




-
土壤重金属污染作为工业化进程的严峻后果,对全球生态系统安全和农业可持续发展构成了持久性威胁[1]。在众多环境修复技术中,基于超积累植物的植物修复技术,因具有成本低、无二次污染以及可原地实施等优点,被公认为一种极具潜力的绿色可持续解决方案[2]。在这一领域,发现于中国的重金属超积累植物——伴矿景天Sedum plumbizincicola,展现出对锌(Zn)和镉(Cd)非凡的富集能力,已成为研究植物重金属耐受与超积累分子机制的珍贵模式材料[3−4]。理解其内在的分子应答机制,是推动该物种在植物修复中实现高效应用,并通过基因工程手段改良其他作物重金属耐受性的理论基础。
植物在应对重金属胁迫时,进化出了一套精密的体内稳态调控网络。其中,将细胞质中过量的重金属离子转运至液泡等细胞器中进行区隔化,是解毒过程的核心环节[5]。而金属耐受蛋白(MTP)作为阳离子扩散辅助蛋白(CDF)家族在植物中的重要代表,在此过程中扮演着关键的运输者角色。其主要定位于液泡膜或细胞质膜,负责将锌、锰、铁等二价阳离子从细胞质中泵出,从而有效维持胞质内金属离子的动态平衡,避免金属毒性。自首个MTP成员在模式植物拟南芥Arabidopsis thaliana中被功能鉴定以来[6],该基因家族在多种植物中的系统发生与功能研究已广泛展开。例如,拟南芥中的AtMTP1和AtMTP3被证实参与锌的液泡区隔化[7−8],而AtMTP8则与锰的耐受密切相关[9]。这些研究表明:MTP家族成员在功能上存在显著的分化,各司其职又相互协作,共同构成了植物应对金属胁迫的复杂防线。近年来,越来越多研究证实MTP基因在植物镉胁迫应答中扮演了关键角色。如水稻Oryza sativa OsMTP11定位于叶部维管细胞,通过TGN-PVC-液泡转运途径将镉螯合于液泡,阻断其向籽粒转运,且启动子区G-box-like基序的拷贝数变异可通过调控转录因子OsIRO2的结合效率影响基因表达强度[10];向日葵 Helianthus annuus HaMTP10属于Mn-CDF亚家族,作为镉反向转运体,在辣木籽提取物(MSE)诱导下高表达,通过增强光合作用和调控镉的细胞内分配,显著降低向日葵镉积累并提升产量[11];红豆杉 Taxus media TmMTP1 (Zn-CDF亚家族)和TmMTP11 (Mn-CDF亚家族)则通过细胞膜定位介导镉的转运与液泡区隔化,其表达受TmMYB16 (抑制)和TmMYB123 (激活)的协同调控,进而增强植株镉耐受性[12]。这些研究表明:MTP家族成员可通过液泡区隔化、组织间转运调控、离子平衡协同维持等多种机制参与镉胁迫响应,但其在超积累植物中针对镉的特异性转运机制仍需深入解析。
然而,当前对MTP家族的认识大多源于拟南芥、水稻等非超积累植物。在这些植物中,MTP蛋白的功能主要是维持微量必需元素的稳态和抵御毒性压力。与之形成鲜明对比的是,在伴矿景天这样的超积累植物中,其生理目标发生了根本性转变——它们需要主动且高效地吸收、转运并储存远超正常生理需求的重金属离子[13]。这一独特的生物学特性预示着:其MTP基因家族可能经历了不同于非超积累植物的进化历程,从而在基因拷贝数、功能分化以及表达调控上形成了独特的适应性特征。但是,伴矿景天中SpMTP基因家族的全基因组系统性分析仍缺乏。
本研究对伴矿景天基因组中的SpMTP基因家族进行了系统的生物信息学与表达谱分析。通过全基因组鉴定,明确其成员组成与基本特性;通过构建包含多个代表性物种的系统进化树,厘清其进化地位与亚族分类;通过分析保守基序、基因结构及共线性关系,揭示其结构保守性与基因组进化机制;最终,通过探究镉胁迫下SpMTP基因在根、茎、叶中的特异性表达模式,筛选出在镉超积累过程中可能发挥关键作用的候选基因。本研究不仅能够填补超积累植物中MTP基因家族系统性研究的空白,深化对植物重金属耐受分子机制的理解,更为未来利用分子育种手段定向改良植物修复效率,或通过基因工程培育重金属安全的作物品种提供了重要的理论依据与基因资源。
-
研究所用的伴矿景天植株在可控温室中水培培养。待无性繁殖的幼苗生根后,采用添加400 μmol·L−1 镉的霍格兰营养液进行胁迫处理,以不含额外重金属的营养液为对照(ck)。处理4 d后,分别收取根、茎、叶组织,立即于液氮中速冻,并于−80℃超低温冰箱保存,用于后续RNA提取。每处理设置3个生物学重复。
-
从Pfam数据库下载CDF家族隐马尔可夫模型(HMM)文件(PF01545)[14]。利用HMMER 3.3.2的hmmsearch程序搜索伴矿景天基因组蛋白序列(来源于转录组数据),期望值(E)的阈值设置为1×10−5。所有候选序列通过SMART和美国国家生物技术信息中心(NCBI)的CDD数据库进行保守结构域验证,仅保留含有完整CDF结构域的序列用于后续分析。利用ExPASy ProtParam工具预测蛋白质的分子量(MW)[15]、等电点(pI)和总平均亲水性(GRAVY)。使用TMHMM Server v.2.0预测跨膜结构域(TMD)[16]。
-
从Phytozome或NCBI数据库下载拟南芥、水稻等6个物种的MTP蛋白序列。使用MEGA 11.0软件,采用邻接法(NJ)构建系统进化树[17],bootstrap重复次数设为1 000。利用MEME Suite 5.4.1在线工具进行保守基序分析[18],最大基序数设为10,基序宽度范围为6~50个氨基酸。基于基因组注释文件(GFF3格式),利用TBtools软件的Visualize Gene Structure功能绘制基因结构图。
-
使用MCScanX软件分析伴矿景天自身基因组内以及与拟南芥、景天科Crassulaceae近缘物种费氏伽蓝菜Kalanchoe fedtschenkoi基因组间的共线性关系[19]。使用BLASTP进行全基因组蛋白序列比对,将比对结果导入MCScanX软件进行共线性区块检测。最终结果利用TBtools中的Advanced Circos功能进行可视化。
-
使用PlantCARE在线工具分析SpMTP基因上游2 000 bp的基因组序列中的顺式作用元件。各元件的数量及位置利用TBtools进行可视化。
-
SpMTP基因的每千碱基转录本每百万映射片段数(FPKM)数据源自中国林业科学研究院亚热带林业研究所发表的转录组测序结果[20]。表达值经Z-score标准化后归一化处理,利用TBtools绘制表达热图。使用TRIzol法提取各组织总RNA,选取4个代表性SpMTP基因,采用实时荧光定量PCR (RT-qPCR)验证转录组数据可靠性。以Actin为内参基因,采用 2−ΔΔCt法计算相对表达量。
-
