




-
草地作为陆地生态系统的重要组成部分,不仅为人类提供了具有直接经济价值的产品,更在调节气候、涵养水源、维持生物多样性与促进碳固存等方面发挥着重要的生态功能[1−2]。合理的草地管理与利用能够有效提升草地生产力,同时维持草地生态系统功能的稳定[3];反之,不合理利用或干扰则会破坏生态系统平衡,进而导致草地退化[4]。草地退化不仅会直接降低其生物多样性水平,更会危及到区域生态安全和草地畜牧业的可持续发展[5−7]。因此,根据草地类型及健康状况,探索合理的管理与利用措施,对于维持草地生态系统稳定具有重要的科学与实践意义。
物种多样性和生产力作为草地生态系统结构和功能的重要指标[8],其变化深刻影响着草地生态系统的稳定性与服务能力[9]。围封和放牧作为常见的草地管理与利用方式,对草地生态系统的影响具有双重性[10−11]。研究表明:放牧干扰或围封主要通过影响草地生产力和物种多样性水平,进而影响草地生态系统结构与功能的稳定[3, 12]。适度放牧可促进草地植物再生,提高草地生产力,同时有利于维持物种多样性水平[13];但是,过度放牧则会导致草地裸露、生产力下降以及物种多样性丧失[14−15]。同样,对于退化草地,合理围封有助于改善土壤质量,提高植被覆盖度、物种多样性和生产力[16−18]。然而,对草地进行不合理围封,则会导致一系列生产和生态问题,例如草地植物多样性降低,土壤碳氮储量减少,凋落物过度积累,影响植物群落更新[19−21]。
目前,关于围封和放牧对草地生态系统影响的研究,大多聚焦于单一管理措施对草地群落结构和功能的影响。例如,围封对于遏制草地退化、促进植被恢复的作用[22],不同围封年限对草地生态系统各指标动态变化的影响[23],以及不同放牧压力梯度下,草地植被组成、物种多样性水平及生产力的变化规律等[24]。关于长期围封与适度放牧如何影响未退化高寒草地的群落结构与功能,仍缺乏充分研究。本研究以青藏高原东缘未退化的高寒草地为研究对象,通过长期围封与自由放牧的对比试验,比较了2种草地管理模式对草地植物群落特征和生产力的影响,旨在探究2种草地管理模式下,草地植物生产力和物种多样性水平是否存在显著差异,是否会导致同一草地群落结构产生分异等科学问题。
-
研究区位于祁连山国家公园东缘天祝藏族自治县甘肃农业大学高山草原生态实验站,海拔为2 960 m。该区域年均气温约为 −1.0 ℃,≥0 ℃的年积温约为1 380 ℃;年平均降水量约为416.0 mm,且主要集中在每年的7—9月,年均蒸发量高达1 592.0 mm,属于典型的高原大陆性季风气候。该地区无明显绝对无霜期,仅分冷、热两季,植物生长期约120~140 d [25]。土壤类型为亚高山草甸土,植被类型为高寒草甸,主要草种有垂穗披碱草Elymus nutans、矮嵩草Kobresia humilis、洽草Koeleria cristata、针茅Stipa krylovii、珠芽蓼Polygonum viviparum和球花蒿Artemisia smithii等[26]。
-
为探究长期围封与适度放牧2种管理模式对未退化高寒草地群落的影响,本研究选取植被生长状况一致良好的未退化草地。自2015年起,将其划分为2个处理区域:一半实施长期围封,另一半根据国家行业标准 NY/T
3647 —2020《草食家畜羊单位换算》[27],全年进行适度放牧(研究区域总面积约17 hm2,划定围封区域面积约3 hm2)。围封处理区域,草地植被保持自然生长状态,无任何人为活动干扰。为统一采样条件,数据采集当年,放牧区草地暂停放牧活动。2025年8月初,将围封草地和放牧草地分别等面积划分为4个小区,于各小区中心位置设置1个50 cm×50 cm的样方(试验布设样方的局部草地面积约0.01 hm2)。调查记录每个样方内所有植物种类,并采用针刺法测定样方总盖度及各物种盖度。此外,在各样方内齐地面剪割地上植物部分,并收集样方内全部凋落物,分别装袋带回实验室,置于60 ℃烘箱内连续烘烤48 h至恒量后称量,以此作为地上生物量与凋落物量的评价指标。 -
物种重要值是综合评价草地群落物种优势地位的指标,计算公式为[3]:重要值=(相对多度+相对盖度+相对地上生物量)/3,其中,相对多度指某个物种的个体数与样方内所有物种个体数的比值,相对盖度指某物种盖度与样方总盖度的比值,相对地上生物量指某物种地上生物量与样方总地上生物量的比值。
用物种等级-多度关系(rank-abundance distribution, RAD)分析围封与放牧草地的群落结构与物种分布均匀度。物种等级指将群落内所有物种按相对多度由高到低排序后的位次;多度采用相对多度(M)表示,即某一物种个体数占群落总个体数的比值[28]。以物种等级为横坐标、log10M为纵坐标进行线性回归;回归斜率用于衡量物种分布均匀度,斜率绝对值越小,物种分布越均匀;斜率绝对值越大,物种分布均匀度越低。
