑杨叶片富集系数最高,达1.00;其次是楸树,达0.71;垂柳叶片的富集系数最小,为0.29。黑杨叶柄的富集系数最高,达0.98;其次是楸树,达0.50;泡桐叶柄的富集系数最小,为0.13。对比叶片和叶柄的富集系数发现:泡桐、楸树和黑杨叶片的富集系数均大于叶柄,悬铃木叶柄的富集系数大于叶片。泡桐叶片富集系数显著高于叶柄(P<0.05),悬铃木、黑杨的叶片与叶柄富集系数差异不显著。
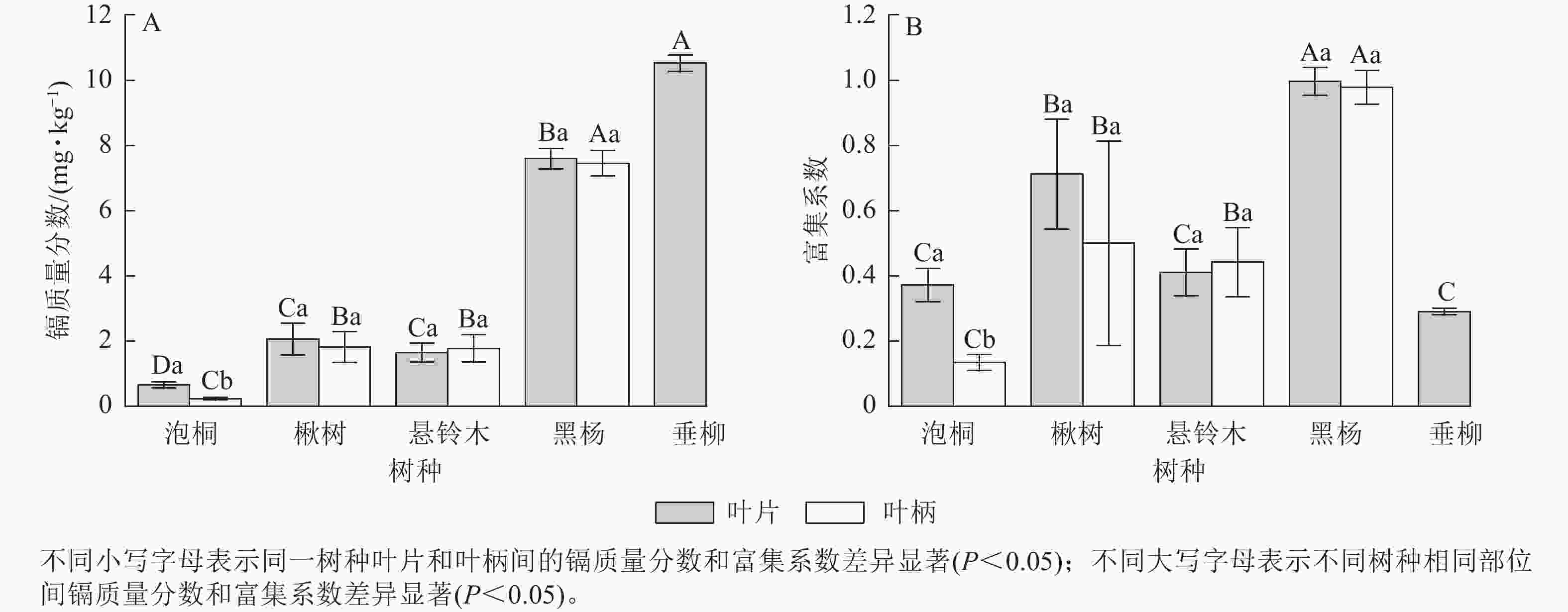
图 1 不同树种叶部的镉质量分数及富集系数
Figure 1. Cd concentration and concentration coefficients in leaves of 5 tree species
-
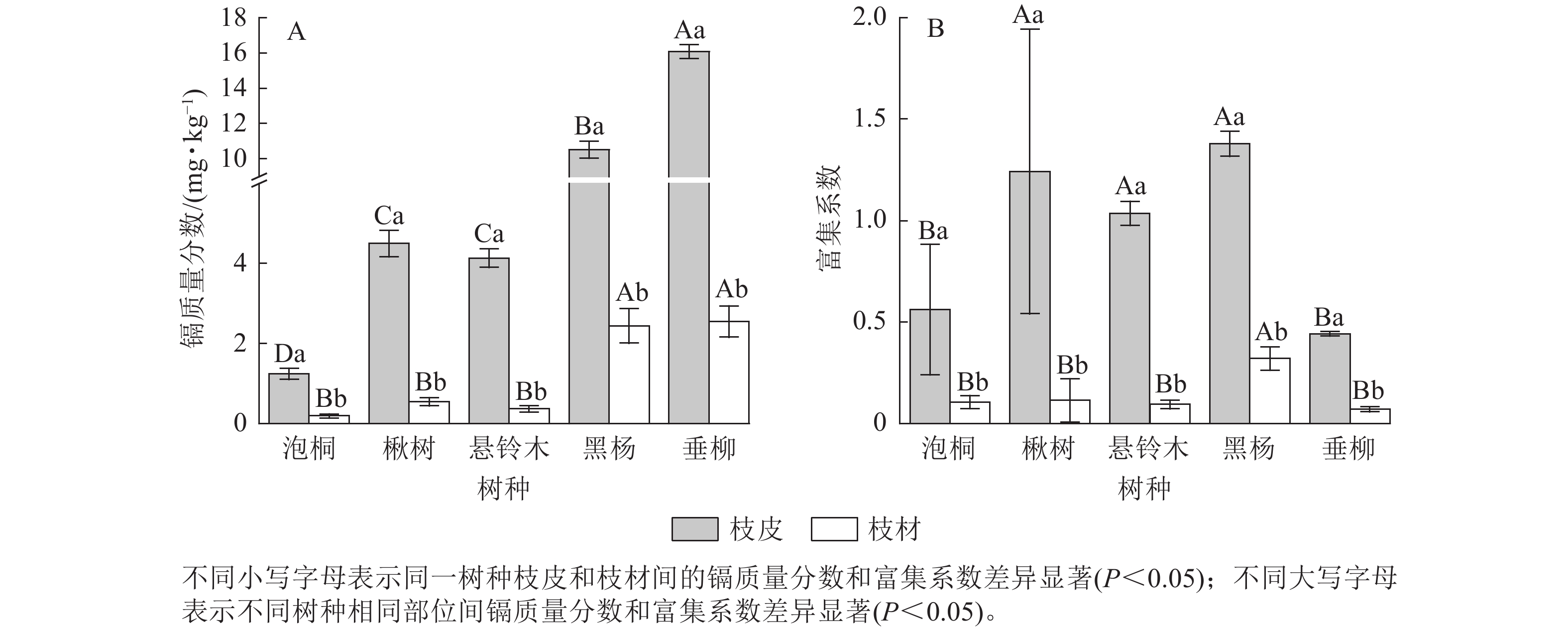
比较5种树种枝部的镉质量分数发现(图2A):垂柳枝皮的镉质量分数最高,达16.08 mg·kg−1;其次是黑杨,达10.51 mg·kg−1;泡桐枝皮的镉质量分数最低,为1.24 mg·kg−1。垂柳枝材的镉质量分数最高,达2.55 mg·kg−1;泡桐枝材的镉质量分数最低,为0.19 mg·kg−1。对比枝皮和枝材的镉质量分数发现:所有树种枝皮的镉质量分数显著高于枝材的镉质量分数(P<0.05)。比较不同树种枝部的富集系数发现(图2B):黑杨枝皮的富集系数最高,达1.38;其次是楸树,达1.24;垂柳枝皮的富集系数最低,为0.44。黑杨枝材的富集系数最高,达0.32,其次是楸树、泡桐和悬铃木,分别为0.11、0.11和0.09;垂柳枝材的富集系数最低,为0.07。对比枝皮和枝材的富集系数发现:所有树种枝皮的富集系数显著高于枝材(P<0.05)。
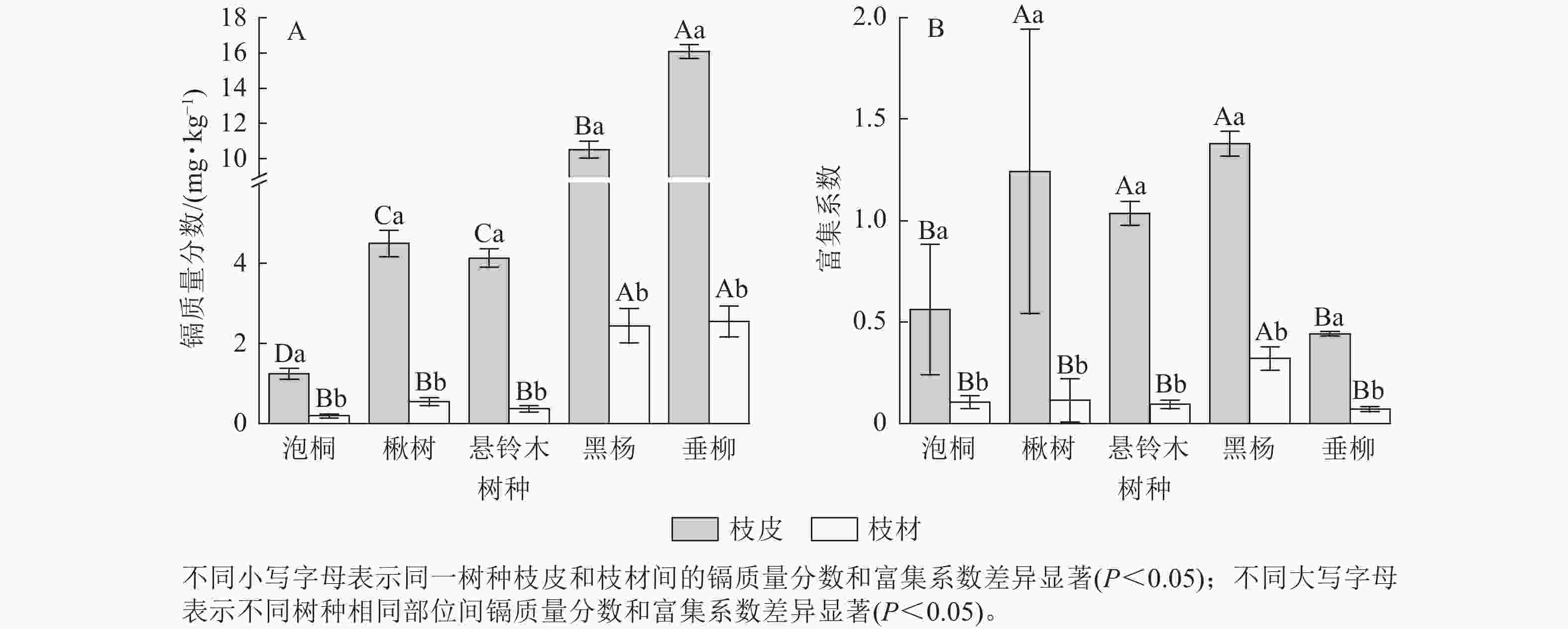
图 2 不同树种枝部的镉质量分数及富集系数
Figure 2. Cd concentration and concentration coefficients in branches of 5 tree species
-
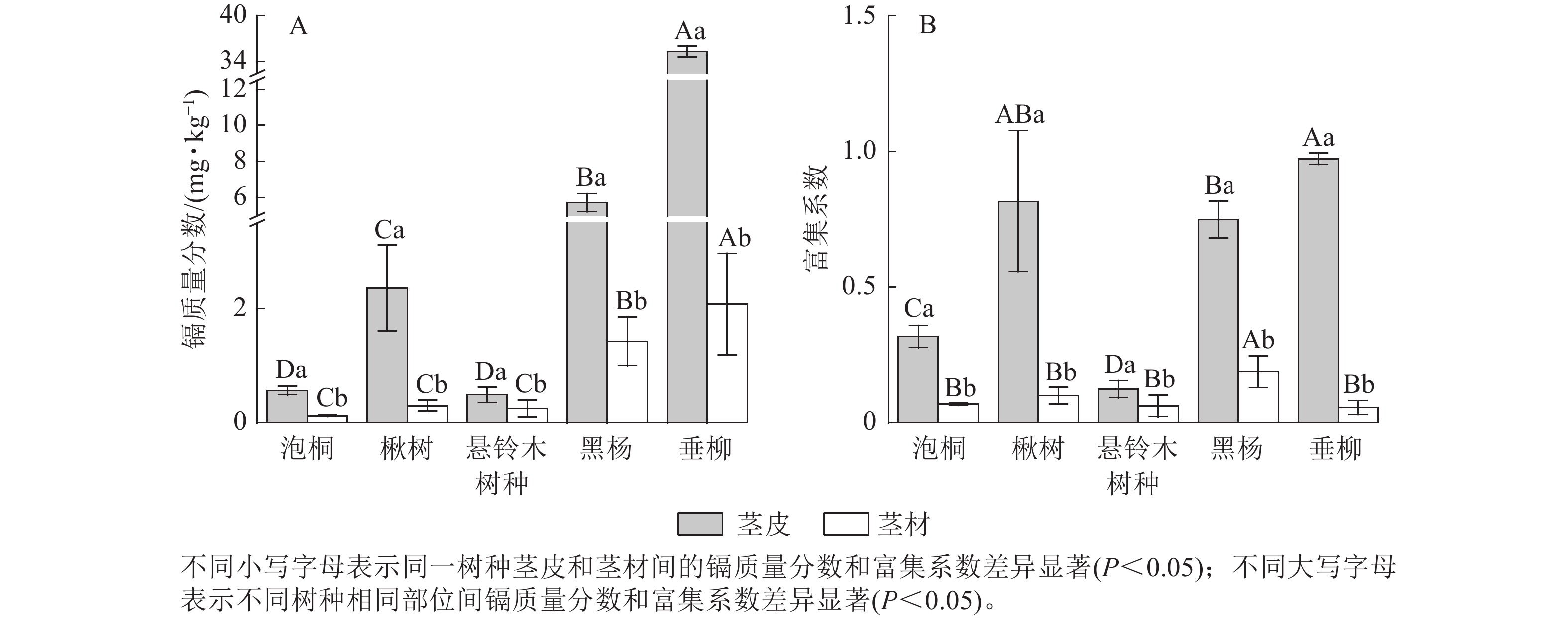
比较5种树种茎部的镉质量分数发现(图3A):垂柳茎皮的镉质量分数最高,达35.30 mg·kg−1;其次是黑杨,达5.73 mg·kg−1;泡桐茎皮的镉质量分数最低,为0.12 mg·kg−1。垂柳茎材的镉质量分数最高,达2.08 mg·kg−1;其次是黑杨,达1.43 mg·kg−1;泡桐茎材的镉质量分数最低,为0.12 mg·kg−1。对比茎皮和茎材的镉质量分数发现:楸树、黑杨和垂柳茎皮的镉质量分数显著高于茎材的镉质量分数(P<0.05),泡桐和悬铃木茎皮与茎材的镉质量分数差异不显著。比较不同树种茎部的富集系数发现(图3B):垂柳茎皮的富集系数最高,达0.97;其次是楸树,达0.82;悬铃木茎皮的富集系数最低,为0.12。黑杨茎材的富集系数最高,达0.19,其次是楸树、泡桐和悬铃木,分别达0.10、0.07和0.06;垂柳茎材的富集系数最低,为0.05。对比茎皮与茎材的富集系数发现:除悬铃木外,所有树种茎皮的富集系数显著高于茎材(P<0.05)。
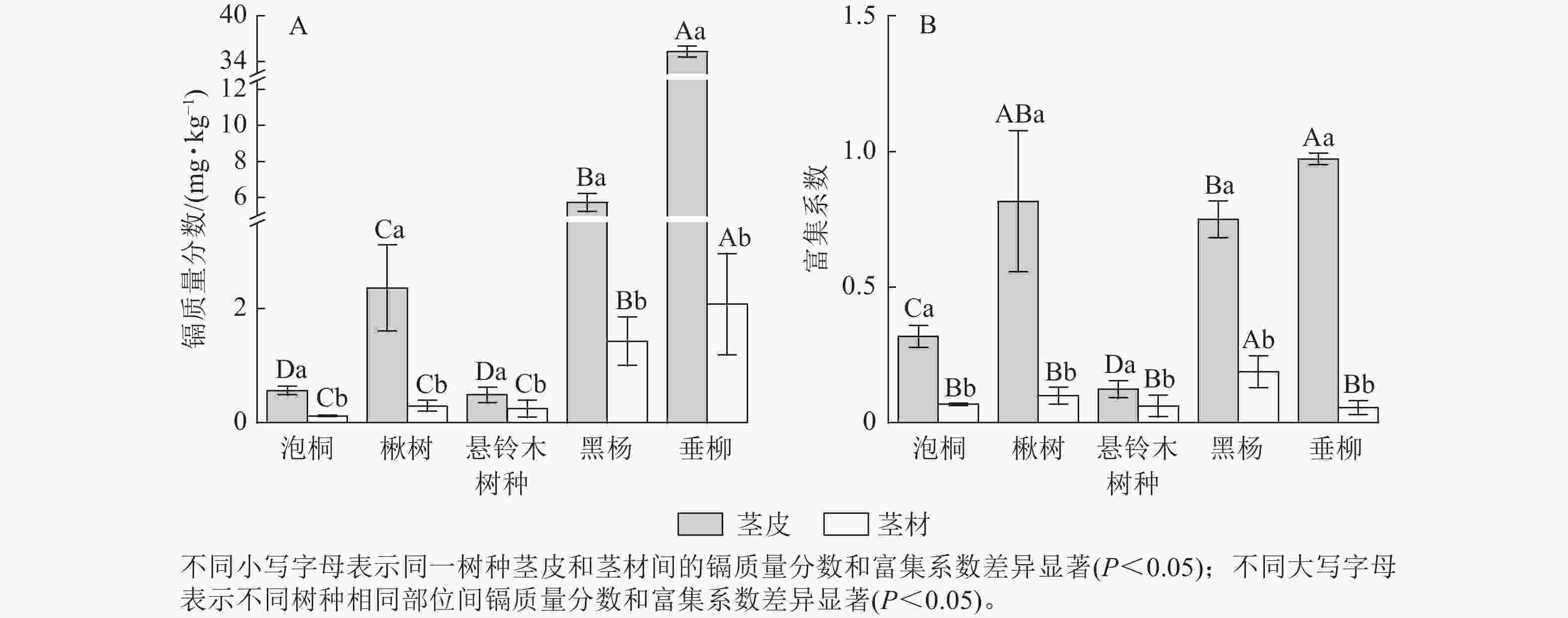
图 3 不同树种茎部的镉质量分数及富集系数
Figure 3. Cd concentration and concentration coefficients in stems of 5 tree species
-
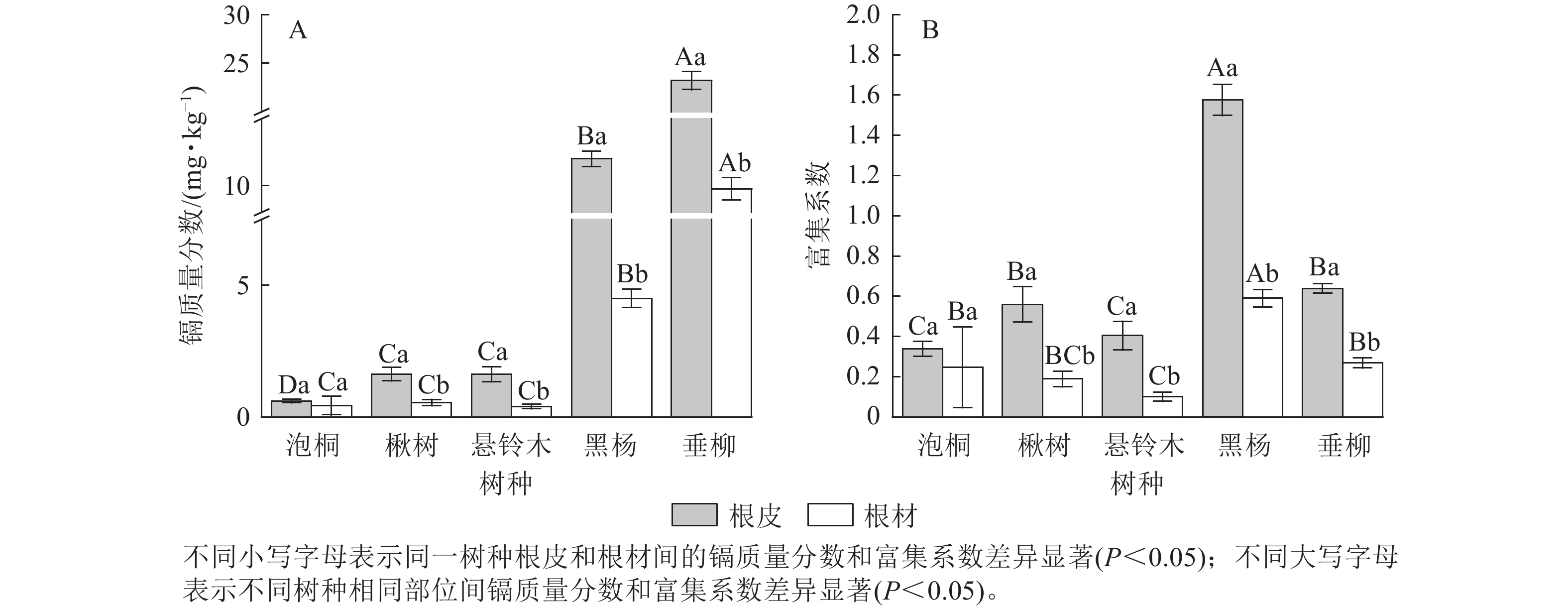
比较5种树种根部的镉质量分数发现(图4A):垂柳根皮的镉质量分数最高,达23.21 mg·kg−1;其次是黑杨,达12.01 mg·kg−1;泡桐根皮的镉质量分数最低,为0.60 mg·kg−1。垂柳根材的镉质量分数最高,达9.76 mg·kg−1;其次是黑杨,达4.50 mg·kg−1,悬铃木根材的镉质量分数最低,为0.39 mg·kg−1。对比根皮和根材的镉质量分数发现:楸树、悬铃木、黑杨和垂柳根皮的镉质量分数显著高于根材的镉质量分数(P<0.05),泡桐根皮与根材的镉质量分数差异不显著。比较不同树种根部的富集系数发现(图4B):黑杨根皮的富集系数最高,达1.58;其次是垂柳,达0.64;泡桐根皮的富集系数最低,为0.34。黑杨根材的富集系数最高,达0.59,其次是垂柳和泡桐,分别达0.27、0.25;悬铃木根材的富集系数最低,为0.10。对比根皮与根材的富集系数发现:除泡桐外,所有树种根皮的富集系数均显著高于根材的富集系数(P<0.05)。
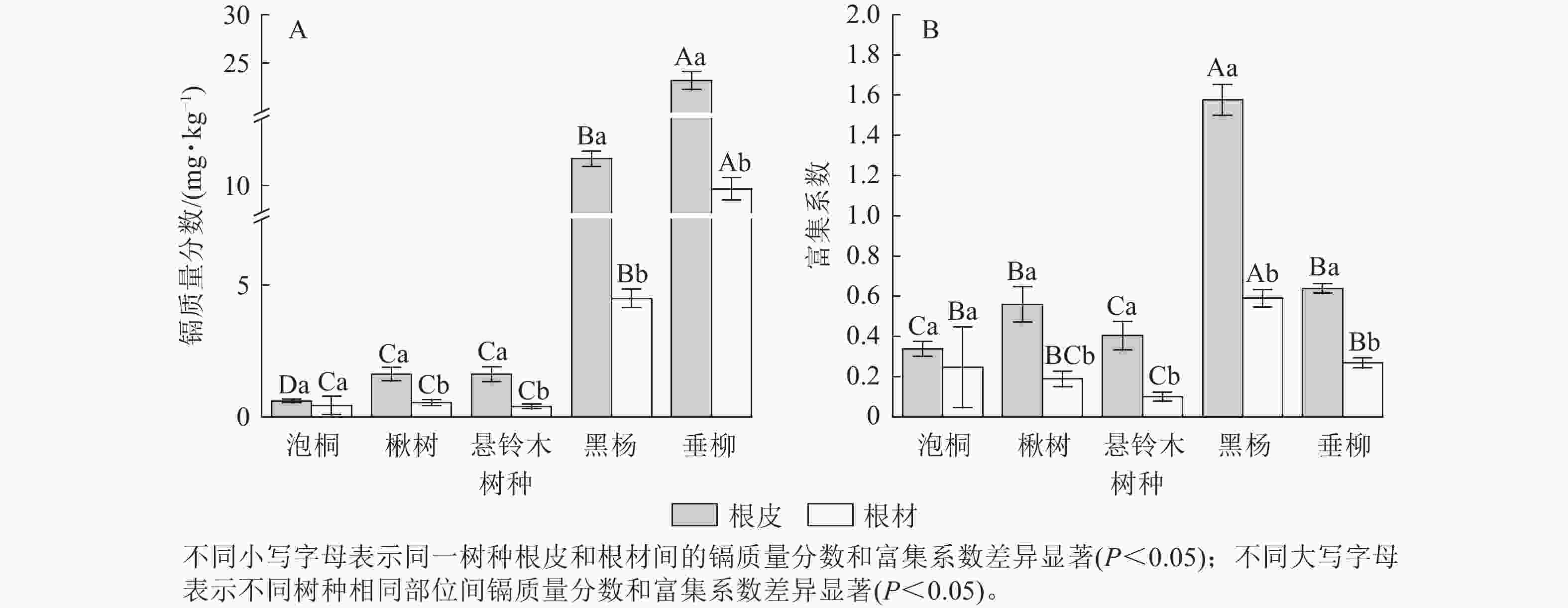