对伴矿景天基因组进行系统性搜索与保守结构域验证,共鉴定出14个非冗余的SpMTP基因,其编码蛋白的基本理化性质如表1所示。14个SpMTP基因的开放阅读框(CDS)长度介于906 bp (SpMTP5.2)和2 499 bp (SpMTP12),编码氨基酸数目为301~832个,其中SpMTP12的氨基酸数目最多(832个),是SpMTP5.2 (301个)的2.77倍;分子量(MW)范围为33.80 kDa (SpMTP5.2)至92.58 kDa (SpMTP12),与氨基酸数目呈正相关。等电点(pI)分布在4.9 (SpMTP11、SpMTP8.1)至8.9 (SpMTP5.2),其中10个基因的pI<7 (酸性蛋白),4个基因的pI>7 (碱性蛋白),表明不同成员可能在细胞内不同pH微环境中发挥功能。亲水性分析(GRAVY)结果显示:除SpMTP7 (−0.028)、SpMTP3 (−0.021)、SpMTP9 (−0.124)、SpMTP6 (−0.001)、SpMTP10 (−0.129)外,其余9个成员的GRAVY均为正值(0.017~0.136),表明大多数SpMTP蛋白为疏水性蛋白,符合膜转运蛋白的结构特征。跨膜结构域(TMD)预测显示:13个成员含有5~6个TMD,仅SpMTP10含有4个TMD,而SpMTP12含有16个TMD,推测其可能具有更复杂的跨膜转运功能。亚细胞定位预测显示:10个成员定位于液泡膜,4个成员(SpMTP5.2、SpMTP9、SpMTP12、SpMTP10)定位于细胞膜/液泡膜,为其参与金属离子跨膜转运提供了结构基础[21]。
表 1 伴矿景天SpMTP基因家族成员基本信息
Table 1. Information of SpMTP gene family in S. plumbizincicola genome
基因名称 基因编号 基因长
度/bp编码区
长度/bp分子量/Da 氨基
酸数不稳定
系数脂肪族
指数总平均
亲水性等电点 跨膜结构
域数量亚细胞定位 SpMTP5.2 evm.model.000001F.133 4 751 906 33 804.93 301 39.95 90.70 0.099 8.9 5 细胞膜/液泡膜 SpMTP11 evm.model.000001F.377 3 009 1 185 44 577.15 394 42.06 105.43 0.069 4.9 6 液泡膜 SpMTP8.2 evm.model.000002F.140 2 722 1 266 46 436.58 421 44.01 107.72 0.120 5.1 6 液泡膜 SpMTP7 evm.model.000002F.531 4 104 1 374 50 592.04 457 39.21 94.09 −0.028 7.8 5 液泡膜 SpMTP3 evm.model.000009F.229 2 814 1 206 44 609.16 401 31.38 106.76 −0.021 5.9 6 液泡膜 SpMTP9 evm.model.000009F.527 3 030 1 191 45 410.30 396 46.02 94.55 −0.124 6.6 6 细胞膜/液泡膜 SpMTP6 evm.model.000010F.138 3 954 1 506 54 669.90 501 42.13 98.30 −0.001 6.8 5 液泡膜 SpMTP12 evm.model.000014F.173 2 499 2 499 92 580.50 832 42.38 98.49 0.072 8.4 16 细胞膜/液泡膜 SpMTP2 evm.model.000015F.32 1 479 1 227 45 123.59 408 40.28 107.03 0.017 6.1 6 液泡膜 SpMTP8.1 evm.model.000078F.209 2 229 1 266 46 436.37 421 45.52 108.43 0.136 4.9 6 液泡膜 SpMTP1 evm.model.000106F.135 1 992 1 176 43 433.16 391 39.09 109.97 0.131 5.9 6 液泡膜 SpMTP10 evm.model.000127F.82 6 121 1 065 40 650.67 354 46.93 91.41 −0.129 6.2 4 细胞膜/液泡膜 SpMTP5.1 evm.model.000138F.18 5 668 1 179 43 832.34 392 48.38 92.73 0.084 7.2 6 液泡膜 SpMTP4 evm.model.000148F.36 2 170 1 197 43 582.79 398 35.91 103.79 0.057 6.0 6 液泡膜 -
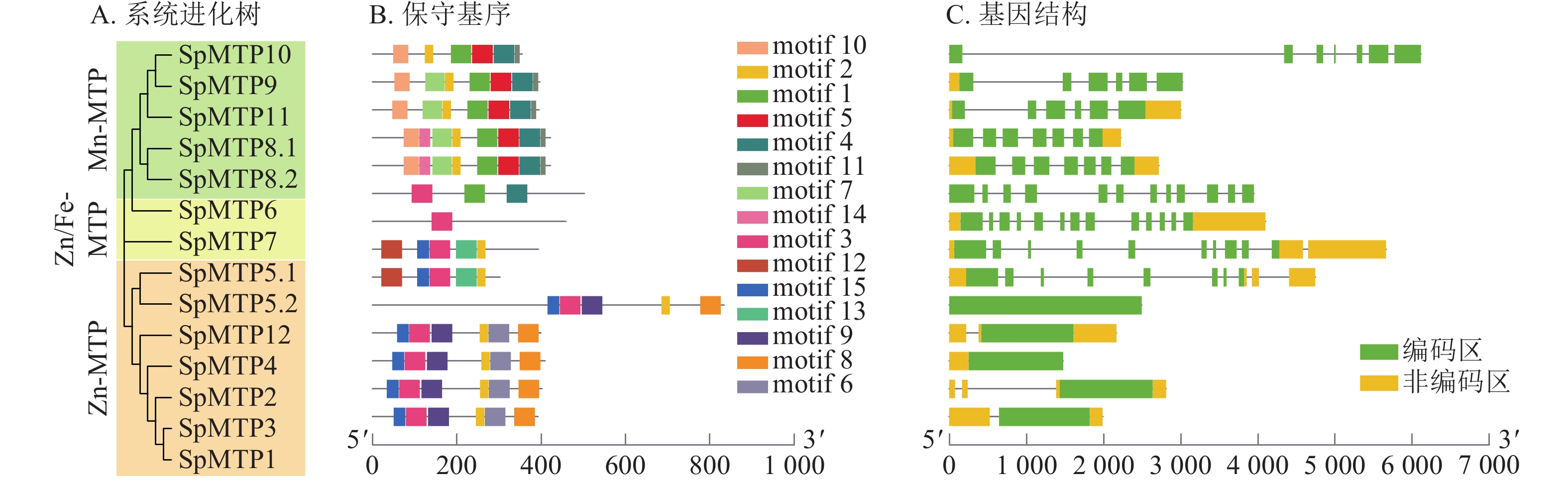
为了探究SpMTP基因的进化关系,将伴矿景天的14个SpMTP蛋白与来自东南景天Sedum alfredii、拟南芥、水稻等6个代表性物种的MTP蛋白一同构建了系统进化树(图1)。系统进化树显示:89个MTP蛋白(含伴矿景天的14个SpMTP)被明确划分为7个亚族。其中,SpMTP1、SpMTP2、SpMTP3、SpMTP4 归属于亚族1,与东南景天SaMTP1、SaMTP2、SaMTP4 及拟南芥AtMTP1、AtMTP2聚集; SpMTP6 属于亚族6,与东南景天SaMTP6及水稻OsMTP6亲缘关系较近;SpMTP5.1、SpMTP5.2归属于亚族5;SpMTP7单独聚类于亚族7;SpMTP8.1、SpMTP8.2属于亚族8;SpMTP9、SpMTP10、SpMTP11归属于亚族11;SpMTP12单独聚类于亚族12。14个SpMTP成员在7个亚族中均有分布,且多数SpMTP成员与东南景天的同源蛋白形成直系同源对,表明二者MTP家族进化高度保守[22]。

图 1 伴矿景天与其他6种植物MTP蛋白的系统进化分析
Figure 1. Phylogenetic analysis of MTP proteins from S. plumbizincicola and other 6 plant species
为进一步揭示其序列与结构特征,对所有SpMTP蛋白进行了保守基序分析(图2B)。结果显示:属于同一进化亚族的成员通常共享相似的保守基序组合,而不同亚族间的基序组成则存在明显差异,这为亚族的功能分化提供了结构线索[23]。所有SpMTP蛋白均含有完整的CDF保守结构域(PF01545),这是金属离子结合与转运的关键功能区域。MEME分析鉴定出15个保守motif (motif 1~15),其中motif 5和10仅存在于Mn-MTP成员中,可能与锰离子特异性结合相关;motif 15为亚族特异性motif,推测与各亚族功能分化相关。同一亚族成员的motif组成顺序和数量高度一致,不同亚族间motif差异显著,表明motif组成与进化分类及功能分化密切相关。与此同时,基因结构分析(图2C)表明:不同亚族的SpMTP基因在外显子-内含子结构上呈现出高度的多样性,其内含子数量为2~12个。系统进化关系相近的基因通常具有相似的基因结构,例如亚族8中的成员大多内含子数量较多且结构复杂,而亚族1中的成员则结构相对简单。这种系统进化、保守基序与基因结构间的高度相关性,共同印证了SpMTP基因家族内部的进化保守性,并为后续功能推测提供了重要依据。