草地群落物种多样性特征选取Margalef丰富度指数(S)、Simpson多样性指数(D)、Shannon-Weiner多样性指数(H)及物种丰富度指数(R)表征 [29−30]。
-
2种处理的物种重要值、物种多样性和生物量指标的差异显著性采用独立样本t检验(independent samples t-test)分析。物种等级-多度线性回归方程及斜率比较分别采用R软件中的lm()和anova()函数进行分析。群落结构的差异采用R软件中主成分分析(PCA,vegan包中的rda()函数)。所有数据分析及绘图均在R 4.5.1软件中进行。
-
草地群落总的植物物种组成以及各功能群的物种组成在围封和放牧处理下均存在差异(表1)。围封草地共有植物12种,赖草Leymus secalinus、垂穗披碱草Elymus nutans等7种植物的重要值高于20.00%;放牧草地共有植物20种,重要值超过20.00%的植物有8种。围封草地和放牧草地共有物种为10种,除青藏薹草Carex moorcroftii外,羊茅Festuca ovina、青海仲彬草Kengyilia kokonorica等9种植物的重要值均在围封草地中较高。其中,围封草地紫花针茅的重要值极显著高于放牧草地(P<0.01),围封草地为37.33%,放牧草地为10.70%;围封草地青海苜蓿的重要值显著高于放牧草地(P<0.05),围封草地为50.73%,比放牧草地高12.90%。赖草的重要值在围封草地与放牧草地均居首位,分别为60.45%与54.33%,围封草地特有物种仅有2种,且均属于杂类草功能群;放牧草地则高达10种,在4个功能群中均有分布。
表 1 草地群落植物物种组成及重要值在围封草地与放牧草地的差异
Table 1. Difference of plant species composition and its important value between fencing and grazing grassland in grassland community
功能群 物种 重要值/% 功能群 物种 重要值/% 围封草地 放牧草地 围封草地 放牧草地 禾本科 赖草 60.45±2.36 54.33±5.25 杂类草 蒲公英 36.95±0.90 19.18±6.39 垂穗披碱草 39.35±1.40 36.25±1.66 球花蒿 4.25±4.25 2.13±2.13 羊茅 20.23±6.81 9.18±5.33 茵陈蒿 − 8.63±4.98 青海仲彬草 21.10±7.16 12.58±7.26 麻花艽 9.38±5.41 2.15±2.15 紫花针茅 37.33±1.44** 10.70±6.34 高山唐松草 2.28±2.28 − 冰草 − 25.45±10.35 棉毛茛 − 34.33±0.18 
− 42.00±2.61 平车前 − 2.50±2.50 草地早熟禾 − 3.28±3.28 翻白委陵菜 − 19.73±6.59 豆科 青海苜蓿 50.73±2.91* 37.83±1.59 高山韭 2.28±2.28 − 黄花棘豆 − 34.03±0.09 鳞茎堇菜 − 18.93±6.31 莎草科 青藏薹草 19.75±6.59 24.48±8.44 矮生嵩草 − 37.70±0.48 说明:数据为平均值±标准误。−.样地内不存在的物种。*表示与放牧草地间存在显著性差异(P<0.05)。**表示与放牧草地间差异极显著(P<0.01)。禾本科Poaceae;豆科Fabaceae;莎草科Cyperaceae。冰草Agropyron cristatum; 
-
由图1可知:围封对草地群落总的植物物种多样性存在显著影响。放牧草地的Margalef指数为2.21,显著高于围封草地的1.42 (P<0.05);围封草地和放牧草地的物种丰富度也存在显著差异(P<0.05),围封草地有8.75种,比放牧草地少5.00种。围封草地的Shannon-Wiener指数和Simpson指数分别为1.44和0.25,放牧草地分别为1.72和0.28,差异不显著。
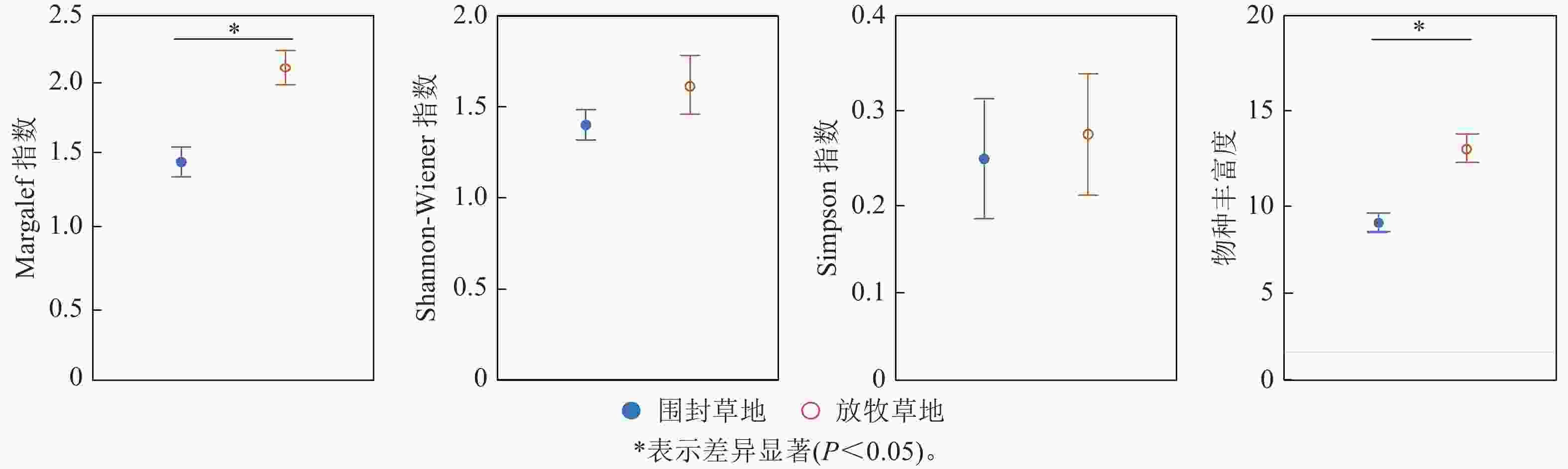
图 1 草地植物群落物种多样性指数在围封草地与放牧草地的差异
Figure 1. Difference of species diversity index in grassland community between fencing and grazing grassland
