




-
土壤作为环境污染物的重要载体,正面临着日益严重的污染挑战。其中,重金属污染已经成为中国当前最突出的土壤污染问题之一[1]。土壤中的砷(As)、铅(Pb)等重金属污染具有持久性、不可逆性和隐蔽性,对土壤质量和土壤生产力造成极大的负面影响。此外,由于水稻Oryza sativa具有较强的重金属积累能力,As和Pb可以通过食物链对人体和其他生物体健康构成严重威胁[2]。
生物质炭是生物质在缺氧或低氧条件下热解的产物,具有发达的孔隙结构和丰富的表面官能团,能够通过物理吸附、离子交换和共沉淀等机制降低土壤中重金属的生物有效性[3]。然而,由于比表面积、成分组成和表面官能团等物理化学特性的限制,其对污染土壤中重金属的吸附能力有限,且选择性较差。YANG等[4]研究发现:由于生物质炭表面带有负电荷,其对以阴离子形式存在的污染物(如As)的钝化能力相对较弱。为了提高生物质炭对重(类)金属复合污染土壤的修复能力,对生物质炭与其他材料进行功能化改性成为了新的研究方向。
铁基材料因其成本低、制备工艺简易、种类多样及较低的毒性,成为生物质炭改性中广泛应用的材料之一[5]。改性材料主要通过离子交换和沉淀作用钝化土壤中的重金属。WEN等[6]研究表明:使用氯化铁改性后的园林废弃物生物质炭可以将As(Ⅲ)氧化为移动性较低的As(V),从而有效减少水稻对As的吸收,改善水稻的生长情况。聚合硫酸铁(polyferric sulfate,PFS)作为一种无机高分子混凝剂,相比其他铁基材料,具有成本低、水解速度快及絮凝体密度大等优点,在吸附去除污水废水中重金属离子方面表现出显著的功效[7]。有研究指出:将活化硅酸与PFS联合使用能够提高其修复效率[8]。目前,关于PFS改性生物质炭的研究主要集中在对水体污染物的去除作用,以及As单一污染土壤的修复应用,而在As和Pb复合污染土壤中的作用机制以及对作物吸收重金属的改善效应的研究相对较少。因此,深入研究PFS改性生物质炭与复合污染土壤中重金属的相互作用及其改善效应,对于提高土壤修复技术的有效性至关重要。同时,随着中国园林绿化面积的增加,园林废弃物的产量亦随之上升,探索更为环保的处理方法对实现园林废弃物的资源化及减少对环境的影响具有重要意义。
鉴于上述研究背景,本研究选用细叶榕Ficus microcarpa枝条为原材料制备生物质炭,并采用聚合硫酸铁{[Fe2(OH)n(SO4)3-n/2]m}进行改性,制得聚合硫酸铁改性细叶榕生物质炭(Fe-FMB),进行水稻盆栽试验,重点探讨这2种生物质炭与土壤中As和Pb的相互作用关系,评估对土壤酶活性的影响,同时考察水稻各器官内As和Pb的富集情况。还研究了2种生物质炭对土壤理化性质及养分的影响,旨在为聚合硫酸铁基生物质炭在重(类)金属复合污染土壤修复中的应用提供理论支持和科学依据。
-
土壤采集:本研究所用土壤采自浙江省绍兴市某铅锌矿区附近的水稻田0~20 cm土层,地理坐标为29°59′N,120°47′E。所采集土壤中As和Pb的质量分数均超过了GB 15618—2018《土壤环境质量标准》规定的风险筛选值(As:30 mg·kg−1;Pb:100 mg·kg−1)。土壤样品去除碎石和植物残体后风干过2 mm筛备用。供试土壤基本理化性质:pH为5.63,有机质为39.30 g·kg−1,阳离子交换量为12.53 cmol·kg−1,速效钾为107.00 mg·kg−1,速效磷为10.10 mg·kg−1,碱解氮为96.26 mg·kg−1,总As为99.73 mg·kg−1,总Pb为447.84 mg·kg−1。供试植物:特早熟晚粳稻品种‘秀水519’ Oryza sativa ‘Xiushui 519’,种子购自临安绿娃种子有限公司。原始生物质炭制备:细叶榕修剪枝条收集于广东省佛山市某公园,经切碎风干处理后,使用小型炭化设备(ECO-8-10,湖州宜可欧环保科技有限公司)在500 ℃限氧条件下热解2 h,制得细叶榕生物质炭(FMB),生物质炭经研磨后过2 mm筛备用。铁改性生物质炭制备:采用酸化熟化法,通过将工业硅酸钠(Na2O·SiO2)与浓硫酸反应,制得SiO2质量浓度为10%的活化硅酸。对细叶榕修剪枝条进行前处理,然后将碎屑按照炭铁质量比20∶1浸泡于聚合硫酸铁溶液中。为了提升聚合硫酸铁及生物质炭的修复效能,按照硅铁质量比1.5∶1.0向溶液中添加活化硅酸[8],充分搅拌后在105 ℃下烘干至恒量。将处理后的生物质置于同一炭化设备中,同样在500 ℃限氧条件下热解2 h,制得聚合硫酸铁改性细叶榕生物质炭(Fe-FMB)。生物质炭经研磨后过2 mm筛备用。
-
水稻盆栽试验在浙江农林大学温室内进行。分别将质量分数为2%的FMB和Fe-FMB与供试土壤充分混合,以未施加生物质炭的土壤作为对照组,分别在苗期、分蘖期、灌浆期和成熟期4个生长阶段进行取样,每个时期设置3个处理,共12个处理,每个处理设置4个重复,称取5 kg相应处理的土壤装入塑料盆中。
试验开始前,向盆中浇水至田间持水量的70%,并保持稳定7 d。之后,根据当地常用农作方式[9],向土壤中施入基肥(尿素、KH2PO4及KCl),按N∶P2O5∶K2O=1.0∶0.5∶1.0的质量比配置(折合为0.429 0、0.192 0和0.211 4 g·kg−1)。将提前培育的5株长势相似的健康水稻秧苗移栽至盆中,移栽10 d后,再向每盆追施0.257 5 g·kg−1的尿素。
整个水稻生长周期(154 d)采用长期淹水灌溉方式,定期向盆中加入去离子水至土壤被水覆盖。为避免环境误差,盆栽的摆放方式采用随机区组排列,并定期调换各区组盆栽的位置。在苗期、分蘖期、灌浆期和成熟期进行破坏性取样,收集水稻不同器官和土壤样品,进行后续分析测定。
-
测定了土壤pH、电导率、阳离子交换量、有效磷、碱解氮、速效钾等基础理化性质[10]。土壤中有效硅及各形态硅用钼蓝比色法测定。土壤中有效砷和铅分别采用NH4H2PO4和二乙三胺五乙酸(DTPA)浸提法提取。
采用荧光微孔板检测技术测定土壤β-1,4-葡萄糖苷酶(β-1,4-glucosidase, BG)、亮氨酸氨基肽酶(leucine amino peptidase, LAP)和酸性磷酸酶(acid phosphatase, PHOS)的酶活性[11]。过氧化氢酶(catalase, CAT)活性采用高锰酸钾滴定法测定。测定生物质炭的pH、电导率、灰分、比表面积及碳(C)和氮(N)等元素,其中元素以质量分数的形式表示。同时通过傅里叶红外光谱仪(FTIR)、X射线衍射分析(XRD)和扫描电子显微镜(SEM)对生物质炭表面官能团及形貌特征进行分析。生物质炭中总铁、总硅用硝酸-氢氟酸-高氯酸三酸消解法测定。
-
水稻收获后使用自来水及离子水冲洗干净,以去除表面残留物,将样品放入105 ℃的烘箱中杀青处理30 min。随后,在65 ℃的条件下烘干至恒量。烘干后的水稻样品记录生物量,研磨过0.25 mm的筛网备用。采用硝酸-过氧化氢消煮法对水稻各器官样品进行预处理,随后采用ICP-MS测定样品中As和Pb的质量分数。
-