图 4 不同树种根部的镉质量分数及富集系数
Figure 4. Cd concentration and concentration coefficients in roots of 5 tree species
-
利用异速生长方程计算出每株树的叶部、枝部、茎部、根部以及整株的生物量(表1),并进一步计算得到不同部位的积累量及积累总量(表2),黑杨和垂柳不同部位镉积累量从大到小依次表现为茎部、根部、枝部、叶部;楸树不同部位镉积累量从大到小依次表现为茎部、枝部、根部、叶部;泡桐和悬铃木不同部位镉积累量从大到小依次表现为枝部、茎部、根部、叶部。
表 1 不同树种不同部位及整株的生物量
Table 1. Biomass of different parts of the tree and the whole plant
树种 叶部生物量/kg 枝部生物量/kg 茎部生物量/kg 根部生物量/kg 整株生物量/kg 泡桐 6.22±0.77 a 28.59±4.73 a 31.54±6.16 bc 17.11±2.88 b 71.72±13.08 b 楸树 2.59±0.63 b 6.86±1.89 c 22.38±6.10 c 10.42±2.74 b 42.25±11.13 b 悬铃木 6.93±2.10 a 15.30±5.51 b 79.63±29.16 a 27.39±7.74 a 181.28±68.93 a 黑杨 2.95±0.66 b 11.32±3.91 bc 48.76±16.96 b 14.83±2.92 b 79.36±24.69 b 垂柳 1.79±0.38 b 7.03±2.14 c 31.64±10.55 bc 12.78±3.64 b 65.15±20.08 b 说明:不同字母表示不同树种间相同部位及整株的生物量差异显著 (P<0.05)。 表 2 不同树种不同部位及整株镉积累量
Table 2. Cd accumulation in different parts and whole trees
树种 叶部 枝部 茎部 根部 积累总量/mg 平均镉质量分
数/(mg·kg−1)积累
量/mg平均镉质量分
数/(mg·kg−1)积累
量/mg平均镉质量分
数/(mg·kg−1)积累
量/mg平均镉质量分
数/(mg·kg−1)积累
量/mg泡桐 0.45 2.80±0.34 c 0.72 20.59±3.40 b 0.34 10.72±2.09 c 0.52 8.89±1.50 c 43.01±7.34 c 楸树 2.05 5.31±1.29 c 2.52 17.29±4.76 b 1.33 29.77±8.12 c 1.08 11.25±2.96 c 63.62±16.85 c 悬铃木 1.71 11.86±3.60 b 2.25 34.42±12.40 b 0.37 29.46±10.79 c 1.00 27.39±7.74 c 103.13±34.52 c 黑杨 7.52 22.19±4.99 a 6.48 73.36±25.37 a 3.58 174.57±60.71 b 8.25 122.38±24.09 b 392.51±115.15 b 垂柳 10.52 18.82±4.05 a 9.32 65.48±19.94 a 18.69 591.29±197.21 a 16.48 210.68±59.93 a 886.28±281.11 a 说明:不同字母表示不同树种间相同部位镉积累量及积累总量差异显著(P<0.05)。 比较镉在5种树种不同部位中镉的相对含量发现(图5):泡桐不同部位镉的相对含量从大到小依次为枝部、茎部、根部、叶部;楸树不同部位镉的相对含量从大到小依次为茎部、枝部、根部、叶部;悬铃木不同部位镉的相对含量从大到小依次为茎部、根部、枝部、叶部;黑杨不同部位镉的相对含量从大到小依次为枝部、根部、茎部、叶部;垂柳不同部位镉的相对含量从大到小依次为茎部、根部、枝部、叶部。总体上看,楸树、悬铃木、垂柳均表现为茎部镉的相对含量最高,叶部镉的相对含量最低。
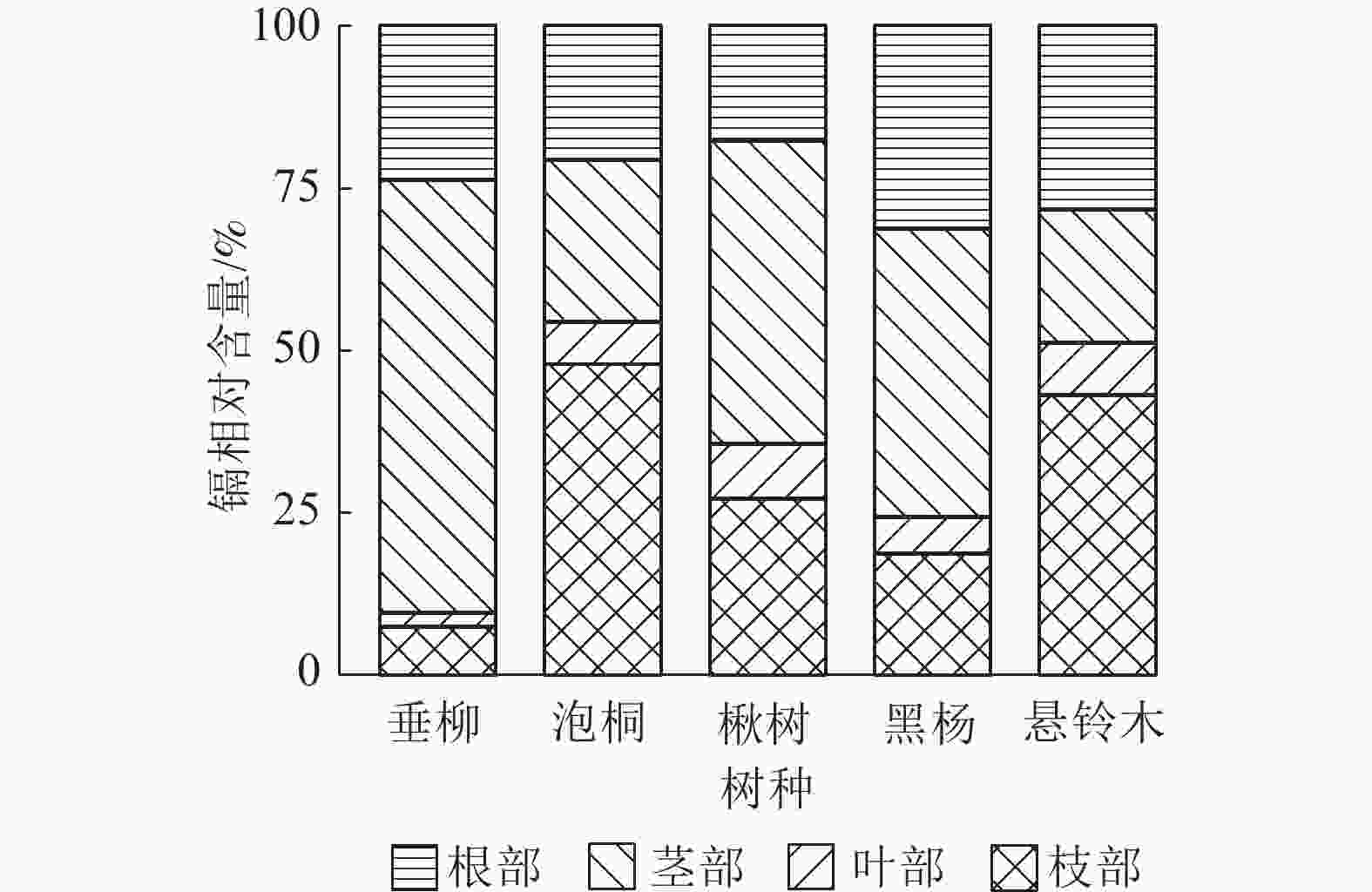
图 5 不同树种不同部位镉的分配
Figure 5. Distribution of cadmium in different parts of 5 tree species
-
本研究表明:不同树种对重金属的富集系数不同,黑杨叶片和叶柄的富集系数均为最大;黑杨枝皮和枝材的富集系数均最大;垂柳茎皮的富集系数最大,黑杨茎材的富集系数最大;黑杨根皮和根材的富集系数均最大。由此可见, 5种供试树种对镉的吸收特征不同。在这些树种中,黑杨和垂柳均有较高的镉积累量,这与前人研究结果类似[11−12],且黑杨和垂柳具有易成活、生长快、生态价值高等优势,因此在镉污染土壤修复中具有较大的推广应用价值。
