





-
叶绿体是植物细胞内的关键细胞器,在光合作用和碳水化合物代谢等过程中扮演重要角色[1]。近年来,随着高通量测序技术的飞速发展,植物叶绿体基因组的研究也在不断深入。被子植物的叶绿体基因组通常呈现为四分体结构,由1个大单拷贝区(large single copy region,LSC)、1个小单拷贝区(small single copy region,SSC)以及2个反向重复区(inverted repeat regions A和B,IRa和IRb)共4部分组成[2]。然而,也有一些植物,如豆科Leguminosae的箭筈豌豆Vicia sativa[3]、三叶草Trifolium[4]等由于缺失1个反向重复序列,因而其叶绿体基因组结构组成为LSC、SSC和IR;而绿潮藻Ulva flexuosa的反向重复序列则完全缺失[5]。叶绿体基因组是母系遗传,具有高度的保守性,不受核基因组的影响,被用来进行植物系统发育、遗传多样性分析等研究[6]。
十字花科Brassicaceae家族十分庞大,约包含351个属3 977个种[7]。芸薹属Brassica作为十字花科植物中的关键成员,包括白菜B. rapa、甘蓝B. oleracea、芥菜B. juncea和油菜B. napus等重要的蔬菜作物和油料作物[8]。白菜作为芸薹属中的一员,经过长期人工选择和环境变化,出现了诸多变种。菜心B. rapa var. parachinensis是白菜种下的一个变种,因口感好、营养丰富而备受消费者青睐,在中国南方地区尤其是华南地区多有栽培[9−11]。然而,菜心与其他白菜变种以及十字花科成员之间的进化关联尚不明确。叶绿体基因组在一定程度上可呈现植物的进化历程,目前针对十字花科植物叶绿体基因组的研究已取得较为丰硕的成果。早在20多年前,模式植物拟南芥Arabidopsis thaliana的叶绿体基因组即已完成解析工作,为后续十字花科植物的遗传进化研究奠定了重要基础[12]。近些年来,随着研究的持续深入,紫菜薹B. rapa var. purpuraria[13]、芥菜[14]、萝卜Raphanus sativus[15]等众多十字花科植物的叶绿体基因组也已相继被研究,然而关于菜心叶绿体基因组的研究还未见报道。本研究以菜心品种‘油青四九’B. rapa var. parachinensis‘Youqing 49’为材料,借助高通量测序技术结合生物信息学方法对其叶绿体基因组进行测序、组装并注释,对其基本结构特征等进行分析,明晰了菜心与其他白菜变种以及十字花科其他成员间的进化关系。研究结果有助于弥补菜心叶绿体基因组研究空白,为十字花科植物系统发育分析及遗传资源利用提供科学依据。
-
本研究所用材料为菜心‘油青四九’,种植于浙江农林大学园艺科学学院气候室(30°26′N, 119°72′E),2024年8月取健康植株的叶片,用锡箔纸包好后置于液氮中速冻,取出后保存于−80 ℃冰箱。使用通用型植物DNA提取试剂盒(D312)提取总DNA,使用Illumina NovaSeq 6000平台进行测序。
-
使用fastp v0.23.4[16]软件对原始数据过滤获得净数据(clean data)。采用GetOrganelle v1.7.7.1[17]软件组装叶绿体基因组,分别运用prodigal v2.6.3[18]、hmmer v3.1b2 [19]和aragorn v1.2.38[20]对叶绿体的编码序列(coding sequence, CDS)注释,预测rRNA和tRNA。叶绿体基因组图谱使用OGDRAW[21]软件绘制。
-
使用vmatch v2.3.09软件鉴定重复序列,相关参数设定为:海明距离为3,最小长度为30 bp,鉴定形式为正向(forward)、回文(palindromic)、反向(reverse)、互补(complement)共4种。通过MISA v1.0工具[22]对简单重复序列(simple sequence repeats, SSRs)进行分析。参数配置为:单碱基重复次数不少于8次,双碱基重复不少于5次,而3碱基、4碱基、5碱基及6碱基重复则要求至少出现3次。
-
在美国国家生物技术信息中心(NCBI)上下载包括拟南芥(KX551970)、埃塞俄比亚芥B. carinata (MW628493)、芥菜(KT581449)、油菜(NC_016734)、黑芥B. nigra (KT878383)、甘蓝(MT721156)、白菜(NC_040849)等7种十字花科植物的叶绿体基因组。利用MAFFT v7.427 [23]对不同物种间的同源基因序列进行了全局比对分析,并通过dnasp5[24]工具计算核苷酸多样性(nucleotide diversity, Pi)。使用集思慧远的云平台工具CPJSdraw将各物种的叶绿体基因组边界信息进行可视化处理,使用Mauve v2.3.1[25]进行基因组比对。
-
从NCBI数据库下载25种十字花科植物的叶绿体基因组序列,外类群选择番木瓜Carica papaya(EU431223),以此来进行系统发育关系的分析。分别使用MAFFT v7.427[23]和RAxML v8.2.10[26]进行多序列比对和进化树构建。
-
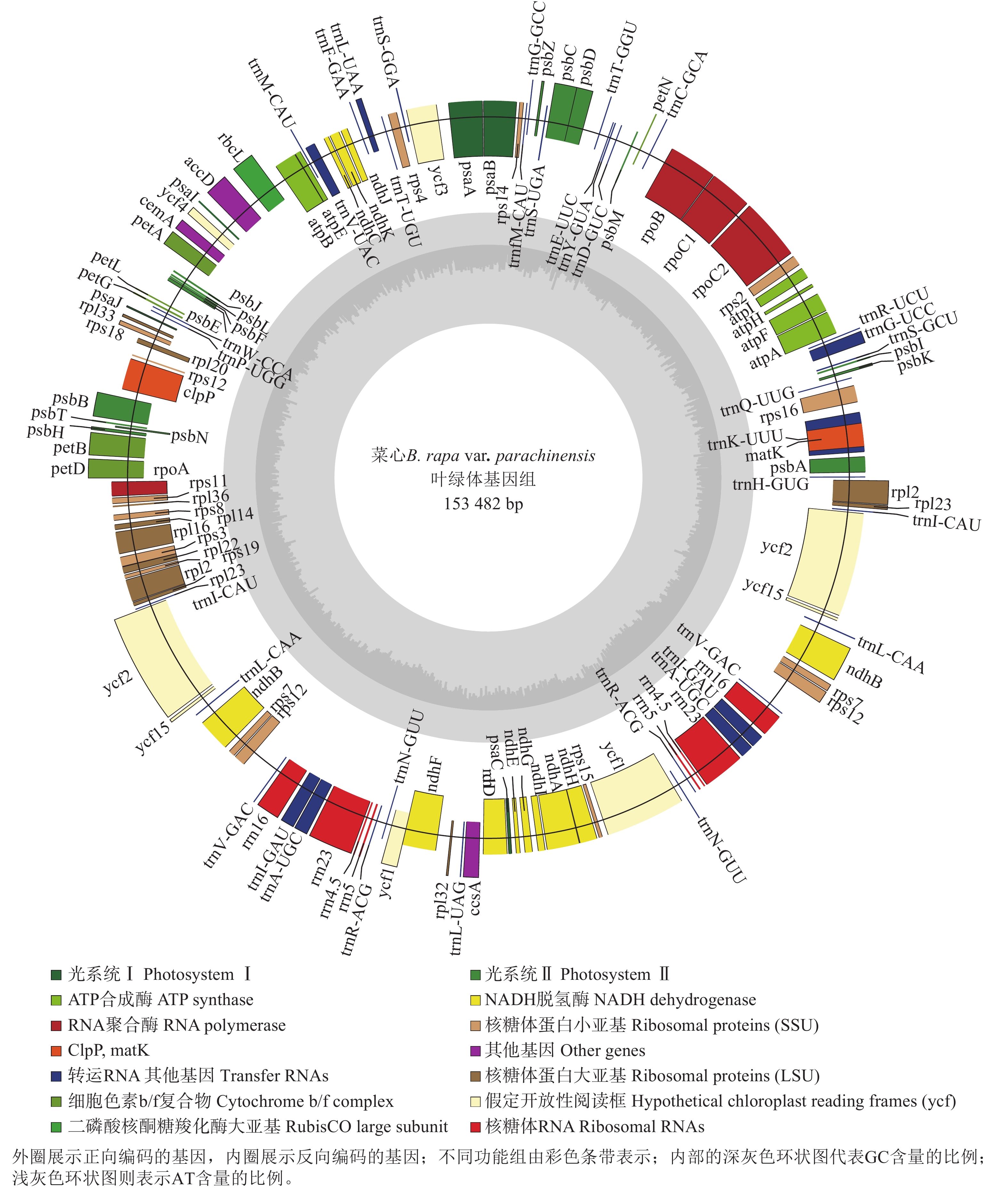
菜心叶绿体基因组(GenBank登录号:PX244600)呈典型的四分体结构,长度为153 482 bp,包含反向重复序列a (IRa)和反向重复序列b (IRb)以及大单拷贝区域(LSC)和小单拷贝区域(SSC),序列长度分别为26 213、26 213、83 281、17 775 bp (图1和表1)。对碱基组成进行分析,结果显示:菜心叶绿体基因组中A、C、G、T的含量分别为31.35%、18.51%、17.85%、32.39%;整个菜心叶绿体基因组中的GC总含量为36.36%,其中IRa和IRb的GC含量均为42.34%,高于LSC (34.12%)和SSC (29.20%)中的GC含量(表1)。
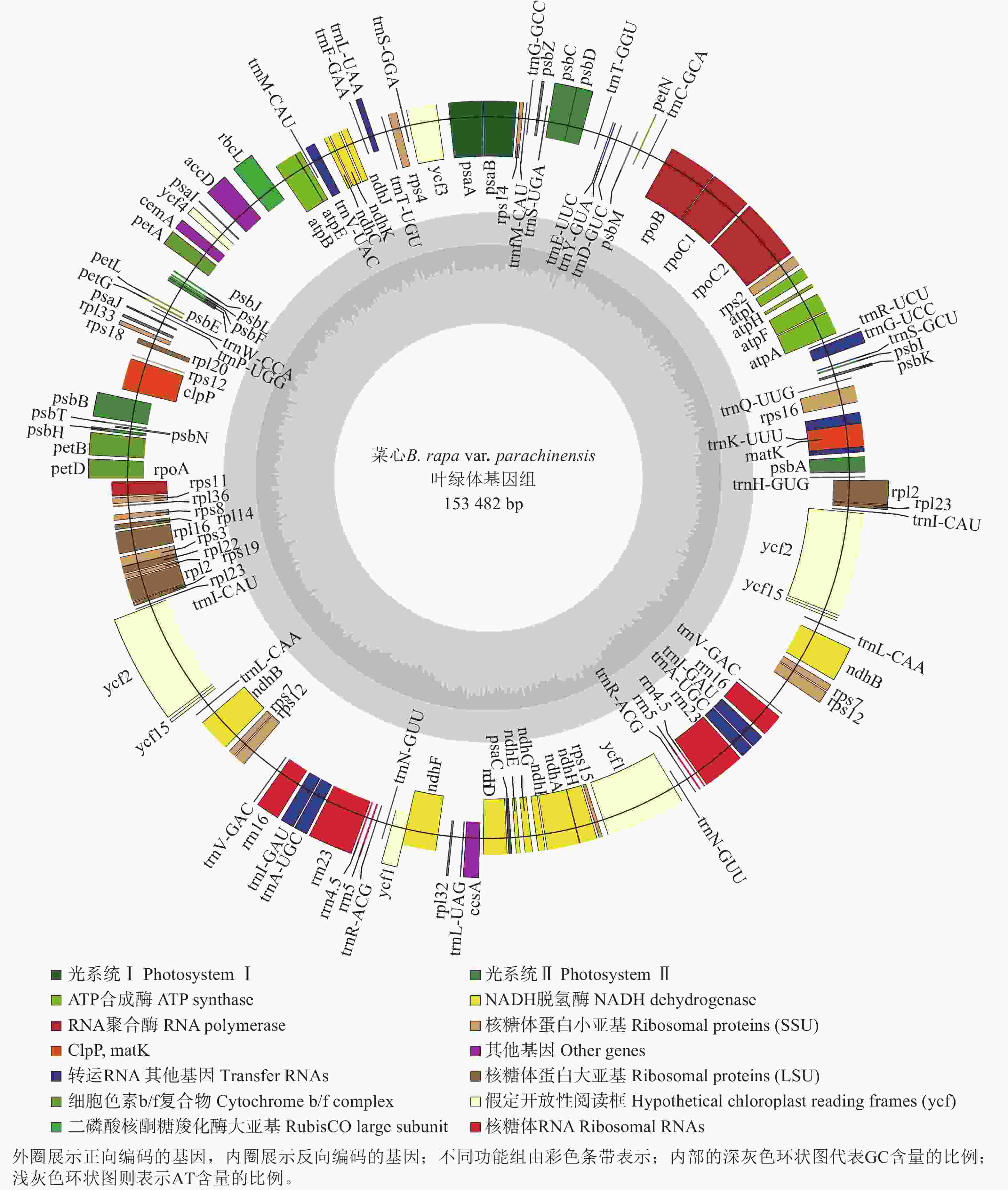
图 1 菜心叶绿体基因组图谱
Figure 1. Map of B. rapa var. parachinensis chloroplast genome
表 1 菜心叶绿体基因组特征
Table 1. Chloroplast genome characteristics of B. rapa var. parachinensis
区域 A含量/% C含量/% G含量/% T含量/% GC含量/% 碱基长度/bp LSC 32.05 17.54 16.58 33.83 34.12 83 281 SSC 35.49 15.16 14.04 35.31 29.20 17 775 IRa 28.91 22.03 20.31 28.76 42.34 26 213 IRb 28.76 20.31 22.03 28.91 42.34 26 213 总量 31.35 18.51 17.85 32.29 36.36 153 482 -
在菜心的叶绿体基因组中,共发现了132个基因,包括86个蛋白质编码基因、37个tRNA基因、8个rRNA基因,以及1个假基因。这些基因的主要功能涉及光合作用和自我复制等过程,此外还有部分基因的功能尚未明确(表2)。在菜心叶绿体基因组中,72个蛋白质编码基因、23个tRNA的拷贝数为1,7个蛋白质编码基因、7个tRNA、4个rRNA的拷贝数为2;11个蛋白质编码基因、8个tRNA的内含子数量为1,4个蛋白质编码基因的内含子数量为2 (表2)。