数据处理和统计分析通过Excel和SPSS 19.0完成。土壤理化性质及重金属有效态的质量分数差异通过单因素方差分析(one-way ANOVA)评估;不同处理之间的差异显著性采用Duncan多重比较确定;采用Pearson相关分析法探究各变量间的相关性。所有的分析结果均以显著性水平P<0.05为标准判断。图表采用Origin 2022制作。
-
由表1可知:Fe-FMB的pH明显低于FMB。这可能是由于浸渍热解过程中生物质炭表面Fe3+的水解作用导致溶液中H+浓度增加,从而增强了溶液的酸性。此外,Fe-FMB显示出更高的灰分和比表面积,这可能是热解过程中铁颗粒与碳质化合物之间碰撞的结果[12]。Fe-FMB的阳离子交换量也明显高于FMB,这可能与其表面含氧官能团的增加有关。
表 1 生物质炭的基本理化性质
Table 1. Selected physicochemical properties of the used biochars
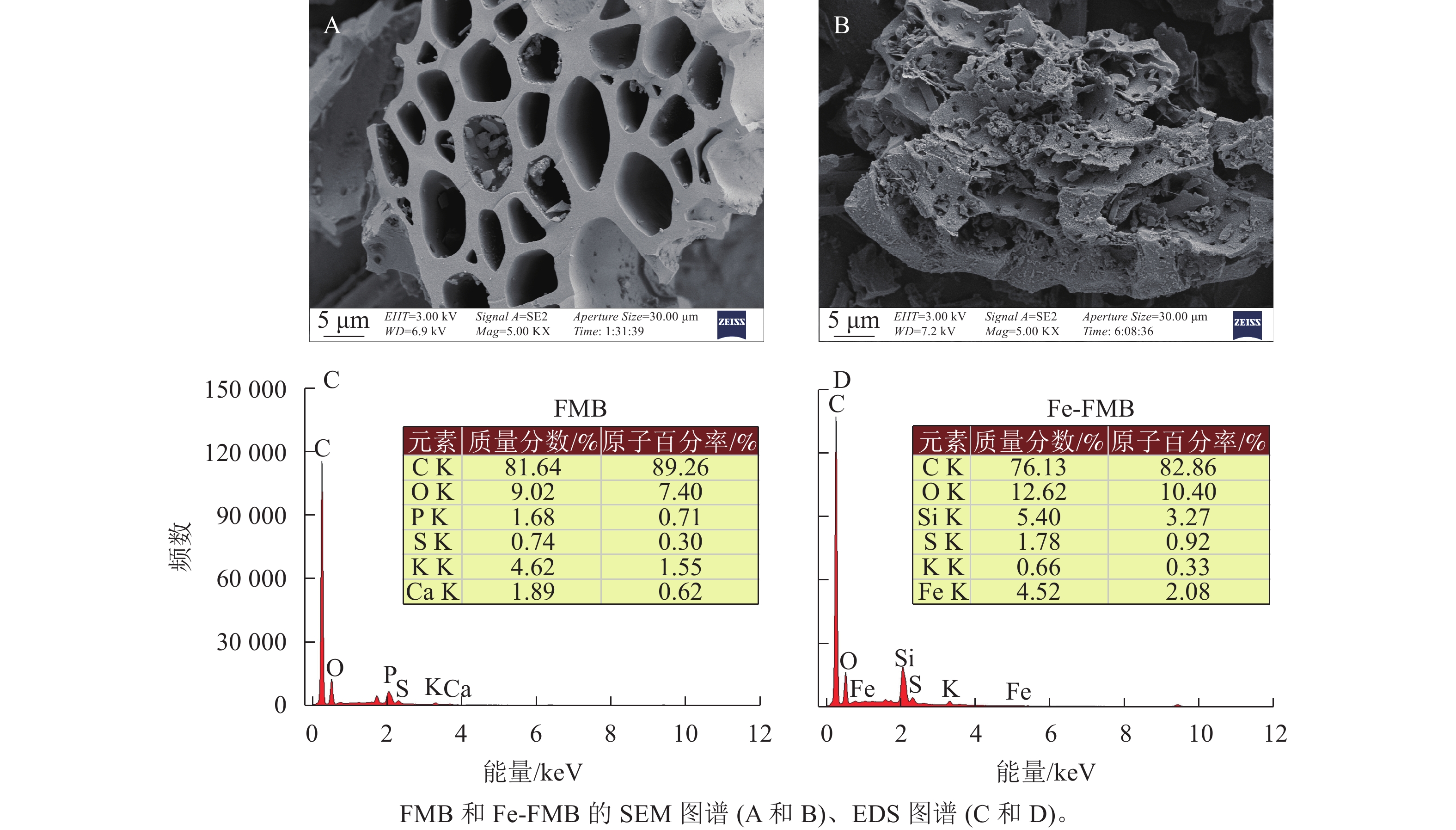
生物质炭 pH 灰分/(g·kg−1) 碳/(g·kg−1) 氢/(g·kg−1) 氮/(g·kg−1) 硅/(g·kg−1) 铁/(g·kg−1) 比表面积/(m2·kg−1) 阳离子交换量/(cmol·kg−1) FMB 9.5 79.3 753 5.0 27.5 1.50 7.89 23.4 14.45 Fe-FMB 5.3 137.8 505 7.5 13.5 7.31 49.32 132.6 30.95 如图1A和B所示:2种生物质炭均展现出均匀排列的管式结构。这一特征可能源于相对较低的热解温度,导致生物质炭本身的导管结构被部分保留下来[13]。相比之下,Fe-FMB的表面更加粗糙,其微孔和管式结构中可见颗粒状物质。根据2种生物质炭的X射线能量色谱仪(EDS)元素分布结果(图1C和D),发现FMB具有更多的钾(K)元素,而在Fe-FMB表面则观察到更多的铁(Fe)、硫(S)和氧(O)元素,Fe-FMB微孔和管式结构颗粒状物质主要由Fe构成的细小颗粒组成。这一结果表明FPS已成功负载于生物质炭表面。
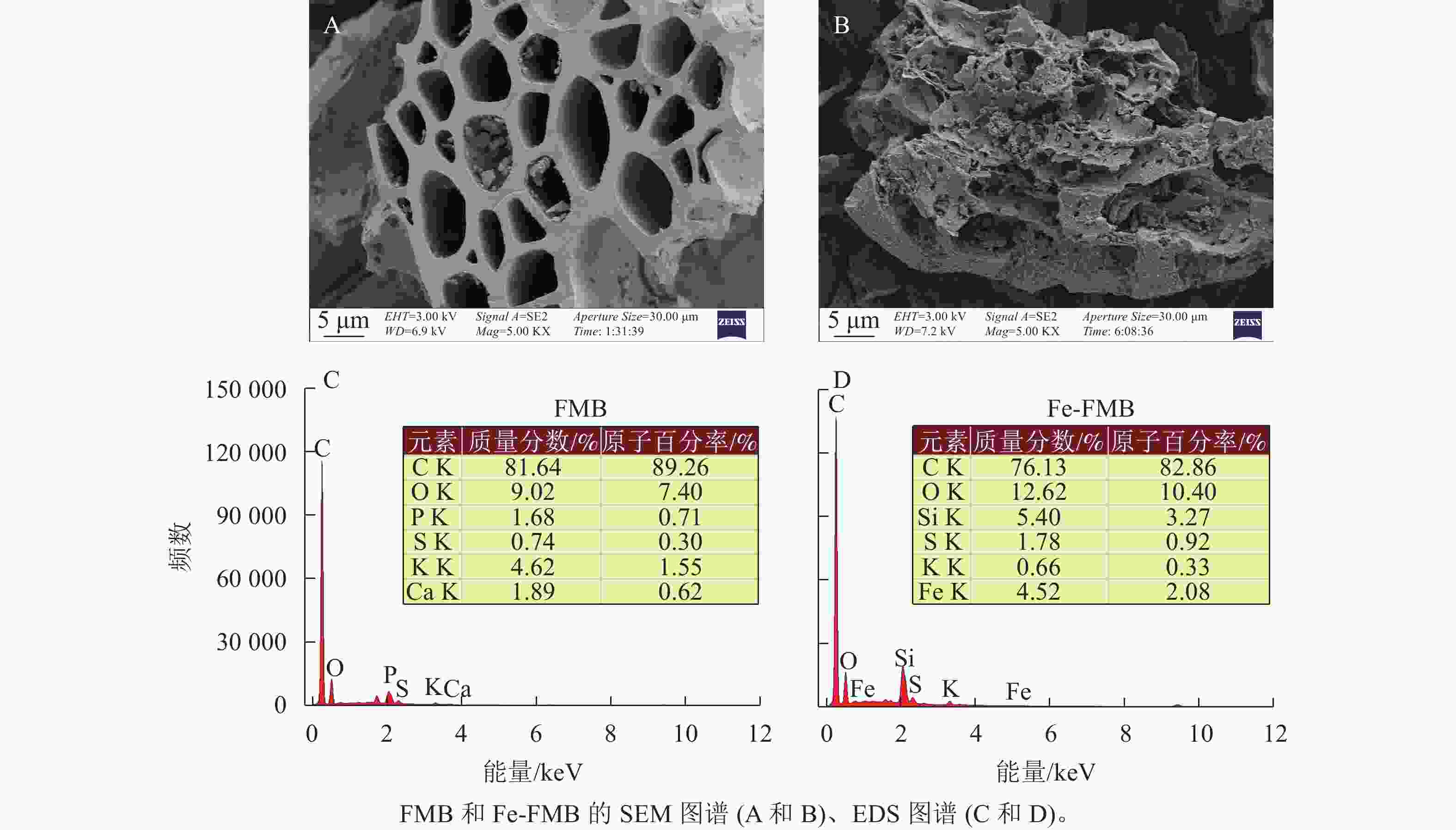
图 1 供试生物质炭的形貌和结构
Figure 1. Morphology and structure of biochar before and after modification
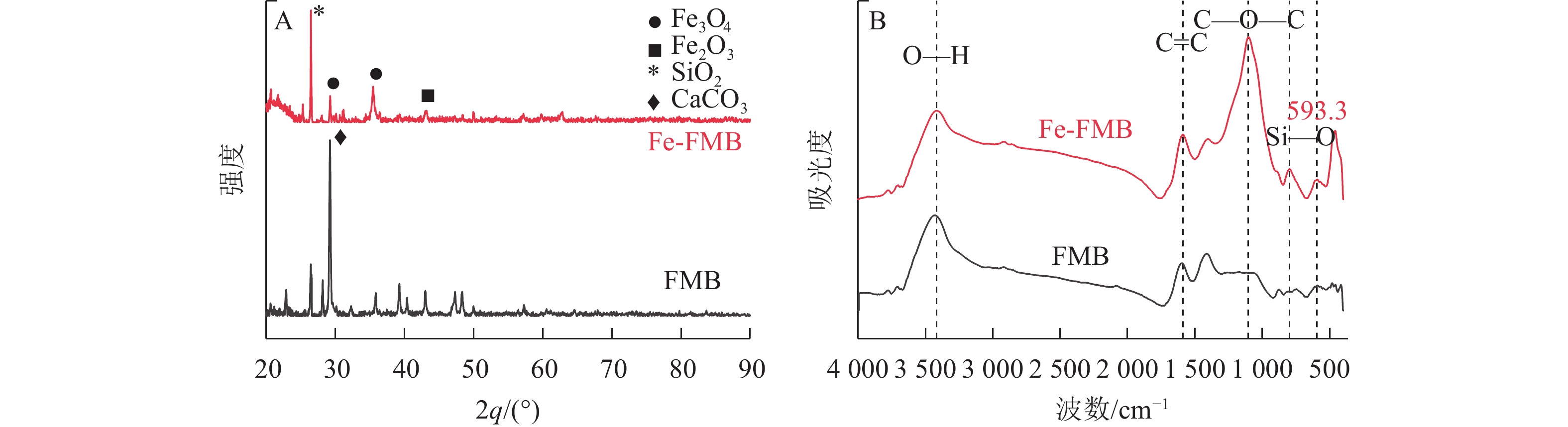
根据X射线衍射分析图谱(图2A),在Fe-FMB的图谱中,在30.15°、35.51°和43.17°处分别观察到了Fe3O4和Fe2O3的特征衍射峰,这些峰值表明Fe成功地负载于生物质炭上,且以Fe3O4为主要形态。同时在Fe-FMB上检测到SiO2特征衍射峰,说明其含有较高的硅(Si)。FTIR分析结果(图2B)显示:Fe-FMB在650~1 000 cm−1以及
1050 cm−1处的特征峰强度高于FMB。这一结果表明:铁改性生物质炭中含氧官能团的伸缩振动增强,含氧官能团数量增加。除此之外,860 cm−1附近的吸收峰可能是由于Si—O官能团的不对称伸缩振动所致[14]。这也证明了铁改性生物质炭中Si质量分数的增加(表1)。在593.3 cm−1处的吸收峰则表明了Fe—O官能团的存在,与Fe—O络合物或铁氧化物的存在相符[15]。
图 2 供试生物质炭的X射线衍射分析图谱(A)和傅里叶红外光谱图(B)
Figure 2. X-ray diffraction (A), and Fourier transform infrared spectra (B) of biochars
-
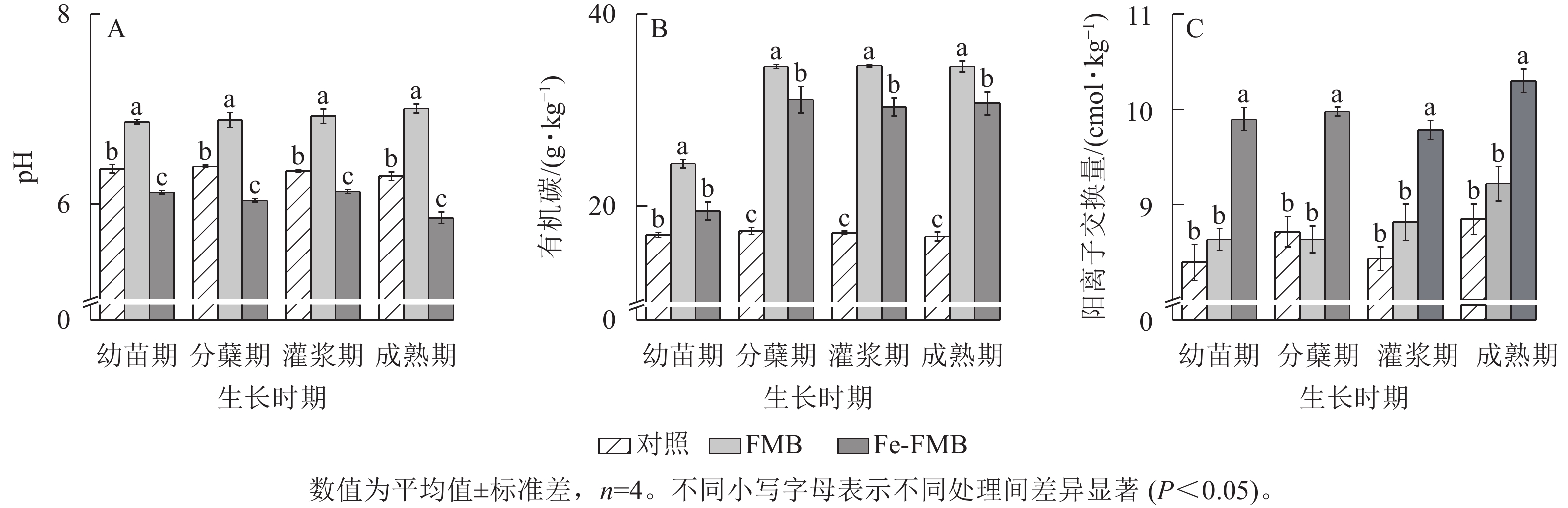
由图3A可见:在水稻生长的各时期,与对照相比,FMB土壤pH提高了0.21~0.49,Fe-FMB土壤pH则显著降低(P<0.05)。与对照相比,2种生物质炭均显著提高了土壤中有机碳质量分数(P<0.05),其中FMB的增幅达43%~105%(图3B)。根据图3C,施用Fe-FMB显著提高了土壤阳离子交换量(1.26~1.57 cmol·kg−1)(P<0.05),在幼苗期增幅最大,与对照相比,增幅达18%。
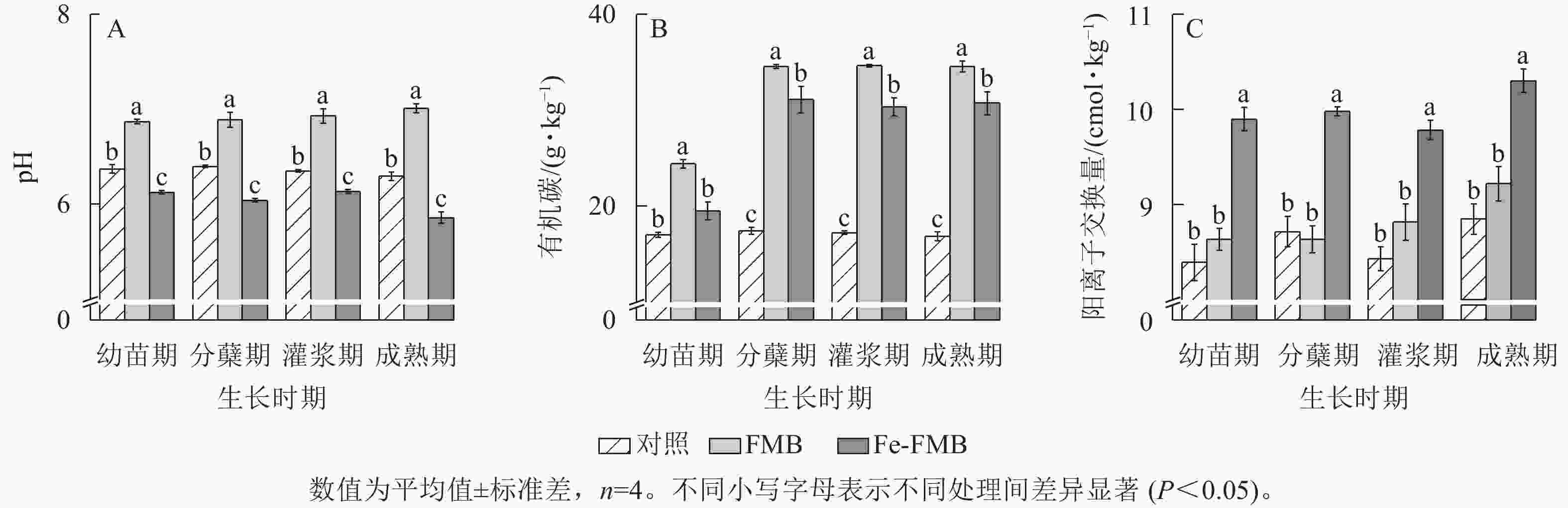
图 3 施入生物质炭对土壤理化性质的影响
Figure 3. Effect of biochar application on soil physiochemitrical properties
-
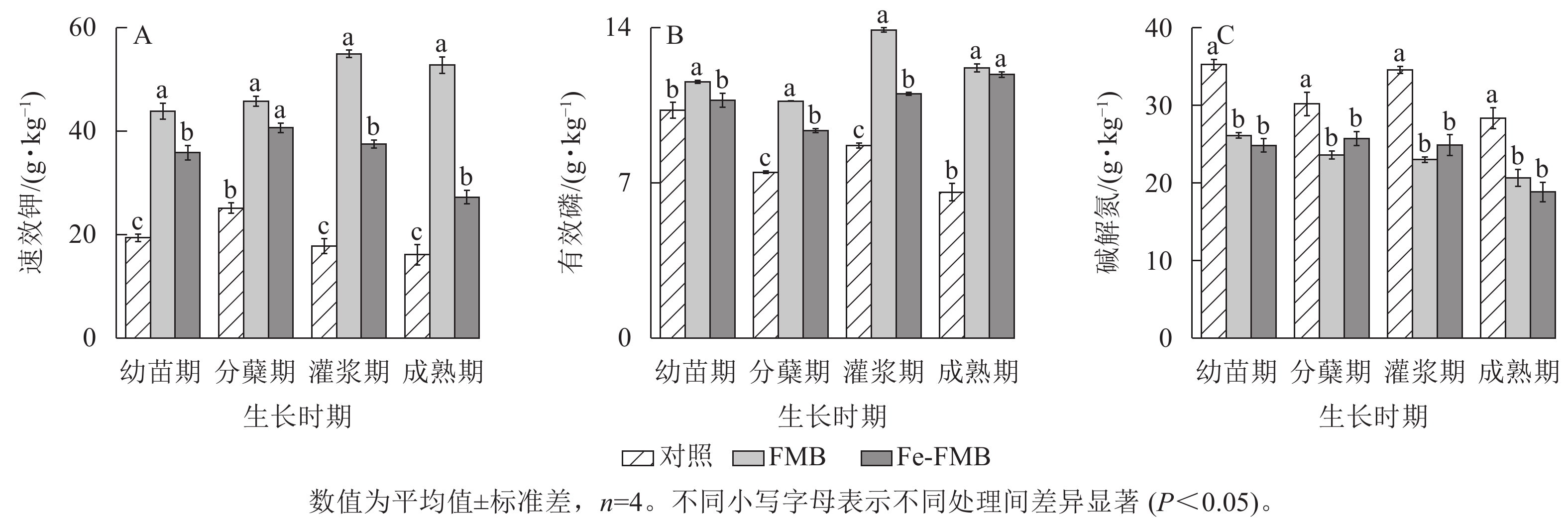
如图4A所示:施用2种生物质炭后,土壤的速效钾质量分数均显著提升(P<0.05)。FMB对速效钾质量分数的提升效果更为显著,与对照相比提升了92%~227%。与对照相比,FMB和Fe-FMB在各个时期均明显提升了土壤中有效磷质量分数,增幅分别为8%~57%和12%~76%(图4B)。施用生物质炭后,土壤的碱解氮质量分数显著下降(图4C,P<0.05)。
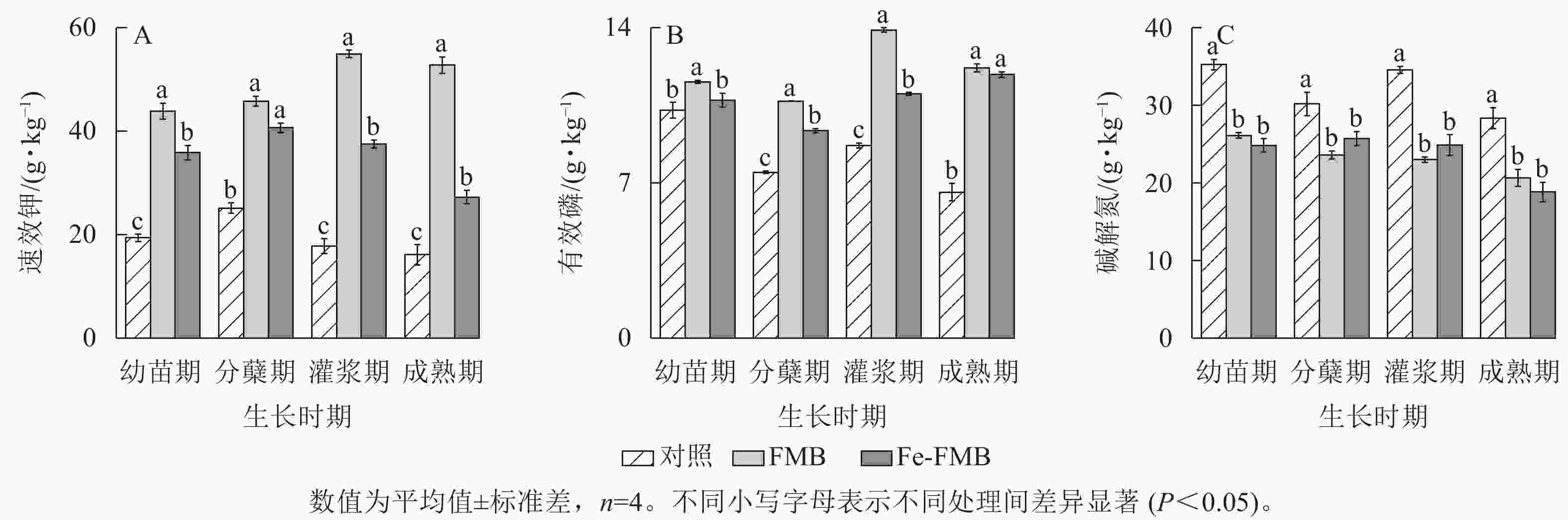
图 4 施入生物质炭对土壤养分的影响
Figure 4. Effect of biochar application on soil N, P and K availability
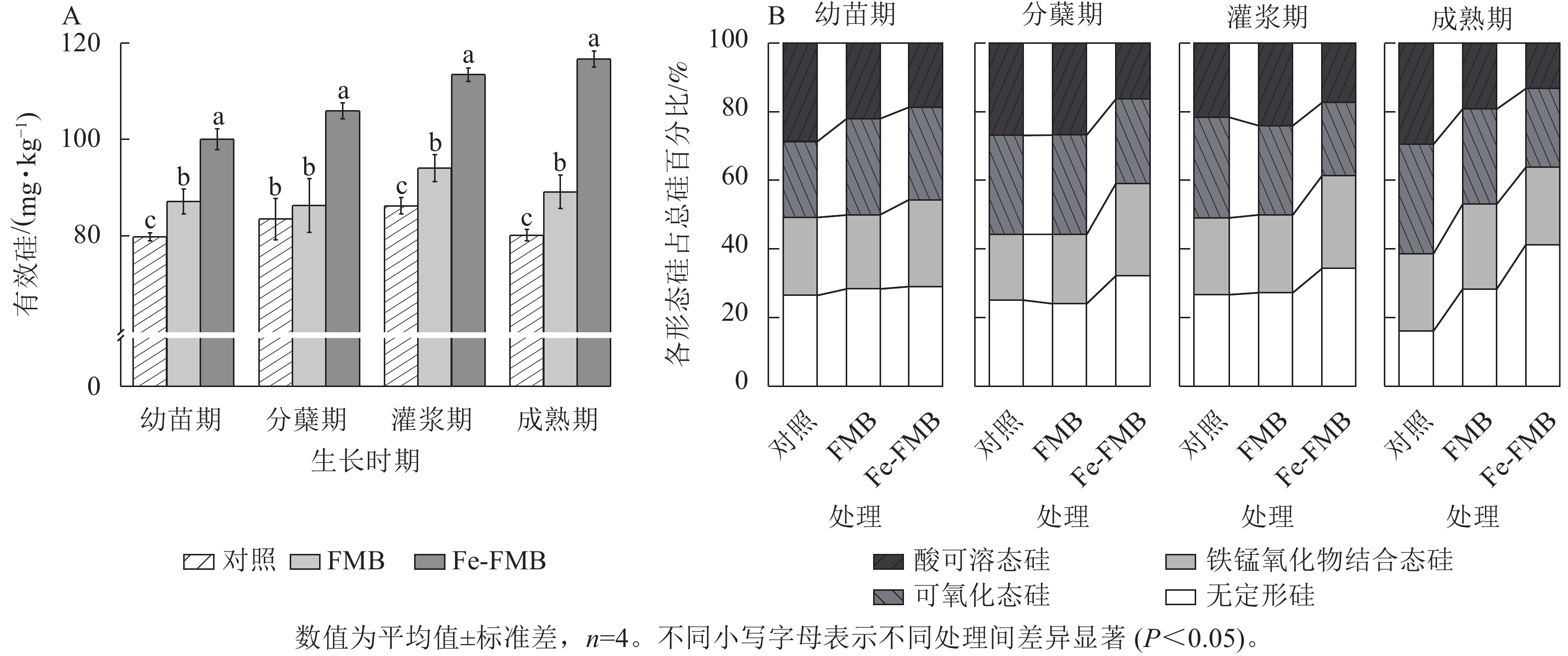
如图5A所示:与对照相比,Fe-FMB显著提升了土壤中有效硅的质量分数(P<0.05),增幅为24%~49%。根据图5B的结果,与对照相比,施用Fe-FMB后,土壤中无定型硅的占比均显著增加(P<0.05),增幅为3%~25%,也显著提高了土壤中铁锰氧化态硅的占比(P<0.05)。
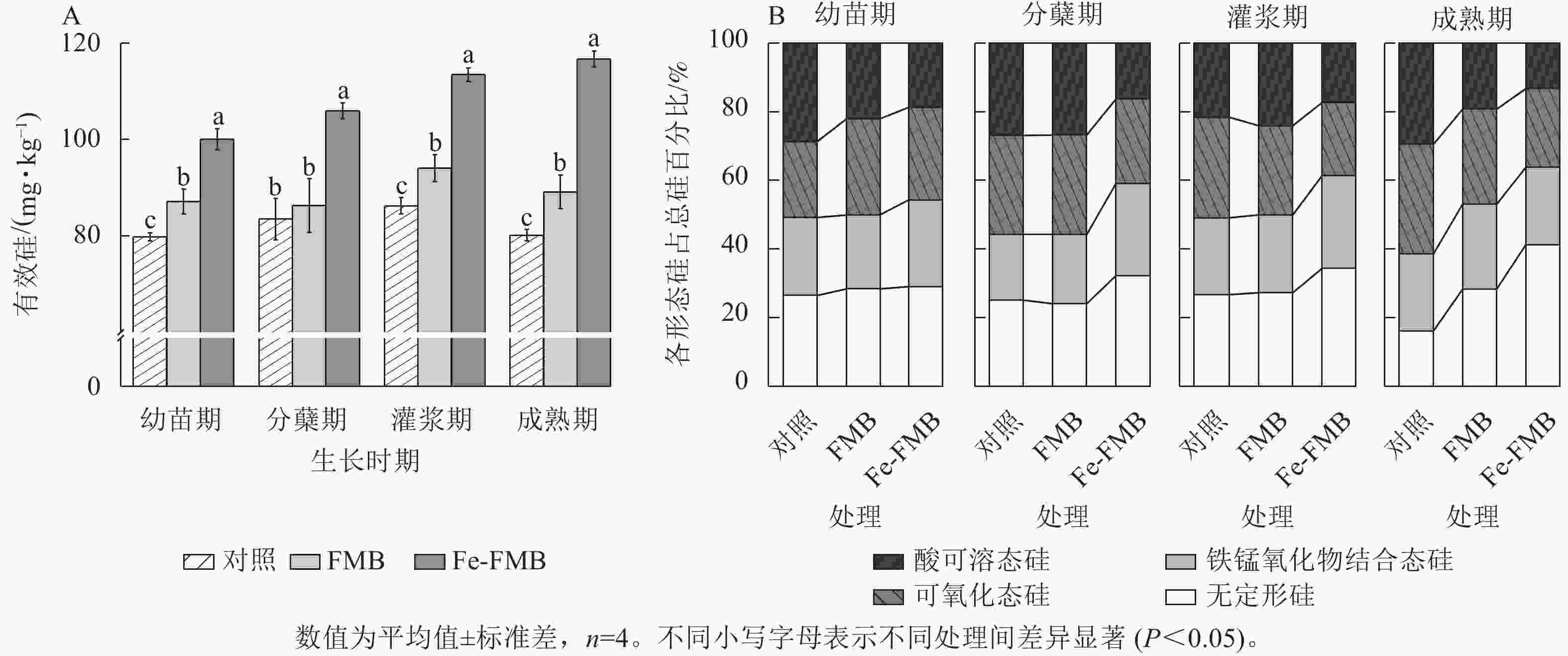
图 5 施用生物质炭对土壤有效硅以及不同形态硅分布的影响
Figure 5. Effect of biochar application on soil Si availability and the fractions of Si in soil
-
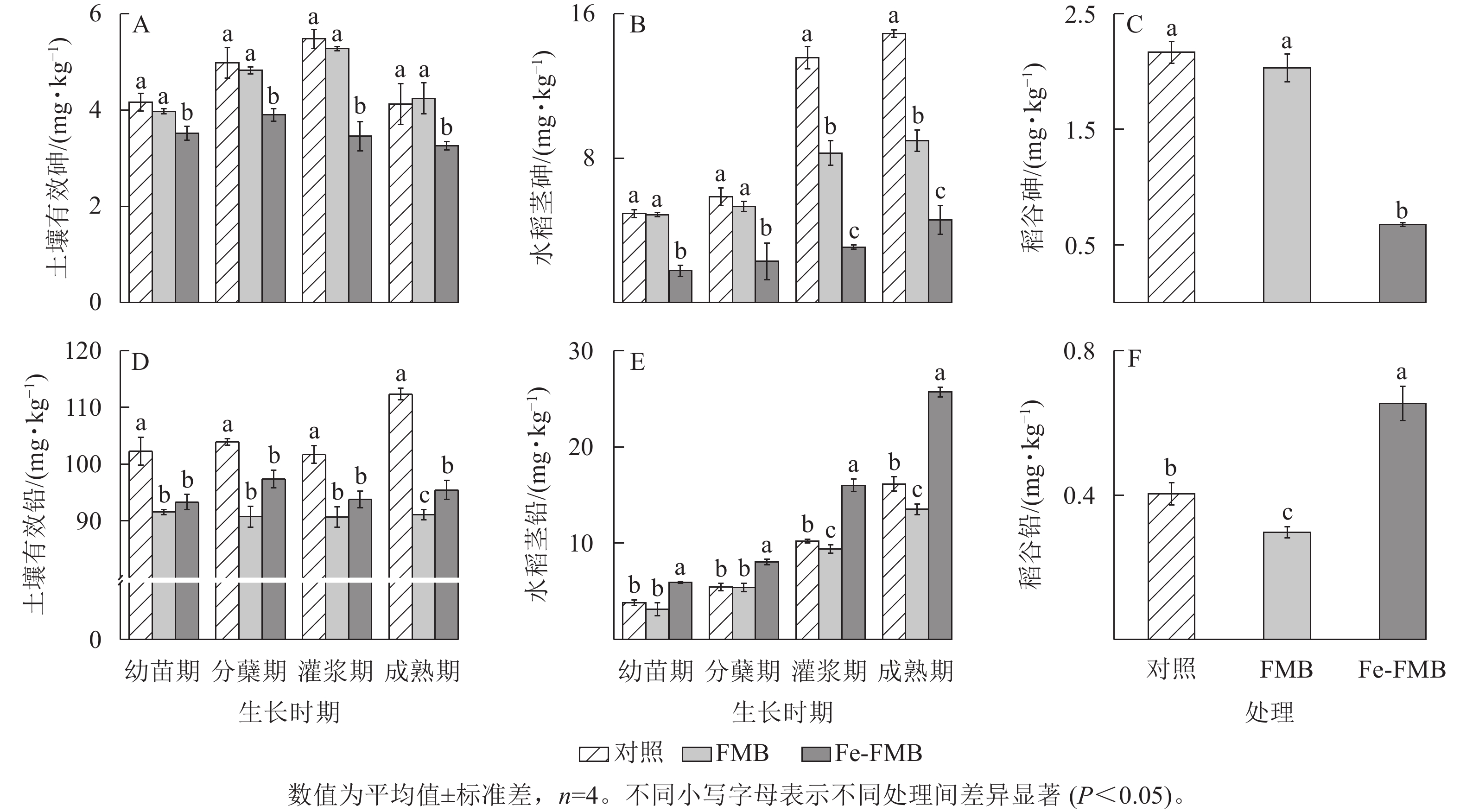
如图6A所示:在水稻生长的各时期,与对照和FMB相比, Fe-FMB显著降低了土壤中有效砷的质量分数(P<0.05),降幅最高达37%,FMB与对照差异不显著。由图6B和图6C可见:Fe-FMB在水稻各生长时期都显著降低了水稻茎、稻谷中的砷质量分数(P<0.05),与对照相比,分别下降了71%~84%和70%;FMB仅在水稻灌浆期和成熟期显著降低了茎的砷质量分数(P<0.05)。
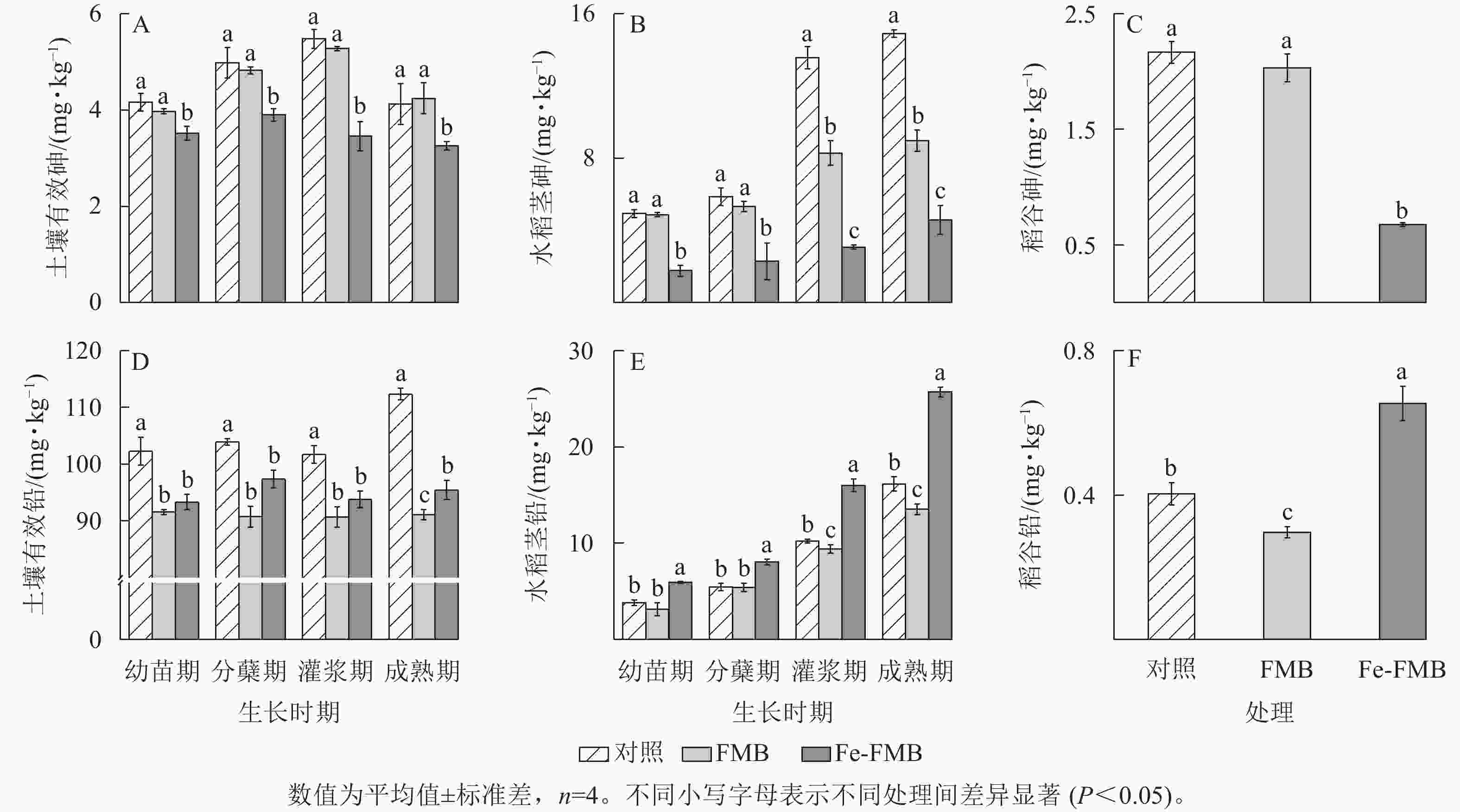
图 6 生物质炭对土壤中砷和铅有效性及在稻谷中富集的影响