积累量在重金属积累的植物中相当可观。镉超积累植物龙葵 Solanum nigrum在镉质量分数为2.00 mg·kg−1的土壤中生长86 d后积累量达0.025 mg·株−1[13];镉超积累植物商陆 Phytolacca acinosa在镉质量分数为5.00 mg·kg−1的土壤中生长60 d后地上部(镉的主要储存部分)的积累量达7.58 mg·株−1[14];镉超积累植物东南景天 Sedum alfredii在镉质量分数为2.47 mg·kg−1的土壤中生长210 d后地上部的积累量达9.60 μg·株−1[15]。树木相比超积累植物拥有更长的生长周期、更大的生物量和更高的单株镉积累量,因此具有较大的镉污染修复潜力。在树木吸收镉的研究方面,前人也做过类似的研究。徐爱春[16]研究发现:旱柳 Salix matsudana在镉质量分数为20 mg·L−1的营养液中培养一段时间后,积累量达25.81 mg·株−1。周洁等[17]研究杂交柳(耳柳Salix aurita×银柳Salix argyracea)在镉质量分数为5.28 mg·kg−1的田间土壤中生长1 a后,镉的最大积累量达73.66 mg·株−1。乔木对镉污染土壤修复后再回收利用具有一定的经济效益,比如重金属的回收、植物能源及建材方面的利用[18]。此外,乔木具有多年生、持续修复、不进入食物链的优势,因此在镉污染土壤中植树是一种较为理想的修复技术。
本研究发现:所有树种的根、茎和枝的镉质量分数均表现为树皮部大于木材部,即从大到小依次为根皮、根材、茎皮、茎材、枝皮、枝材。可见,乔木具有优先把镉积累到皮部的特性,这种现象在其他研究中也有发现。例如,唐丽清等[19]研究发现:北京市台基厂大街行道树国槐 Sophora japonica的树皮中镉质量分数高于树干。RODRÍGUEZ等[20]研究表明:西班牙发电厂附近的地中海松Pinus halepensis树皮中镉质量分数高于树干。田胜尼等[21]研究指出:镉质量分数在腺柳 Salix chaenomeloides根皮部最高,达1 438.919 μg·kg−1,乔木中最低,为228.065 μg·kg−1。这可能是乔木的解毒机制导致的。镉主要通过土壤溶液进入根系,并从木质部转移到树皮。由于树皮代谢相对旺盛[22],且树皮中富含有助于与二价离子结合的萜类、软木脂、脂肪酸酶、氨基酸等物质,这些化学物质与镉离子能够结合,生成稳定的络合物,达到解毒目的。而树干中主要为纤维素、半纤维素和木质素,与镉离子能够结合的物质较少,所以只有少部分镉积累在树干中[23]。
-
对河南某处镉污染土壤上栽植的5种树种调查发现:所有树木根皮、茎皮和枝皮均显著大于其对应的根材、茎材和枝材的镉质量分数,这可能是乔木本身的解毒机制造成的。在这些树种中,黑杨和垂柳均有较高的镉积累量,其中黑杨多数部位(除茎皮外)的富集系数均大于其他树种,因此在镉污染土壤修复中具有较大的优势。此外,采用乔木进行镉污染土壤修复时,木材中镉质量分数可达到较高水平,因此在后期木材的加工使用过程中应注意其对人体的健康风险。
Cd accumulation characteristics of different greening tree species
-
摘要:
目的 乔木对重金属具有富集能力,但是目前对其积累特征尚不明确,一定程度上限制了乔木修复重金属污染土壤技术的推广应用。探讨5种乔木绿化树种对镉的富集特征。 方法 分析了镉污染土壤上栽植的泡桐Paulownia fortunei、楸树Catalpa bungei、悬铃木Platanus acerifolia、黑杨Populus nigra 和垂柳Salix babylonica等树木不同部位对镉的富集特征,并分析了不同树种对镉的积累总量。 结果 供试树种对镉均具有一定的积累能力,单株对镉的积累量为43.01~886.28 mg·株−1,其中垂柳对镉的积累总量最大,达886.28 mg·株−1;其次是黑杨,达392.51 mg·株−1。不同树种不同部位对镉的富集能力也不同,泡桐、楸树、悬铃木、黑杨和垂柳不同部位的富集系数分别为0.07~0.56、0.10~1.24、0.06~1.04、0.32~1.58、0.06~0.97。其中黑杨多数部位(除茎皮外)的富集系数均大于其他树种。所有树种根皮、茎皮和枝皮的镉质量分数均显著(P<0.05)大于其对应的根材、茎材和枝材。所有树种不同部位镉质量分数与土壤镉质量分数均呈极显著正相关(P<0.01)。 结论 乔木主要将镉积累到枝部和茎部,其中泡桐、楸树和悬铃木主要积累在枝部,黑杨和垂柳主要积累在茎部。此外,黑杨和垂柳具有较高的镉积累量,是今后镉污染土壤修复中的首选树种。