





-
叶绿体基因组为植物质体遗传体系之一,是由大单拷贝区(LSC)、小单拷贝区(SSC)以及穿插在它们之间的一对碱基组成相同、排列方向相反的反向重复区(IR)构成的环状双链四分体,通常编码约130个主要与光合作用以及叶绿体自身复制相关的基因[1−2]。叶绿体基因组是研究植物物种鉴别、进化生物学和遗传多样性的新思路,并且已有较多的应用实例。SONG等[3]比较了27种安息香属Styrax植物的叶绿体基因组序列,筛选出6个高突变序列,其中ycf1b和trnT-trnL是安息香属物种鉴别的特异性DNA条形码。LIU等[4]利用叶绿体基因组序列探讨了11种燕麦属Avena植物的系统进化关系,支持燕麦属的单系性,并且认为该属包括2个遗传支系。JO等[5]基于饲料作物大花野豌豆Vicia bungei的叶绿体简单重复序列(SSR)开发出232个分子标记,并将其中39个分子标记应用于野豌豆属Vicia中7个物种的遗传多样性分析。
了哥王Wikstroemia indica是瑞香科Thymelaeaceae荛花属Wikstroemia常绿灌木植物,全株有毒,分布于中国广东、广西、海南、福建、湖南、浙江、云南、贵州、四川以及台湾等大部分省区,多见于海拔1 500 m以下开旷的林下或石山上,野生时一般分布在山坡灌木丛中或路边、村边等[6]。了哥王的干燥根及根皮可入药,能清热解毒、消肿散结、止痛,多用来治疗支气管炎、肺炎、乳腺炎等多种炎症以及跌打损伤[7],已被开发成了哥王片、了哥王胶囊、了哥王颗粒、祛伤消肿酊等制剂。了哥王含有黄酮、木质素、香豆素、甾体、挥发油等化学成分,具有抗炎、抗菌、抗病毒、抗肿瘤等药理活性[8−9]。了哥王根叶煮汁可作杀虫剂,茎皮纤维可以制作高级纸张和人造棉[6]。了哥王还是一种观赏性园林植物[10]。因此,了哥王具有较高的药用、经济和社会价值。
了哥王原以野生采收为主要来源,随着市场需求的快速增加,了哥王遭到过度开采,不少产地的野生蕴藏量急剧下降,出现资源匮乏甚至枯竭的情况[11]。本研究分析了哥王叶绿体基因组序列的结构和基因组成、密码子偏好性、SSR等特征,比较了哥王与近缘植物的叶绿体基因组序列差异,并基于叶绿体基因组探究了哥王的进化位置,旨在为了哥王的品种鉴定、育种栽培、遗传背景和系统进化等资源保护和可持续利用提供科学依据。
-
了哥王植株于2022年6月采自广西壮族自治区钦州市钦北区,经广东医科大学天然药物研究与开发重点实验室吴科锋研究员鉴定。取了哥王植株的新鲜叶片,使用北京康为世纪生物科技公司生产的磁珠法植物DNA提取试剂盒提取了哥王的总DNA,随后采用美国Illumina公司生产的NexteraXT DNA测序文库制备试剂盒构建了哥王的叶绿体基因组测序文库。
-
通过美国Illumina公司的Novaseq 6000高通量测序平台完成了哥王叶绿体基因组测序。测得的原始序列经滤除试剂盒附带的接头序列和含有无法确定碱基的劣质序列后,获得合格的可分析读序。采用SPAdes软件对可分析读序进行序列拼接,并运用plastid genome annotator软件对序列中含有的基因进行功能注释。拼接和注释后的了哥王叶绿体基因组序列和基因信息提交至GenBank保存(序列登记号为OQ831641),并提交到OGDRAW在线工具(
https://chlorobox.mpimp-golm.mpg.de/OGDraw.html )绘制物理图谱。 -
参照黄琼林[12]的研究,采用CodonW v1.3软件计算了哥王叶绿体基因组全部密码子的相对同义密码子使用情况(RSCU)值,评价其使用频率,RSCU>1为高频使用密码子。参照吴民华等[13]的研究,使用MISA软件(
https://webblast.ipk-gatersleben.de/misa/ )检索了哥王的SSR,分析其类型、组成基序、数量和分布等特点。 -
利用IRscope在线软件(
https://irscope.shinyapps.io/irapp/ )比较了哥王与同属植物细轴荛花W. nutans (MW393702)、荛花W. canescens (MW073911)、头序荛花W. capitate (MW073909)以及同科植物长柱瑞香Daphne championii (MT648376)叶绿体基因组IR边界(即IR贴近相邻LSC或SSC的碱基)的位置差异。采用设置为Shuffle-LAGAN模式的mVISTA软件(https://genome.lbl.gov/vista/index.shtml )进行前述5种瑞香科植物叶绿体基因组的多重序列比对,明确它们之间的碱基差异。 -
为探究了哥王在荛花属和瑞香科内的进化关系,在GenBank中下载已公开发布的瑞香科及其相关类群菱科Trapaceae、桃金娘科Myrtaceae植物叶绿体基因组序列,并以与了哥王亲缘关系较远的单子叶植物高良姜Alpinia officinarum的叶绿体基因序列作为外群,将所有下载序列与本研究测得的了哥王叶绿体基因组序列一起提交到MAFFT软件进行多重比对及对齐,随后导入设置为GTRGAMMA模型的RAxML软件构建系统进化树。
-
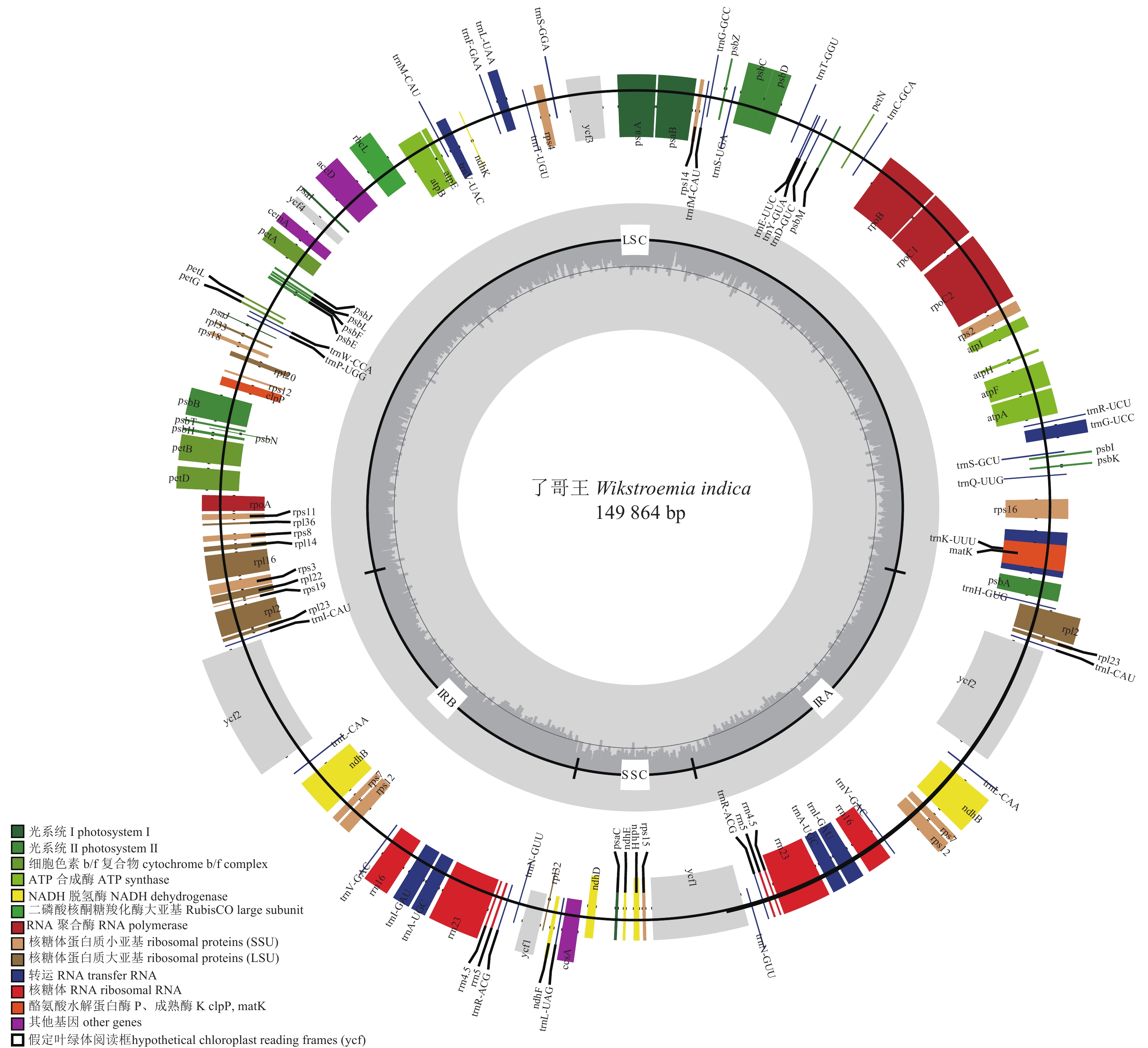