Figure 6. Effect of biochar application on the availability of As and Pb in soil and their accumulation in rice straw
据图6D可知:与对照相比,施用2种生物质炭均显著降低了土壤中有效铅质量分数(P<0.05),其中FMB在成熟期对铅的固定效果最好,降幅达24%。使用FMB显著降低了水稻植株各部分的铅质量分数(图6E和图6F,P<0.05),Fe-FMB处理的水稻植株各部分的铅质量分数升高。
-
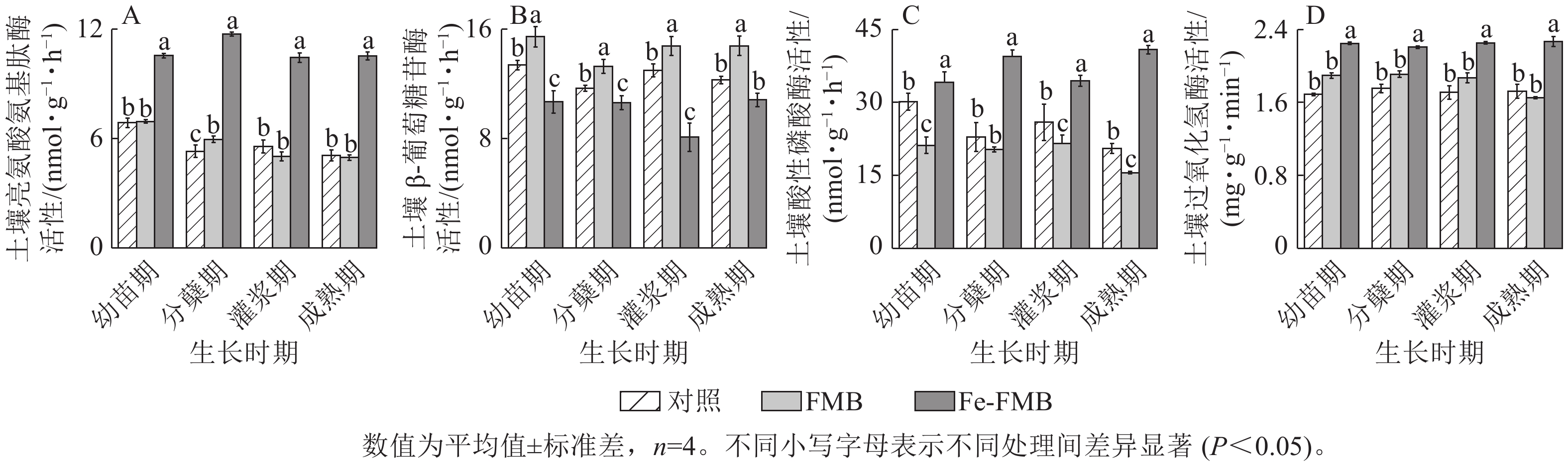
由图7可见:与对照相比,FMB显著提高了土壤中β-葡萄糖苷酶的活性(P<0.05),同时对酸性磷酸酶的活性也有一定抑制作用。Fe-FMB则在提升土壤中除β-葡萄糖苷酶外其他酶的活性方面表现出显著效果(P<0.05),与对照相比,最高增幅分别达121%、99%及33%。
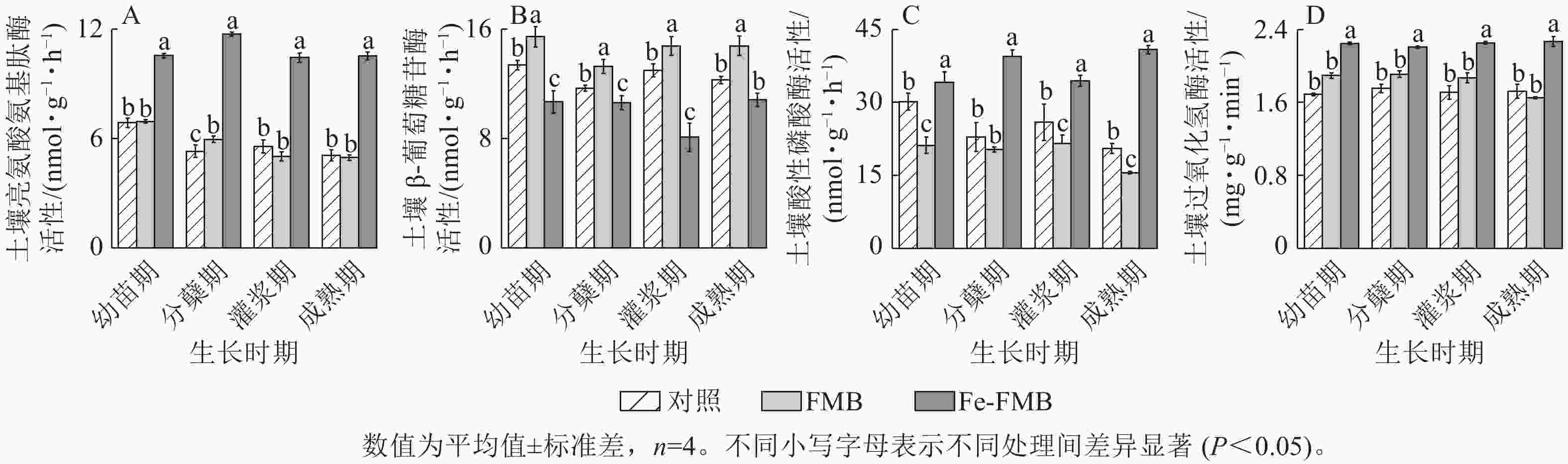
图 7 生物质炭对土壤酶活性的影响
Figure 7. Effect of biochars on soil enzyme activities
-
由表2可知:与对照相比,2种生物质炭均显著提高了水稻的生物量和稻谷产量(P<0.05),Fe-FMB处理的提升效果又显著高于FMB(P<0.05)。
表 2 生物质炭对水稻生物量(干质量)及稻谷产量的影响
Table 2. Impact of biochar on rice biomass (dry weight) and grain yield
处理组 不同生长时期水稻生物量(干质量)/g 稻谷产量/g 幼苗期 分蘖期 灌浆期 成熟期 对照 0.76±0.05 c 1.56±0.21 c 4.81±0.86 c 6.20±0.56 c 6.11±0.11 c FMB 1.68±0.15 b 2.93±0.29 b 6.54±0.42 b 7.54±0.22 b 8.42±0.47 b Fe-FMB 2.35±0.20 a 4.34±0.37 a 7.96±0.40 a 11.41±0.77 a 11.01±0.23 a 说明:不同小写字母表示同一时期内不同处理间差异显著(P<0.05)。 -
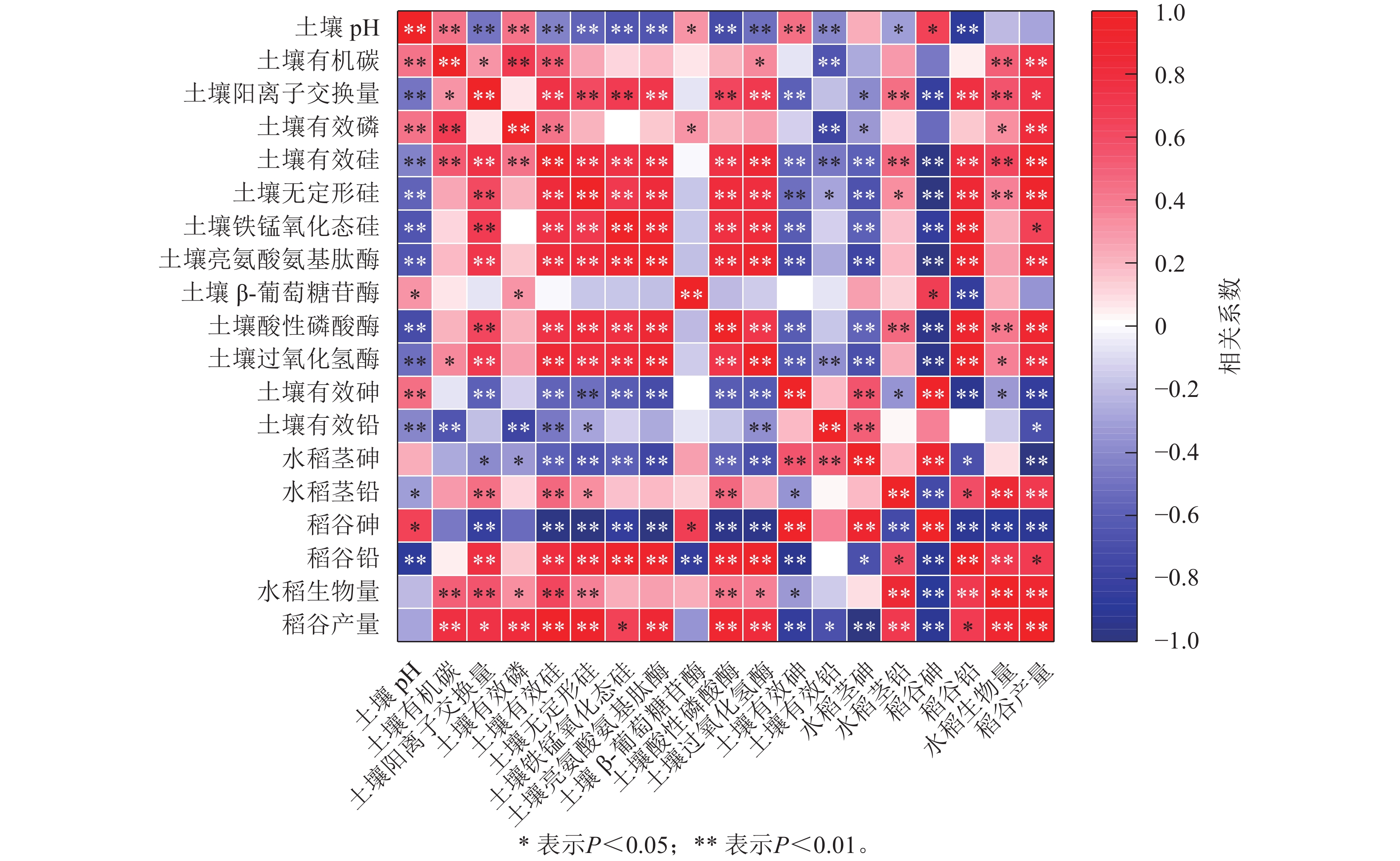
相关性分析(图8)表明:土壤有效砷的质量分数与土壤pH呈极显著正相关(P<0.01);而土壤中有效铅的质量分数则与土壤pH及有效磷质量分数呈极显著负相关(P<0.01);土壤pH与β-葡萄糖苷酶活性呈显著正相关(P<0.05);水稻植株生物量和稻谷产量与土壤有效硅及酶活性呈极显著正相关(P<0.01)。

图 8 不同指标的Pearson相关性分析
Figure 8. Pearson correlation coefficients among different parameters
-
本研究中,原始生物质炭呈碱性,施入土壤后可直接提高土壤的pH,生物质炭中的碳酸盐、硅酸盐和碳酸氢盐与土壤中的H+发生反应,也进一步提高了土壤pH[16]。土壤pH的提升降低了与磷的吸附解析相关的阳离子(Al3+、Ca2+)的活性,增加了土壤磷的有效性[17]。原始生物质炭较高的钾水平使其在提升土壤速效钾方面效果更好。生物质炭的施入调节了土壤的碳氮比,导致有效氮水平的降低[18]。此外,生物质炭中的酚类化合物促进了土壤的呼吸作用,进而加速了氮的流失[19]。
铁改性生物质炭由于其本身较低的pH,施入土壤后降低了土壤pH。铁改性生物质炭表面含有更多的含氧官能团,这些官能团在土壤溶液中吸附H+及其他阳离子,从而提高了土壤阳离子交换量[20]。另外铁改性生物质炭提高了土壤磷酸酶活性,促进了土壤有机残留物中磷的释放,提高了土壤有效磷水平。铁改性生物质炭较高的硅质量分数导致更多的硅溶出,提升了土壤无定型硅的水平[21]。无定型硅通过水解作用溶解于土壤中,为植物生长提供有效硅,间接提高了土壤有效硅水平[22]。胡祖武等[23]研究也表明:施用富含硅的竹叶生物质炭,可以显著增加水稻土壤有效硅和铁锰氧化态硅质量分数,从而显著提高水稻产量。
本研究中原始生物质炭能提高β-葡萄糖苷酶的活性,这可能是施用原始生物质炭后土壤pH上升的原因。YANG等[24]研究也发现:施用稻壳炭和猪炭后土壤酶活性提高,主要是由土壤pH升高导致。酸性磷酸酶和过氧化氢酶通常作为监测土壤砷污染的敏感指标。本研究中铁改性生物质炭能显著提升这2种酶的活性,表明铁改性生物质炭有效缓解了砷对酶的胁迫。但TANG等[25]研究发现:土壤中有效砷的增加可能增强酶活性点位与底物的协同性,提高过氧化氢酶的活性,与本研究结果不完全一致。这可能是因为铁改性生物质炭提高了土壤有机碳质量分数,增强了土壤好氧微生物的代谢能力,最终提高了过氧化氢酶的活性[26]。根据TRIPATHI等[27]的研究,硅可以提升水稻根际氧化能力,从而有效激活水稻根际过氧化氢酶活性,所以土壤有效硅质量分数显著上升也是过氧化氢酶活性增强的原因。
-
本研究使用聚合硫酸铁对细叶榕炭进行改性后发现,铁改性生物质炭施用后显著降低了土壤pH,促进了阴离子形态的砷通过静电吸附作用被固定,从而降低了砷的有效性[9]。此外,改性后生物质炭表面的官能团(—COOH和—OH)质子化增强,提升了砷与官能团间的络合作用[6]。铁改性生物质炭中的含铁化合物与${\mathrm{AsO}}_4^{3-} $和${\mathrm{HAsO}}_3^{2-} $等阴离子形成稳定的Fe-O-As沉淀,降低了土壤有效砷的质量分数[28]。本研究中原始生物质炭对土壤有效砷质量分数的影响不显著,可能是因为原始生物质炭较高的pH使其对阴离子形态的砷吸附效果较差。2种生物质炭均降低了土壤铅的有效性,主要原因为:首先,原始生物质炭提高了土壤pH,使土壤中的Pb(Ⅱ)与OH−反应产生沉淀,从而降低铅的迁移率[29];其次,2种生物质炭均显著提升了土壤中有效磷质量分数,可能促进Pb2+与${\mathrm{PO}}_4^{3-} $形成不溶性磷酸盐沉淀,如β-Pb9(PO4)6和Pb5(PO4)3[30]。而铁改性生物质炭降低了有效铅质量分数,这可能与土壤有效硅的升高有关,土壤中部分水溶性硅可以与铅产生难溶的沉淀,从而将铅转化为有效性较低的残渣态。
在水稻砷/铅积累方面,原始生物质炭降低了水稻中铅的富集,铁改性生物质炭减少了植株中砷的富集,但增加了稻谷中铅的质量分数。这可能是由于铁改性生物质炭施入土壤后,形成的难溶化合物有效降低了植物可吸收的砷[6]。水稻植株中的砷质量分数与土壤有效硅质量分数及无定形硅占比呈显著负相关,这可能是因为长期淹水环境下亚砷酸主要通过硅转运体被植物吸收,且土壤中的硅主要以水化无定型的SiO2·6H2O的形式存在,即As(Ⅲ)与土壤无定型硅共用吸收通道,因此无定型硅质量分数的升高可以通过竞争转运体的方式限制水稻对砷的吸收[31]。从根际吸收方面来看,铁改性生物质炭施入土壤后,根际逐渐形成铁膜,可能吸附土壤中的水溶性砷,限制水稻对砷的吸收[32]。原始生物质炭将土壤中可交换态铅转化为碳酸盐结合态和残渣态,从而减少了植物对铅的吸收。而在铁改性生物质炭施用后,水稻植株各部分的铅质量分数升高。根据相关研究[33],在高质量分数铅条件下,水稻对铁和铅的吸收可能存在协同作用,铁改性生物质炭引入的外源铁也可能是导致水稻植株中更高铅质量分数的原因。基于水稻对铅的吸收特性,推测土壤中硅质量分数的提升也可能导致水稻植株内铅质量分数有所提高。对水稻叶面喷施硅肥后,其茎叶中的铅质量分数显著提高,这与本研究结果一致。同样,陈新红等[34]研究也发现:铅在水稻各器官中的分布比例受到不同器官及处理的影响。