表 2 菜心叶绿体基因组编码基因注释信息
Table 2. Gene annotation of the chloroplast genome of B. rapa var. parachinensis
基因分类 基因分组 基因列表 光合作用 光系统Ⅰ亚基 psaA、psaB、psaC、psaI、psaJ 光系统Ⅱ亚基 psbA、psbB、psbC、psbD、psbE、psbF、psbH、psbI、psbJ、psbK、psbL、psbM、psbN、psbT、psbZ NADH脱氢酶亚基 ndhA*、ndhB*(2)、ndhC、ndhD、ndhE、ndhF、ndhG、ndhH、ndhI、ndhJ、ndhK 色素细胞b/f复合物亚基 petA、petB*、petD*、petG、petL、petN ATP合成酶亚基 atpA、atpB、atpE、atpF*、atpH、atpI Rubisco大亚基 rbcL 复制基因 核糖体蛋白大亚基 rpl14、rpl16*、rpl2*(2)、rpl20、rpl22、rpl23(2)、rpl32、rpl33、rpl36 核糖体蛋白小亚基 rps11、rps12**(2)、rps14、rps15、rps16*、rps18、rps19、rps2、rps3、rps4、rps7(2)、rps8 RNA聚合酶亚基 rpoA、rpoB、rpoC1*、rpoC2 核糖体RNAs rrn16(2)、rrn23(2)、rrn4.5(2)、rrn5(2) 转运RNAs trnA-UGC*(2)、trnC-GCA、trnD-GUC、trnE-UUC、trnF-GAA、trnG-GCC、trnG-UCC*、trnH-GUG、trnI-CAU(2)、trnI-GAU*(2)、trnK-UUU*、trnL-CAA(2)、trnL-UAA*、trnL-UAG、trnM-CAU、trnN-GUU(2)、trnP-UGG、trnQ-UUG、trnR-ACG(2)、trnR-UCU、trnS-GCU、trnS-GGA、trnS-UGA、trnT-GGU、trnT-UGU、trnV-GAC(2)、trnV-UAC*、trnW-CCA、trnY-GUA、trnfM-CAU 其他基因 成熟酶 matK 蛋白酶 clpP** 被膜蛋白 cemA 乙酰辅酶A羧化酶 accD 色素细胞C合成酶 ccsA 未知功能基因 保守的开放阅读框 #ycf1、ycf1、ycf15(2)、ycf2(2)、ycf3**、ycf4 说明:(2)代表多拷贝基因;*表示带1个内含子的基因;**表示带2个内含子的基因;#表示假基因。 -
分析菜心叶绿体基因组发现:总计22 799个密码子被用于编码氨基酸,其中编码亮氨酸(Leu)的密码子最为常用,数量高达2 421个;其次为编码异亮氨酸(Ile)和丝氨酸(Ser)的密码子,数量分别为1 980和1 699个(表3)。对其同义密码子使用度(relative synonymous codon usage, RSCU)进行分析,发现RSCU大于1的密码子有31个,其中以A或者U结尾的有29个;RSCU小于1的密码子有34个,其中以G或者C结尾的有31个;编码色氨酸(Trp)的密码子只有1个(UGG),因此其RSCU等于1;AUG密码子不仅是起始密码子,还编码甲硫氨酸(Met),因此RSCU最高,为6.97;其次为编码Leu的UUA和编码丙氨酸(Ala)的GCU,RSCU分别为2.26和1.89;RSCU最低的为编码Met的GUG和UUG,为0.01 (表3)。
表 3 菜心叶绿体基因组同义密码子使用度(RSCU)分析
Table 3. Relative synonymous codon usage (RSCU) analysis of B. rapa var. parachinensis
氨基酸 密码子 数量 RSCU 氨基酸 密码子 数量 RSCU 氨基酸 密码子 数量 RSCU Ter UAA 48 1.82 Ile AUC 350 0.53 Arg AGG 126 0.57 Ter UAG 19 0.72 Ile AUU 1005 1.52 Arg CGA 296 1.35 Ter UGA 12 0.46 Lys AAA 997 1.57 Arg CGC 96 0.44 Ala GCA 348 1.12 Lys AAG 276 0.43 Arg CGG 102 0.47 Ala GCC 177 0.57 Leu CUA 323 0.80 Arg CGU 311 1.42 Ala GCG 131 0.42 Leu CUC 149 0.37 Ser AGC 100 0.35 Ala GCU 586 1.89 Leu CUG 139 0.34 Ser AGU 358 1.26 Cys UGC 62 0.46 Leu CUU 498 1.23 Ser UCA 340 1.20 Cys UGU 207 1.54 Leu UUA 872 2.16 Ser UCC 235 0.83 Asp GAC 170 0.39 Leu UUG 440 1.09 Ser UCG 156 0.55 Asp GAU 698 1.61 Met AUG 512 6.97 Ser UCU 510 1.80 Glu GAA 928 1.55 Met GUG 1 0.01 Thr ACA 367 1.23 Glu GAG 270 0.45 Met UUG 1 0.01 Thr ACC 212 0.71 Phe UUC 401 0.59 Asn AAC 244 0.44 Thr ACG 123 0.41 Phe UUU 967 1.41 Asn AAU 855 1.56 Thr ACU 490 1.64 Gly GGA 624 1.61 Pro CCA 266 1.16 Val GUA 455 1.45 Gly GGC 153 0.39 Pro CCC 161 0.70 Val GUC 153 0.49 Gly GGG 254 0.66 Pro CCG 124 0.54 Val GUG 169 0.54 Gly GGU 519 1.34 Pro CCU 366 1.60 Val GUU 475 1.52 His CAC 125 0.49 Gln CAA 642 1.57 Trp UGG 396 1.00 His CAU 383 1.51 Gln CAG 175 0.43 Tyr UAC 155 0.37 Ile AUA 625 0.95 Arg AGA 384 1.75 Tyr UAU 687 1.63 -
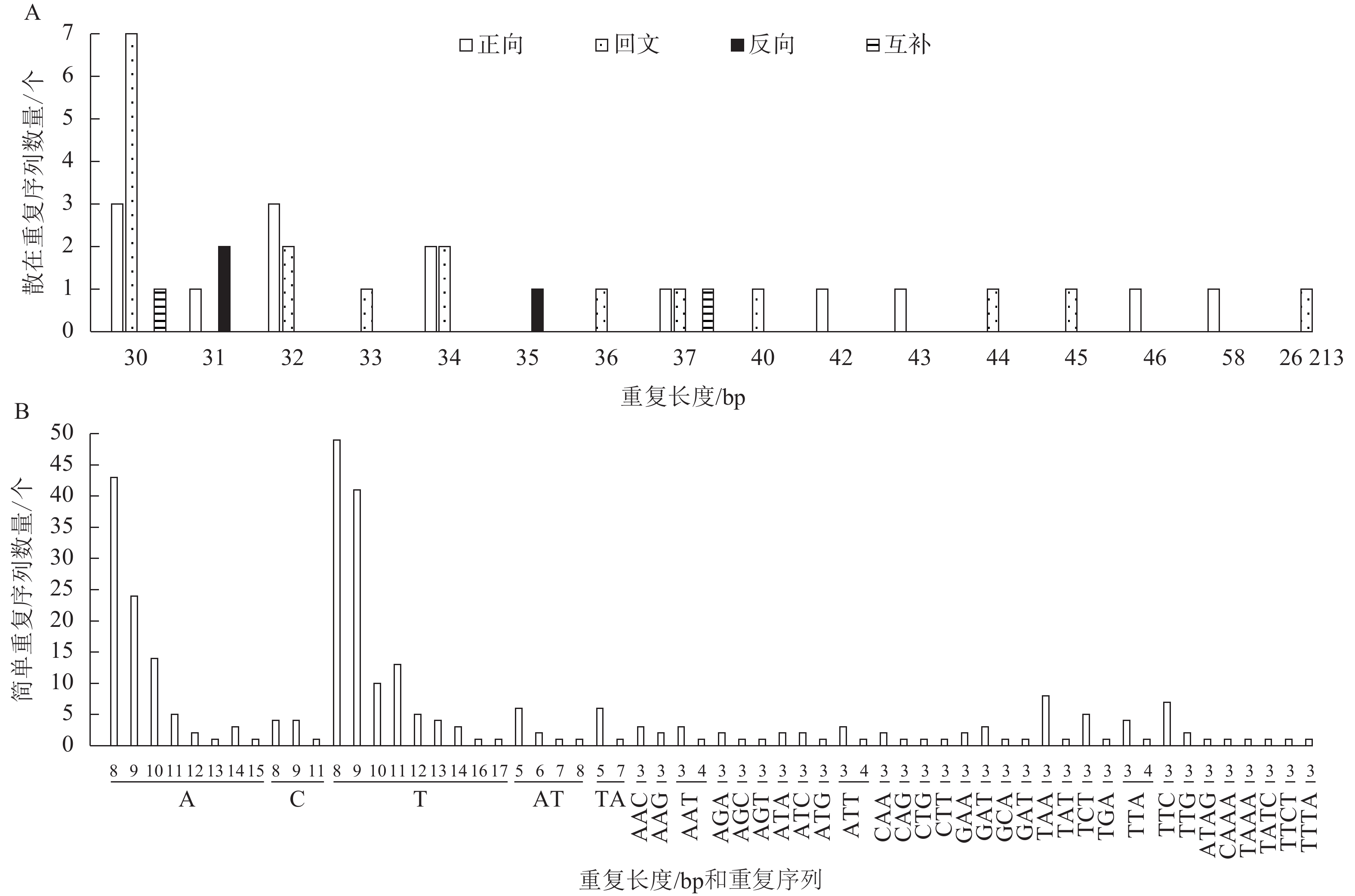
散在重复序列在基因组里以分散的形式存在。菜心叶绿体基因组中包括正向重复14个、回文重复18个、反向重复3个、互补重复2个共37个散在重复序列。其中36个散在重复序列长度为30~58 bp,长度为30 bp的散在重复序列数量最多,共有11个,其次是长度为32 bp的散在重复序列,共有5个;此外还有1个最长的散在重复序列,长度为26 213 bp (图2A)。
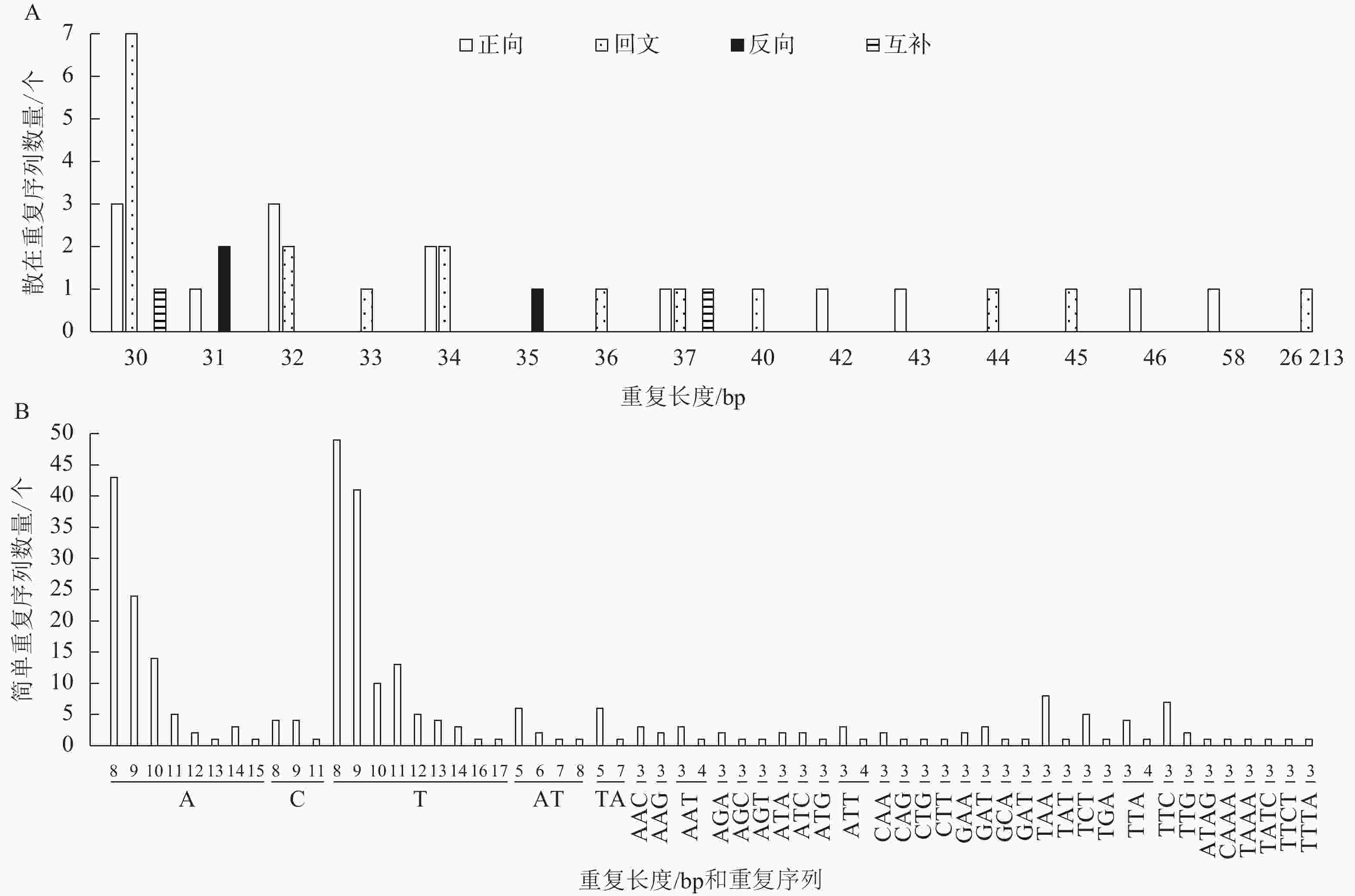
图 2 菜心叶绿体基因组散在重复序列(A)和简单重复序列(B)分析