如图1所示:了哥王叶绿体基因组由位于1~86 347 bp的LSC、86 348~112 805 bp的IRA、112 806~123 407 bp的SSC和123 408~149 864 bp的IRB依顺时针排列而成,呈现为149 864 bp的环状双链四分体分子。了哥王叶绿体基因组的GC含量较低,仅为37.5%。
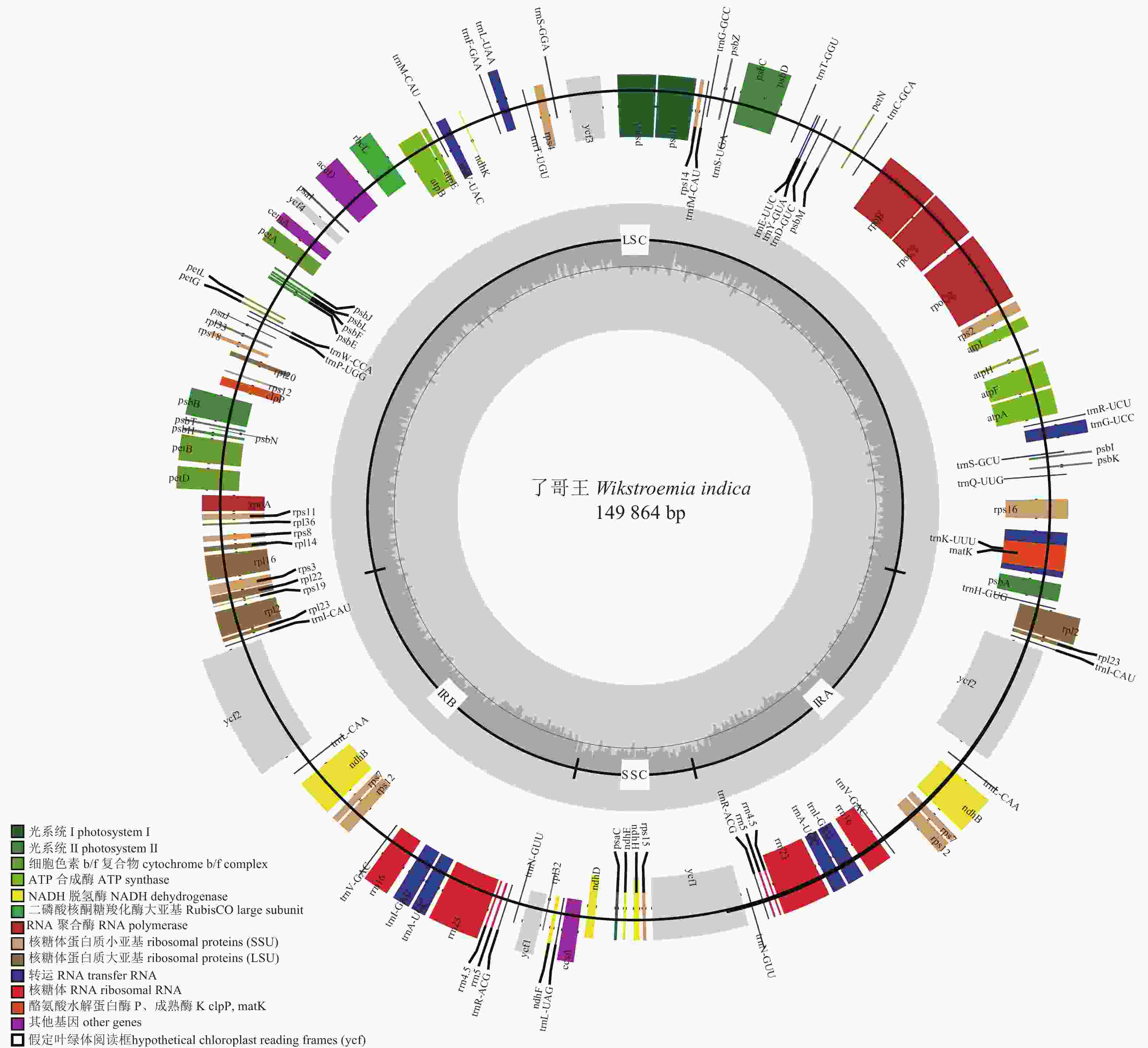
图 1 了哥王叶绿体基因组
Figure 1. Chloroplast geome of W. indica
从表1可见:了哥王叶绿体共有124个基因,包括79个蛋白质编码基因、8个核糖体RNA基因和37个转运RNA基因。这些基因中有14个基因(trnK-UUU、rps16、trnG-UCC、atpF、rpoC1、trnL-UAA、trnV-UAC、petB、petD、rpl16、rpl2、ndhB、trnI-GAU、trnA-UGC)含有1个内含子,1个基因(ycf3)存在2个内含子;17个基因(ndhB、rps7、rpl2、rpl23、ycf1、ycf2、rrn4.5、rrn5、rrn16、rrn23、trnI-CAU、trnL-CAA、trnV-GAC、trnI-GAU、trnA-UGC、trnR-ACG、trnN-GUU)具有2个拷贝。
表 1 了哥王叶绿体基因组基因组成和功能注释
Table 1. Gene composition and annotation in W. indica chloroplast genome
基因类别 基因功能 基因名称 蛋白质编码基因 ATP合成酶 atpA, atpB, atpE, atpF1, atpH, atpI 细胞色素b/f复合物 petA, petB1, petD1, petG, petL, petN NADH脱氢酶 ndhB1*, ndhD, ndhE, ndhF, ndhH, ndhK 光合系统Ⅰ psaA, psaB, psaC, psaI, psaJ 光合系统Ⅱ psbA, psbB, psbC, psbD, psbE, psbF, psbH, psbI, psbJ, psbK, psbL, psbM, psbN, psbT, psbZ 核糖体蛋白质小亚基 rps2, rps3, rps4, rps7*, rps8, rps11, rps12#, rps14, rps15, rps161, rps18, rps19 核糖体蛋白质大亚基 rpl21*, rpl14, rpl161, rpl20, rpl22, rpl23*, rpl32, rpl33, rpl36 RNA 聚合酶 rpoA, rpoB, rpoC11, rpoC2 假定叶绿体阅读框 ycf1*, ycf2*, ycf32, ycf4 其他基因 matK, rbcL, cemA, accD, ccsA, clpP 核糖体RNAs rrn4.5*, rrn5*, rrn16*, rrn23* 转运RNAs trnH-GUG, trnK-UUU1, trnM-CAU, trnI-CAU*, trnV-UAC1, trnF-GAA, trnL-UAA1, trnT-UGU, trnS- GGA, trnfM-CAU, trnG-GCC, trnS-UGA, trnT-GGU, trnE-UUC, trnY-GUA, trnD-GUC, trnC-GCA, trnR-UCU, trnG-UCC1, trnS-GCU, trnQ-UUG, trnW-CCA, trnP-UGG, trnL-CAA*, trnV-GAC*, trnI-GAU1*, trnA-UGC1*, trnR-ACG*, trnN-GUU*, trnL-UAG 说明:上标1表示含有1个内含子,上标2表示含有2个内含子,#表示反式剪接基因,*表示双拷贝基因。 -
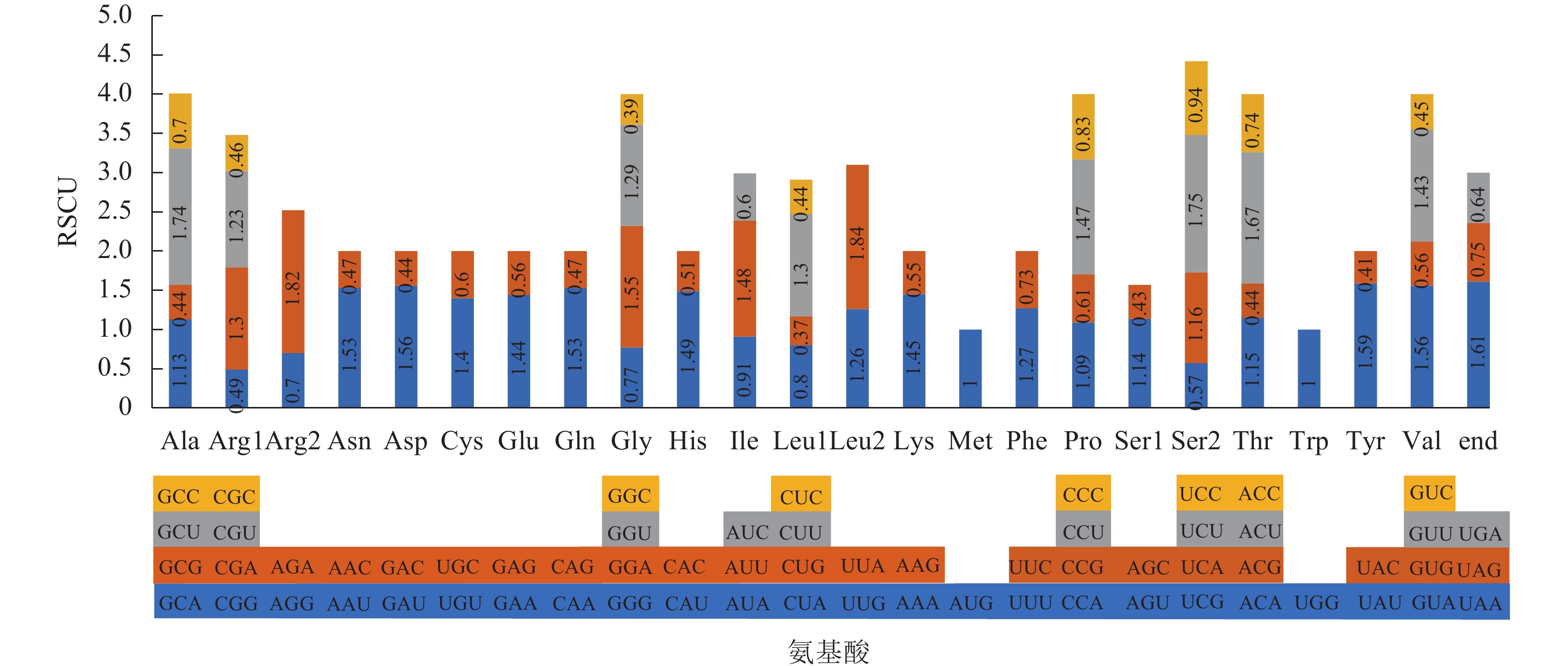
通过CodonW软件,在了哥王叶绿体基因组的编码区共找到24 180个密码子,包括24 100个氨基酸编码密码子和80个终止密码子。如图2所示:61种氨基酸编码密码子编码20种氨基酸,终止密码子有3种。编码相同氨基酸的多种密码子即称为同义密码子,除了甲硫氨酸(Met)和色氨酸(Trp)外,其余18种氨基酸均有2~6种同义密码子。除了AUG (单独编码Met)和UGG (单独编码Trp)的RSCU为1外,有30种密码子的RSCU>1,为高频使用密码子;其余32种密码子的RSCU<1,属于低频使用密码子。在30种高频使用密码子中,有29种的第3位碱基是A/T,表明了哥王叶绿体基因组偏好使用A/T结尾的密码子。
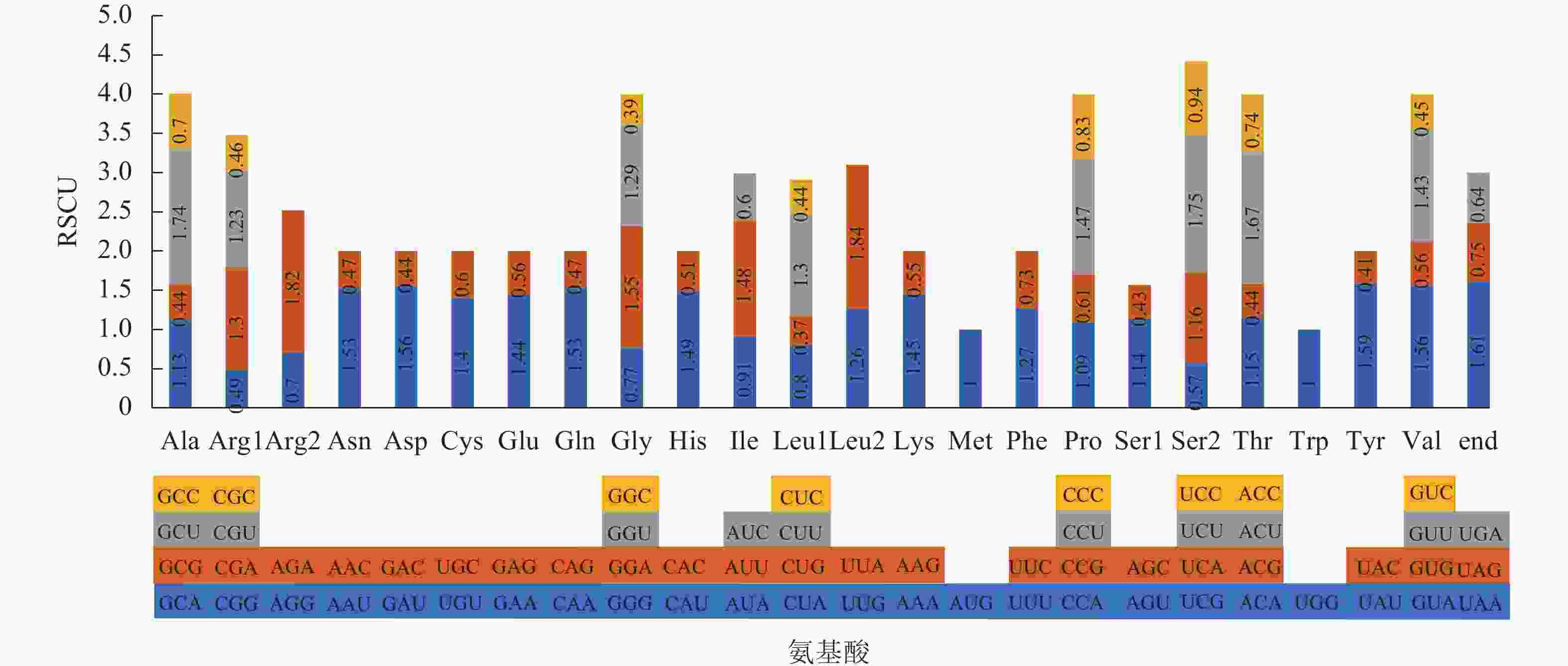
图 2 了哥王叶绿体基因组密码子的RSCU值
Figure 2. RSCU value of all codon in W. indica chloroplast genome
-