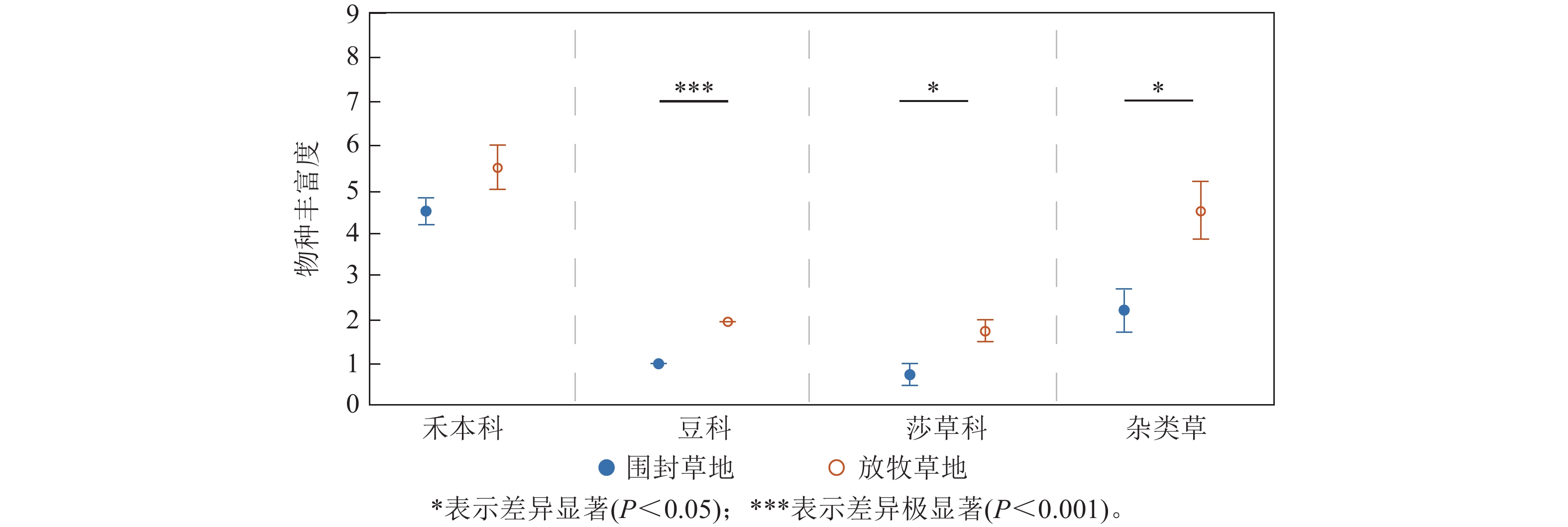
各功能群的物种丰富度对于2种处理的响应存在差异(图2)。其中,禾本科的物种丰富度在围封草地和放牧草地差异不显著,在围封草地有4.50种,比放牧草地少1.00种;豆科植物的物种丰富度在围封草地和放牧草地具有极显著差异(P<0.001),在围封草地为1种,在放牧草地为2种;放牧草地莎草科和杂类草的物种丰富度均显著高于围封草地(P<0.05),在放牧草地分别为1.75和4.50种,在围封草地分别为0.75和2.25种。
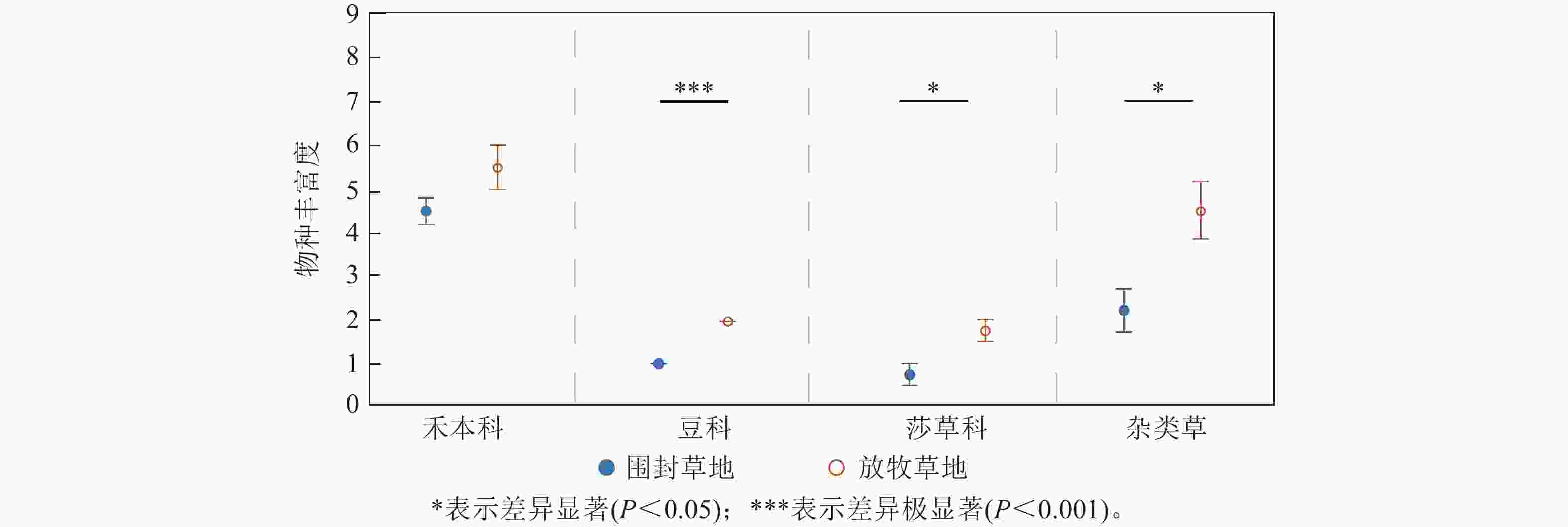
图 2 草地群落各功能群物种丰富度在围封草地和放牧草地的差异
Figure 2. Difference of species richness of each functional group in grassland community between fencing and grazing grassland
-
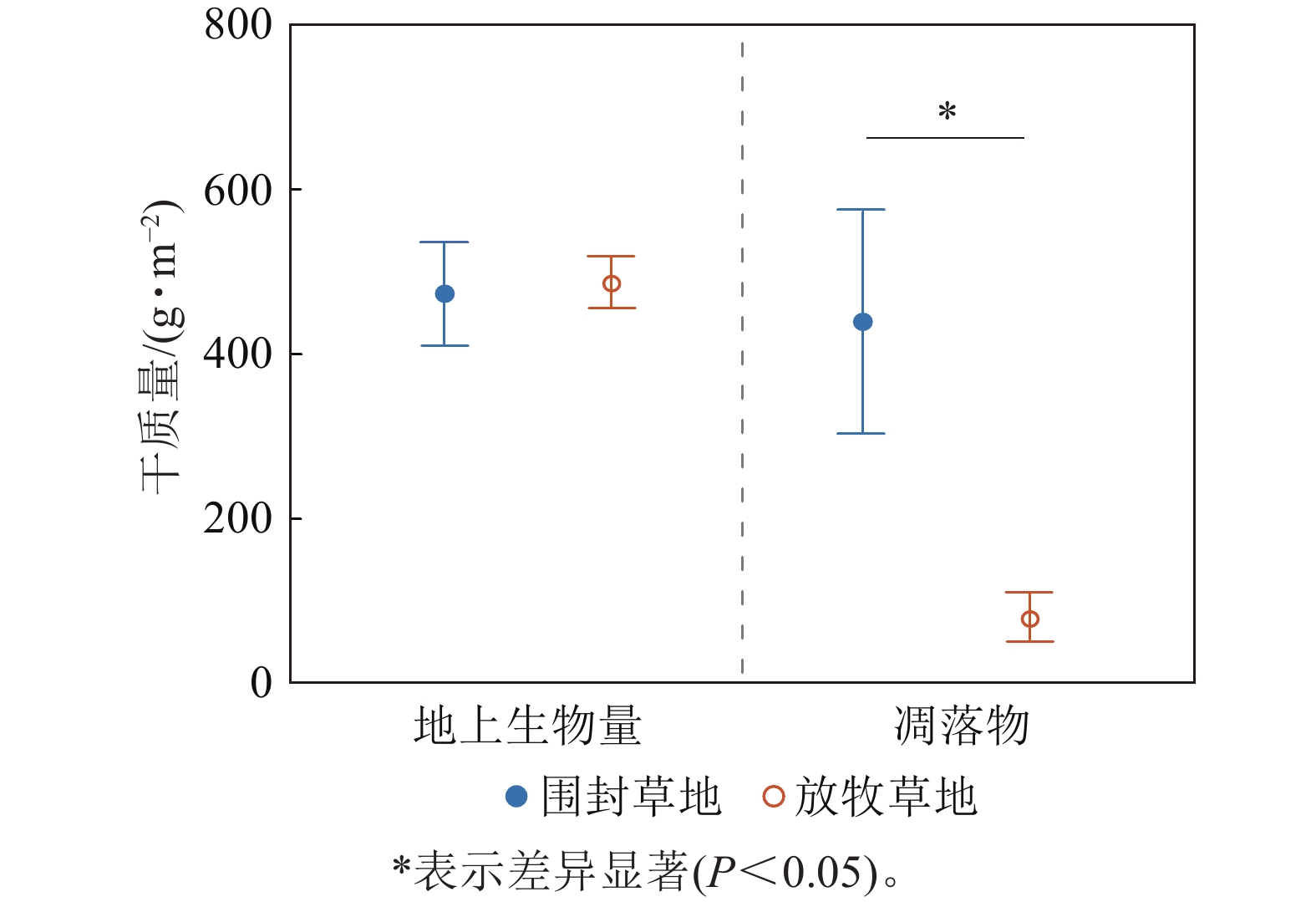
总体来看,草地群落总地上生物量在围封草地和放牧草地未表现出显著差异(P=0.84),围封草地的地上生物量干质量为468.91 g·m−2,放牧草地为483.18 g·m−2;但凋落物干质量在围封草地和放牧草地存在显著差异(P=0.04),围封草地凋落物干质量为434.82 g·m−2,约为放牧草地的5.5倍(图3)。
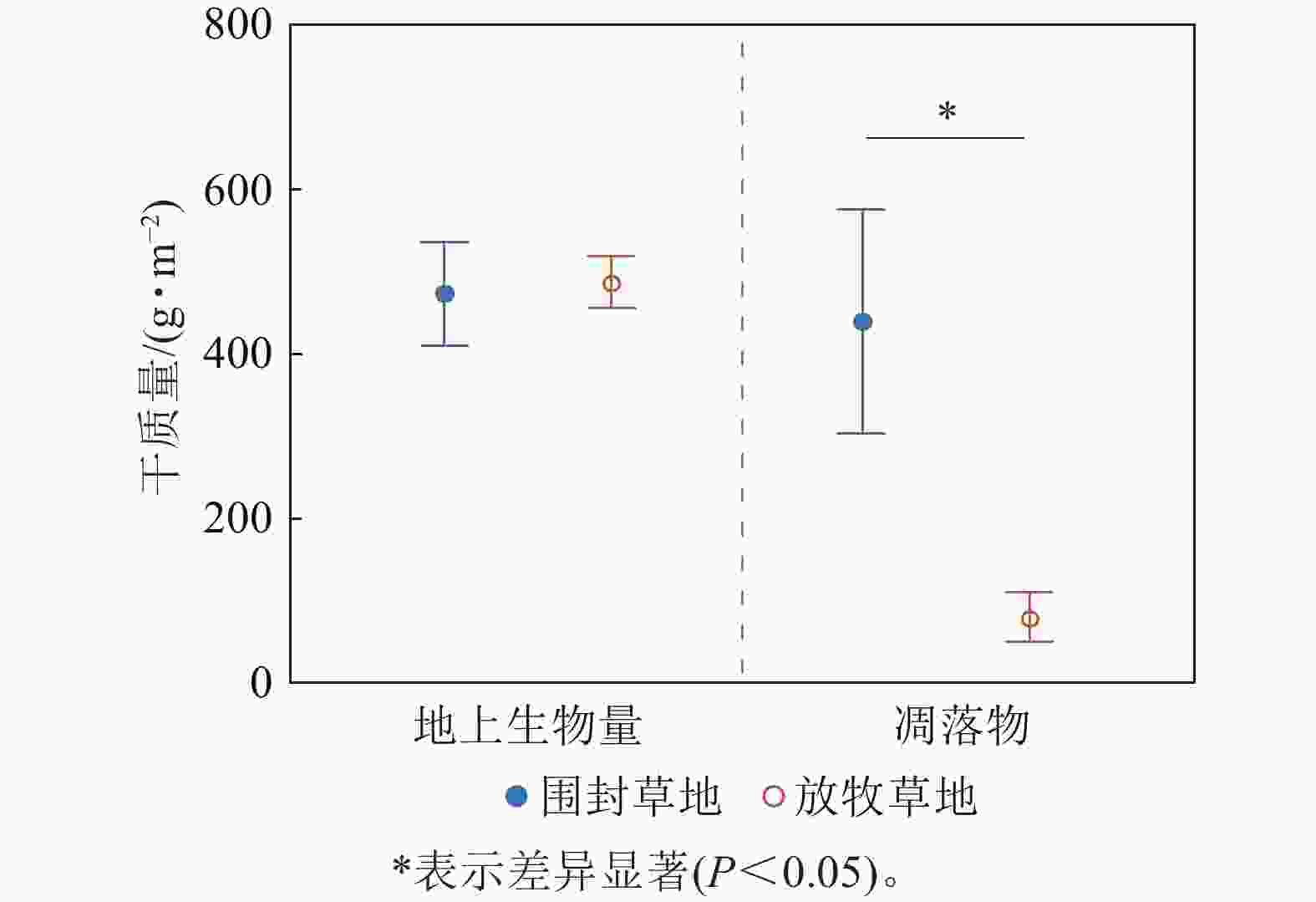
图 3 草地群落地上生物量与凋落物干质量在围封草地与放牧草地的差异
Figure 3. Difference of aboveground biomass and litter dry weight of grassland community between fencing and grazing grassland
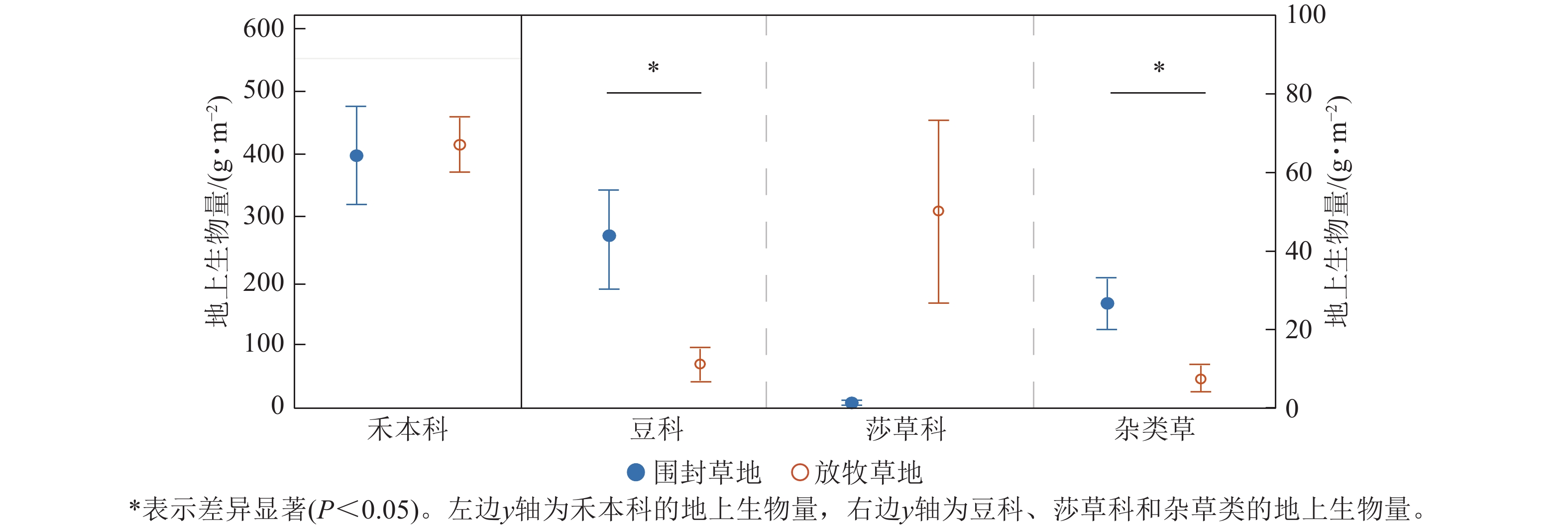
由图4可知:各功能群的地上生物量在围封草地和放牧草地表现出不同的结果。禾本科的地上生物量是总地上生物量的主要部分,但其在围封草地和放牧草地不存在显著差异(P=0.85),围封草地为397.51 g·m−2,占围封草地总地上生物量的84.77%,放牧草地为414.26 g·m−2,占放牧草地总地上生物量的85.74%。豆科与杂类草的地上生物量均表现为围封草地显著高于放牧草地(P<0.05),在围封草地分别为43.49和26.58 g·m−2,在放牧草地分别为11.23和7.63 g·m−2。莎草类则呈相反趋势,但差异未达显著水平。
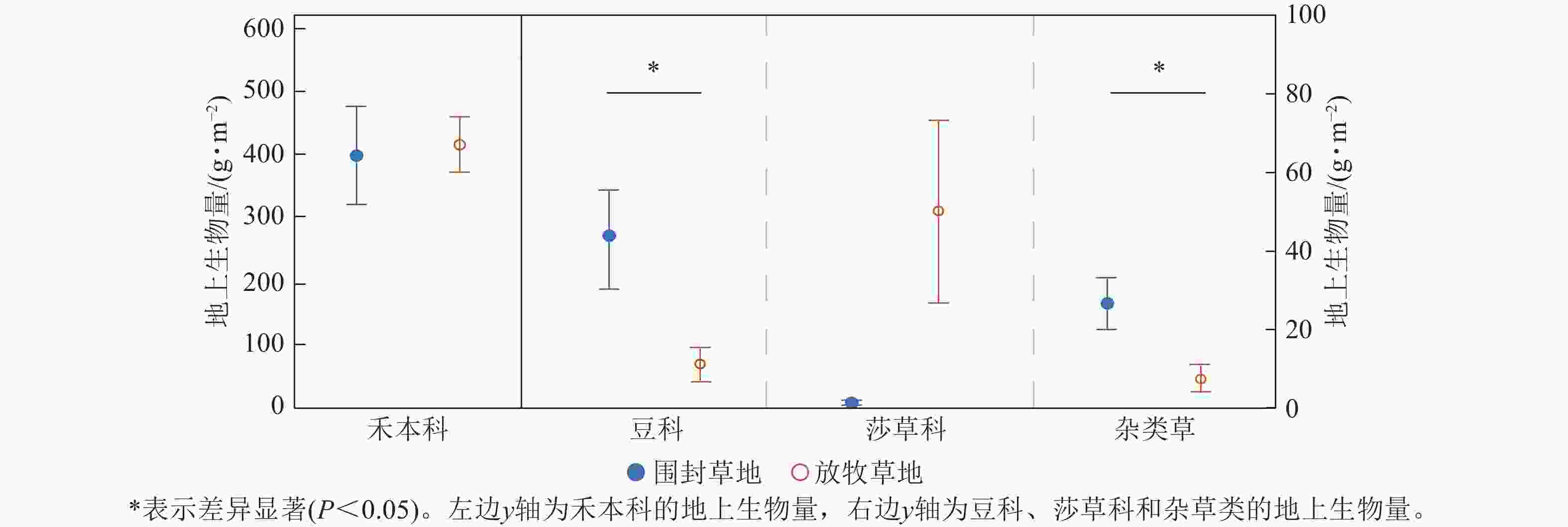
图 4 草地群落各功能群地上生物量在围封草地和放牧草地的差异
Figure 4. Difference of aboveground biomass of each functional group in grassland community between fencing and grazing grassland
-
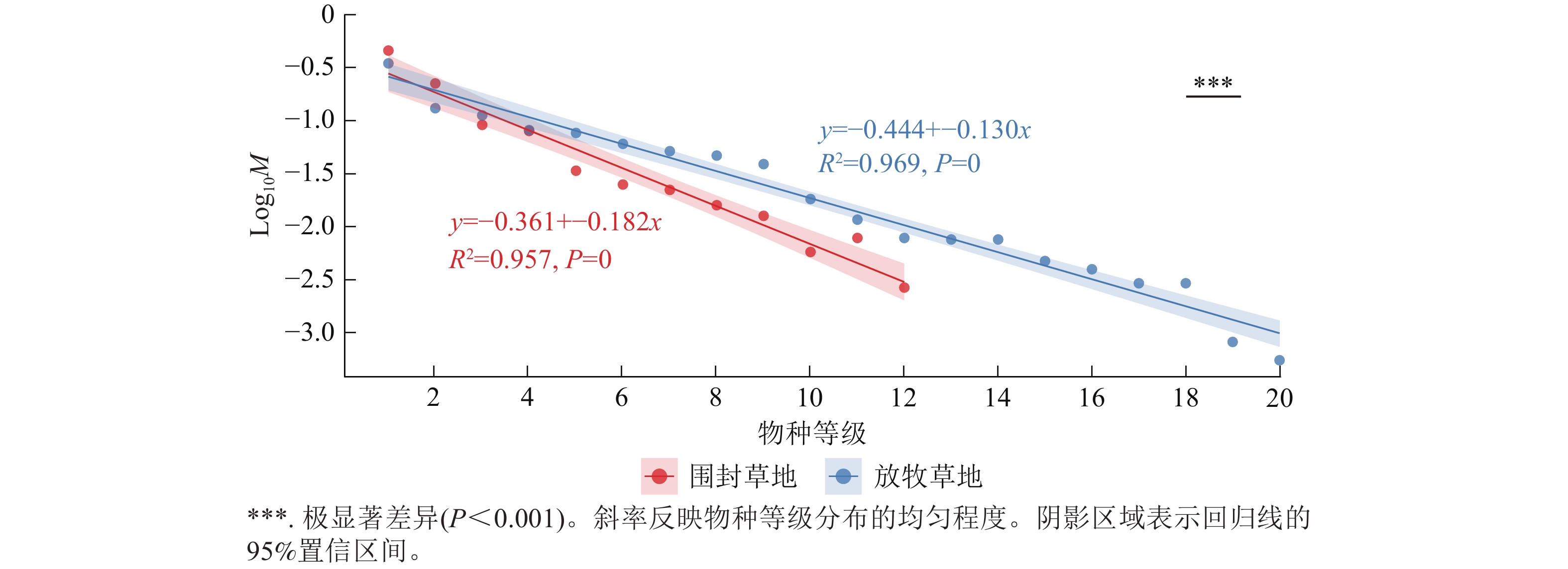
回归分析表明:草地植物群落物种分布均匀度在围封草地和放牧草地存在显著差异(图5)。围封草地和放牧草地的回归模型分别可以解释 95.7% 与 96.9% 的数据变异,拟合效果良好。其中,围封草地的回归斜率为−0.182,放牧草地为−0.130,两者差异极显著(P<0.001)。
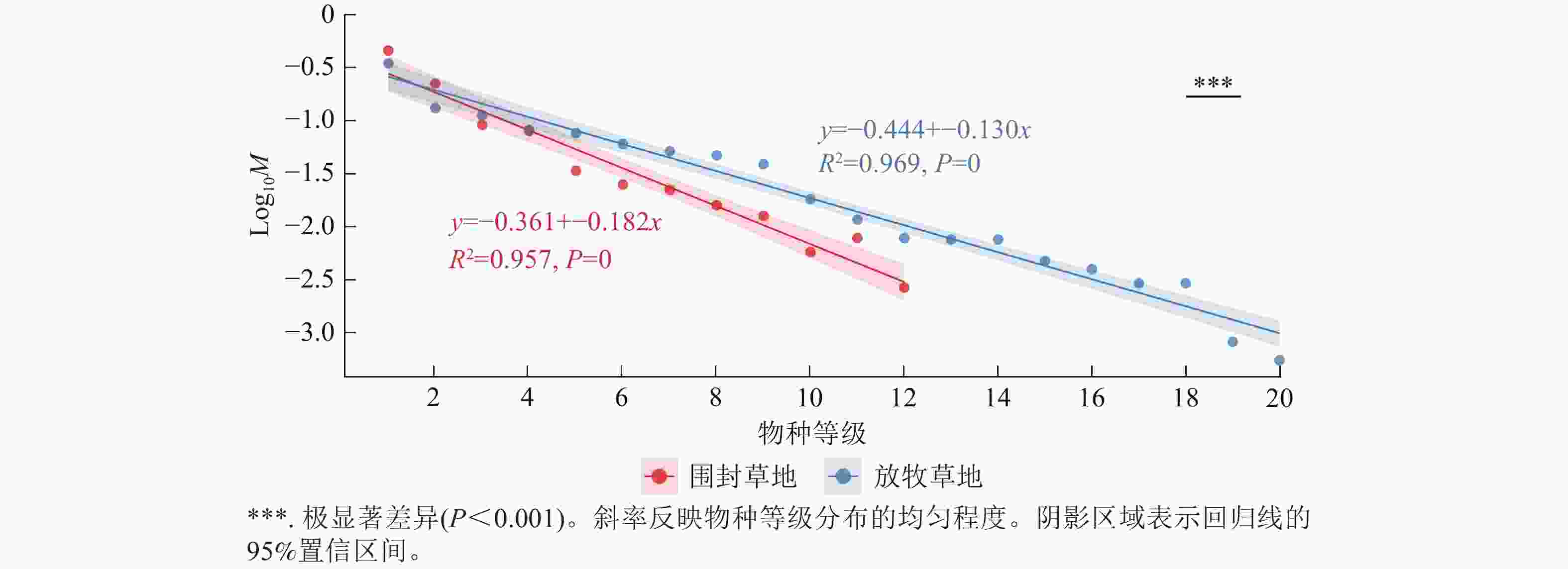
图 5 围封草地和放牧草地的植物群落物种等级-多度图
Figure 5. Plant community species level-abundance map of fencing and grazing grassland
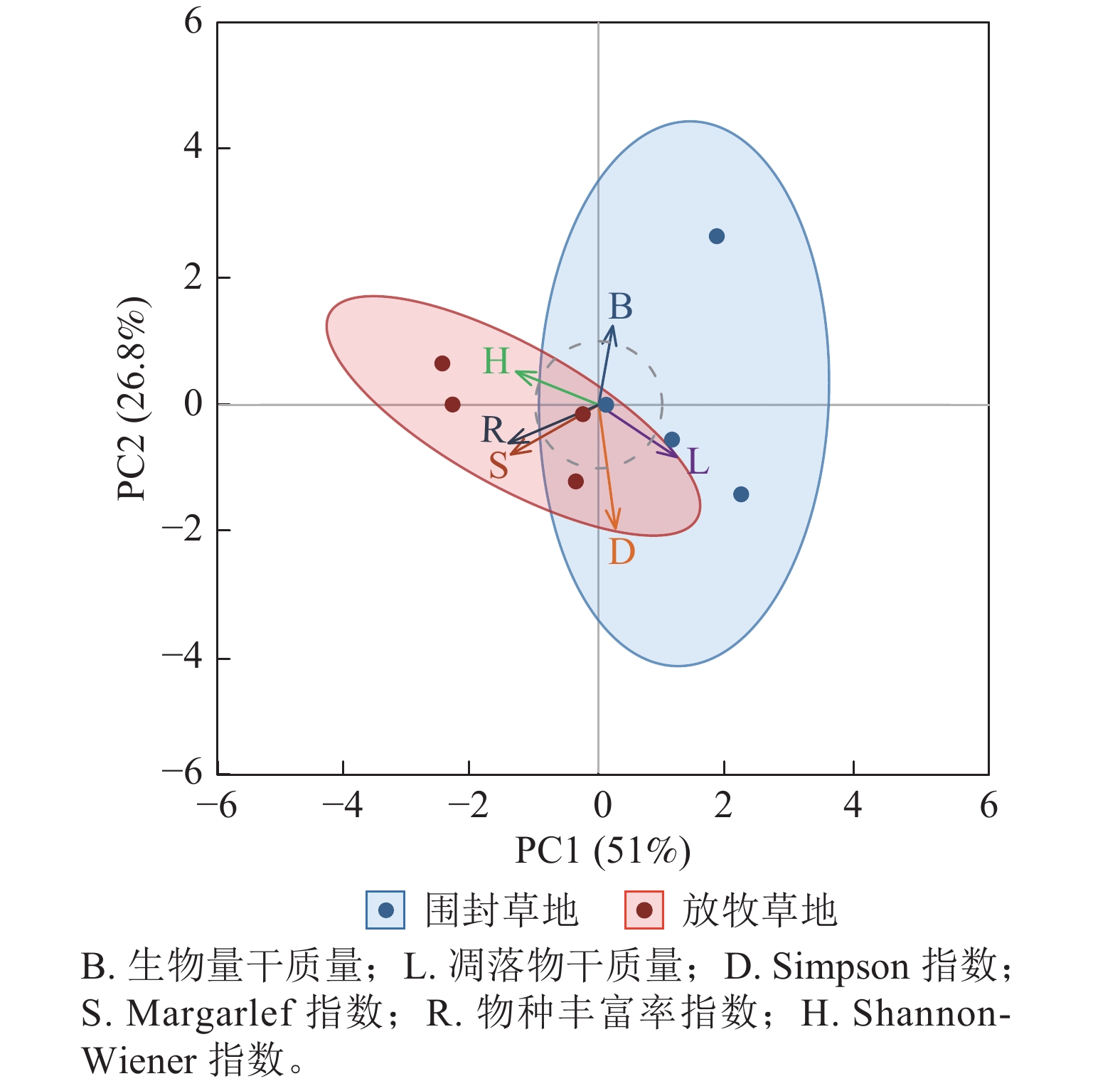
主成分分析表明:围封与放牧处理下草地群落结构存在明显分异,其中,围封更有利于生产力的提升,而放牧则更有利于物种多样性的维持(图6)。第1主成分(PC1)与第2主成分(PC2)的方差贡献率分别为 51.0% 与 26.8%,累计解释方差为 77.8%。围封草地样本主要分布于 PC1 正方向与 PC2 中高值区域,与生物量、凋落物干质量高度相关。放牧草地样本则集中在 PC1 负方向与 PC2 中低值区域,与物种丰富度、Margalef指数和 Shannon-Wiener指数紧密关联。