图5表2参23 Abstract:Objective While trees can accumulate heavy metals, their accumulation characteristics are currently unclear; this, to some degree, restricts the application of the technology of using trees to remediate heavy metal-contaminated soil. This study was designed to explore the cadmium (Cd) accumulation capacities of five greening tree species. Method The Cd accumulation characteristics of different parts of various greening trees (Paulownia fortunei, Catalpa bungei, Platanus acerifolia, Populus nigra, and Salix babylonica) planted in Cd-contaminated soil were analyzed, and the total amounts of Cd in the trees were determined. Result All the tested trees had a certain Cd accumulation capacity, and the cumulative amount of Cd per tree ranged from 43.01 to 886.28 mg·plant−1. Among the trees, S. babylonica had the highest accumulation amount of Cd (886.28 mg·plant−1), followed by Populus nigra (392.51 mg·plant−1). The Cd accumulation ability varied among different tree species and parts, with concentration coefficients ranging from 0.07 to 0.56, 0.10 to 1.24, 0.06 to 1.04, 0.32 to 1.58, and 0.06 to 0.97 for different parts of Paulownia fortunei, C. bungei, Platanus acerifolia, Populus nigra, and S. babylonica, respectively. The concentration coefficients of most parts (except for the stem bark) of Populus nigra were higher than those of the other trees. The Cd concentrations in the root bark, stem bark, and branch bark of all trees were significantly higher than those in their corresponding root wood, stem wood, and branch wood (P<0.05). A highly significant positive correlation was found between the Cd contents in different parts of trees and those in the soil (P<0.01). Conclusion The tested tree species accumulated Cd mainly in their branches and stems. Paulownia fortunei, C. bungei, and Platanus acerifolia accumulated Cd mainly in their branches, while Populus nigra and S. babylonica accumulated Cd mainly in their stems. Populus nigra and S. babylonica displayed higher Cd concentrations, which are the preferred tree species for the remediation of Cd-contaminated soil. [Ch, 5 fig. 2 tab. 23 ref.] -