Figure 2. Analysis of scattered sequence repeats (A) and simple sequence repeats (B) in the chloroplast genome of B. rapa var. parachinensis
SSRs是由1~6个核苷酸组成的短序列作为重复单元串联而成的一类序列。在菜心叶绿体基因组中共存在315个SSRs,其中LSC含有197个,SSC含有72个,IR含有46个;在这些SSRs中,单核苷酸重复、双核苷酸重复、3核苷酸重复和4核苷酸重复的数量分别为229、17、63和6个。进一步分析显示:在全部315个SSRs中,占比排名前3位的类型依次为T(8)、A(8)、T(9),分别为15.56% (49)、13.65% (43)、13.0% (41)(图2B)。
-
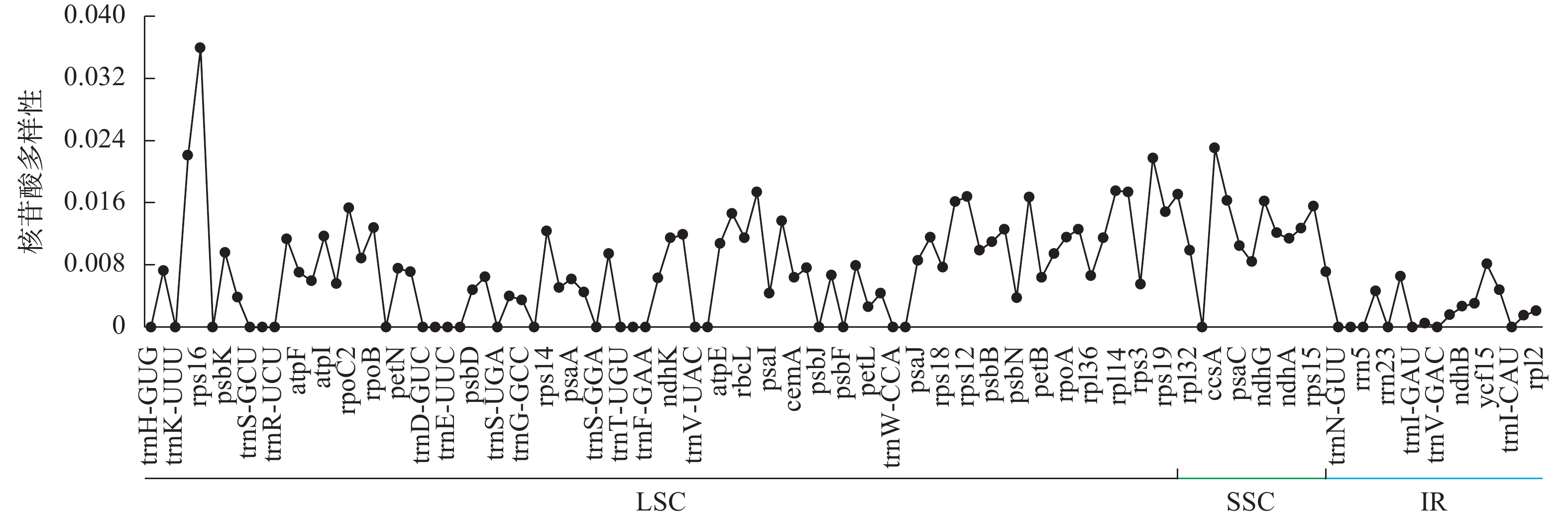
菜心、拟南芥、芥菜等8种十字花科植物的叶绿体基因组平均核苷酸多样性为0.007 2 (图3)。SSC区域的平均核苷酸多样性最高(0.012 3),LSC次之(0.007 4),IR区域的核苷酸多样性最低(0.002 1)。利用核苷酸多样性筛选出4个高变异区域(Pi≥0.02),发现LSC区域含有3个位点,分别为rps16、matK、rpl22,SSC区域含有1个位点,为ccsA,其中变异度最高的位点为rps16。
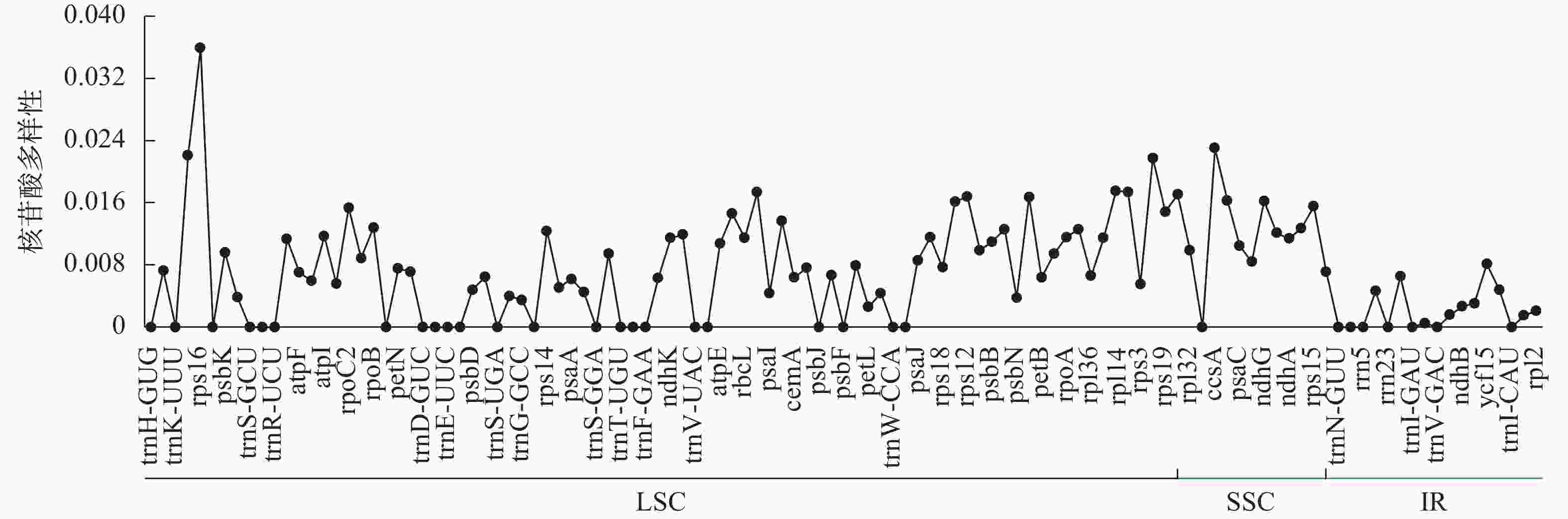
图 3 基因核苷酸多样性折线图
Figure 3. Line chart of gene nucleotide diversity value
在植物叶绿体基因组的进化过程中,导致其大小差异的主要因素是IR边界的扩张与收缩。边界分析结果表明:菜心等8种十字花科植物的叶绿体基因组均存在4个边界,即JLB (LSC/IRb)、JSB (IRb/SSC)、JSA (SSC/IRa)、JLA (IRa/LSC)。rps19、rpl2、ycf1、ndhF、trnN和trnH是位于IR边界附近的主要基因。在这8种十字花科植物中,JLB边界均位于rps19基因编码区内,且在不同植物叶绿体基因组中仅存在1~2 bp的位置差异;JSB边界均位于ycf1和ndhF基因编码区内,两者有36~37 bp的重叠区域,其中ycf1基因的大部分区域位于IRb,只有2~3 bp位于SSC;JSA边界均位于ycf1基因编码区内,其中
1027 ~1030 bp位于IRa,4 271~4 358 bp位于SSC;trnH-GUG均位于LSC,距离JLA边界2~30 bp (图4)。以上结果显示菜心等8种十字花科植物叶绿体基因组高度保守,IR边界总体变化幅度较小,仅涉及少数基因。
图 4 8种十字花科植物的IR/SC边界分析
Figure 4. IR/SC boundary analysis of 8 Brassicaceae crops
-
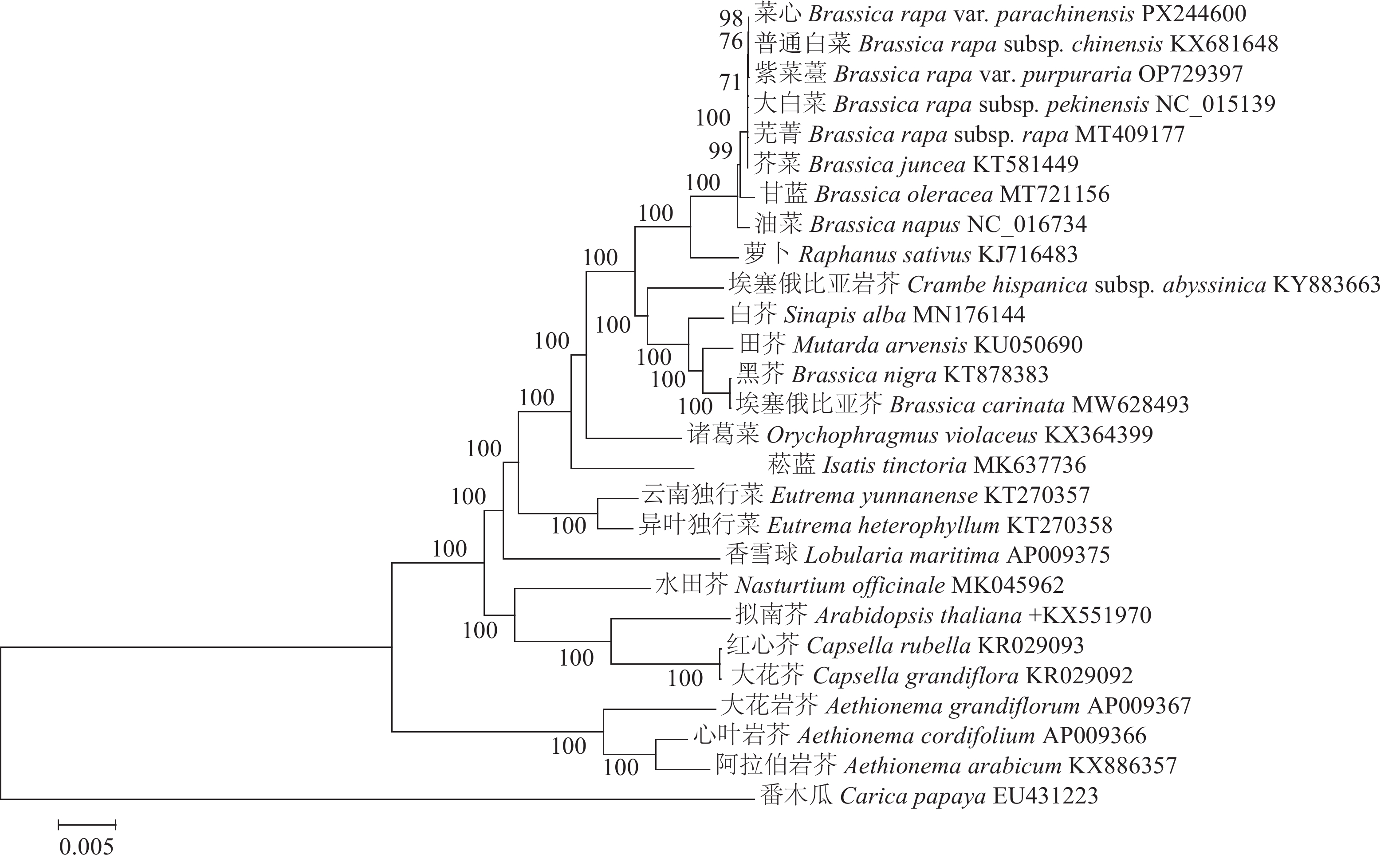
为了更全面地了解十字花科植物之间的进化关系,从NCBI数据库中下载了25种十字花科植物的叶绿体基因组数据。在此基础上,以番木瓜作为外类群构建系统发育树。结果显示:与菜心亲缘关系最近的为普通白菜B. rapa subsp. chinensis,其次为紫菜薹、大白菜B. rapa subsp. pekinensis、芜菁B. rapa subsp. rapa、芥菜、甘蓝、油菜等芸薹属作物;萝卜与菜心的亲缘关系紧随其后,表明在十字花科植物中,萝卜是与菜心等芸薹属作物进化上接近的外属作物,反之,同为芸薹属的黑芥B. nigra和埃塞俄比亚芥B. carinata与菜心的亲缘关系则相对较远。番木瓜作为外类群,与菜心的亲缘关系最远(图5)。
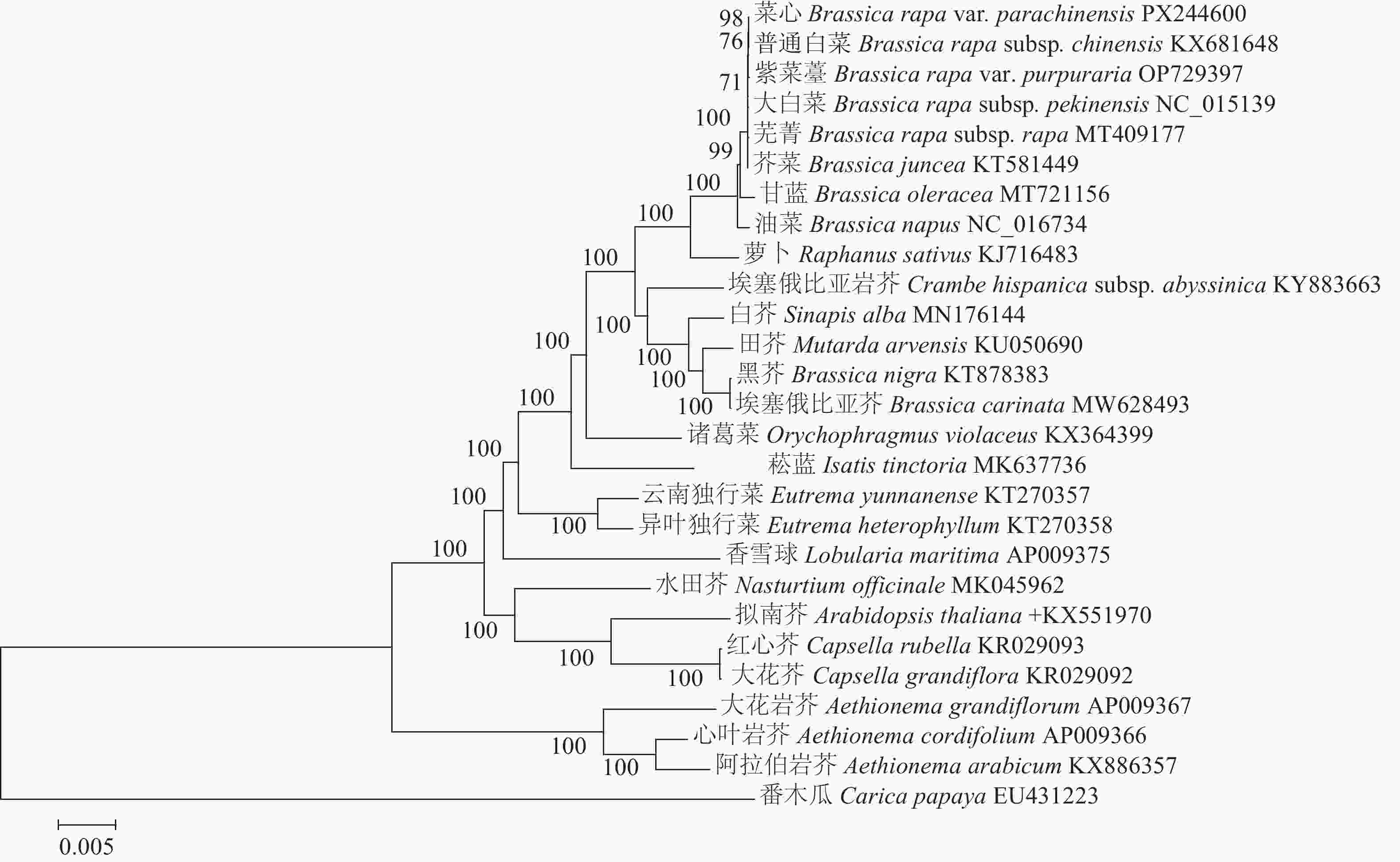
图 5 基于叶绿体基因组序列构建的系统发育树
Figure 5. Phylogenetic tree constructed based on chloroplast genome sequences
-
叶绿体是植物遗传系统中的半自主性细胞器,其基因组的基因数量、基因组成和基因排列比线粒体基因组和核基因组更为保守[27]。本研究利用高通量测序分析了菜心的叶绿体基因组,发现菜心叶绿体基因组呈典型的四分体结构,全长为153 482 bp,GC含量为36.36%,与白菜类蔬菜中的普通白菜、大白菜、紫菜薹的叶绿体基因组长度和GC含量一致;比芜菁叶绿体基因组短139 bp,GC含量高了0.02%[13];与芸薹属中的芥菜、黑芥、埃塞俄比亚芥等作物以及模式植物拟南芥的叶绿体基因组长度和GC含量差距也不明显[12, 14, 28−29]。由此可见,在白菜及芸薹属内,各个物种的叶绿体基因组相对保守,这个结果与ZHAO等[30]的研究结果一致。
菜心叶绿体基因组共鉴定出132个基因,其中45个基因与光合作用相关,与已发表的芸薹属物种相比,其基因种类和数量未表现出显著差异,表明叶绿体基因组核心功能基因在进化过程中高度保守[31]。在植物的进化历程中,不同密码子的使用通常显示出一定的偏好性。在菜心叶绿体基因组中,使用频率最高的氨基酸为Leu,使用频率最低的氨基酸为半胱氨酸(Cys),这与凤梨Ananas comosus var. comosus[32]、宝华玉兰Magnolia zenii[33]等植物的研究结果一致。相对同义密码子使用频率(RSCU)是通过比较某个特定密码子的实际出现频率与其理论预期频率来定义的,是评估密码子偏好性的有效工具,RSCU大于1代表该密码子的使用具有明显偏好[34]。在菜心叶绿体基因组中,绝大多数(93.55%) RSCU大于1的密码子以A或者U结尾,绝大多数(86.84%) RSCU小于1的密码子以G或者C结尾,类似现象在被子植物的叶绿体基因组中普遍存在,说明叶绿体基因组的密码子使用频率也有高度的保守性[35−36]。SSRs被广泛用作构建遗传连锁图谱、种群遗传分析等。ZHAO等[30]在苤蓝B. oleracea var. gongylodes叶绿体基因组中共鉴定出290个SSRs,WU等[37]在冬季油菜中发现了288个SSRs,本研究在菜心叶绿体基因组中共发现315个SSRs,可为研究芸薹属作物的遗传多样性提供潜在候选分子标记。
核苷酸多样性是用来衡量一个群体内遗传变异程度的重要指标,其值越高说明群体内的遗传多样性越丰富,能够为种群遗传学提供潜在的分子标记[38]。本研究利用菜心、拟南芥等8种十字花科植物进行比较分析,发现8个物种叶绿体基因组的平均核苷酸多样性为0.007 2,不同区域平均核苷酸多样性从大到小依次为LSC (0.007 4)、SSC (0.012 3)、IR (0.002 1),说明IR相比其他2个区域更为保守;其中核苷酸多样性最高的前4个位点依次为位于LSC区域的rps16,SSC区域的ccsA,LSC区域的matK、rpl22,这些高度变异位点可以用作十字花科物种鉴定的分子标记。在植物基因组进化过程中,IR区的扩张或收缩是叶绿体基因组结构变异的主要驱动力,可为物种鉴定和系统发育研究提供分子依据[39]。对包括菜心在内的8个十字花科物种的IR边界进行分析,发现IR边界的差异主要跟rps19、ycf1、ndhF、trnH的位置有关,但整体变化较小,说明菜心等十字花科植物的叶绿体基因组比较保守。