铁改性生物质炭显著提升了水稻的生物量与产量,原因是硅可以增加植株机械强度,提高水稻光合效率,从而促进水稻生长,增加稻谷产量。因此,水稻植株生物量及产量的增加可能是由于施用铁改性生物质炭后增加了土壤有效硅及无定型硅。同时,根据刘松涛等[35]的研究,土壤中的硅可以与砷竞争转运通道以及激活植物抗氧化系统,从而提高植物在重金属毒害下的产量。所以,铁改性生物质炭对砷和铅的吸附钝化作用以及对土壤中无定型硅质量分数的提高可能减少了重金属对植物生长的胁迫,这从土壤有效砷质量分数及稻谷中砷质量分数与水稻植株生物量及无定型硅均呈负相关的结果中可以看出。阮麟乔等[36]的研究也发现了相似的结果:在砷污染土壤中施用Fe3O4改性的生物质炭能够通过降低砷质量分数来促进水稻正常合成叶绿素,从而促进水稻的生长。
-
本研究表明:施用原始生物质炭提高了土壤pH和有效磷质量分数,而铁改性生物质炭则在增加土壤有机碳质量分数和提高土壤硅有效性方面表现更佳,且对水稻植株生物量及稻谷产量的提升作用尤为显著。2种生物质炭对于土壤酶活性的影响各异,这主要取决于土壤中的重金属有效性。在降低土壤中有效砷及减少水稻植株中砷富集方面,铁改性生物质炭表现更好,进而提高了水稻的生物量和产量。但铁改性生物质炭也增加了水稻茎叶及稻谷中铅质量分数,原始生物质炭则在降低土壤铅有效态质量分数及减少其在水稻植株中富集方面更为有效。
总体而言,原始生物质炭更适用于单一铅污染的土壤修复,而聚合硫酸铁改性生物质炭则更适用于修复砷铅复合污染的土壤。由于铁改性生物质炭提高了稻谷中铅的水平,在重金属砷铅复合污染土壤中对铅的转运和富集机制仍需进一步研究。
Effect of iron-modified biochars on soil nutrients and bioavailability of As and Pb
-
摘要:
目的 探讨原始及铁改性生物质炭对复合污染农田土壤中养分及砷(As)、铅(Pb)生物有效性的影响。 方法 分别采用质量分数为2%的原始细叶榕Ficus microcarpa生物质炭(FMB)和聚合硫酸铁改性细叶榕生物质炭(Fe-FMB)与受砷铅复合污染土壤混合,以未施加生物质炭的土壤作为对照,进行水稻Oryza sativa盆栽试验,测定不同生长阶段土壤养分、土壤酶活性以及水稻各器官和土壤中As和Pb的质量分数。 结果 与对照组相比,Fe-FMB显著提高了土壤中磷(P)、硅(Si)等养分的有效性(P<0.05),显著改变了土壤中硅(Si)的形态分布,主要增加了无定型硅(增加25.2%)和铁锰氧化态硅质量分数(增加11.1%)。Fe-FMB对于土壤和水稻稻谷中As的钝化效果更为显著(P<0.05);而FMB在钝化土壤Pb方面表现更佳,对Pb有效性最高降低了24.9%。此外,Fe-FMB还显著提升了土壤中亮氨酸氨基肽酶、酸性磷酸酶及过氧化氢酶的活性(P<0.05),增幅分别达121.1%、99.1%及33.2%。Pearson’s相关性分析结果表明,土壤酶活性与pH及土壤As有效性显著相关(P<0.05),说明施用生物质炭可通过调节土壤pH及As有效性来影响土壤酶活性。 结论 原始细叶榕生物质炭适用于修复单一Pb污染土壤。相比之下,铁改性生物质炭在修复砷铅复合污染土壤方面展现出更好的应用前景。图8表2参36 Abstract:Objective To investigate the effects of raw and iron-modified biochar on the nutrient content and bioavailability of arsenic (As) and lead (Pb) in con-contaminated agricultural soil. Method An experiment using rice potted in soil mixed with 2% raw Ficus microcarpa biochar (FMB) and Polyferric Sulfate (iron)-modified biochar (Fe-FMB) was conducted, no biochar soil as control. Soil nutrient availability, soil enzyme activity, rice biomass and As and Pb concentrations in various plant organs at different growth stages were measured. The bioavailable As and Pb in the soil were determined using the NH4H2PO4 and DTPA extraction methods, respectively. Result The results indicated that, compared to the control, Fe-FMB significantly enhanced the availability of nutrients such as phosphorus (P) and silicon (Si) in the soil and significantly altered the distribution of Si forms in the soil (P<0.05), primarily increasing the content of amorphous silicon (by 25.2%) and iron-manganese oxidized silicon (by 11.1%). Fe-FMB was more effective in immobilizing soil As (P<0.05), reducing it by 37.9% compared to the control, while original biochar (FMB) was more effective for soil Pb immobilization, reducing it by 24.9%. Application of Fe-FMB led to a 67.2% reduction in As content in rice grains as compared to the control. Furthermore, Fe-FMB