以设定的参数通过MISA软件查找,在了哥王叶绿体基因组中发现93个SSR,由72个单核苷酸、11个二核苷酸、4个三核苷酸、5个四核苷酸以及1个五核苷酸组成(表2)。在各类SSR中,A/T、AT/AT、AAT/ATT、AAAT/ATTT、AATAG/ATTCT分别是优势基序,数量依次是72、9、3、3、1个,分别占单核苷酸的100.0%、二核苷酸的81.2%、三核苷酸的75.0%、四核苷酸的60.0%和五核苷酸的100.0%。可见,由A/T及组合形成的基序数量最多,分别包括72个单核苷酸、9个二核苷酸、3个三核苷酸、4个四核苷酸,共占SSR总数的94.6%。因此,了哥王以A/T及组合形成的SSR居多,与其叶绿体基因组低GC含量的情况相符。在分布区域上来看,了哥王SSR主要分布在LSC,而SSC和IR较少;从所有位置上来看,这些SSR主要位于基因间隔区(IGS),基因内含子次之,编码区(CDS)则最少。说明了哥王叶绿体SSR分布广泛且不均匀,多态性较为丰富。
表 2 了哥王叶绿体基因组SSR位点的统计
Table 2. Summaries of SSR loci in W. indica chloroplast genome
类型 重复基序 数量 分布区域 所在位置 LSC SSC IR IGS Intron CDS 单核苷酸 A/T 72 56 8 8 48 14 10 二核苷酸 AT/AT 9 9 0 0 5 3 1 AC/GT 1 1 0 0 1 0 0 AG/CT 1 1 0 0 1 0 0 三核苷酸 AAT/ATT 3 1 0 2 1 2 0 AAG/CTT 1 1 0 0 0 1 0 四核苷酸 AAAT/ATTT 3 2 1 0 1 1 1 AATC /ATTG 1 0 1 0 0 0 1 AATT/AATT 1 1 0 0 0 0 1 五核苷酸 AATAG/ATTCT 1 1 0 0 1 0 0 合计 93 73 10 10 58 21 14 说明:LSC为大单拷贝区;SSC为小单拷贝区;IR为反向重复区;IGS基因间隔区;Intron为内含子;CDS为编码区。 -
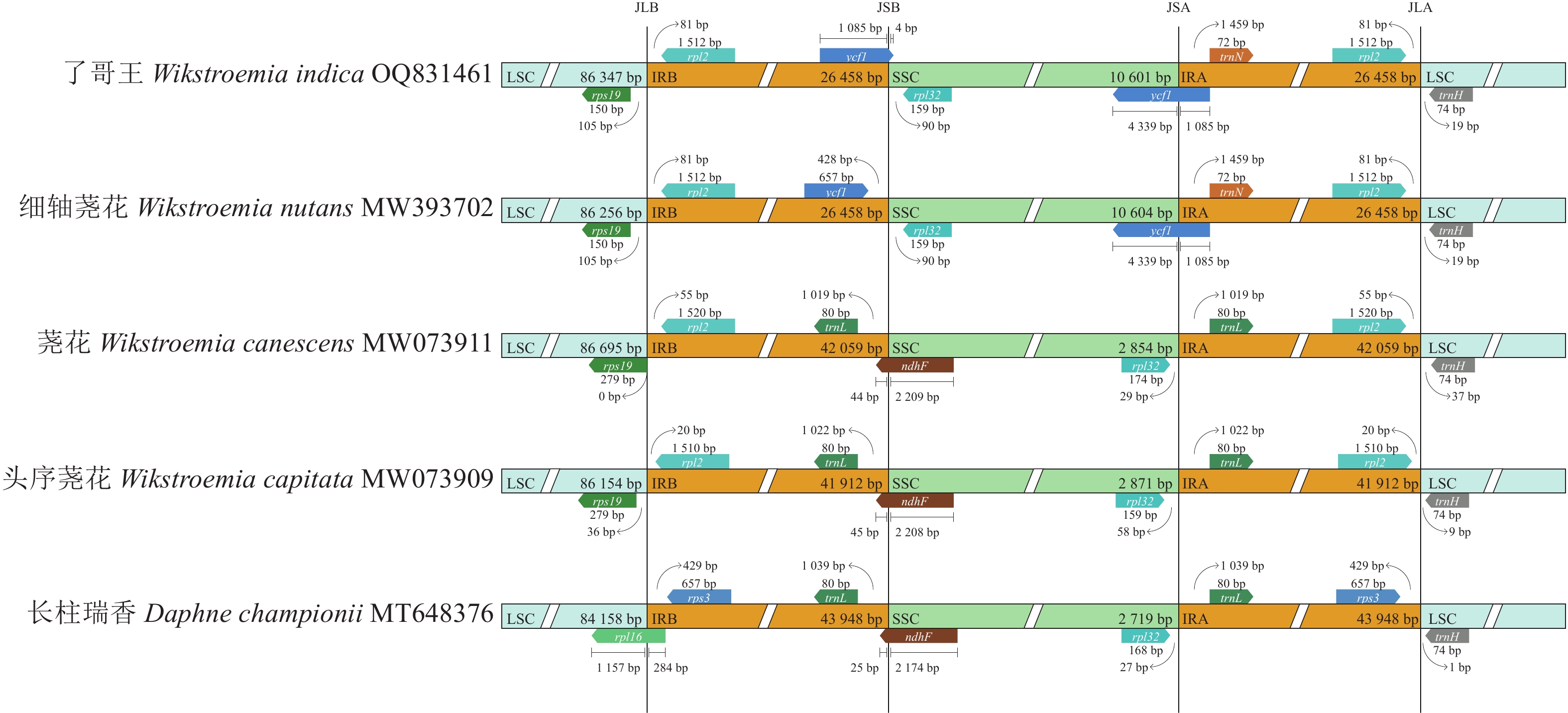
叶绿体基因组的2个IR在邻近LSC、SSC的位置形成边界,分别称作JLB (IRB-LSC)、JSB (IRB-SSC)、JSA (IRA-SSC)和JLA (IRA-LSC)。如图3所示:了哥王等4种荛花属植物的JLB均处于rpl19和rpl2之间的IGS,瑞香属Daphne植物长柱瑞香的JLB则在rpl16内。了哥王的JSB在ycf1内,细轴荛花的JSB在ycf1和rpl132之间的IGS,其余植物的JSB则在ndhF内。了哥王和细轴荛花的JSA都在ycf1内,且都在距离该基因5′端的1 085 bp处,其余植物的JSA则在rpl132和trnL之间的IGS。了哥王等4种荛花属植物的JLA均落在rpl2和trnH之间的IGS,长柱瑞香的JLA则位于rps3和trnH之间的IGS。由此可见,了哥王与其他4种植物的IR边界存在较为明显的差异(尤其是JSB),说明叶绿体基因组在了哥王等瑞香科植物进化过程中发生了不同程度的IR扩张和收缩。
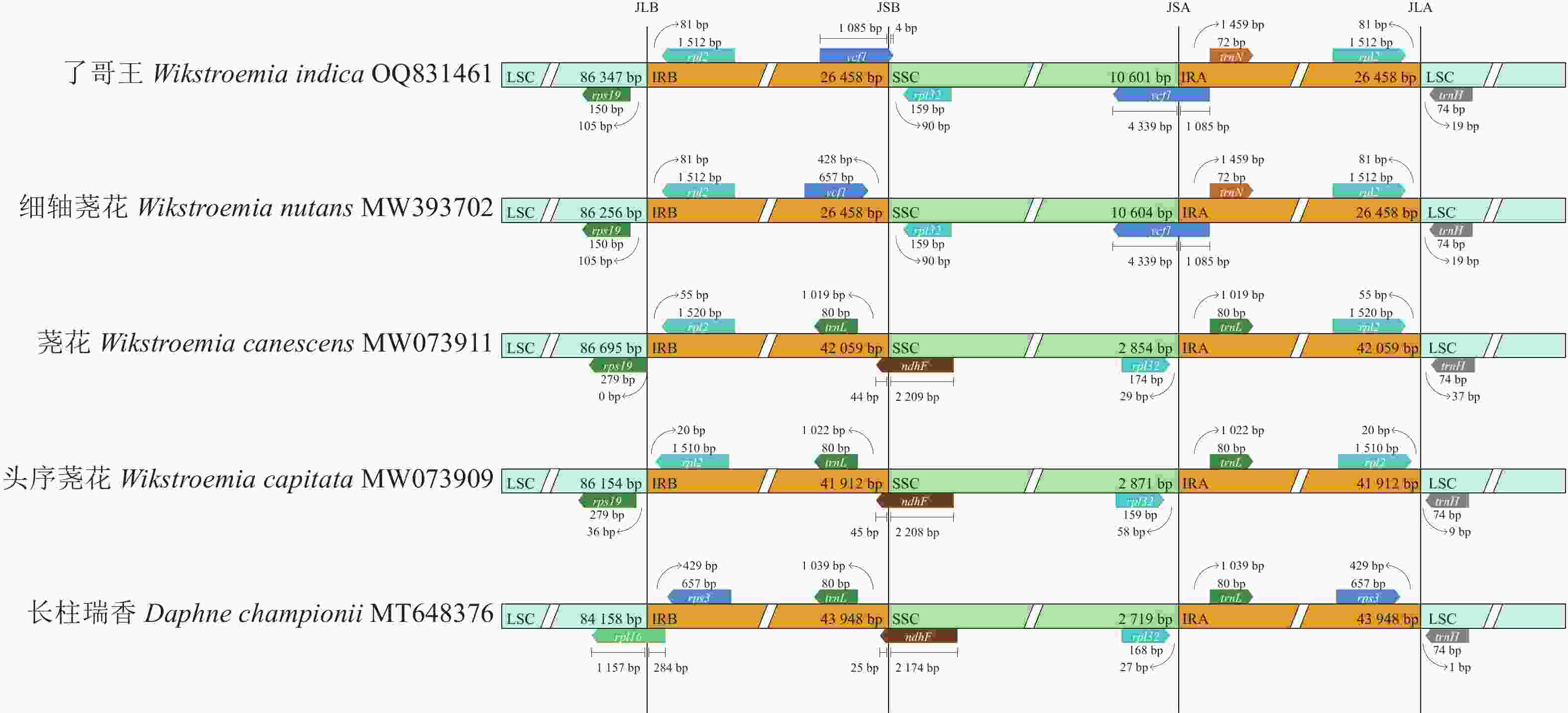
图 3 瑞香科5种植物叶绿体基因组IR边界的差异分析
Figure 3. IR boundaries divergence of chloroplast genomes from 5 Thymelaeaceae plants
-
如图4所示:了哥王与细轴荛花的序列相似度最高,仅在psbE-petL、atpF-atpH等IGS看到少量的碱基突变,并且编码区的序列完全一致。了哥王与另外2种荛花属植物荛花、头序荛花有着较明显的序列差异,几乎所有的IGS、内含子等非编码区序列均可见碱基突变,而且psbA、trnK-UUU、rpoC2、rpoC1、rpoB、accD、rpoA等多个基因也可见变异。了哥王与不同属的长柱瑞香则存在更加显著的序列差异,说明叶绿体基因序列差异与物种的亲缘关系密切相关。
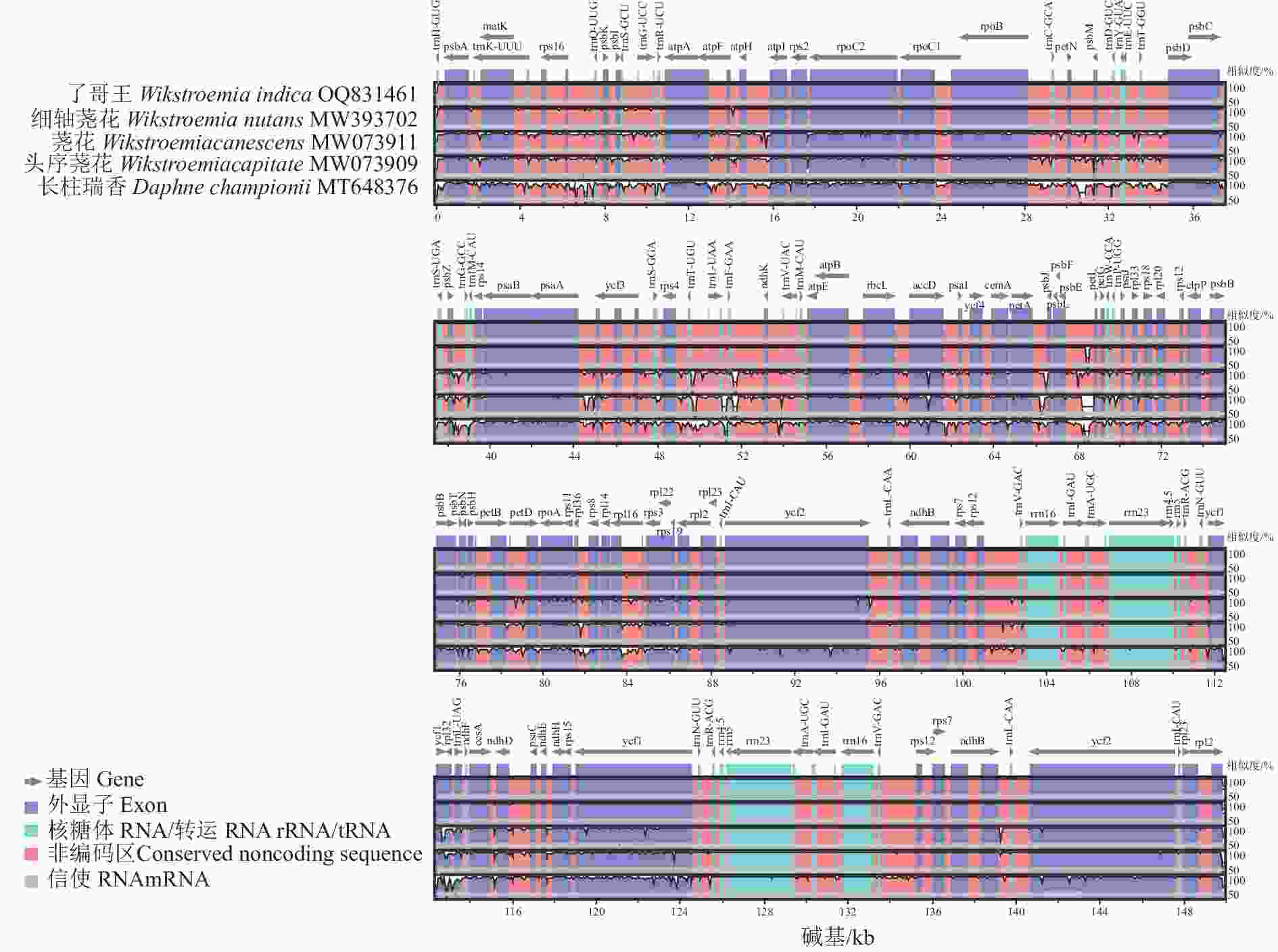
图 4 瑞香科5种植物叶绿体基因组的多重序列比对
Figure 4. Multiple alignment of chloroplast genome sequences from 5 Thymelaeaceae plants
-
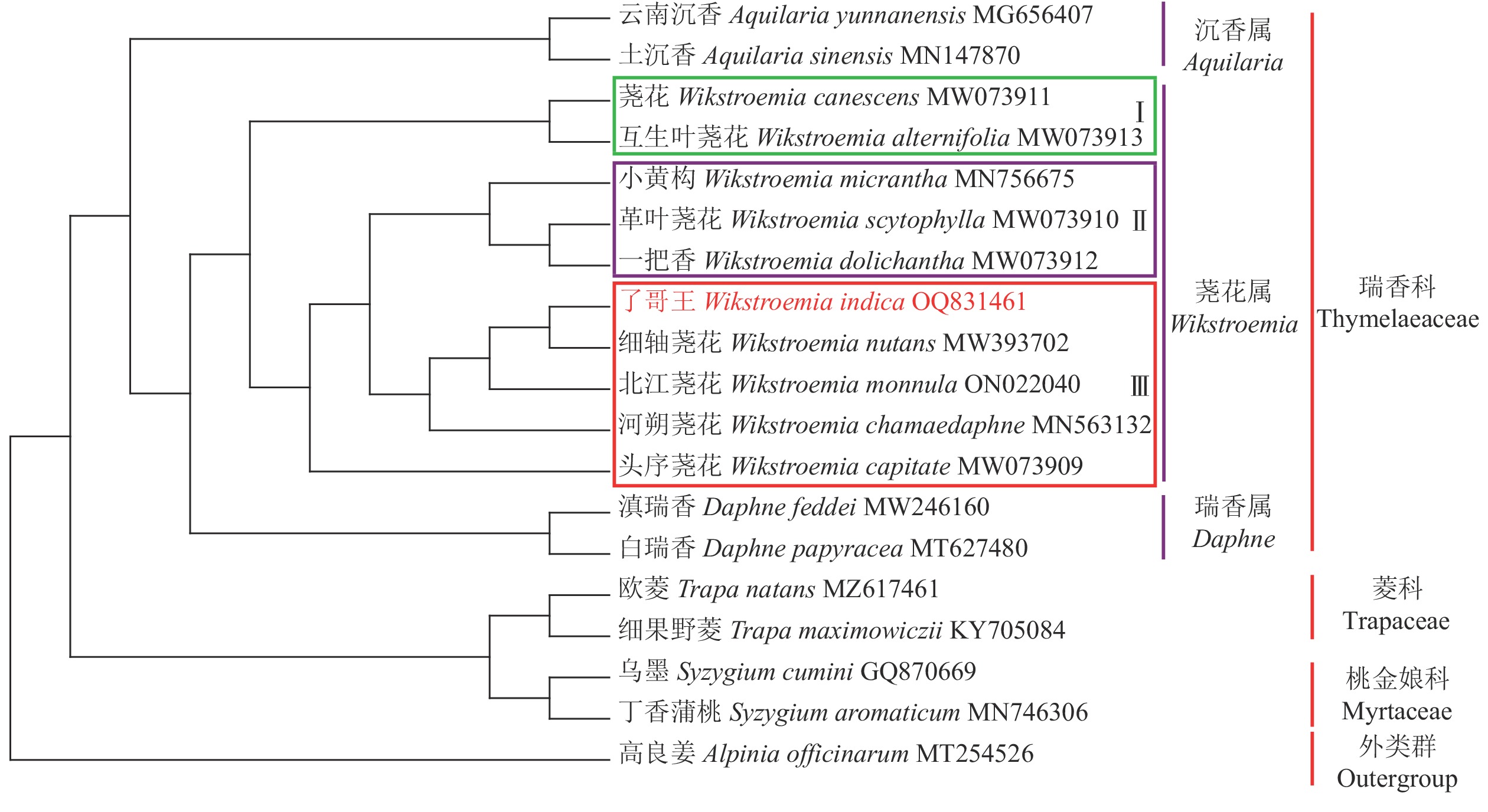
从图5可见:在科水平上,瑞香科、菱科、桃金娘科均形成独立的分支,并与外群高良姜分开。在瑞香科内,沉香属Aquilaria、荛花属和瑞香属各自形成分支,其中荛花属与瑞香属聚在一起,说明这2个属的亲缘关系更近。在荛花属内,9个物种大致聚成3个类群,类群Ⅰ包括荛花和互生叶荛花Wikstroemia canescens,类群Ⅱ含有小黄构W. micrantha、革叶荛花W. scytophylla和一把香W. dolichantha,类群Ⅲ则有了哥王、细轴荛花、北江荛花W. monnula、河朔荛花W. chamaedaphne和头序荛花。了哥王与细轴荛花最先聚成一个分支,表明它们具有最近的亲缘关系。
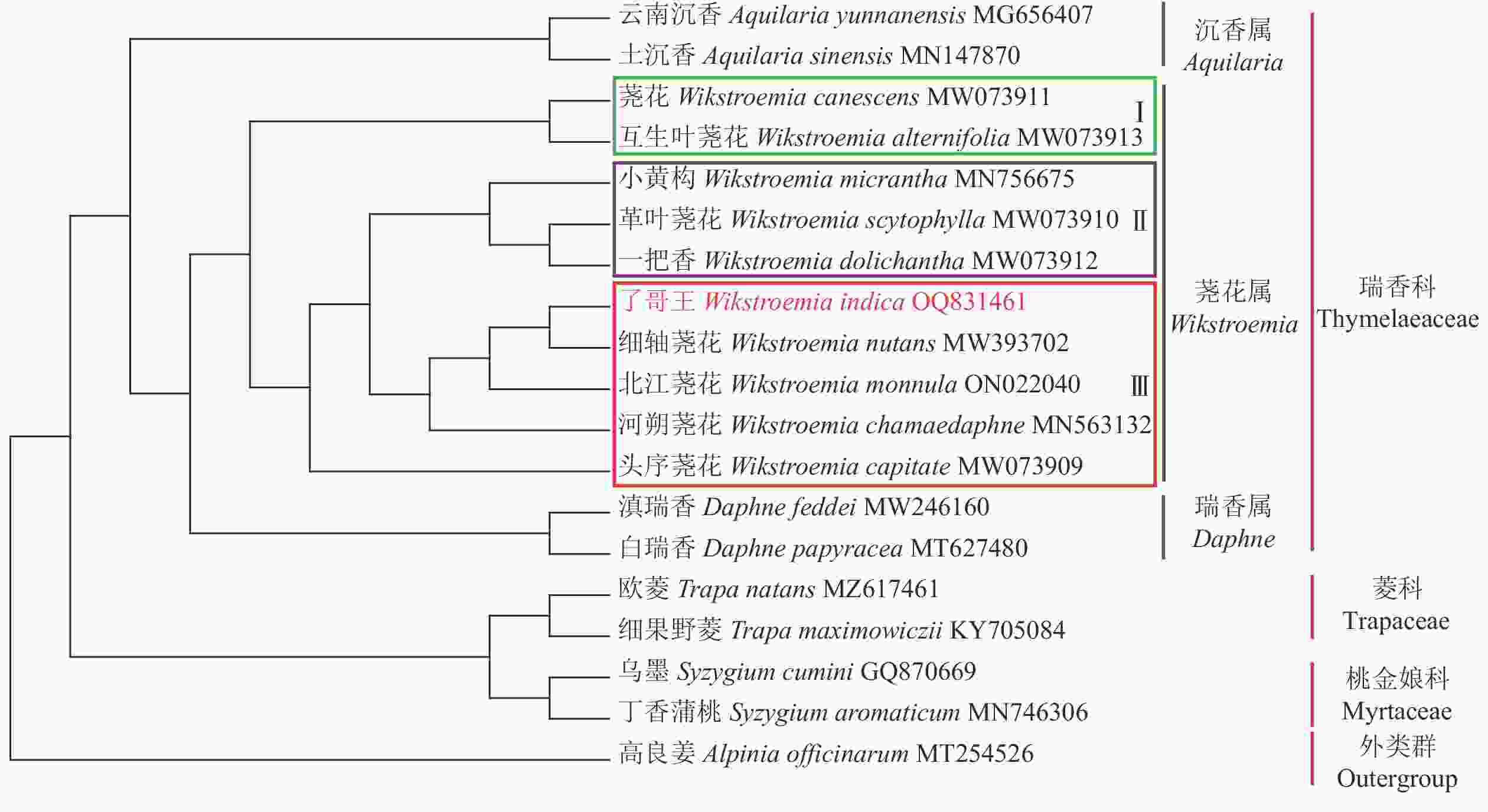
图 5 了哥王等植物的系统进化树
Figure 5. Phylogenetic tree among various plants including W. indica
-