Key words:
- trees /
- cadmium /
- total amount of accumulation /
- concentration coefficient /
- soil remediation
-
图 1 不同树种叶部的镉质量分数及富集系数
Figure 1 Cd concentration and concentration coefficients in leaves of 5 tree species
图 2 不同树种枝部的镉质量分数及富集系数
Figure 2 Cd concentration and concentration coefficients in branches of 5 tree species
图 3 不同树种茎部的镉质量分数及富集系数
Figure 3 Cd concentration and concentration coefficients in stems of 5 tree species
图 4 不同树种根部的镉质量分数及富集系数
Figure 4 Cd concentration and concentration coefficients in roots of 5 tree species
图 5 不同树种不同部位镉的分配
Figure 5 Distribution of cadmium in different parts of 5 tree species
表 1 不同树种不同部位及整株的生物量
Table 1. Biomass of different parts of the tree and the whole plant
树种 叶部生物量/kg 枝部生物量/kg 茎部生物量/kg 根部生物量/kg 整株生物量/kg 泡桐 6.22±0.77 a 28.59±4.73 a 31.54±6.16 bc 17.11±2.88 b 71.72±13.08 b 楸树 2.59±0.63 b 6.86±1.89 c 22.38±6.10 c 10.42±2.74 b 42.25±11.13 b 悬铃木 6.93±2.10 a 15.30±5.51 b 79.63±29.16 a 27.39±7.74 a 181.28±68.93 a 黑杨 2.95±0.66 b 11.32±3.91 bc 48.76±16.96 b 14.83±2.92 b 79.36±24.69 b 垂柳 1.79±0.38 b 7.03±2.14 c 31.64±10.55 bc 12.78±3.64 b 65.15±20.08 b 说明:不同字母表示不同树种间相同部位及整株的生物量差异显著 (P<0.05)。  下载: 导出CSV
下载: 导出CSV
表 2 不同树种不同部位及整株镉积累量
Table 2. Cd accumulation in different parts and whole trees
树种 叶部 枝部 茎部 根部 积累总量/mg 平均镉质量分
数/(mg·kg−1)积累
量/mg平均镉质量分
数/(mg·kg−1)积累
量/mg平均镉质量分
数/(mg·kg−1)积累
量/mg平均镉质量分
数/(mg·kg−1)积累
量/mg泡桐 0.45 2.80±0.34 c 0.72 20.59±3.40 b 0.34 10.72±2.09 c 0.52 8.89±1.50 c 43.01±7.34 c 楸树 2.05 5.31±1.29 c 2.52 17.29±4.76 b 1.33 29.77±8.12 c 1.08 11.25±2.96 c 63.62±16.85 c 悬铃木 1.71 11.86±3.60 b 2.25 34.42±12.40 b 0.37 29.46±10.79 c 1.00 27.39±7.74 c 103.13±34.52 c 黑杨 7.52 22.19±4.99 a 6.48 73.36±25.37 a 3.58 174.57±60.71 b 8.25 122.38±24.09 b 392.51±115.15 b 垂柳 10.52 18.82±4.05 a 9.32 65.48±19.94 a 18.69 591.29±197.21 a 16.48 210.68±59.93 a 886.28±281.11 a 说明:不同字母表示不同树种间相同部位镉积累量及积累总量差异显著(P<0.05)。
下载: 导出CSV
-