significantly increased the activities of leucine aminopeptidase, acid phosphatase, and catalase (P<0.05), with maximum increases of 121.1%, 99.1%, and 33.2%, respectively. Pearson correlation analysis showed that soil enzyme activity was significantly related to pH and As availability (P<0.05), indicating that biochar application can regulate soil enzyme activity by influencing soil pH and As bioavailability. Conclusion While F. microcarpa biochar is effective in remediating soils contaminated with Pb only, it is not suitable for the treatment of soils co-contaminated with As and Pb. On the other hand, iron-modified biochar shows a better prospect for remediating soils co-contaminated with As and Pb. [Ch, 8 fig. 2 tab. 36 ref.] -
Key words:
- biochar /
- heavy metal /
- soil remediation /
- silicon /
- soil enzyme
-
图 1 供试生物质炭的形貌和结构
Figure 1 Morphology and structure of biochar before and after modification
图 2 供试生物质炭的X射线衍射分析图谱(A)和傅里叶红外光谱图(B)
Figure 2 X-ray diffraction (A), and Fourier transform infrared spectra (B) of biochars
图 3 施入生物质炭对土壤理化性质的影响
Figure 3 Effect of biochar application on soil physiochemitrical properties
图 4 施入生物质炭对土壤养分的影响
Figure 4 Effect of biochar application on soil N, P and K availability
图 5 施用生物质炭对土壤有效硅以及不同形态硅分布的影响
Figure 5 Effect of biochar application on soil Si availability and the fractions of Si in soil
图 6 生物质炭对土壤中砷和铅有效性及在稻谷中富集的影响
Figure 6 Effect of biochar application on the availability of As and Pb in soil and their accumulation in rice straw
图 8 不同指标的Pearson相关性分析
Figure 8 Pearson correlation coefficients among different parameters
表 1 生物质炭的基本理化性质
Table 1. Selected physicochemical properties of the used biochars
生物质炭 pH 灰分/(g·kg−1) 碳/(g·kg−1) 氢/(g·kg−1) 氮/(g·kg−1) 硅/(g·kg−1) 铁/(g·kg−1) 比表面积/(m2·kg−1) 阳离子交换量/(cmol·kg−1) FMB 9.5 79.3 753 5.0 27.5 1.50 7.89 23.4 14.45 Fe-FMB 5.3 137.8 505 7.5 13.5 7.31 49.32 132.6 30.95  下载: 导出CSV
下载: 导出CSV
表 2 生物质炭对水稻生物量(干质量)及稻谷产量的影响
Table 2. Impact of biochar on rice biomass (dry weight) and grain yield
处理组 不同生长时期水稻生物量(干质量)/g 稻谷产量/g 幼苗期 分蘖期 灌浆期 成熟期 对照 0.76±0.05 c 1.56±0.21 c 4.81±0.86 c 6.20±0.56 c 6.11±0.11 c FMB 1.68±0.15 b 2.93±0.29 b 6.54±0.42 b 7.54±0.22 b 8.42±0.47 b Fe-FMB 2.35±0.20 a 4.34±0.37 a 7.96±0.40 a 11.41±0.77 a 11.01±0.23 a 说明:不同小写字母表示同一时期内不同处理间差异显著(P<0.05)。
下载: 导出CSV
-
[1] 余涛, 蒋天宇, 刘旭, 等. 土壤重金属污染现状及检测分析技术研究进展 [J]. 中国地质, 2021, 48(2): 460 − 476. YU Tao, JIANG Tianyu, LIU Xu, et al. Research progress in current status of soil heavy metal pollution and analysis technology [J]. Geology in China, 2021, 48(2): 460 − 476. [2] LIU Lin, HAN Jialiang, XU Zhidong, et al. Dietary exposure assessment of cadmium, arsenic, and lead in market rice from Sri Lanka [J]. Environmental Science and Pollution Research, 2020, 27(34): 42704 − 42712. [3] 吴萍萍, 李录久, 李敏. 生物炭负载铁前后对复合污染土壤中Cd、Cu、As淋失和形态转化的影响研究 [J]. 