研究发现:了哥王叶绿体基因组由LSC、SSC和两者之间的一对IR形成,呈现为陆生植物叶绿体基因组典型的环状双链四分体结构[14−15]。了哥王叶绿体基因组长度为149 864 bp,也介于被子植物叶绿体基因组110~160 kb范围内[16]。了哥王叶绿体基因组的GC含量为37.5%,与被子植物如瑞香科瑞香属[17]、豆科Leguminosae和蝶形亚科Papilionoideae[18]、兰科Orchidaceae[19]等物种的叶绿体基因组普遍存在的较低GC含量相符。叶绿体基因组作为执行光合作用的细胞器,容易受到光照等自然因素造成的选择压力的影响,在长期的进化过程中,叶绿体基因组可能发生了碱基替换和基因重组,导致其GC含量降低。此外,较低GC含量的DNA更容易解链,推测可以促进叶绿体基因组的重组,从而更好适应自然选择。在基因数量和组成上,了哥王叶绿体基因组共有124个基因,以与光合作用相关的基因及叶绿体自身复制所需的基因为主,也符合被子植物叶绿体基因组的基因构成[20]。因此,了哥王叶绿体基因组具有植物叶绿体基因组的共性特征。
植物在自然选择、碱基突变的作用下会形成一套与其自身进化相适应的常用密码子,以执行编码氨基酸或终止氨基酸翻译的功能。本研究在了哥王叶绿体基因组发现30种高频使用密码子(RSCU>1),包括29种氨基酸编码密码子和1种终止密码子,这些高频使用密码子比其他同义密码子更常被使用。除了编码亮氨酸(Leu)的UUG,其他高频密码子的末位碱基都是A/T。而且,在编码Leu的6种同义密码子中,UUG是使用频率较高的密码子(RSCU=1.26),排第3位,最高的是UUA (RSCU=1.84),其次是CUU (RSCU=1.30)。由此可见,了哥王叶绿体基因组编码20种氨基酸以及终止密码子最常用都是末位碱基为A/T的密码子,具有一定的密码子使用偏好性。相同的密码子偏好性也出现在金银花大毛花Lonicera japonica ‘Damaohua’[21]、高良姜[12]、菠萝Ananas comosus[22]等植物中。
SSR为一种具有丰富多态性的显性DNA标记,是研究植物品种鉴别、多样性分析以及构建遗传图谱、辅助分子育种的有效手段。利用MISA软件对了哥王叶绿体SSR进行定位和统计,发现了哥王含93个叶绿体SSR,且以A或T碱基及其组成的基序,尤其是polyA或polyT单核苷酸为优势类型,以分布在LSC和IGS为主,与北陵鸢尾Iris typhifolia[23]、露兜树Pandanus tectorius[13]、闭鞘姜Helenia speciosa [24]等植物的SSR特点相同。
IR被认为是叶绿体基因组中序列最为保守的区域,但IR经常会发生扩张和收缩,从而引起IR乃至整个叶绿体基因组长度的变化。因此,IR边界是比较物种叶绿体基因组差异的重要指标。本研究发现:了哥王与细轴荛花同属近缘植物的IR边界特别是SSC和IRB之间的JSB具有明显的不同。并且全序列比对显示:了哥王与细轴荛花同属近缘植物在IGS (如psbE-petL、atpF-atpH)等非编码区具有碱基变异。这些结果提示叶绿体基因组序列可以为了哥王及其近缘物种的鉴别提供依据。此外,系统进化树也直观地展示了了哥王的进化位置及其与相关近缘植物的亲缘关系,聚类效果良好。
-
本研究结果表明:了哥王叶绿体基因组为149 864 bp的环状双链四分体分子,GC含量为37.5%,含124个基因;偏好使用以A/T结尾的密码子;SSR以polyA或polyT单核苷酸为主。了哥王与细轴荛花具有最近的亲缘关系。
Analysis on chloroplast genome of Wikstroemia indica
-
摘要:
目的 阐明药用植物了哥王Wikstroemia indica的叶绿体基因组结构特点及系统进化地位,为了哥王的资源保护和可持续利用提供科学依据。 方法 采用Illumina测序平台进行了哥王叶绿体基因组测序,并通过生物信息技术和软件进行序列拼接、注释以及比对和系统进化分析。 结果 了哥王叶绿体基因组全长为149 864 bp,由86 347 bp的大单拷贝区(LSC)、10 601 bp的小单拷贝区(SSC)以及穿插在它们之间均为26 458 bp的一对反向重复区(IR)构成,具有环状双链四分体结构,包含124个基因。在了哥王叶绿体基因组中共找到64种24 180个密码子,其中30种为高频使用密码子,高频使用密码子中又有29种是以A/T结尾;搜索到93个简单重复序列(SSR),其中单核苷酸重复居多(72个),且以A或T及两者组合形成的基序为优势基序。了哥王与近缘植物的叶绿体基因组IR边界存在较为明显的变异。序列比较和系统进化树显示了哥王与同属细轴荛花W. nutans具有最高的序列同源性。 结论 了哥王叶绿体基因组具有植物叶绿体基因组的典型结构,有密码子使用偏好性,含多态性较为丰富的SSR,且与细轴荛花的亲缘关系最近。