基于叶绿体基因组,利用最大似然法对菜心进行系统发育分析,为菜心在十字花科中的进化位置提供了依据。本研究结果显示:菜心与普通白菜、紫菜薹、大白菜、芜菁等白菜类蔬菜亲缘关系最近,其次为芥菜、甘蓝、油菜等芸薹属作物,而同为芸薹属的黑芥和埃塞俄比亚芥与菜心的亲缘关系则较远。WANG等[31]基于叶绿体基因组对芥蓝B. oleracea var. alboglabra等十字花科作物进行系统发育分析也发现了同样的现象,芥蓝与油菜、白菜、芥菜、甘蓝聚为一个类群,黑芥和埃塞俄比亚芥聚为另一个类群。
-
菜心叶绿体基因组呈典型的四分体结构,全长为153 482 bp,包含86个蛋白质编码基因、37个tRNA基因和8个rRNA基因,基因功能主要为光合作用、自我复制等;其基因组密码子多以A/U结尾,共有37个散在重复序列,315个SSR;系统发育分析结果显示:普通白菜与菜心的亲缘关系最近。本研究有助于深入理解菜心的系统发育关系,可为菜心及其他十字花科植物的遗传改良、育种策略等提供理论依据。
Characteristics of the chloroplast genome and phylogenetic analysis of flowering Chinese cabbage
-
摘要:
目的 对菜心Brassica rapa var. parachinensis叶绿体基因组进行组装、注释和分析,揭示其与其他十字花科Brassicaceae植物的系统进化关系。 方法 利用Illumina NovaSeq 6000平台对菜心叶绿体全基因组测序,利用生物信息学的手段分析其叶绿体基因组结构、重复序列、核苷酸多样性、密码子偏好性,以及结合其他25种十字花科植物叶绿体基因组序列构建系统发育关系。 结果 菜心叶绿体基因组全长为153 482 bp,GC含量为36.36%,是典型的四分体结构;共注释到132个编码基因,其中86个蛋白质编码基因、37个tRNA基因、8个rRNA基因,以及1个假基因;密码子偏好性分析结果显示:亮氨酸的使用频率最高,相对同义密码子使用度大于1的密码子有31个,多以A或者U结尾;共有37个散在重复序列,315个简单重复序列,其中单核苷酸重复占比最高,为72.70%;小单拷贝区的核苷酸多样性最高,反向重复区的核苷酸多样性最低;系统发育分析结果显示:菜心与其他白菜类蔬菜亲缘关系最近。 结论 菜心叶绿体基因组为典型的四分体结构,相对保守,与普通白菜B. rapa var. chinensis、紫菜薹B. rapa var. purpuraria、大白菜B. rapa subsp. pekinensis等亲缘关系密切。图5表3参39 Abstract:Objective This study aims to assemble, annotate, and analyze the chloroplast genome of flowering Chinese cabbage (Brassica rapa var. parachinensis), and to elucidate its phylogenetic relationships with other Brassicaceae crops. Method The complete chloroplast genome of flowering Chinese cabbage was sequenced using the Illumina NovaSeq 6000 platform. Bioinformatics approaches were employed to analyze its genome structure, repetitive sequences, nucleotide diversity, codon usage bias, and phylogenetic relationships by constructing phylogenetic trees with chloroplast genome sequences of other 25 Brassicaceae species. Result The chloroplast genome of flowering Chinese cabbage is 153 482 bp in length with a GC content of 36.36%, exhibiting a typical quadripartite structure. In total, 132 functional genes had been identified and annotated, comprising 86 protein-coding genes, 37 tRNA genes, 8 rRNA genes, and 1 pseudogene. Codon usage bias analysis revealed that leucine (Leu) is the most frequently used amino acid, with 31 codons showing a relative synonymous codon usage (RSCU) value greater than 1, predominantly ending with A or U. Repetitive sequence analysis detected 37 dispersed repeats and 315 simple sequence repeats (SSRs), with mononucleotide repeats predominating (72.70% of total SSRs). The nucleotide diversity of small single copy region (SSC) is the highest, while that of inverted repeat region (IR) is the lowest. Phylogenetic analysis revealed that flowering Chinese cabbage exhibits the closest genetic relationship with leafy Chinese cabbage varieties. Conclusion The chloroplast genome of flowering Chinese cabbage exhibits a conserved quadripartite structure and demonstrates close phylogenetic relationships with B. rapa var. chinensis, B. rapa var. purpuraria, and B. rapa subsp. pekinensis. [Ch, 5 fig. 3 tab. 39 ref.] -
Key words:
- flowering Chinese cabbage /
- chloroplast genome /
- sequence feature /
- systematic evolution
-
图 2 菜心叶绿体基因组散在重复序列(A)和简单重复序列(B)分析