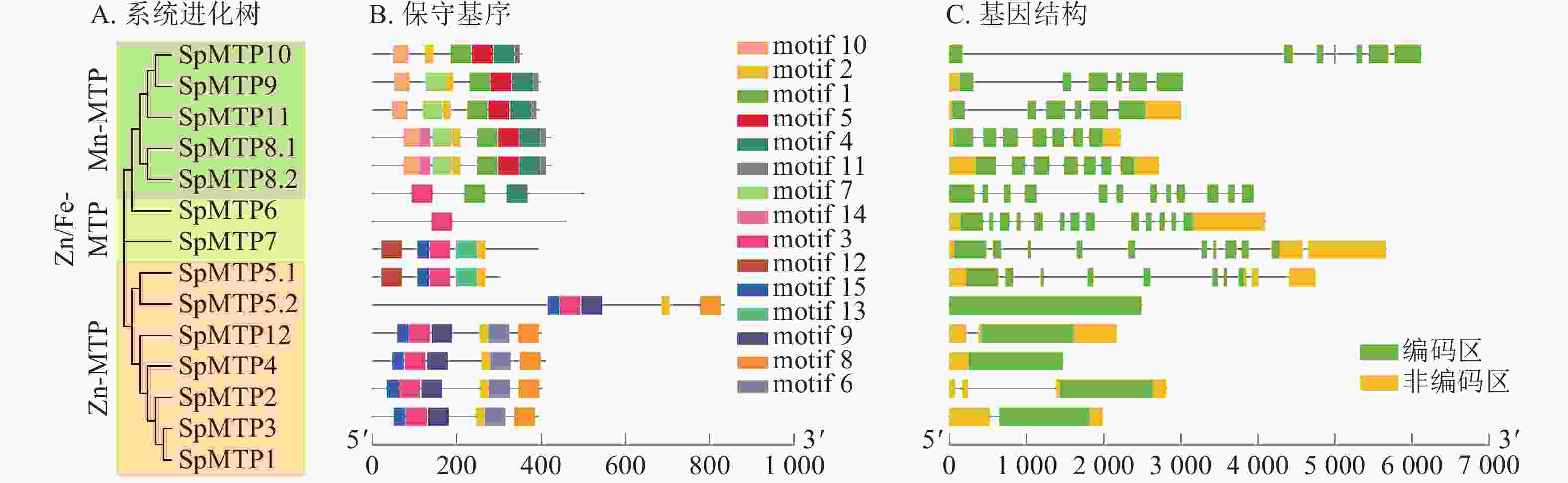
图 2 系统进化关系、保守基序及基因结构
Figure 2. Phylogenic relationship, conserved motif, gene structure analyses
-
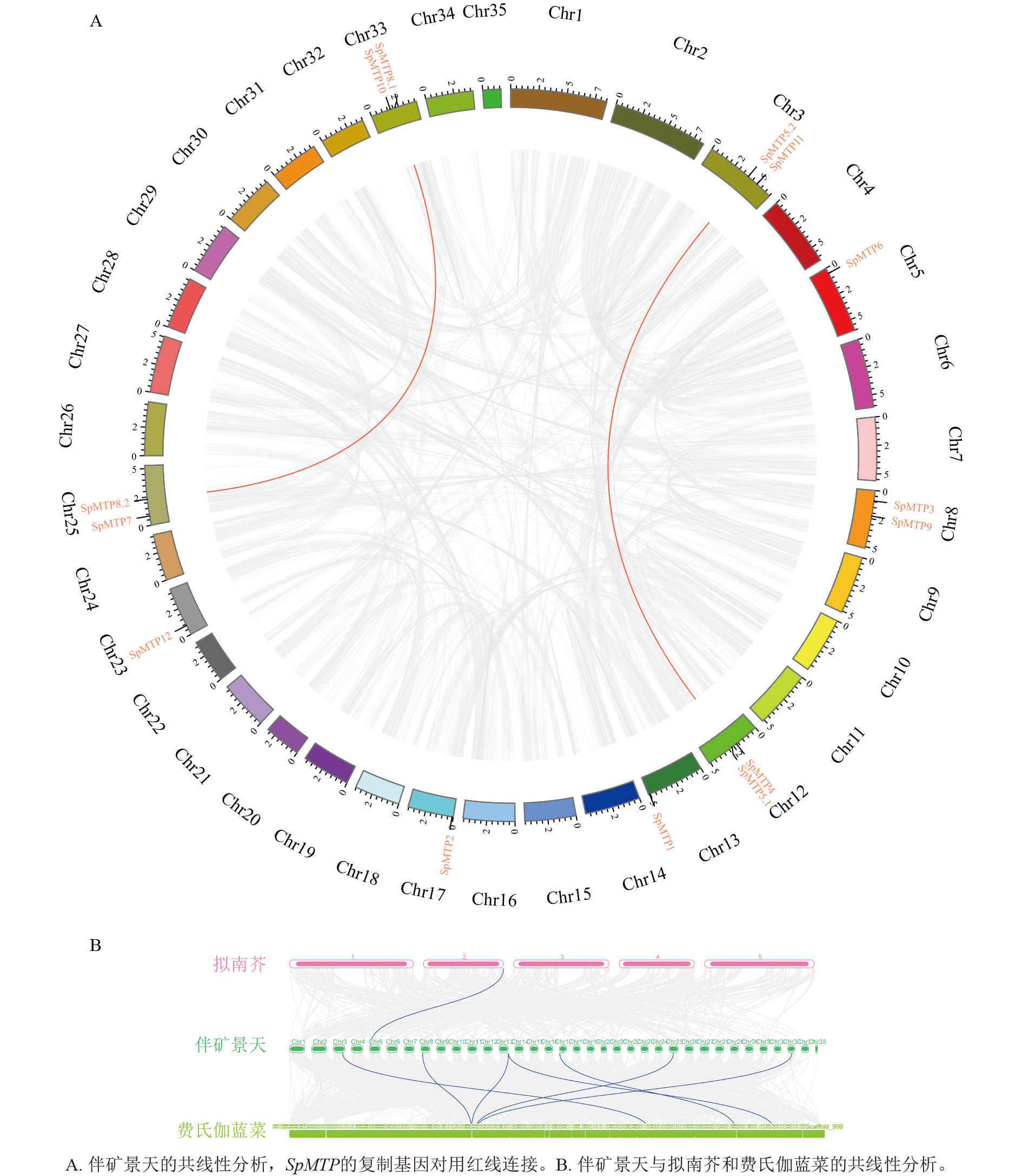
为探讨SpMTP基因家族的进化起源,分析了伴矿景天自身及其与拟南芥、费氏伽蓝菜之间的基因组共线性关系(图3)。自身共线性分析显示:伴矿景天基因组中存在2对SpMTP基因的共线性同源对,分别为SpMTP5.1/SpMTP5.2和SpMTP8.1/SpMTP8.2 (图3A),这2对基因均位于不同染色体上,推测为片段复制产生的旁系同源基因。此外,未检测到串联重复产生的SpMTP基因簇,表明片段复制是伴矿景天SpMTP家族扩张的主要驱动因素。共线性分析结果显示:多个SpMTP基因在伴矿景天与费氏伽蓝菜基因组间存在显著的共线性区块,这表明这些基因对可能起源于2个物种分化之前的共同祖先基因。相比之下,与拟南芥基因组之间存在共线性关系的SpMTP基因数量较少(1对),结合物种内共线性分析结果推测:伴矿景天中部分SpMTP基因(如上述 2 对片段复制基因)可能是其谱系特异性扩张的结果,这种扩张可能为其适应重金属胁迫环境提供了遗传基础[24]。
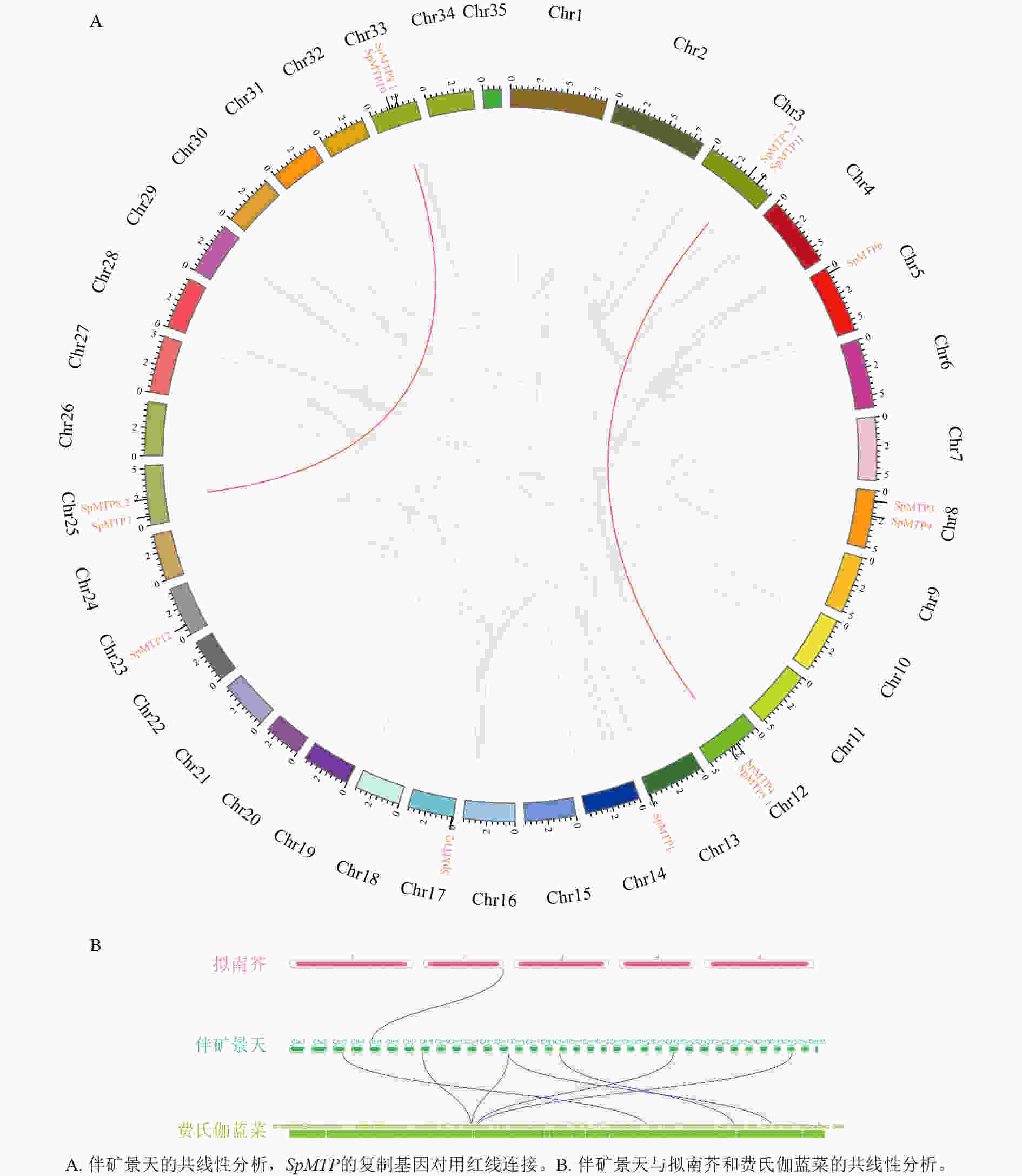
图 3 不同植物物种间SpMTP基因家族的共线性分析图
Figure 3. Synteny map of SpMTP gene family in different plant species
-
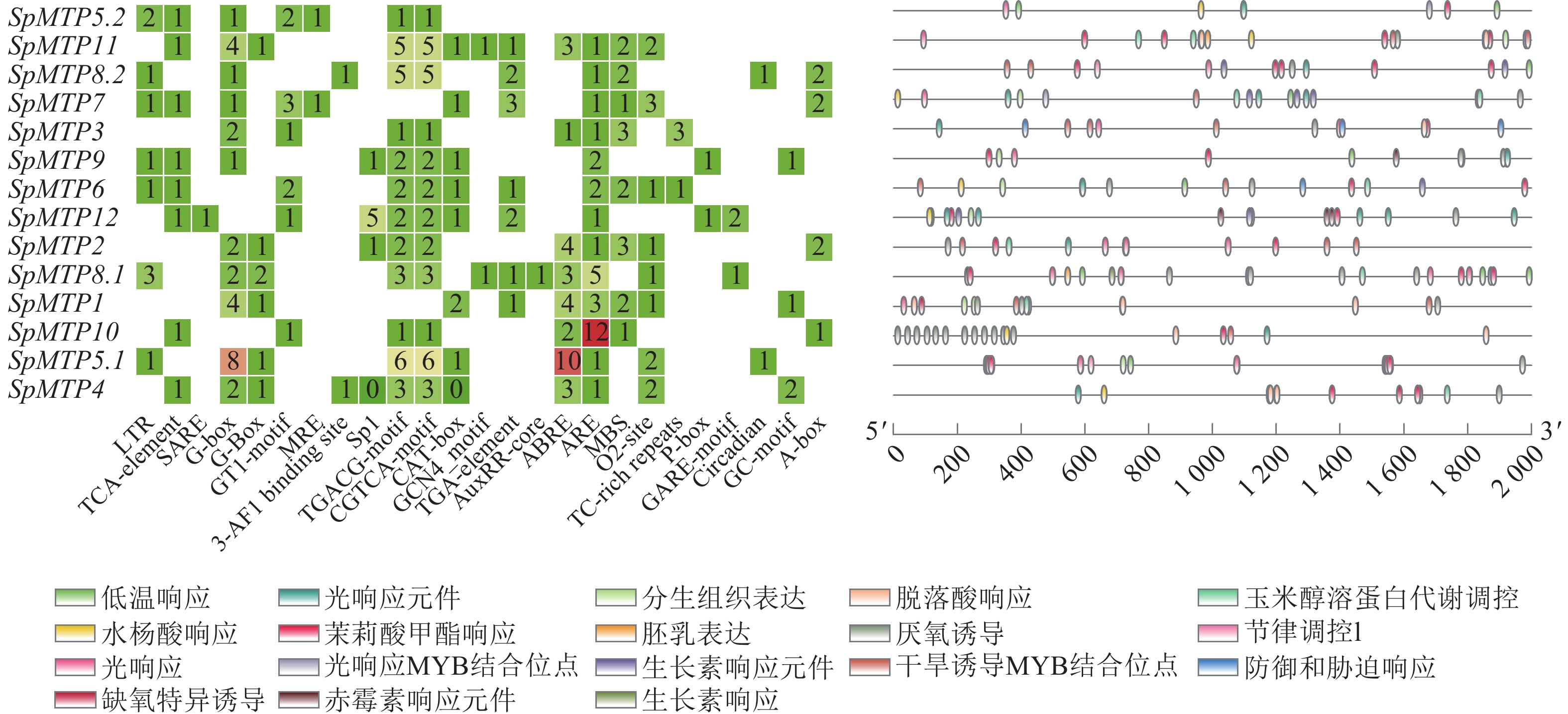
对14个SpMTP基因启动子序列的顺式作用元件进行系统分析,结果显示其含有丰富多样的调控元件(图4)。在环境胁迫响应元件中,所有基因均含有低温响应元件,其中SpMTP5.2、SpMTP11和SpMTP8.2的启动子区域该元件分布尤为密集。光响应元件在SpMTP7、SpMTP3和SpMTP9等基因的启动子中出现频率最高,而厌氧特异性诱导元件则在SpMTP6、SpMTP12和SpMTP2中显著富集。激素响应元件的分析表明:茉莉酸甲酯响应元件在SpMTP8.1、SpMTP1和SpMTP10中含量丰富。生长素响应元件主要分布于SpMTP5.1、SpMTP4和SpMTP5.2的启动子区域,而赤霉素响应元件在SpMTP11和SpMTP8.2中密度较高。脱落酸响应元件在超过85%的SpMTP基因启动子中被检测到,其中SpMTP3、SpMTP9和SpMTP6的含量最为突出。在组织特异性表达调控方面,分生组织表达元件主要存在于SpMTP7、SpMTP3和SpMTP9的启动子中,而胚乳表达元件则在SpMTP6、SpMTP12和SpMTP2中显著富集。此外,参与特定生理过程调控的元件分析显示:干旱诱导相关的MYB结合位点在SpMTP5.2、SpMTP11和SpMTP8.2中分布密集,昼夜节律控制元件主要存在于SpMTP7、SpMTP3和SpMTP9的启动子区域,而防御与胁迫响应元件在SpMTP6、SpMTP12和SpMTP2中含量最为丰富。
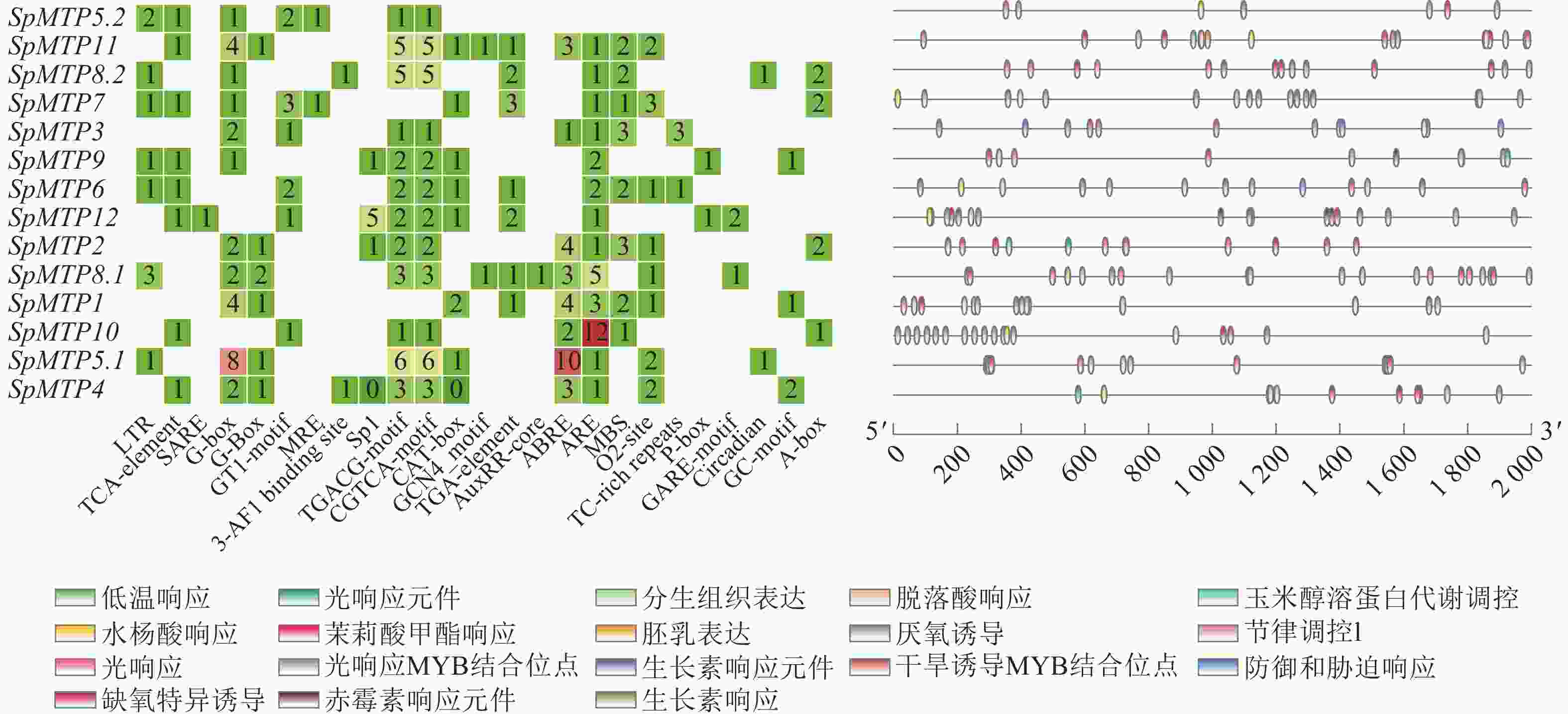
图 4 SpMTP家族基因启动子区域顺式作用元件分析
Figure 4. Analysis of cis-acting elements in the promoters of SpMTPs
这些顺式作用元件在不同SpMTP基因启动子中的组合模式存在明显差异,形成了各基因独特的调控特征。例如,SpMTP5.2的启动子同时富含低温响应、生长素响应和干旱诱导相关的MYB结合位点;SpMTP11的启动子则密集分布低温响应、赤霉素响应和干旱诱导相关元件;而SpMTP6启动子显著富集厌氧特异性诱导、脱落酸响应和防御与胁迫响应元件。这种元件组合的多样性为SpMTP基因在重金属胁迫下表现出的复杂表达模式提供了分子基础[25]。
-
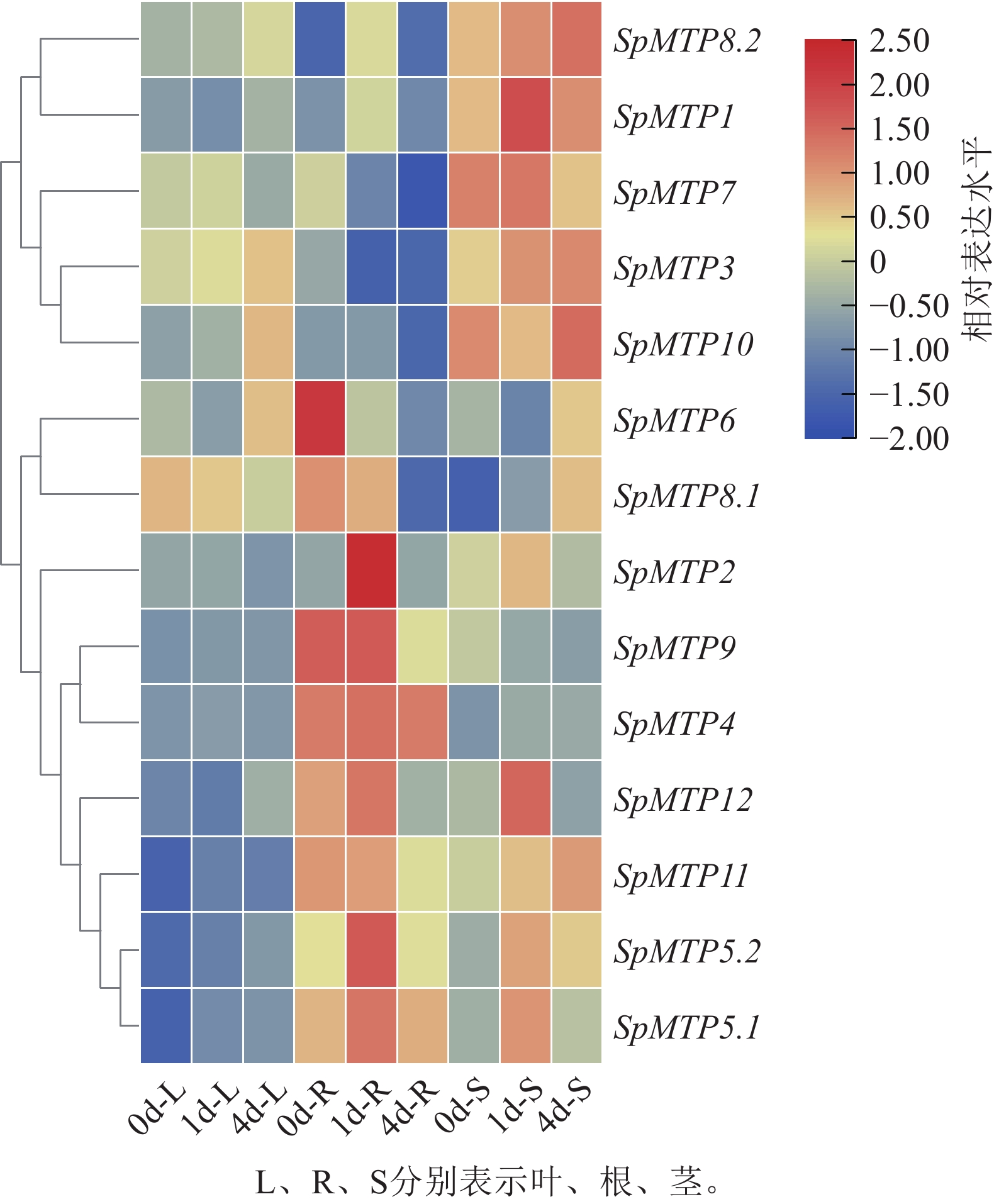
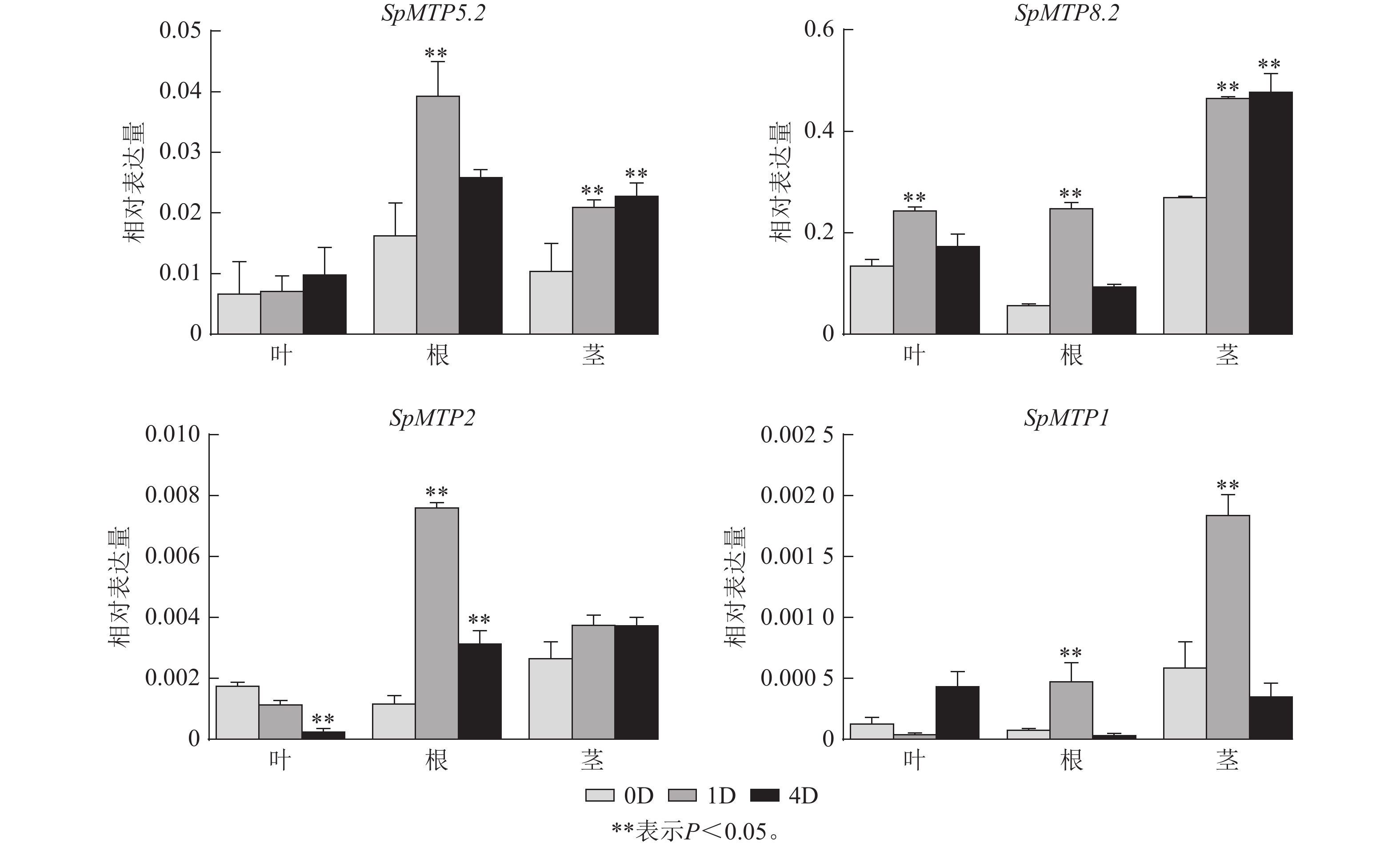
为深入解析SpMTP基因在镉胁迫应答中的功能分化,系统分析了14个SpMTP基因在根、茎、叶3种组织中不同时间点(0、1、4 d)的表达特征(图5),并选取4个覆盖不同亚族、且在转录组中呈现显著时空表达差异的核心基因(SpMTP5.2、SpMTP8.2、SpMTP2、SpMTP1)进行RT-qPCR验证。已发表的伴矿景天镉胁迫转录组数据[20]显示:测序数据均满足要求,可用于本研究的基因表达分析。验证结果显示:所选4个SpMTP基因在不同组织和镉胁迫时间点的表达趋势与转录组FPKM变化趋势高度一致(图6),证实前期转录组数据在本研究实验体系中具有良好的可靠性,可用于后续SpMTP基因表达模式的深入分析。在此基础上,结合转录组FPKM的系统分析,SpMTP家族基因在镉胁迫下呈现显著的组织表达特异性和时间动态响应规律。
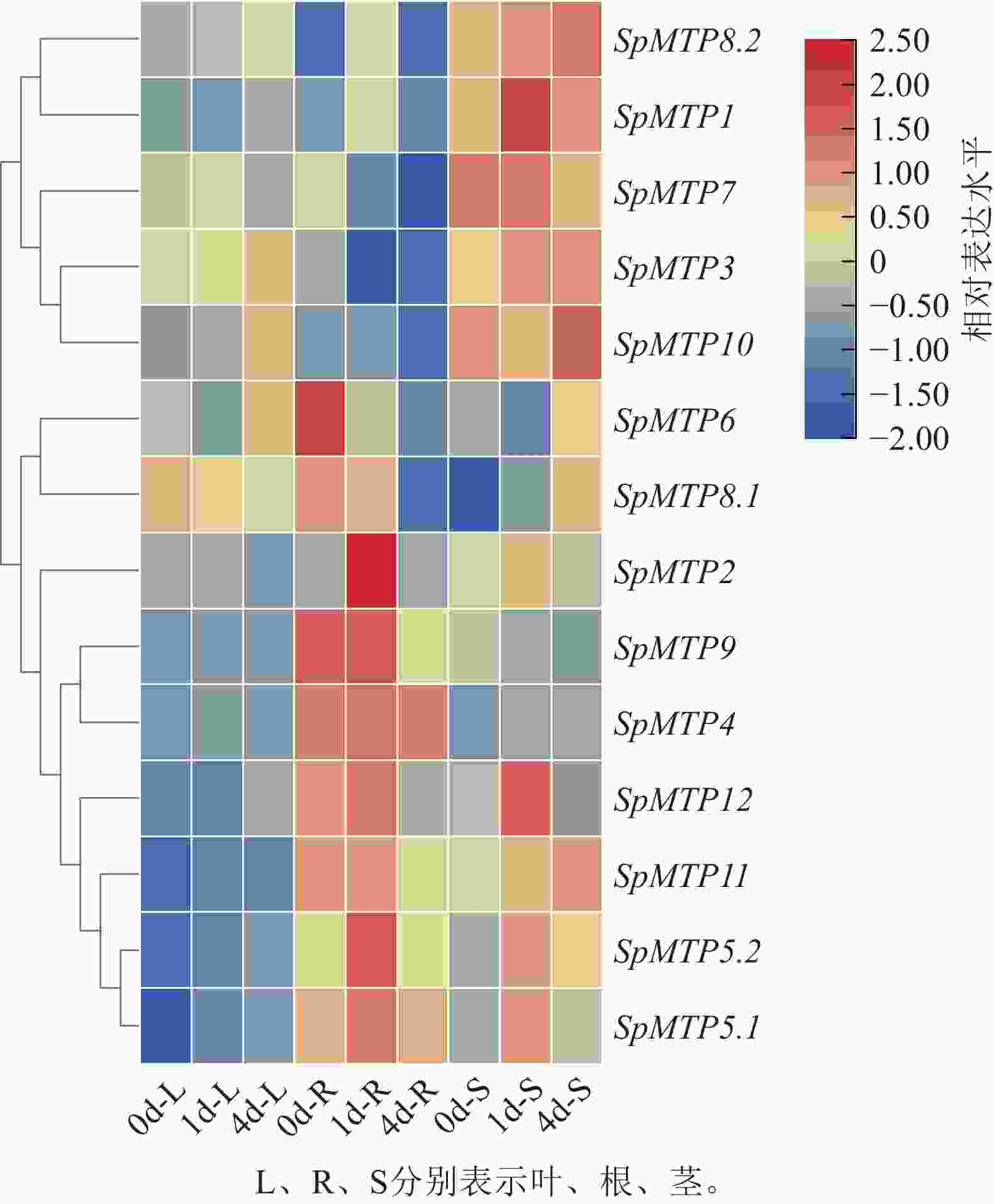
图 5 伴矿景天中14个SpMTP基因在过量金属胁迫下的表达水平
Figure 5. Expression levels of 14 SpMTP genes exposed to excess metals in S. plumbizincicola
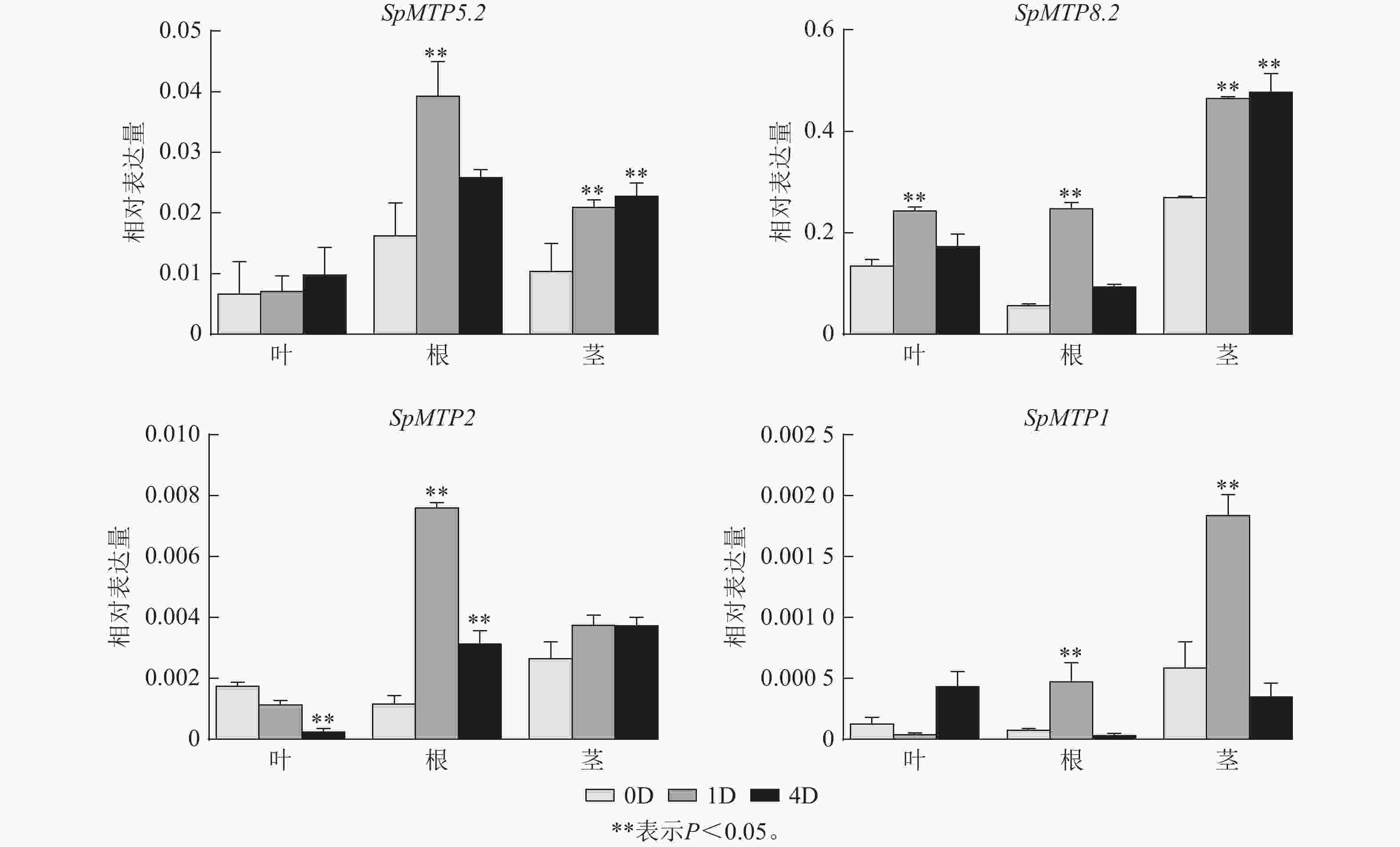
图 6 伴矿景天中4个SpMTP基因的RT-qPCR验证
Figure 6. RT-qPCR of 4 SpMTP genes in S. plumbizincicola
在组织表达特异性方面,不同SpMTP基因在根、茎、叶中展现出清晰的表达偏好。其中,SpMTP9在根部特异性高表达,其表达水平显著高于其他组织,暗示其在根部可能参与镉离子的吸收或初生解毒过程。SpMTP3在叶、茎部高表达,随着胁迫时间的延长,茎中表达量持续上升,预示其在叶片镉离子区隔化中的作用。SpMTP8.2在根、茎、叶中均有表达,该基因在根部对镉胁迫响应最显著,茎部表达量随胁迫时间逐步升高,叶部表达水平整体偏低。这表明SpMTP8.2主要在根部响应镉胁迫,同时参与茎部的转运过程。