图5表2参24 Abstract:Objective This study is aimed to clarify the structure traits of chloroplast genome and phylogenetic status of medicinal plant Wikstroemia indica, providing a scientific basis for the resource protection and sustainable utilization of W. indica. Method An Illumina high-throughput sequencing platform was employed to acquire the chloroplast genome sequence which was then assembled, annotated, compared and analyzed using bioinformatics technologies and software. Result The chloroplast genome of W. indica was an annular quadripartite molecule with a length of 149 864 bp and composed of a large single copy (LSC, 86 347 bp), a small single copy (SSC, 10 601 bp) and a couple of inverted repeat (IR, 26 458 bp) and 124 genes were annotated in the chloroplast genome. In the chloroplast genome there were a total of 24 180 codon, which were divided into 64 types of which 30 types of codon were used frequently and 29 of them ended with A/T. Altogether 93 chloroplast simple sequence repeat (SSR) were identified from the chloroplast genome with Mononucleotide being the predominate type taking up a number of 72 and the motifs formed by A/T contributing the most and obvious variations were observed in IR boundary between W. indica and closely related plants whereas sequence alignment and phylogenetic tree revealed that W. indica shared high sequence identity with W. nutans. Conclusion The chloroplast genome of W. indica exhibited typical structure of plant chloroplast genome, and possessed codon preference and several SSRs with rich polymorphism. Additionally, the chloroplast genome of W. indica showed the closest similarity to W. nutans. [Ch, 5 fig. 2 tab. 24 ref.] -
图 2 了哥王叶绿体基因组密码子的RSCU值
Figure 2 RSCU value of all codon in W. indica chloroplast genome
图 3 瑞香科5种植物叶绿体基因组IR边界的差异分析
Figure 3 IR boundaries divergence of chloroplast genomes from 5 Thymelaeaceae plants
图 4 瑞香科5种植物叶绿体基因组的多重序列比对
Figure 4 Multiple alignment of chloroplast genome sequences from 5 Thymelaeaceae plants
表 1 了哥王叶绿体基因组基因组成和功能注释
Table 1. Gene composition and annotation in W. indica chloroplast genome
基因类别 基因功能 基因名称 蛋白质编码基因 ATP合成酶 atpA, atpB, atpE, atpF1, atpH, atpI 细胞色素b/f复合物 petA, petB1, petD1, petG, petL, petN NADH脱氢酶 ndhB1*, ndhD, ndhE, ndhF, ndhH, ndhK 光合系统Ⅰ psaA, psaB, psaC, psaI, psaJ 光合系统Ⅱ psbA, psbB, psbC, psbD, psbE, psbF, psbH, psbI, psbJ, psbK, psbL, psbM, psbN, psbT, psbZ 核糖体蛋白质小亚基 rps2, rps3, rps4, rps7*, rps8, rps11, rps12#, rps14, rps15, rps161, rps18, rps19 核糖体蛋白质大亚基 rpl21*, rpl14, rpl161, rpl20, rpl22, rpl23*, rpl32, rpl33, rpl36 RNA 聚合酶 rpoA, rpoB, rpoC11, rpoC2 假定叶绿体阅读框 