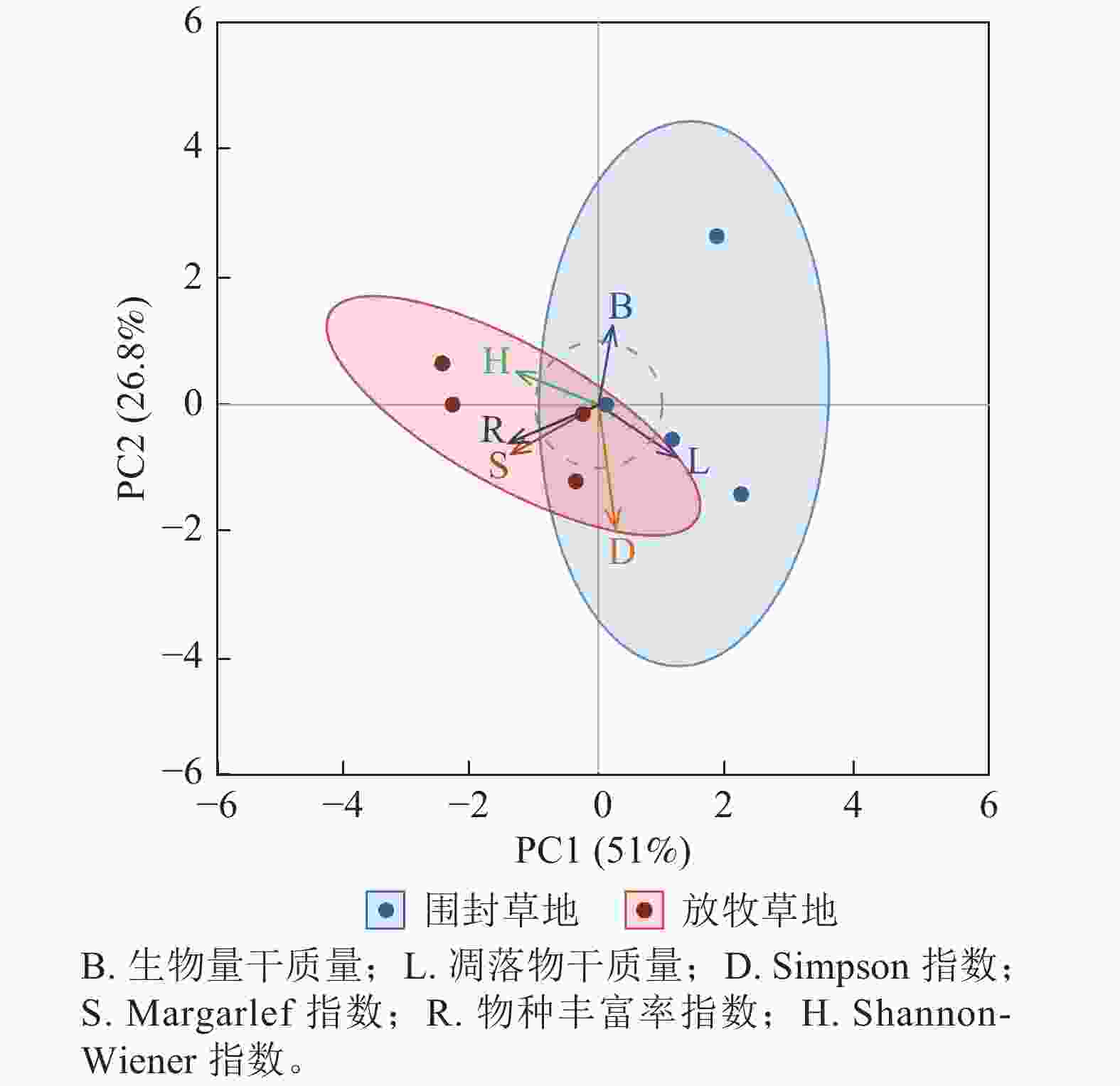
图 6 草地群落结构在围封草地与放牧草地的综合差异
Figure 6. Comprehensive difference of grassland community structure between fencing and grazing grassland
-
物种多样性作为生物多样性的核心组分,其对草地群落的结构与功能(如生产力与生态系统稳定性)产生关键的影响[31−32]。本研究发现:高寒草地群落经长期围封显著降低了总的物种多样性水平,这与“中度干扰假说”的预期相符。中度干扰理论的核心机制在于干扰排除后草地群落竞争格局发生改变。在放牧缺失下,草地群落主要受2种相互关联的抑制过程主导:一方面是物理化学抑制,凋落物的大量积累会抑制植物的定居与生长[33],从而降低群落的物种丰富度[34]。另一方面是生物竞争抑制,在长期缺乏家畜活动干扰的情况下,草地植物种间竞争关系会更为激烈,这将会导致植物群落中竞争力较弱的物种被排除在系统之外甚至消失[35];相反,在放牧草地,适度放牧作为一种中等强度干扰,可通过抑制草地优势种过度生长,为更多物种提供生存机会,从而维持了较高的物种多样性水平[36]。
另外,不同功能群物种多样性对围封的响应存在差异。禾本科植物作为草地的优势功能群,其物种丰富度在2种处理下无显著差异。传统研究认为:在放牧草地中,多年生禾草因家畜长期采食而生长受限[37],即合理的放牧会抑制禾草类植物的生长[38];相反,在围封草地,凋落物长期积累也会抑制其生长[39]。这2种压力可能会相互抵消,导致禾本科植物的物种丰富度在围封和放牧处理下无显著差异。放牧草地豆科、莎草科与杂类草的物种多样性均显著高于围封草地,其原因可能有以下2点。首先,适度放牧干扰会影响草地群落的资源再分配。根据生态位互补假说,当家畜采食减少多年生禾草的数量时,群落中个体较小的植物可获得更多光资源与养分,从而更有利于放牧草地中豆科、莎草科等功能群的建立与维持[40]。其次,在围封条件下,凋落物的长期积累会对各功能群产生不同程度的抑制作用。较厚的凋落物层通过物理阻隔和释放化感物质会抑制植物的种子萌发与幼苗建成,但是,其对低矮物种的抑制作用明显高于禾草类植物[41−42]。
-
物种多样性是维持草地生态系统稳定和生产力的基础[43]。草地植物地上生物量作为生态系统生产力的重要指标,是整个生态系统运行的能量基础和营养物质来源[44]。本研究表明:长期围封虽降低了总的物种多样性,但群落总的地上生物量并未随之降低,这一结果与经典生态学理论中有关物种多样性与生产力呈正相关的普遍认知[45]不尽一致。这一“解耦”现象可能源于多重生态过程的抵消效应。首先,研究样地在围封前处于非退化状态,不同于多数以退化草地为起点开展的相关研究,其初始条件可能削弱了围封对生产力的提升效应。其次,如前文所讨论,围封和放牧可能会通过不同的途径对草地的生产力产生抑制作用。在围封草地,凋落物积累通过改变微环境和资源可利用性抑制植物生长,从而降低草地的生产力[46];在放牧草地,家畜的采食与践踏等行为同样会抑制植物生长[47],导致放牧草地生物量降低。这2种反向抑制力的平衡,可能导致总的地上生物量在围封和放牧处理下未出现显著差异。
另外,值得关注的是,围封草地中豆科与杂类草的物种多样性虽较低,但其生物量却相对较高。这一看似矛盾的现象可能源于凋落物的筛选作用与竞争释放效应(competitive release effect),即凋落物积累抑制了多数竞争力较弱物种的定居,而存活下来的少数豆科与杂类草物种可能因种间竞争减弱而获得更多资源,从而表现出更高的个体生物量[48]。此外,禾本科功能群的地上生物量在2种处理草地间无显著差异,这一结果再次反映了围封与放牧可能对禾草生长均产生一定的抑制效应,即放牧通过采食直接降低其生物量,而围封则通过凋落物积累间接抑制其更新与生长。两者在一定程度上相互抵消,最终导致禾草生物量未发生显著变化。
-
本研究的物种多度分布模型表明:围封显著降低了草地植物群落物种均匀度。在围封草地,植物物种总数的减少以及资源向少数优势种的集中,导致物种间个体数量分配更为不均,即会降低草地群落物种均匀度。这一规律在贺聪等[49]于青海海北的研究中得到了印证:长期的围封使得禾草在群落中的占比持续增加,而豆科、莎草、杂类草等其他功能群的占比则逐渐降低。主成分分析结果进一步在多维特征空间直观印证了这种分异。这种分异意味着,长期围封与适度放牧并非简单导致草地退化与恢复,而是指向了草地的不同发展方向。凋落物积累作为关键的物理和化学因素,导致其在2种处理下的差异可能归因于以下机制:高寒地区低温条件会显著抑制围封草地凋落物的分解,导致有机物质大量滞留[50];相反,放牧草地中家畜的采食与践踏活动能够加速凋落物的破碎与分解,从而减少凋落物积累[51]。此外,这一群落分异结果与本研究有关围封降低物种多样性但维持了地上生物量,而放牧促进物种多样性的结果相印证,表明围封与放牧通过差异化途径影响草地群落的结构与功能。