从时间动态响应来看,SpMTP基因对镉胁迫的应答呈多样化的时序特征。部分基因如SpMTP8.2和SpMTP5.2在胁迫早期(1 d)即被快速诱导,在根部表现出显著的表达上调,体现了其对镉胁迫的快速响应能力。而SpMTP10则表现为典型的后期响应基因,在叶部于胁迫后期(4 d)被明显诱导,暗示其参与长期胁迫适应过程。此外,SpMTP3在叶部呈现持续上调的表达模式,在整个胁迫期间维持高表达水平,进一步支持其在重金属区隔化中的核心地位。
直系同源基因间的表达模式比较揭示了潜在的功能分化。SpMTP9与其直系同源基因SaMTP9虽均在根部高表达,但在其他组织中的表达特征存在明显差异,提示二者可能经历了功能特异化进化。同样,SpMTP3与SaMTP9虽同为高表达基因,但前者表现出更强的叶部表达特异性,反映了超积累植物特有的表达调控适应性。
综合表达特征与进化分析的结果,SpMTP基因家族在镉胁迫响应中形成了明确的功能分工体系:以SpMTP9为代表的根部特异表达基因可能负责镉离子的初始吸收,以SpMTP8.2为核心的茎部优势表达基因专门负责镉的维管运输,而以SpMTP3为主导的叶部高表达基因则承担最终的液泡区隔化功能。这种基于组织特异性与时间动态的精细表达调控网络,为伴矿景天高效镉超积累能力的形成奠定了重要的分子基础,也为后续关键基因的功能验证与研究方向的确定提供了科学依据。
-
MTP基因家族在植物重金属稳态维持中扮演着核心角色,然而其在超积累植物中的系统性进化与功能分化仍知之甚少。本研究在超积累植物伴矿景天中开展了SpMTP基因家族的全基因组鉴定与系统分析,揭示了其在进化、结构及表达调控上的独特特征,为理解植物重金属超积累的分子机制提供了新的视角[26]。
-
本研究发现SpMTP基因家族在进化上被分为7个主要亚族,且成员广泛分布其中(图1)。这一分布模式与拟南芥、水稻等模式植物类似,表明CDF/MTP蛋白家族的进化在植物界具有高度的保守性[27−28]。然而,SpMTP与同属非积累植物的东南景天中的MTP蛋白亲缘关系最近,且二者的直系同源基因在序列上高度相似,但在表达模式上存在显著的组织特异性分化。这种序列保守但功能分化的现象,暗示在景天属Sedum植物谱系中,MTP基因可能经历了适应性进化,通过表达调控分化驱动其功能向增强重金属耐受与转运的方向特化[29]。
共线性分析进一步为SpMTP家族的进化提供了证据。伴矿景天与费氏伽蓝菜之间广泛的共线性基因对共7对,说明这些基因在景天科内部物种分化前已存在,为景天科植物共有的保守基因。而相较于拟南芥,部分SpMTP基因缺乏直接的共线性同源物,这可能源于物种分化后的基因丢失,或是伴矿景天出现了谱系特异性的基因复制,后者影响更为显著。基因复制是基因家族扩展和新功能产生的重要驱动力[30]。在超积累植物中,基因复制可能为产生具有更强金属转运能力或新底物特异性的MTP旁系同源物提供了遗传素材。这一点在保守基序和基因结构分析中得到了支持:同一亚族内的高度保守性体现了功能约束,而不同亚族间的显著差异则为其功能分化奠定了结构基础。
-
本研究通过对SpMTP基因在镉胁迫下表达模式的系统解析,揭示了该基因家族成员在重金属超积累过程中存在的精细功能分工。表达谱分析显示:不同SpMTP基因在根、茎、叶组织中呈现出截然不同的表达特征,且其响应镉胁迫的时间动态各具特色,共同构成了一个协同高效的重金属转运与解毒网络。
在根部,SpMTP9表现出显著的组织特异性高表达,其在胁迫早期虽未表现出明显的上调趋势,但始终维持较高的表达水平,暗示该基因可能负责根部镉离子的基础吸收或初生解毒。其同源基因SaMTP9在东南景天根部也呈现持续高表达特征,进一步支持SpMTP9在根部重金属稳态维持中的功能。结合其液泡膜定位预测,SpMTP9蛋白可能通过将根部吸收的镉离子暂存于液泡中,来避免细胞质毒害,同时为后续向地上部的运输提供缓冲。
在运输环节,SpMTP8.2在茎部持续表达且随胁迫时间缓慢上调,结合其液泡膜定位预测,推测其可能通过茎部细胞内的液泡区隔化调节镉离子动态平衡,为离子向地上部的高效转运提供缓冲,其具体转运功能需后续功能验证实验证实。
在最终储存部位,SpMTP3在叶部的持续高表达特征与伴矿景天叶片镉富集特性高度契合。结合其液泡膜定位预测,推测其可能参与叶部镉离子的液泡区隔化过程,其转运活性需通过异源表达等实验进一步验证。
此外,不同基因的时序表达特征进一步丰富了其在超积累过程中的功能内涵。SpMTP5.2等基因可能参与伴矿景天对镉的早期快速响应;而SpMTP10等基因的后期诱导则可能参与长期胁迫下的生理代谢调节。这种时间维度上的功能互补,确保了植物在整个胁迫周期内都能维持高效的镉积累与解毒能力。
-
本研究通过全基因组鉴定,明确伴矿景天含有14个SpMTP基因,这些基因编码的蛋白均含有完整的CDF保守结构域,可划分为7个亚族,片段复制是其家族扩张的主要驱动因素。SpMTP基因家族在进化上与东南景天SaMTP家族高度保守,但在表达模式上存在超积累植物特有的组织特异性分化。镉胁迫下,SpMTP基因呈现显著的时空表达特征:SpMTP9在根部持续高表达,可能参与镉离子的吸收与初生解毒;SpMTP8.2在根部胁迫早期上调,茎部持续表达,推测负责镉离子的长途运输;SpMTP3在叶部显著上调,可能承担液泡区隔化储存功能。这些基因通过时空协同作用,构成了伴矿景天镉超积累的分子基础。本研究为解析超积累植物MTP基因家族的进化与功能分化提供了理论依据,筛选出的SpMTP3、SpMTP8.2、SpMTP9等候选基因可为后续植物修复技术改良提供基因资源。
Genome-wide identification and expression patterns in response to cadmium stress of SpMTP gene family in Sedum plumbizincicola
-
摘要:
目的 金属耐受蛋白(MTP)参与植物重金属耐受与转运过程,伴矿景天Sedum plumbizincicola为镉超积累植物,其MTP基因家族的系统特征与镉胁迫响应机制尚未明确。鉴定该家族成员并解析其结构、进化及镉(Cd)胁迫表达规律,可为阐明伴矿景天镉超积累分子机制提供理论依据。 方法 采用生物信息学方法,基于阳离子扩散辅助蛋白(CDF)家族保守结构域(PF01545)鉴定SpMTP基因;通过MEGA、MEME、TBtools等软件分析其系统进化、保守基序、基因结构及共线性关系;利用PlantCARE预测启动子顺式作用元件;结合RNA-seq数据分析14个SpMTP基因在镉胁迫(400 μmol·L−1)下根、茎、叶组织(0、1、4 d)的表达特征,并通过实时荧光定量PCR (RT-qPCR)验证部分基因表达可靠性。 结果 共鉴定出14个SpMTP基因,编码蛋白含4~16个跨膜结构域,多定位于液泡膜或细胞膜;系统进化分析将其划分为7个亚族,与近缘非积累植物东南景天S. alfredii同源蛋白亲缘关系最近。共线性分析显示:复制是SpMTP家族扩张的主要驱动因素;启动子区域富含胁迫响应、激素信号及组织特异性表达相关顺式作用元件。