Figure 2 Analysis of scattered sequence repeats (A) and simple sequence repeats (B) in the chloroplast genome of B. rapa var. parachinensis
图 5 基于叶绿体基因组序列构建的系统发育树
Figure 5 Phylogenetic tree constructed based on chloroplast genome sequences
表 1 菜心叶绿体基因组特征
Table 1. Chloroplast genome characteristics of B. rapa var. parachinensis
区域 A含量/% C含量/% G含量/% T含量/% GC含量/% 碱基长度/bp LSC 32.05 17.54 16.58 33.83 34.12 83 281 SSC 35.49 15.16 14.04 35.31 29.20 17 775 IRa 28.91 22.03 20.31 28.76 42.34 26 213 IRb 28.76 20.31 22.03 28.91 42.34 26 213 总量 31.35 18.51 17.85 32.29 36.36 153 482  下载: 导出CSV
下载: 导出CSV
表 2 菜心叶绿体基因组编码基因注释信息
Table 2. Gene annotation of the chloroplast genome of B. rapa var. parachinensis
基因分类 基因分组 基因列表 光合作用 光系统Ⅰ亚基 psaA、psaB、psaC、psaI、psaJ 光系统Ⅱ亚基 psbA、psbB、psbC、psbD、psbE、psbF、psbH、psbI、psbJ、psbK、psbL、psbM、psbN、psbT、psbZ NADH脱氢酶亚基 ndhA*、ndhB*(2)、ndhC、ndhD、ndhE、ndhF、ndhG、ndhH、ndhI、ndhJ、ndhK 色素细胞b/f复合物亚基 petA、petB*、petD*、petG、petL、petN ATP合成酶亚基 atpA、atpB、atpE、atpF*、atpH、atpI Rubisco大亚基 rbcL 复制基因 核糖体蛋白大亚基 rpl14、rpl16*、rpl2*(2)、rpl20、rpl22、rpl23(2)、rpl32、rpl33、rpl36 核糖体蛋白小亚基 rps11、rps12**(2)、rps14、rps15、rps16*、rps18、rps19、rps2、rps3、rps4、rps7(2)、rps8 RNA聚合酶亚基 rpoA、rpoB、rpoC1*、rpoC2 核糖体RNAs rrn16(2)、rrn23(2)、rrn4.5(2)、rrn5(2) 转运RNAs trnA-UGC*(2)、trnC-GCA、trnD-GUC、trnE-UUC、trnF-GAA、trnG-GCC、trnG-UCC*、trnH-GUG、trnI-CAU(2)、trnI-GAU*(2)、trnK-UUU*、trnL-CAA(2)、trnL-UAA*、trnL-UAG、trnM-CAU、trnN-GUU(2)、trnP-UGG、trnQ-UUG、trnR-ACG(2)、trnR-UCU、trnS-GCU、trnS-GGA、trnS-UGA、trnT-GGU、trnT-UGU、trnV-GAC(2)、trnV-UAC*、trnW-CCA、trnY-GUA、trnfM-CAU 其他基因 成熟酶 matK 蛋白酶 clpP** 被膜蛋白 cemA 乙酰辅酶A羧化酶 accD 色素细胞C合成酶 ccsA 未知功能基因 保守的开放阅读框 #ycf1、ycf1、ycf15(2)、ycf2(2)、ycf3**、ycf4 说明:(2)代表多拷贝基因;*表示带1个内含子的基因;**表示带2个内含子的基因;#表示假基因。
下载: 导出CSV
表 3 菜心叶绿体基因组同义密码子使用度(RSCU)分析
Table 3. Relative synonymous codon usage (RSCU) analysis of B. rapa var. parachinensis
氨基酸 密码子 数量 RSCU 氨基酸 密码子 数量 RSCU 氨基酸 密码子 数量 RSCU Ter UAA 48 1.82 Ile AUC 350 0.53 Arg AGG 126 0.57 Ter UAG 19 0.72 Ile AUU 1005 1.52 Arg CGA 296 1.35 Ter UGA 12 0.46 Lys AAA 997 1.57 Arg CGC 96 0.44 Ala GCA 348 1.12 Lys AAG 276 0.43 Arg CGG 102 0.47 Ala GCC 177 0.57 Leu CUA 323 0.80 Arg CGU 311 1.42 Ala GCG 131 0.42 Leu CUC 149 0.37 Ser AGC 100 0.35 Ala GCU 586 1.89 Leu CUG 139 0.34 Ser AGU 358 1.26 Cys UGC 62 0.46 Leu CUU 498 1.23 Ser UCA 340 1.20 Cys UGU 207 1.54 Leu UUA 872 2.16 Ser UCC 235 0.83 Asp GAC 170 0.39 Leu UUG 440 1.09 Ser UCG 156 0.55 Asp GAU 698 1.61 Met AUG 512 6.97 Ser UCU 510 1.80 Glu GAA 928 1.55 Met GUG 1 0.01 Thr ACA 367 1.23 Glu GAG 270 0.45 Met UUG 1 0.01 Thr ACC 212 0.71 Phe UUC 401 0.59 Asn AAC 244 0.44 Thr ACG 123 0.41 Phe UUU 967 1.41 Asn AAU 855 1.56 Thr ACU 490 1.64 Gly GGA 624 1.61 Pro CCA 266 1.16 Val GUA 455 1.45 Gly GGC 153 0.39 Pro CCC 161 0.70 Val GUC 153 0.49 Gly GGG 254 0.66 Pro CCG 124 0.54 Val GUG 169 0.54 Gly GGU 519 1.34 Pro CCU 366 1.60 Val GUU 475 1.52 His CAC 125 0.49 Gln CAA 642 1.57 Trp UGG 396 1.00 His CAU 383 1.51 Gln CAG 175 0.43 Tyr UAC 155 0.37 Ile AUA 625 0.95 Arg AGA 384 1.75 Tyr UAU 687 1.63
下载: 导出CSV
-