[1] 李杨, 丁振军, 王星蒙. 葫芦岛地区土壤镉元素空间含量分布特征及污染评价研究[J]. 环境科学与管理, 2022, 47(10): 166 − 170. LI Yang, DING Zhenjun, WANG Xingmeng. Content distribution characteristics and pollution evaluation of cadmium in soil, Huludao area [J]. Environmental Science and Management, 2022, 47(10): 166 − 170. [2] 顾继光, 周启星, 王新. 土壤重金属污染的治理途径及其研究进展[J]. 应用基础与工程科学学报, 2003, 11(2): 143 − 151. GU Jiguang, ZHOU Qixing, WANG Xin. Reused path of heavy metal pollution in soils and its research advance [J]. Journal of Basic Science and Engineering, 2003, 11(2): 143 − 151. [3] 张云霞, 周浪, 肖乃川, 等. 鬼针草(Bidens pilosa L. )对镉污染农田的修复潜力[J]. 生态学报, 2020, 40(16): 5805 − 5813. ZHANG Yunxia, ZHOU Lang, XIAO Naichuan, et al. Remediation potential of B. pilosa L. in cadmium-contaminated farmland [J]. Acta Ecologica Sinica, 2020, 40(16): 5805 − 5813. [4] 柏佳, 谭长吟, 曹雪莹, 等. 3种有机酸对伴矿景天修复效率及土壤微生物数量的影响[J]. 水土保持学报, 2020, 34(2): 318 − 324. BAI Jia, TAN Changyin, CAO Xueying, et al. Effect of three organic acids on the remediation efficiency of Sedum plumbizincicola and soil microbial quantity [J]. Journal of Soil and Water Conservation, 2020, 34(2): 318 − 324. [5] 张永超, 陈存根, 钟发明, 等. 重金属铅、镉在白榆中分布规律和累积特性研究[J]. 西北林学院学报, 2011, 26(5): 6 − 11. ZHANG Yongchao, CHEN Cungen, ZHONG Faming, et al. Distribution and accumulation of lead and cadmium in Ulmus pumila [J]. Journal of Northwest Forestry University, 2011, 26(5): 6 − 11. [6] 曲豪杰, 张涵丹, 谭志超, 等. 纳塔栎和柳叶栎对铅锌矿区污染土壤的修复潜力分析: 田间试验[J]. 生态学报, 2020, 40(19): 6969 − 6981. QU Haojie, ZHANG Handan, TAN Zhichao, et al. The remediation potential of Quercus nuttallii and Quercus phellos to heavy metalpolluted soil in lead-zinc mining area: a field test [J]. Acta Ecologica Sinica, 2020, 40(19): 6969 − 6981. [7] 李培坤, 牛帅, 王雪颖, 等. 河南农业大学文化路校区校园植物调查与分析[J]. 河南农业大学学报, 2017, 51(2): 243 − 249. LI Peikun, NIU Shuai, WANG Xueying, et al. lnvestigation and analysis of campus plants in Wenhua Road Campus of Henan Agricultural University [J]. Journal of Henan Agricultural University, 2017, 51(2): 243 − 249. [8] 张欣. 基于树木年轮信息的北京园林树木碳固定研究[D]. 