表达谱分析表明:SpMTP基因呈现显著时空特异性,SpMTP9在根部持续高表达,SpMTP8.2在根部早期诱导上调,SpMTP3在叶部持续高表达,三者分别参与镉的吸收、转运及液泡区隔化。RT-qPCR验证显示:所选基因表达趋势与RNA-seq数据高度一致,证实转录组数据可靠。 结论 伴矿景天SpMTP基因家族通过进化扩张与功能分化,形成了根-茎-叶协同的镉胁迫响应网络,其组织特异性表达特征是伴矿景天镉超积累能力的重要分子基础。图6表1参30 Abstract:Objective Metal tolerance protein (MTP) is involved in heavy metal tolerance and transport in plant. As a cadmium (Cd) hyperaccumulator, Sedum plumbizincicola has poorly characterized MTP family features and Cd response mechanisms. This study identified SpMTP family members, and analyzed their structure, evolution and expression profiles under Cd stress, to provide theoretical support for exploring the molecular mechanisms of Cd hyperaccumulation in S. plumbizincicola. Method Bioinformatics methods were used to identify SpMTP genes based on the conserved domain (PF01545) of the cation diffusion facilitator (CDF) family. Phylogenetic analysis, conserved motif identification, gene structure analysis, and synteny analysis were performed using MEGA, MEME, and TBtools software. Cis-acting elements in the promoter regions were predicted by PlantCARE. RNA-seq technology was used to analyze the expression profiles of 14 SpMTP genes in roots, stems, and leaves of S. plumbizincicola under Cd stress (400 μmol·L−1) at different time points (0, 1, and 4 d), and real-time quantitative PCR (RT-qPCR) was used to verify the expression reliability of selected genes. Result A total of 14 non-redundant SpMTP genes were identified in S. plumbizincicola. The encoded proteins contained 4 to 16 transmembrane domains (TMDs) and were mostly localized to the vacuolar membrane or cell membrane. Phylogenetic analysis classified these SpMTP proteins into 7 distinct subfamilies, with the closest evolutionary relationship to orthologs in S. alfredii. Synteny analysis indicated that segmental duplication events were the main driver of SpMTP family expansion. Promoter analysis revealed a variety of cis-acting elements related to stress response, hormone signaling, and tissue-specific expression. Expression profiling showed that SpMTP genes exhibited distinct spatiotemporal expression patterns under Cd stress: SpMTP9 was consistently highly expressed in roots, SpMTP8.2 was rapidly induced in roots at the early stage of stress, and SpMTP3 was consistently highly expressed in leaves, suggesting their specialized roles in Cd uptake, translocation, and vacuolar sequestration, respectively. The RT-qPCR results confirmed the reliability of RNA-seq data. Conclusion SpMTP gene family in S. plumbizincicola has formed a coordinated Cd stress response network in roots, stems, and leaves through evolutionary expansion and functional differentiation, and its tissue-specific expression pattern serves as an important molecular basis for the Cd hyperaccumulation capacity of S. plumbizincicola. [Ch, 6 fig. 1 tab. 30 ref.] -
Key words:
- Sedum plumbizincicola /
- SpMTP /
- gene family /
- evolutionary analysis /