生物多样性是生态系统抵抗力、恢复力以及适应环境变化能力的基础[52]。在本研究中,围封与放牧处理对于草地地上生物量的影响并不明显,但显著影响了植物多样性水平。根据该结果可知,在非退化或轻度退化的高寒草地上,长期围封可能削弱其生态系统的抵抗力与恢复力,并非是一种可持续的草地管理策略。基于本研究的结果与“中度干扰假说”[53−54],对于未退化草地,应采取放牧压力适中的管理措施,如季节性轮牧、划区轮牧或适度割草等。这不仅能够维持较高的物种多样性,还能保障草地生态系统的生产功能与稳定。
-
本研究比较研究了长期围封与适度放牧对未退化高寒草地生态系统的影响。结果表明:围封显著降低了草地群落的物种多样性,具体表现为Margalef指数与物种丰富度的下降,其中豆科、莎草科及杂类草功能群的响应尤为明显。草地群落总地上生物量对于围封处理的响应不明显,但改变了不同功能群间的生物量分配,如增加了豆科与杂类草的生物量积累。同时,围封改变了草地群落物种多度分配格局,导致群落均匀度降低,并形成高生产积累的资源利用模式。适度放牧有利于草地群落维持较高的物种多样性水平与群落结构和功能的稳定。因此,对于未退化或轻度退化的高寒草地,长期围栏封育并非理想的草地管理策略,不利于草地生态系统的健康与稳定。
Differentiation of plant community characteristics in undegraded alpine grasslands under enclosure and grazing treatments
-
摘要:
目的 探索科学合理的草地管理模式,维持草地生态系统结构和功能稳定。 方法 以青藏高原东缘高寒草地为对象,通过长期围封与适度放牧进行对比试验,探究了2种管理方式对草地植物群落结构、物种多样性及生产力的影响。 结果 ①与放牧相比,长期围封显著降低了草地群落的植物多样性(P<0.05),其中豆科Fabaceae、莎草科Cyperaceae和杂类草功能群的物种丰富度下降最为明显。②围封未显著改变草地总地上生物量,但显著改变了群落各功能群间的生物量分配,如显著促进了豆科与杂类草的生物量积累(P<0.05)。同时,围封也导致凋落物大量积累(约为放牧草地的5.5倍)。③相比适度放牧草地,长期围封极显著降低了草地植物物种的均匀度(P<0.001),导致植物群落结构和功能发生明显分异。 结论 长期围封驱动草地植物群落向高生产力,即生物量与凋落物积累的方向发展,而放牧则有利于维持较高的物种多样性水平。对于未退化高寒草地,适度放牧更有利于维持物种多样性和生态系统功能的稳定。图6表1参54 Abstract:Objective The objective is to explore a scientific and reasonable grassland management model to maintain the stability of grassland ecosystem structure and function. Method A comparative experiment involving long-term enclosure and moderate grazing was conducted to investigate the effects of the two management practices on grassland plant community structure, species diversity, and productivity in an alpine grassland on the eastern margin of the Qinghai-Tibet Plateau. Result (1) Compared with grazing, long-term enclosure significantly reduced overall plant diversity of grassland communities (P< 0.05), with the most significant declines observed in the