[1] 熊瑶, 童凌斐, 曹岚, 等. 4种忍冬属植物叶绿体基因组结构特征及系统发育分析[J]. 药学学报, 2024, 59(11): 3164−3171. XIONG Yao, TONG Lingfei, CAO Lan, et al. Structural characteristics and phylogenetic analysis of chloroplast genomes of four species of Lonicera[J]. Acta Pharmaceutica Sinica, 2024, 59(11): 3164−3171. DOI: 10.16438/j.0513-4870.2024-0306. [2] YIN Dongmei, WANG Yun, ZHANG Xingguo, et al. Development of chloroplast genome resources for peanut (Arachis hypogaea L.) and other species of Arachis[J]. Scientific Reports, 2017, 7: 11649. DOI: 10.1038/s41598-017-12026-x. [3] 温静, 郭茂伟, 朱琳, 等. 黄花型扁蓿豆叶绿体基因组及其系统发育分析[J]. 中国草地学报, 2024, 46(8): 10−18. WEN Jing, GUO Maowei, ZHU Lin, et al. Analysis of chloroplast genome and phylogeny of yellow flowered type Medicago ruthenica[J]. Chinese Journal of Grassland, 2024, 46(8): 10−18. DOI: 10.16742/j.zgcdxb.20230367. [4] SVEINSSON S, CRONK Q. Evolutionary origin of highly repetitive plastid genomes within the clover genus (Trifolium)[J]. BMC Evolutionary Biology, 2014, 14(1): 228. DOI: 10.1186/s12862-014-0228-6. [5] CAI Chuner, WANG Lingke, ZHOU Lingjie, et al. Complete chloroplast genome of green tide algae Ulva flexuosa (Ulvophyceae, Chlorophyta) with comparative analysis[J]. PLoS One, 2017, 12(9): e0184196. DOI: 10.1371/journal.pone.0184196. [6] DANIELL H, LIN C S, YU Ming, et al. Chloroplast genomes: diversity, evolution, and applications in genetic engineering[J]. Genome Biology, 2016, 17(1): 134. DOI: 10.1186/s13059-016-1004-2. [7] LIU Liangmin, DU Xinyu, GUO Cen, et al. Resolving robust phylogenetic relationships of core Brassicaceae using genome skimming data[J]. Journal of Systematics and Evolution, 2021, 59(3): 442−453. DOI: 10.1111/jse.12666. [8] YANG Jinghua, LIU Dongyuan, WANG Xiaowu, et al. The genome sequence of allopolyploid Brassica juncea and analysis of differential homoeolog gene expression influencing selection[J]. Nature Genetics, 2016, 48(10): 1225−1232. DOI: 10.1038/ng.3657. [9] 李晓晴, 杨佳雨, 陈少明, 等. 氮肥与生物炭、有机肥配施对菜心生长及养分积累的影响[J]. 植物营养与肥料学报, 2024, 30(11): 2150−2160. LI Xiaoqing, YANG Jiayu, CHEN Shaoming, et al. Effects of nitrogen fertilizer combined with biochar or organic fertilizer on the growth and nutrient accumulation of Chinese flowering cabbage [Brassica campestris L. ssp. chinensis (L.) var. utilis Tsen et Lee][J]. Journal of Plant Nutrition and Fertilizers, 2024, 30(11): 2150−2160. DOI: 10.11674/zwyf.2024221. [10] 原远, 王春政, 周贤玉, 等. 六种菜心氨基酸组成及营养价值评价[J]. 南方农业学报, 2019, 50(10): 2271−2277. YUAN Yuan, WANG Chunzheng, ZHOU Xianyu, et al. Amino acid composition and nutritional value evaluation of six kinds of flowering Chinese cabbages (Brassica campestris L. ssp. chinensis var. utilis Tsen et Lee)[J]. Journal of Southern Agriculture, 2019, 50(10): 2271−2277. DOI: 10.3969/j.issn.2095-1191.2019.10.17. [11] 朱云娜, 杨庆庆, 寇尔丰, 等. 水杨酸处理对菜心生长发育及其品质的影响[J]. 北方园艺, 2020(22): 53−59. ZHU Yunna, YANG Qingqing, KOU Erfeng, et al. Effects of salicylic acid on plant growth development and qualities in flowering Chinese cabbage[J]. Northern Horticulture, 2020(22): 53−59. DOI: 10.11937/bfyy. 20200707. [12] SATO S, NAKAMURA Y, KANEKO T, et al. Complete structure of the chloroplast genome of Arabidopsis thaliana[J]. DNA Research, 1999, 6(5): 283−290. DOI: 10.1093/dnares/6.5.283. [13] 王传之, 周贤玉, 李扬眉, 等. 紫菜薹叶绿体全基因组序列及其系统发育分析[J]. 