保定: 河北农业大学, 2021. ZHANG Xin. Research on Landscape Trees Carbon Fixation in Beijing Based on Tree Rings Information [D]. Baoding: Hebei Agricultural University, 2021. [9] 罗云建, 王效科, 逯非. 中国主要林木生物量模型手册[M]. 北京: 中国林业出版社, 2015. LUO Yunjian, WANG Xiaoke, LU Fei. Comprehensive Database of Biomass Regressions for China’s Tree Species [M]. Beijing: China Forestry Publishing House, 2015. [10] 周国逸, 尹光彩, 唐旭利, 等. 中国森林生态系统碳储量——生物量方程[M]. 北京: 科学出版社, 2018. ZHOU Guoyi, YIN Guangcai, TANG Xuli, et al. Carbon Storage-Biomass Equation of Forest Ecosystems in China [M]. Beijing: Science Press, 2018. [11] 张晓丽, 翟飞飞, 李伟, 等. 27个柳树无性系对镉的吸收分配特性[J]. 林业科学, 2017, 53(4): 9 − 17. ZHANG Xiaoli, ZHAI Feifei, LI Wei, et al. Characteristics of cadmium absorption and distribution in 27 willow clones [J]. Scientia Silvae Sinicae, 2017, 53(4): 9 − 17. [12] 郑青雅, 马云璐, 戎恭, 等. 2年生杨树吸收和富集土壤镉特性研究[J]. 安徽农业大学学报, 2023, 50(2): 297 − 302. ZHENG Qingya, MA Yunlu, RONG Gong, et al. Study on the uptake and enrichment of heavy metal Cd in two-year old poplar trees [J]. Journal of Anhui Agricultural University, 2023, 50(2): 297 − 302. [13] 李志贤, 冯涛, 陈章, 等. 镉胁迫对龙葵镉的吸收积累及生理响应的影响[J]. 水土保持学报, 2017, 31(5): 328 − 333. LI Zhixian, FENG Tao, CHEN Zhang, et al. Effects of different levels Cd stress on Cd uptake and physiological response of Solanum nigrum L. [J]. Journal of Soil and Water Conservation, 2017, 31(5): 328 − 333. [14] 聂发辉, 吴彩斌, 吴双桃. 商陆对镉的富集特征[J]. 浙江林学院学报, 2006, 23(4): 400 − 405. NIE Fahui, WU Caibin, WU Shuangtao. Characteristics of Phytolacca acinosa accumulating Cd [J]. Journal of Zhejiang Forestry College, 2006, 23(4): 400 − 405. [15] 刘波, 陈绩, 马嘉伟, 等. 交流电场和水分对褪色柳-东南景天混栽修复镉污染土壤的影响[J]. 浙江农林大学学报, 2021, 38(6): 1238 − 1244. LIU Bo, CHEN Ji, MA Jiawei, et al. Effects of AC electric field and soil moisture on phytoremediation of cadmium contaminated soil by mixed planting of Salix discolor and Sedum alfredii [J]. Journal of Zhejiang A&F University, 2021, 38(6): 1238 − 1244. [16] 徐爱春. 柳树无性系镉积累和生理变化规律研究[D]. 北京: 中国林业科学研究院, 2007. XU Aichun. Study on Cd Accumulation and Physiological Change Rule in Clones of Salix [D]. Beijing: China Academy of Forestry, 2007. [17] 周洁, 陈庆生, 王保松, 等. 