ycf1*, ycf2*, ycf32, ycf4 其他基因 matK, rbcL, cemA, accD, ccsA, clpP 核糖体RNAs rrn4.5*, rrn5*, rrn16*, rrn23* 转运RNAs trnH-GUG, trnK-UUU1, trnM-CAU, trnI-CAU*, trnV-UAC1, trnF-GAA, trnL-UAA1, trnT-UGU, trnS- GGA, trnfM-CAU, trnG-GCC, trnS-UGA, trnT-GGU, trnE-UUC, trnY-GUA, trnD-GUC, trnC-GCA, trnR-UCU, trnG-UCC1, trnS-GCU, trnQ-UUG, trnW-CCA, trnP-UGG, trnL-CAA*, trnV-GAC*, trnI-GAU1*, trnA-UGC1*, trnR-ACG*, trnN-GUU*, trnL-UAG 说明:上标1表示含有1个内含子,上标2表示含有2个内含子,#表示反式剪接基因,*表示双拷贝基因。  下载: 导出CSV
下载: 导出CSV
表 2 了哥王叶绿体基因组SSR位点的统计
Table 2. Summaries of SSR loci in W. indica chloroplast genome
类型 重复基序 数量 分布区域 所在位置 LSC SSC IR IGS Intron CDS 单核苷酸 A/T 72 56 8 8 48 14 10 二核苷酸 AT/AT 9 9 0 0 5 3 1 AC/GT 1 1 0 0 1 0 0 AG/CT 1 1 0 0 1 0 0 三核苷酸 AAT/ATT 3 1 0 2 1 2 0 AAG/CTT 1 1 0 0 0 1 0 四核苷酸 AAAT/ATTT 3 2 1 0 1 1 1 AATC /ATTG 1 0 1 0 0 0 1 AATT/AATT 1 1 0 0 0 0 1 五核苷酸 AATAG/ATTCT 1 1 0 0 1 0 0 合计 93 73 10 10 58 21 14 说明:LSC为大单拷贝区;SSC为小单拷贝区;IR为反向重复区;IGS基因间隔区;Intron为内含子;CDS为编码区。
下载: 导出CSV
-
[1] 邢少辰, CLARKE JIHONG LIU. 叶绿体基因组研究进展[J]. 生物化学与生物物理进展, 2008, 35(1): 21 − 28. XING Shaochen, CLARKE J L. Progress in chloroplast genome analysis [J]. Progress in Biochemistry and Biophysics, 2008, 35(1): 21 − 28. [2] DANIELL H, LIN C S, YU Ming, et al. Chloroplast genomes: diversity, evolution, and applications in genetic engineering [J/OL]. Genome Biology, 2016, 17(1): 134[2023-07-01]. doi: 10.1186/s13059-016-1004-2. [3] SONG Yun, ZHAO Wenjun, XU Jin, et al. Chloroplast genome evolution and species identification of Styrax (Styracaceae) [J/OL]. Biomed Research International, 2022, 2022: 5364094[2023-07-01]. doi: 10.1155/2022/5364094. [4] LIU Qing, LI Xiaoyu, LI Mingzhi, et al. Comparative chloroplast genome analyses of Avena: insights into evolutionary dynamics and phylogeny [J/OL]. BMC Plant Biology, 2020, 20(1): 406[2023-07-01]. doi: 10.1186/s12870-020-02621-y. [5] JO I H, HAN S, SHIM D, et al. Complete chloroplast genome of the inverted repeat-lacking species Vicia bungei and development of polymorphic simple sequence repeat markers [J/OL]. Frontiers in Plant Science, 2022, 13: 891783[2023-07-01]. doi: 10.3389/fpls.2022.891783. [6] 中国植物志编辑委员会. 中国植物志(52卷)[M]. 北京: 科学出版社, 1995. Committee of Flora of China. Flora of China (Volume 52) [M]. Beijing: Science Press, 1995. [7] 广东省食品药品监督管理局. 