环境科学学报, 2017, 37 (10): 3959 − 3967. WU Pingping, LI Lujiu, LI Min. Effects of biochar and Fe-loaded biochar on the leaching and fraction transformation of Cd, Cu and As in multi-contaminated soil [J]. Acta Scientiae Circumstantiae, 2017, 37 (10): 3959 − 3967. [4] YANG Xing, HINZMANN M, PAN He, et al. Pig carcass-derived biochar caused contradictory effects on arsenic mobilization in a contaminated paddy soil under fluctuating controlled redox conditions [J/OL]. Journal of Hazardous Materials, 2022, 421 : 126647[2024-01-30]. doi: 10.1016/j.jhazmat.2021.126647. [5] 宋佩佩, 马文静, 王军, 等. 铁改性生物炭的制备及其在重金属污染土壤修复技术中的应用进展 [J]. 环境工程学报, 2022, 16 (12): 4018 − 4036. SONG Peipei, MA Wenjing, WANG Jun, et al. Preparation of iron-modified biochar and its application in heavy metal contaminated soils [J]. Chinese Journal of Environmental Engineering, 2022, 16 (12): 4019 − 4036. [6] WEN Er, YANG Xing, CHEN Hanbo, et al. Iron-modified biochar and water management regime-induced changes in plant growth, enzyme activities, and phytoavailability of arsenic, cadmium and lead in a paddy soil [J/OL]. Journal of Hazardous Materials, 2021, 407 : 124344[2024-01-30]. doi:/10.1016/j.jhazmat.2020.124344. [7] 秦怀婷, 曾小林, 刘程琳. 聚合硫酸铁的制备、改性及应用研究进展 [J]. 工业水处理, 2023, 43(7): 53 − 61. QIN Huaiting, ZENG Xiaolin, LIU Chenglin. Research progress on preparation, modification, and application of polyferric sulfate [J]. Industrial Water Treatment, 2023, 43(7): 53 − 61. [8] 谢登科. 聚合硫酸铁和活化硅酸复配除浊效果对比 [J]. 资源节约与环保, 2019(7): 71 − 72 XIE Dengke. The comparative effectiveness of composite coagulants containing polymeric ferric sulfate and activated silica sol for turbidity removal [J]. Resource Conservation and Environmental Protection, 2019(7): 71 − 72 [9] 戴志楠, 杨兴, 陈翰博, 等. 原始及铁改性生物质炭对污染土壤中As、Pb生物有效性和微生物群落结构的影响 [J]. 环境科学学报, 2022, 42(7): 456 − 465. DAI Zhinan, YANG Xing, CHEN Hanbo, et al. Effect of raw and iron-modified biochars on the bioavailability of As and Pb and functional diversity of the microbial community in soils [J]. Acta Scientiae Circumstantiae, 2022, 42(7): 456 − 465. [10] 鲁如坤. 土壤农业化学分析方法[M]. 北京: 中国农业科技出版社, 2000. LU Rukun. Methods for Agrochemical Analysis of Soil [M]. Beijing: China Agricultural Science and Technology Press, 2000. [11] 包建平, 袁根生, 董方圆, 等. 生物质炭与秸秆施用对红壤有机碳组分和微生物活性的影响 [J]. 土壤学报, 2020, 57(3): 721 − 729. BAO Jianping, YUAN Gensheng, DONG Fangyuan, et al. Effects of biochar application and straw returning on organic carbon fractionations and microbial activities in a red soil [J]. Acta Pedologica Sinica, 2020, 57(3): 721 − 729 [12] TAN Guangcai, MAO Yi, WANG Hongyuan, et al. A comparative study of arsenic(Ⅴ), tetracycline and nitrate ions adsorption onto magnetic biochars and activated carbon [J]. Chemical Engineering Research and Design, 2020, 159: 582 − 591. [13] 顾绍茹, 杨兴, 陈翰博, 等. 小龙虾壳炭和细叶榕枝条炭对土壤养分及镉和铅生物有效性的影响 [J]. 