species richness of legumes, sedges, and forbs. (2) Enclosure did not significantly alter total aboveground biomass, but it significantly shifted biomass allocation among functional groups, significantly promoting biomass accumulation in legumes and forbs (P<0.05). Additionally, enclosure led to substantial litter accumulation (about 5.5 times that of grazed grasslands). (3) Long-term enclosure significantly reduced species evenness compared with moderate grazing (P<0.001), resulting in a significant differentiation in plant community structure and function. Conclusion Long-term enclosure drives grassland plant communities toward high productivity characterized by increased biomass and litter accumulation, while grazing is beneficial for the maintenance of a high level of species diversity. For non-degraded alpine grasslands, moderate grazing is more conducive to maintaining species diversity and stabilizing ecosystem functions. [Ch, 6 fig. 1 tab. 54 ref.] -
Key words:
- alpine grassland /
- long-term enclosure /
- moderate grazing /
- community structure /
- species diversity
-
图 1 草地植物群落物种多样性指数在围封草地与放牧草地的差异
Figure 1 Difference of species diversity index in grassland community between fencing and grazing grassland
图 2 草地群落各功能群物种丰富度在围封草地和放牧草地的差异
Figure 2 Difference of species richness of each functional group in grassland community between fencing and grazing grassland
图 3 草地群落地上生物量与凋落物干质量在围封草地与放牧草地的差异
Figure 3 Difference of aboveground biomass and litter dry weight of grassland community between fencing and grazing grassland
图 4 草地群落各功能群地上生物量在围封草地和放牧草地的差异
Figure 4 Difference of aboveground biomass of each functional group in grassland community between fencing and grazing grassland
图 5 围封草地和放牧草地的植物群落物种等级-多度图
Figure 5 Plant community species level-abundance map of fencing and grazing grassland
图 6 草地群落结构在围封草地与放牧草地的综合差异
Figure 6 Comprehensive difference of grassland community structure between fencing and grazing grassland
表 1 草地群落植物物种组成及重要值在围封草地与放牧草地的差异