广东省中药材标准(第1册)[M]. 广州: 广东科技出版社, 2019. Guangdong Medical Product Administration. Standard for Traditional Chinese Medicine in Guangdong Province (Volume 1) [M]. Guangzhou: Guangdong Science & Technology Press, 2019. [8] 尹震花, 张娟娟, 陈林, 等. 了哥王化学成分与生物活性研究进展[J]. 中草药, 2018, 49(8): 1964 − 1976. YIN Zhenhua, ZHANG Juanjuan, CHEN Lin, et al. Research progress on chemical constituents and biological activities of Wikstroemia indica [J]. Chinese Traditional and Herbal Drugs, 2018, 49(8): 1964 − 1976. [9] 郑传奇, 冯果, 李伟, 等. 了哥王生物活性及抗肿瘤作用研究进展[J]. 微量元素与健康研究, 2020, 37(3): 46 − 48. ZHENG Chuanqi, FENG Guo, LI Wei, et al. Research progress on bioactivity and anti-tumor effects of the Wikstroemia indica [J]. Studies of Trace Elements and Health, 2020, 37(3): 46 − 48. [10] 张群, 陈颖, 曹晟烜, 等. 了哥王叶功能性状特征及其对土壤因子的响应[J]. 亚热带植物科学, 2023, 52(1): 60 − 66. ZHANG Qun, CHEN Ying, CAO Shengxuan, et al. Characterization of leaf functional traits of Wikstroemia indica and its responses to soil factors [J]. Subtropical Plant Science, 2023, 52(1): 60 − 66. [11] 房海灵, 邓绍勇, 朱培林. 不同引发处理对了哥王种子萌发的影响[J]. 南方林业科学, 2016, 44(2): 8 − 11. FANG Hailing, DENG Shaoyong, ZHU Peiling. Effects of seed priming with different agents on seed germination of Wikstroemia indica [J]. South China Forestry Science, 2016, 44(2): 8 − 11. [12] 黄琼林. 高良姜叶绿体基因组测序与特征分析[J]. 热带作物学报, 2021, 42(1): 1 − 6. HUANG Qionglin. Complete sequencing and analysis of chloroplast genome from Alpinia officinarum Hance [J]. Chinese Journal of Tropical Crop, 2021, 42(1): 1 − 6. [13] 吴民华, 邹振宁, 叶晓霞, 等. 露兜树叶绿体基因组结构与序列特征分析[J]. 中药新药与临床药理, 2023, 34(1): 115 − 122. WU Minhua, ZOU Zhenning, YE Xiaoxia, et al. Structure and sequence characteristics of chloroplast genome from Pandanus tectorius [J]. Traditional Chinese Drug Research and Clinical Pharmacology, 2023, 34(1): 115 − 122. [14] BENDICH A J. Circular chloroplast chromosomes: the grand illusion [J]. Plant Cell, 2004, 16(7): 1661 − 1666. [15] ZHANG Yanjun, DU Liuwen, LIU Ao, et al. The complete chloroplast genome sequences of five epimedium species: lights into phylogenetic and taxonomic analyses [J/OL]. Frontiers in Plant Science, 2016, 7: 306[2023-07-01]. doi: 10.3389/fpls.2016.00306. [16] DANIELL H, LEE S B, GREVICH J, et al. Complete chloroplast genome sequences of Solanum bulbocastanum, Solanum lycopersicum and comparative analyses with other Solanaceae genomes [J]. Theoretical and Applied Genetics, 2006, 112(8): 1503 − 1518. [17] 钱少娟. 