- cadmium stress
-
图 1 伴矿景天与其他6种植物MTP蛋白的系统进化分析
Figure 1 Phylogenetic analysis of MTP proteins from S. plumbizincicola and other 6 plant species
图 2 系统进化关系、保守基序及基因结构
Figure 2 Phylogenic relationship, conserved motif, gene structure analyses
图 3 不同植物物种间SpMTP基因家族的共线性分析图
Figure 3 Synteny map of SpMTP gene family in different plant species
图 4 SpMTP家族基因启动子区域顺式作用元件分析
Figure 4 Analysis of cis-acting elements in the promoters of SpMTPs
图 5 伴矿景天中14个SpMTP基因在过量金属胁迫下的表达水平
Figure 5 Expression levels of 14 SpMTP genes exposed to excess metals in S. plumbizincicola
表 1 伴矿景天SpMTP基因家族成员基本信息
Table 1. Information of SpMTP gene family in S. plumbizincicola genome
基因名称 基因编号 基因长
度/bp编码区
长度/bp分子量/Da 氨基
酸数不稳定
系数脂肪族
指数总平均
亲水性等电点 跨膜结构
域数量亚细胞定位 SpMTP5.2 evm.model.000001F.133 4 751 906 33 804.93 301 39.95 90.70 0.099 8.9 5 细胞膜/液泡膜 SpMTP11 evm.model.000001F.377 3 009 1 185 44 577.15 394 42.06 105.43 0.069 4.9 6 液泡膜 SpMTP8.2 evm.model.000002F.140 2 722 1 266 46 436.58 421 44.01 107.72 0.120 5.1 6 液泡膜 SpMTP7 evm.model.000002F.531 4 104 1 374 50 592.04 457 39.21 94.09 −0.028 7.8 5 液泡膜 SpMTP3 evm.model.000009F.229 2 814 1 206 44 609.16 401 31.38 106.76 −0.021 5.9 6 液泡膜 SpMTP9 evm.model.000009F.527 3 030 1 191 45 410.30 396 46.02 94.55 −0.124 6.6 6 细胞膜/液泡膜 SpMTP6 evm.model.000010F.138 3 954 1 506 54 669.90 501 42.13 98.30 −0.001 6.8 5 液泡膜 SpMTP12 evm.model.000014F.173 2 499 2 499 92 580.50 832 42.38 98.49 0.072 8.4 16 细胞膜/液泡膜 SpMTP2 evm.model.000015F.32 1 479 1 227 45 123.59 408 40.28 107.03 0.017 6.1 6 液泡膜 SpMTP8.1 evm.model.000078F.209 2 229 1 266 46 436.37 421 45.52 108.43 0.136 4.9 6 液泡膜 SpMTP1 evm.model.000106F.135 1 992 1 176 43 433.16 391 39.09 109.97 0.131 5.9 6 液泡膜 SpMTP10 evm.model.000127F.82 6 121 1 065 40 650.67 354 46.93 91.41 −0.129 6.2 4 细胞膜/液泡膜 SpMTP5.1 evm.model.000138F.18 5 668 1 179 43 832.34 392 48.38 92.73 0.084 7.2 6 液泡膜 SpMTP4 evm.model.000148F.36 2 170 1 197 43 582.79 398 35.91 103.79 0.057 6.0 6 液泡膜  下载: 导出CSV
下载: 导出CSV
-
[1] RIZWAN M, ALI S, REHMAN M Z U, et al. A critical review on the effects of zinc at toxic levels of cadmium in plants[J]. Environmental Science and Pollution Research, 2019, 26(7): 6279−6289. DOI: 10.1007/s11356-019-04174-6. [2] MAHAR A, WANG Ping, ALI A, et al. Challenges and opportunities in the phytoremediation of heavy metals contaminated soils: a review[J]. Ecotoxicology and Environmental Safety, 2016, 126: 111−121. DOI: 10.1016/j.ecoenv.2015.12.023. [3] ZHU Xia, TU Chen, ZHOU Jiawen, et al. Cadmium phytoextraction by Sedum alfredii and Sedum plumbizincicola: mechanisms, challenges and prospects[J]. International Journal of Phytoremediation, 2025, 27(6): 852−860. DOI: 10.1080/15226514.2025.2451714. [4] PENG Jiashi, WANG Yuejun, DING Ge, et al. A pivotal role of cell wall in cadmium accumulation in the Crassulaceae hyperaccumulator Sedum plumbizincicola[J]. Molecular Plant, 2017, 10(5): 771−774. DOI: 10.1016/j.molp.2016.12.007. [5] SHARMA S S, DIETZ K J, MIMURA