Table 1. Difference of plant species composition and its important value between fencing and grazing grassland in grassland community
功能群 物种 重要值/% 功能群 物种 重要值/% 围封草地 放牧草地 围封草地 放牧草地 禾本科 赖草 60.45±2.36 54.33±5.25 杂类草 蒲公英 36.95±0.90 19.18±6.39 垂穗披碱草 39.35±1.40 36.25±1.66 球花蒿 4.25±4.25 2.13±2.13 羊茅 20.23±6.81 9.18±5.33 茵陈蒿 − 8.63±4.98 青海仲彬草 21.10±7.16 12.58±7.26 麻花艽 9.38±5.41 2.15±2.15 紫花针茅 37.33±1.44** 10.70±6.34 高山唐松草 2.28±2.28 − 冰草 − 25.45±10.35 棉毛茛 − 34.33±0.18 − 42.00±2.61 平车前 − 2.50±2.50 草地早熟禾 − 3.28±3.28 翻白委陵菜 − 19.73±6.59 豆科 青海苜蓿 50.73±2.91* 37.83±1.59 高山韭 2.28±2.28 − 黄花棘豆 − 34.03±0.09 鳞茎堇菜 − 18.93±6.31 莎草科 青藏薹草 19.75±6.59 24.48±8.44 矮生嵩草 − 37.70±0.48 说明:数据为平均值±标准误。−.样地内不存在的物种。*表示与放牧草地间存在显著性差异(P<0.05)。**表示与放牧草地间差异极显著(P<0.01)。禾本科Poaceae;豆科Fabaceae;莎草科Cyperaceae。冰草Agropyron cristatum;  下载: 导出CSV
下载: 导出CSV
-
[1] 白永飞, 黄建辉, 郑淑霞, 等. 草地和荒漠生态系统服务功能的形成与调控机制[J]. 植物生态学报, 2014, 38(2): 93−102. BAI Yongfei, HUANG Jianhui, ZHENG Shuxia, et al. Drivers and regulating mechanisms of grassland and desert ecosystem services[J]. Chinese Journal of Plant Ecology, 2014, 38(2): 93−102. DOI: 10.3724/SP.J.1258.2014.00009. [2] BAI Yongfei, COTRUFO M F. Grassland soil carbon sequestration: current understanding, challenges, and solutions[J]. Science, 2022, 377(6606): 603−608. DOI: 10.1126/science.abo2380. [3] 张扬建, 朱军涛, 沈若楠, 等. 放牧对草地生态系统影响的研究进展[J]. 植物生态学报, 2020, 44(5): 553−564. ZHANG Yangjian, ZHU Juntao, SHEN Ruonan, et al. Research progress on the effects of grazing on grassland ecosystem[J]. Chinese Journal of Plant Ecology, 2020, 44(5): 553−564. DOI: 10.17521/cjpe.2019.0314. [4] 慕宗杰. 草地生态系统的保护及治理对策[J]. 畜牧与饲料科学, 2009, 30(2): 48−50. MU Zongjie. Protection and control countermeasures of giassland ecosystem[J]. Animal Husbandry and Feed Science, 2009, 30(2): 48−50. DOI: 10.3969/j.issn.1672-5190.2009.02.022. [5] CHEN Xiaolong, LI Xu, SHAN Xiaofeng, et al. The effects of sand-fixing agents and Trichoderma longibrachiatum on soil quality and alfalfa growth in wind-sand soil[J]. Agriculture, 2025, 15(23): 2463. DOI: 10.3390/agriculture15232463. [6] 侯星辰, 鲁绍伟, 向昌林, 等. 