瑞香属的叶绿体基因组特征与系统学研究[D]. 昆明: 云南师范大学, 2021. QIAN Shaojuan. Chloroplast Genome Characteristics and Phylogeny of the Genus Daphne [D]. Kunming: Yunnan Normal University, 2021. [18] ZHAO Xueli, ZHU Zhangming. Comparative genomics and phylogenetic analyses of Christia vespertilionis and Urariopsis brevissima in the Tribe Desmodieae (Fabaceae: Papilionoideae) based on complete chloroplast genomes [J/OL]. Plants, 2020, 9(9): 1116[2023-07-01]. doi: 10.3390/plants9091116. [19] 刘浩宇, 刘力宽, 王郑雷, 等. 兰科植物叶绿体基因组研究进展[J]. 中国野生植物资源, 2023, 42(7): 73 − 79. LIU Haoyu, LIU Likuan, WANG Zhenglei, et al. Research progress on chloroplast genome of Orchidaceae [J]. Chinese Wild Plant Resources, 2023, 42(7): 73 − 79. [20] 宋菊, 龙月红, 林丽梅, 等. 五加科植物叶绿体基因组结构与进化分析[J]. 中草药, 2017, 48(24): 5070 − 5075. SONG Ju, LONG Yuehong, LIN Limei, et al. Analysis on structure and phylogeny of chloroplast genomes in Araliaceae species [J]. Chinese Traditional and Herbal Drugs, 2017, 48(24): 5070 − 5075. [21] 张加强, 刘慧春, 王杰, 等. 金银花大毛花叶绿体基因组密码子的偏好性分析[J]. 浙江农业学报, 2023, 35(4): 821 − 830. ZHANG Jiaqiang, LIU Huichun, WANG Jie, et al. Analysis on codon usage patterns of chloroplast genome of Lonicera japonica cv. Damaohua [J]. Acta Agriculturae Zhejiangensis, 2023, 35(4): 821 − 830. [22] 杨祥燕, 蔡元保, 谭秦亮, 等. 菠萝叶绿体基因组密码子偏好性分析[J]. 热带作物学报, 2022, 43(3): 439 − 446. YANG Xiangyan, CAI Yuanbao, TAN Qinliang, et al. Analysis of codon usage bias in the chloroplast genome of Ananas comosus [J]. Chinese Journal of Tropical Crops, 2022, 43(3): 439 − 446. [23] 卢政阳, 于凤扬, 肖月娥, 等. 北陵鸢尾叶绿体基因组及其特征分析[J]. 草地学报, 2023, 31(6): 1656 − 1664. LU Zhengyang, YU Fengyang, XIAO Yue’ e, et al. Complete chloroplast genome and characteristics analysis of Iris typhifolia [J]. Acta Agrestia Sinica, 2023, 31(6): 1656 − 1664. [24] 吴永飞, 杨雪莲, 颜丽, 等. 闭鞘姜叶绿体基因组特征及其系统发育分析[J]. 种子, 2023, 42(4): 10 − 17. WU Yongfei, YANG Xuelian, YAN Li, et al. Analysis on chloroplast genome characteristics and phylogeny of Helenia speciosa [J]. Seed, 2023, 42(4): 10 − 17. -

-
链接本文:
https://zlxb.zafu.edu.cn/article/doi/10.11833/j.issn.2095-0756.20230412
 点击查看大图
点击查看大图
计量
- 文章访问数: 1675
- HTML全文浏览量: 662
- PDF下载量: 22
- 被引次数: 0