镉超富集柳树对镉吸收、积累及分配特性的田间试验研究[J]. 江苏林业科技, 2023, 50(1): 1 − 6. ZHOU Jie, CHEN Qingsheng, WANG Baosong, et al. Characteristics of cadmium absorption, accumulation and distribution in cadmium-tolerant willows of field test [J]. Journal of Jiangsu Forestry Science &Technology, 2023, 50(1): 1 − 6. [18] 钟斌, 陈俊任, 彭丹莉, 等. 速生林木对重金属污染土壤植物修复技术研究进展[J]. 浙江农林大学学报, 2016, 33(5): 899 − 909. ZHONG Bin, CHEN Junren, PENG Danli, et al. Research progress of heavy metal phytoremediation technology of fast-growing forest trees in soil [J]. Journal of Zhejiang A&F University, 2016, 33(5): 899 − 909. [19] 唐丽清, 邱尔发, 韩玉丽, 等. 不同径级国槐行道树重金属富集效能比较[J]. 生态学报, 2015, 35(16): 5353 − 5363. TANG Liqing, QIU Erfa, HAN Yuli, et al. Variation of heavy metal enrichment efficiency in roadside trees of Sophora japonica L. with different diameters at breast height [J]. Acta Ecologica Sinica, 2015, 35(16): 5353 − 5363. [20] RODRÍGUEZ M, JOSE A, GUTIERREZ, et al. Wood and bark of Pinus halepensis as archives of heavy metal pollution in the Mediterranean Region [J]. Environmental Pollution, 2018, 239: 438 − 447. [21] 田胜尼, 张静, 孙庆业, 等. 铜尾矿自然定居腺柳对重金属吸收及分布的研究[J]. 农业环境科学学报, 2013, 32(9): 1771 − 1777. TIAN Shengni, ZHANG Jing, SUN Qingye, et al. Heavy metals absorption and distribution by Salix chaenomeloides settled naturally on the copper tailings [J]. Journal of Agro-Environment Science, 2013, 32(9): 1771 − 1777. [22] 王荣芬, 邱尔发, 唐丽清, 等. 行道树毛白杨树干中重金属元素分布[J]. 生态学报, 2014, 34(15): 4212 − 4222. WANG Rongfa, QIU Erfa, TANG Liqing, et al. Heavy metals distribution in roadside tree trunks under a heavy urban transportation condition [J]. Acta Ecologica Sinica, 2014, 34(15): 4212 − 4222. [23] GONZALEZ-OREJA J A, ROZAS M A, ALKORTA I, et al. Dendroremediation of heavy metal polluted soils [J]. Reviews on Environmental Health, 2008, 23(3): 223 − 234. -

-
链接本文:
https://zlxb.zafu.edu.cn/article/doi/10.11833/j.issn.2095-0756.20230630
 点击查看大图
点击查看大图
计量
- 文章访问数: 1295
- HTML全文浏览量: 365
- PDF下载量: 25
- 被引次数: 0