T. Vacuolar compartmentalization as indispensable component of heavy metal detoxification in plants[J]. Plant, Cell & Environment, 2016, 39(5): 1112−1126. DOI: 10.1111/pce.12706. [6] van der ZAAL B J, NEUTEBOOM L W, PINAS J E, et al. Overexpression of a novel Arabidopsis gene related to putative zinc-transporter genes from animals can lead to enhanced zinc resistance and accumulation[J]. Plant Physiology, 1999, 119(3): 1047−1056. DOI: 10.1104/pp.119.3.1047. [7] KOBAE Y, UEMURA T, SATO M H, et al. Zinc transporter of Arabidopsis thaliana AtMTP1 is localized to vacuolar membranes and implicated in zinc homeostasis[J]. Plant and Cell Physiology, 2004, 45(12): 1749−1758. DOI: 10.1093/pcp/pci015. [8] ARRIVAULT S, SENGER T, KRÄMER U. The Arabidopsis metal tolerance protein AtMTP3 maintains metal homeostasis by mediating Zn exclusion from the shoot under Fe deficiency and Zn oversupply[J]. The Plant Journal, 2006, 46(5): 861−879. DOI: 10.1111/j.1365-313X.2006.02746.x. [9] EROGLU S, MEIER B, von WIRÉN N, et al. The vacuolar manganese transporter MTP8 determines tolerance to iron deficiency-induced chlorosis in Arabidopsis[J]. Plant Physiology, 2016, 170(2): 1030−1045. DOI: 10.1104/pp.15.01194. [10] LIU Peng, SUN Liang, ZHANG Yu, et al. The metal tolerance protein OsMTP11 facilitates cadmium sequestration in the vacuoles of leaf vascular cells for restricting its translocation into rice grains[J]. Molecular Plant, 2024, 17(11): 1733−1752. DOI: 10.1016/j.molp.2024.09.012. [11] LI Jia, ABBAS M, DESOKY E M, et al. Analysis of metal tolerance protein (MTP) family in sunflower (Helianthus annus L.) and role of HaMTP10 as cadmium antiporter under moringa seed extract[J]. Industrial Crops and Products, 2023, 202: 117023. DOI: 10.1016/j.indcrop.2023.117023. [12] FENG Shangguo, HOU Kailin, ZHANG Hongshan, et al. Investigation of the role of TmMYB16/123 and their targets (TmMTP1/11) in the tolerance of Taxus media to cadmium[J]. Tree Physiology, 2023, 43(6): 1009−1022. DOI: 10.1093/treephys/tpad019. [13] LIU Mingying, FENG Ying, WANG Miao, et al. Sedum alfredii Hance: a cadmium and zinc hyperaccumulating plant[J]. Ecotoxicology and Environmental Safety, 2025, 290: 117588. DOI: 10.1016/j.ecoenv.2024.117588. [14] MISTRY J, CHUGURANSKY S, WILLIAMS L, et al. Pfam: the protein families database in 2021[J]. Nucleic Acids Research, 2021, 49(D1): D412−D419. DOI: 10.1093/nar/gkaa913. [15] GASTEIGER E, HOOGLAND C, GATTIKER A, et al. Protein identification and analysis tools on the ExPASy server[M]//WALKER J M. The Proteomics Protocols Handbook. Totowa: Humana Press, 2005: 571−607. [16] KROGH A, LARSSON B, von HEIJNE G, et al. Predicting transmembrane protein topology with a hidden Markov model: application to complete genomes[J]. Journal of Molecular Biology, 2001, 305(3): 567−580. DOI: 10.1006/jmbi.2000.4315. [17] KUMAR S, STECHER G, LI M, et al. MEGA X: molecular evolutionary genetics analysis across computing