西北农业学报, 2024, 33(8): 1483−1494. WANG Chuanzhi, ZHOU Xianyu, LI Yangmei, et al. Chloroplast genome sequence and phylogenetic analysis of Brassica rapa var. purpuraria[J]. Acta Agriculturae Boreali-occidentalis Sinica, 2024, 33(8): 1483−1494. DOI: 10.7606/j.issn.1004-1389.2024.08.009. [14] PRABHUDAS S K, RAJU B, KANNAN THODI S, et al. The complete chloroplast genome sequence of Indian mustard (Brassica juncea L.)[J]. Mitochondrial DNA Part A, 2016, 27(6): 4622−4623. DOI: 10.3109/19401736.2015.1101586. [15] JEONG Y M, CHUNG W H, MUN J H, et al. De novo assembly and characterization of the complete chloroplast genome of radish (Raphanus sativus L. )[J]. Gene, 2014, 551(1): 39−48. DOI: 10.1016/j.gene.2014.08.038. [16] CHEN Shifu, ZHOU Yanqing, CHEN Yaru, et al. Fastp: an ultra-fast all-in-one FASTQ preprocessor[J]. Bioinformatics, 2018, 34(17): i884−i890. DOI: 10.1093/bioinformatics/bty560. [17] ANTON B, SERGEY N, DMITRY A , et al. SPAdes: a new genome assembly algorithm and its applications to single-cell sequencing[J]. Journal of Computational Biology, 2012, 19(5): 455–477. DOI: 10.1089/cmb.2012.0021. [18] HYATT D, CHEN G L, LoCASCIO P F, et al. Prodigal: prokaryotic gene recognition and translation initiation site identification[J]. BMC Bioinformatics, 2010, 11(1): 119. DOI: 10.1186/1471-2105-11-119. [19] MISTRY J, FINN R D, EDDY S R, et al. Challenges in homology search: HMMER3 and convergent evolution of coiled-coil regions[J]. Nucleic Acids Research, 2013, 41(12): e121. DOI: 10.1093/nar/gkt263. [20] LASLETT D, CANBACK B. ARAGORN a program to detect tRNA genes and tmRNA genes in nucleotide sequences[J]. Nucleic Acids Research, 2004, 32(1): 11−16. DOI: 10.1093/nar/gkh152. [21] GREINER S, LEHWARK P, BOCK R. OrganellarGenomeDRAW (OGDRAW) version 1.3.1: expanded toolkit for the graphical visualization of organellar genomes[J]. Nucleic Acids Research, 2019, 47(W1): W59−W64. DOI: 10.1093/nar/gkz238. [22] BEIER S, THIEL T, MÜNCH T, et al. MISA-web: a web server for microsatellite prediction[J]. Bioinformatics, 2017, 33(16): 2583−2585. DOI: 10.1093/bioinformatics/btx198. [23] NAKAMURA T, YAMADA K D, TOMII K, et al. Parallelization of MAFFT for large-scale multiple sequence alignments[J]. Bioinformatics, 2018, 34(14): 2490−2492. DOI: 10.1093/bioinformatics/bty121. [24] LIBRADO P, ROZAS J. DnaSP v5: a software for comprehensive analysis of DNA polymorphism data[J]. Bioinformatics, 2009, 25(11): 1451−1452. DOI: 10.1093/bioinformatics/btp187. [25] DARLING A C E, MAU B, BLATTNER F R, et al. Mauve: multiple alignment of conserved genomic sequence with rearrangements[J]. Genome Research, 2004, 14(7): 1394−1403. DOI: 10.1101/gr.2289704. [26] SILVESTRO D, MICHALAK I. raxmlGUI: a graphical front-end for RAxML[J]. Organisms Diversity & Evolution, 2012, 12(4): 335−337. DOI: 10.1007/s13127-011-0056. [27] 王杰, 贺文闯, 向坤莉, 等. 基因组时代的植物系统发育研究进展[J]. 