浙江农林大学学报, 2023, 40(1): 176 − 187. GU Shaoru, YANG Xing, CHEN Hanbo, et al. Effects of biochar from Procambarus clarkia shells and Ficus microcarpa branches on soil nutrients and bioavailability of Cd and Pb [J]. Journal of Zhejiang A&F University, 2023, 40(1): 176 − 187. [14] ZHANG Jingyi, ZHOU Hang, GU Jiacun, et al. Effects of nano-Fe3O4-modified biochar on iron plaque formation and Cd accumulation in rice (Oryza sativa L. ) [J/OL]. Environmental Pollution, 2020, 260 : 113970[2024-01-30]. doi: 10.1016/j.envpol.2020.113970. [15] HAO Bian, JIANG Wan, MUHAMMAD T, et al. Computational study and optimization experiment of nZVI modified by anionic and cationic polymer for Cr(Ⅵ) stabilization in soil: kinetics and response surface methodology (RSM) [J/OL]. Environmental Pollution, 2021, 276 : 116745[2024-01-30]. doi:10.1016/j.envpol.2021.116745. [16] GUL S, WHALEN J K, THOMAS B W, et al. Physico-chemical properties and microbial responses in biochar-amended soils: mechanisms and future directions [J]. Agriculture, Ecosystems & Environment, 2015, 206 : 46 − 59. [17] 占亚楠, 王智, 孟亚利. 生物炭提高土壤磷素有效性的整合分析 [J]. 应用生态学报, 2020, 31(4): 1185 − 1193. ZHAN Yanan, WANG Zhi, MENG Yali. Biochar addition improve soil phosphorus availability: a meta-analysis [J]. Chinese Journal of Applied Ecology, 2020, 31(4): 1185 − 1193. [18] 何绪生, 张树清, 佘雕, 等. 生物炭对土壤肥料的作用及未来研究 [J]. 中国农学通报, 2011, 27(15): 16 − 25. HE Xusheng, ZHANG Shuqing, SHE Diao, et al. Effects of biochar on soil and fertilizer and future research [J]. Chinese Agricultural Science Bulletin, 2011, 27(15): 16 − 25. [19] DEENIK J L, MCCLELLAN A T, UEHARA G. Biochar volatile matter content effects on plant growth and nitrogen transformations in a tropical soil [C]//[s.l.] Western Nutrient Management Conference, Salt Lake City, 2009, 8 : 26 − 31. [20] WANG Shuai, LI Bo, ZHU Hanhua, et al. Long-term effects of biochar on trace metals accumulation in rice grain: a 7-year field experiment [J/OL]. Agriculture, Ecosystems & Environment, 2021, 315: 107446[2024-01-30]. doi: 10.1016/j.agee.2021.107446. [21] WANG Yaofeng, XIAO Xin, Chen Baoliang. Biochar impacts on soil silicon dissolution kinetics and their interaction mechanisms [J/OL]. Scientific Reports, 2018, 8 (1): 8040[2024-01-30]. doi:10.1038/s41598-018-26396-3. [22] SCHALLER J, PUPPE D, BUSSE J, et al. Silicification patterns in wheat leaves related to ontogeny and soil silicon availability under field conditions [J]. Plant and Soil, 2022, 477(1/2): 9 − 23. [23] 胡祖武, 吴多基, 吴建富, 等. 富硅生物炭有效提高红壤性稻田土壤不同形态硅含量及水稻产量 [J]. 植物营养与肥料学报, 2022, 28(8): 1421 − 1429. HU Zuwu, WU Duoji, WU Jianfu, et al. Silicon-rich biochar effectively increases the availability of soil silicon and rice yield in reddish paddy soil [J]. Journa of Plant Nutrition and Fertilizers, 2022, 28(8): 1421 − 1429. [24] YANG Xing, WEN Er, GE Chengjun, et al. Iron-modified phosphorus- and silicon-based biochars exhibited various influences on arsenic, cadmium, and lead accumulation in rice and enzyme activities in a paddy soil [J/OL]. Journal of Hazardous Materials, 2023, 443 : 130203[2024-01-30]. doi:10.1016/j.jhazmat.2022.130203. [25] TANG Jiayi, ZHANG Lihua, ZHANG Jiachao, et al. Physicochemical features, metal availability and enzyme activity in heavy metal-polluted soil remediated by biochar and compost [J/OL]. Science of the Total Environment, 2020, 701 : 134751[2024-01-30]. doi: 10.1016/j.scitotenv.2019.134751. [26] 王文慧, 蒋志慧, 张纪, 等. 