platforms[J]. Molecular Biology and Evolution, 2018, 35(6): 1547−1549. DOI: 10.1093/molbev/msy096. [18] BAILEY T L, JOHNSON J, GRANT C E, et al. The MEME suite[J]. Nucleic Acids Research, 2015, 43(W1): W39−W49. DOI: 10.1093/nar/gkv416. [19] WANG Yupeng, TANG Haibao, DEBARRY J D, et al. MCScanX: a toolkit for detection and evolutionary analysis of gene synteny and collinearity[J]. Nucleic Acids Research, 2012, 40(7): e49−e49. DOI: 10.1093/nar/gkr1293. [20] HAN Xiaojiao, YIN Hengfu, SONG Xixi, et al. Integration of small RNAs, degradome and transcriptome sequencing in hyperaccumulator Sedum alfredii uncovers a complex regulatory network and provides insights into cadmium phytoremediation[J]. Plant Biotechnology Journal, 2016, 14(6): 1470−1483. DOI: 10.1111/pbi.12512. [21] MONTANINI B, BLAUDEZ D, JEANDROZ S, et al. Phylogenetic and functional analysis of the cation diffusion facilitator (CDF) family: improved signature and prediction of substrate specificity[J]. BMC Genomics, 2007, 8(1): 107. DOI: 10.1186/1471-2164-8-107. [22] ZHANG Jie, MARTINOIA E, LEE Y. Vacuolar transporters for cadmium and arsenic in plants and their applications in phytoremediation and crop development[J]. Plant and Cell Physiology, 2018, 59(7): 1317−1325. DOI: 10.1093/pcp/pcy006. [23] EMMS D M, KELLY S. OrthoFinder: phylogenetic orthology inference for comparative genomics[J]. Genome Biology, 2019, 20(1): 238. DOI: 10.1186/s13059-019-1832-y. [24] HANIKENNE M, TALKE I N, HAYDON M J, et al. Evolution of metal hyperaccumulation required cis-regulatory changes and triplication of HMA4[J]. Nature, 2008, 453(7193): 391−395. DOI: 10.1038/nature06877. [25] YAMAGUCHI-SHINOZAKI K, SHINOZAKI K. Transcriptional regulatory networks in cellular responses and tolerance to dehydration and cold stresses[J]. Annual Review of Plant Biology, 2006, 57: 781−803. DOI: 10.1146/annurev.arplant.57.032905.105444. [26] KRÄMER U. Metal hyperaccumulation in plants[J]. Annual Review of Plant Biology, 2010, 61(1): 517−534. DOI: 10.1146/annurev-arplant-042809-112156. [27] GUSTIN J L, ZANIS M J, SALT D E. Structure and evolution of the plant cation diffusion facilitator family of ion transporters[J]. BMC Evolutionary Biology, 2011, 11(1): 76. DOI: 10.1186/1471-2148-11-76. [28] COLANGELO E P, GUERINOT M L. Put the metal to the petal: metal uptake and transport throughout plants[J]. Current Opinion in Plant Biology, 2006, 9(3): 322−330. DOI: 10.1016/j.pbi.2006.03.015. [29] VERBRUGGEN N, HERMANS C, SCHAT H. Molecular mechanisms of metal hyperaccumulation in plants[J]. New Phytologist, 2009, 181(4): 759−776. DOI: 10.1111/j.1469-8137.2008.02748.x. [30] CONANT G C, WOLFE K H. Turning a hobby into a job: how duplicated genes find new functions[J]. Nature Reviews Genetics, 2008, 9(12): 938−950. DOI: 10.1038/nrg2482. -

-
链接本文:
https://zlxb.zafu.edu.cn/article/doi/10.11833/j.issn.2095-0756.20260147
 点击查看大图
点击查看大图
计量
- 文章访问数: 11
- HTML全文浏览量: 3
- PDF下载量: 0
- 被引次数: 0