浙江农林大学学报, 2023, 40(1): 227−236. WANG Jie, HE Wenchuang, XIANG Kunli, et al. Advances in plant phylogeny in the genome era[J]. Journal of Zhejiang A&F University, 2023, 40(1): 227−236. DOI: 10.11833/j.issn.2095-0756.20220313. [28] SEOL Y J, KIM K, KANG S H, et al. The complete chloroplast genome of two Brassica species, Brassica nigra and B. oleracea[J]. Mitochondrial DNA Part A, DNA Mapping, Sequencing, and Analysis, 2017, 28(2): 167−168. DOI: 10.3109/19401736.2015.1115493. [29] ZHU Bin, GAO Zuomin, QIAN Fang, et al. The complete chloroplast genome of a purple Ethiopian rape (Brassica carinata: Brassicaceae) from Guizhou Province, China and its phylogenetic analysis[J]. Mitochondrial DNA Part B, 2021, 6(7): 1821−1823. DOI: 10.1080/23802359.2021.1926365. [30] ZHAO Mengliang, WU Yanxun, REN Yanjing. Complete chloroplast genome sequence structure and phylogenetic analysis of kohlrabi (Brassica oleracea var. gongylodes L. )[J]. Genes, 2024, 15(5): 550. DOI: 10.3390/genes15050550. [31] WANG Yilin, LIANG Qiannan, ZHANG Chenlu, et al. Sequencing and analysis of complete chloroplast genomes provide insight into the evolution and phylogeny of Chinese kale (Brassica oleracea var. alboglabra)[J]. International Journal of Molecular Sciences, 2023, 24(12): 10287. DOI: 10.3390/ijms241210287. [32] REDWAN R M, SAIDIN A, KUMAR S V. Complete chloroplast genome sequence of MD-2 pineapple and its comparative analysis among nine other plants from the subclass Commelinidae[J]. BMC Plant Biology, 2015, 15: 196. DOI: 10.1186/s12870-015-0587-1. [33] LI Yongfu, SYLVESTER S P, LI Meng, et al. The complete plastid genome of Magnolia zenii and genetic comparison to Magnoliaceae species[J]. Molecules, 2019, 24(2): 261. DOI: 10.3390/molecules24020261. [34] DURET L. tRNA gene number and Codon usage in the C. elegans genome are co-adapted for optimal translation of highly expressed genes[J]. Trends in Genetics, 2000, 16(7): 287−289. DOI: 10.1016/S0168-9525(00)02041-2. [35] 朱梦飞, 胡迎峰, 师雪芹. 濒危植物新绒苔叶绿体基因组特征及系统发育位置分析[J]. 浙江农林大学学报, 2025, 42(1): 55−63. ZHU Mengfei, HU Yingfeng, SHI Xueqin. Characterization and phylogenetic location analysis of chloroplast of the endangered plant Neotrichocolea bissetii[J]. Journal of Zhejiang A&F University, 2025, 42(1): 55−63. DOI: 10.11833/j.issn.2095-0756.20240356. [36] 蒋思思, 袁军, 周文君, 等. 薄壳山核桃(Carya illinoinensis)叶绿体基因组及其特征分析[J]. 园艺学报, 2022, 49(8): 1772−1784. JIANG Sisi, YUAN Jun, ZHOU Wenjun, et al. Complete chloroplast genome sequence and characteristics analysis of Carya illinoinensis[J]. Acta Horticulturae Sinica, 2022, 49(8): 1772−1784. DOI: 10.16420/j.issn.0513-353x.2021-0389. [37] WU Junyan, MA Xuecai, MA Li, et al. Complete chloroplast genome sequence and phylogenetic analysis of winter oil rapeseed (Brassica rapa L.)[J]. Mitochondrial DNA. Part B, Resources, 2021, 6(3): 723−731. DOI: 10.1080/23802359.2020.1860697. [38] 王云生, 黄宏文, 王瑛. 植物分子群体遗传学研究动态[J]. 遗传, 2007, 29(10): 1191−1198. WANG Yunsheng, HUANG Hongwen, WANG Ying. Recent progress in plant molecular population genetics[J]. Hereditas, 2007, 29(10): 1191−1198. DOI: 10.16288/j.yczz.2007.10.005. [39] XU Jin, LIU Chen, SONG Yun, et al. Comparative analysis of the chloroplast genome for four Pennisetum species: molecular structure and phylogenetic relationships[J]. Frontiers in Genetics, 2021, 12: 687844. DOI: 10.3389/fgene.2021.687844. -

-
链接本文:
https://zlxb.zafu.edu.cn/article/doi/10.11833/j.issn.2095-0756.20250279
 点击查看大图
点击查看大图
计量
- 文章访问数: 1070
- HTML全文浏览量: 535
- PDF下载量: 159
- 被引次数: 0