生物炭对大豆根际土壤酶活性及产量的影响 [J]. 中国土壤与肥料, 2023(6): 147 − 153. WANG Wenhui, JIANG Zhihui, ZHANG Ji, et al. The effect of biochar on soybean rhizosphere soil enzyme activity and yield [J]. Chinese Journal of Soil and Fertilizer, 2023(6): 147 − 153. [27] TRIPATHI P, TRIPATHI R D, SINGH R P, et al. Silicon mediates arsenic tolerance in rice (Oryza sativa L. ) through lowering of arsenic uptake and improved antioxidant defence system [J]. Ecological Engineering, 2013, 52: 96 − 103. [28] XUE Qin, RAN Ying, TAN Yunzhi, et al. Arsenite and arsenate binding to ferrihydrite organo-mineral coprecipitate: Implications for arsenic mobility and fate in natural environments [J]. Chemosphere, 2019, 224: 103 − 110. [29] LI Jianhong, WANG Shanli, ZHANG Jing, et al. Coconut-fiber biochar reduced the bioavailability of lead but increased its translocation rate in rice plants: elucidation of immobilization mechanisms and significance of iron plaque barrier on roots using spectroscopic techniques [J/OL]. Journal of Hazardous Materials, 2020, 389: 122117[2024-01-30]. doi: 10.1016/j.jhazmat.2020.122117. [30] PENG Dinghua, WU Bin, TAN Hang, et al. Effect of multiple iron-based nanoparticles on availability of lead and iron, and micro-ecology in lead contaminated soil [J]. Chemosphere, 2019, 228: 44 − 53. [31] SEYFFERTH A L, AMARAL D, LIMMER M A, et al. Combined impacts of Si-rich rice residues and flooding extent on grain As and Cd in rice [J]. Environment International, 2019, 128: 301 − 309. [32] YU Zhihong, QIU Weiwen, WANG Fei, et al. Effects of manganese oxide-modified biochar composites on arsenic speciation and accumulation in an indica rice (Oryza sativa L. ) cultivar [J]. Chemosphere, 2017, 168: 341 − 349. [33] 黄冬芬, 王志琴, 刘立军. 铅胁迫下不同品种水稻对铅与微量元素的吸收分配差异 [J]. 热带作物学报, 2009, 30(12): 25 − 28. HUANG Dongfen, WANG Zhiqing, LIU Lijun. Differences in absorption and distribution of Pb and trace elements in different rice varieties under lead stress [J]. Chinese Journal of Tropical Crops, 2009, 30(12): 25 − 28. [34] 陈新红, 叶玉秀, 潘国庆, 等. 杂交水稻不同器官重金属铅浓度与累积量 [J]. 中国水稻科学, 2014, 28(1): 57 − 64. CHEN Xinhong, YE Yuxiu, PAN Guoqing, et al. Concentration and accumulation of lead in different organs of hybrid rice [J]. Chinese Rice Science, 2014, 28(1): 57 − 64. [35] 刘松涛, 李茜, 王小玲. 硅素营养对水稻抗重金属毒害的研究进展 [J]. 湖北农业科学, 2017, 56(3): 405 − 408, 417. LIU Songtao, LI Qian, WANG Xiaoling. Progress in research of the resistance to heavy metal toxicity in rice by silicon nutrient [J]. Hubei Agricultural Sciences, 2017, 56(3): 405 − 408, 417. [36] 阮麟乔, 梁美娜, 丁艳梅, 等. 施加Fe3O4/桑树杆生物炭对土壤砷形态和水稻砷含量的影响 [J]. 环境科学, 2023, 44(8): 4468 − 4478. RUAN Lingqiao, LIANG Meina, DING Yanmei, et al. Application of Fe3O4/mulberry stem biochar effects on soil arsenic species and rice arsenic content [J]. Environmental Science, 2023, 44(8): 4468 − 4478. -

-
链接本文:
https://zlxb.zafu.edu.cn/article/doi/10.11833/j.issn.2095-0756.20240171
 点击查看大图
点击查看大图
计量
- 文章访问数: 1759
- HTML全文浏览量: 656
- PDF下载量: 33
- 被引次数: 0