不同母质温带草地植物群落多样性对人为干扰的响应[J]. 应用生态学报, 2022, 33(8): 2153−2160. HOU Xingchen, LU Shaowei, XIANG Changlin, et al. Responses of plant community diversity to human disturbance in temperate grassland with different soil parent materials[J]. Chinese Journal of Applied Ecology, 2022, 33(8): 2153−2160. DOI: 10.13287/j.1001-9332.202208.015. [7] 白永飞, 赵玉金, 王扬, 等. 中国北方草地生态系统服务评估和功能区划助力生态安全屏障建设[J]. 中国科学院院刊, 2020, 35(6): 675−689. BAI Yongfei, ZHAO Yujin, WANG Yang, et al. Assessment of ecosystem services and ecological regionalization of grasslands support establishment of ecological security barriers in northern China[J]. Bulletin of the Chinese Academy of Sciences, 2020, 35(6): 675−689. DOI: 10.16418/j.issn.1000-3045.20200515003. [8] 李禄军, 曾德慧, 于占源, 等. 氮素添加对科尔沁沙质草地物种多样性和生产力的影响[J]. 应用生态学报, 2009, 20(8): 1838−1844. LI Lujun, ZENG Dehui, YU Zhanyuan, et al. Effects of nitrogen addition on grassland species diversity and productivity in Keerqin Sandy Land[J]. Chinese Journal of Applied Ecology, 2009, 20(8): 1838−1844. DOI: 10.13287/j.1001-9332.2009.0302. [9] 郑伟, 董全民, 李世雄, 等. 放牧强度对环青海湖高寒草原群落物种多样性和生产力的影响[J]. 草地学报, 2012, 20(6): 1033−1038. ZHENG Wei, DONG Quanmin, LI Shixiong, et al. Impact of grazing intensities on community biodiversity and productivity of alpine grassland in Qinghai Lake Region[J]. Acta Agrestia Sinica, 2012, 20(6): 1033−1038. [10] GANGULI A C, O’ROURKE M E. How vulnerable are rangelands to grazing?[J]. Science, 2022, 378(6622): 834. DOI: 10.1126/science.add4278. [11] XIANG Mingxue, WU Junxi, DUO L, et al. The effects of grazing and fencing on grassland productivity and diversity in alpine grassland ecosystem in the Tibetan highland[J]. Global Ecology and Conservation, 2023, 44: e02495. DOI: 10.1016/j.gecco.2023.e02495. [12] BELLARD C, BERTELSMEIER C, LEADLEY P, et al. Impacts of climate change on the future of biodiversity[J]. Ecology Letters, 2012, 15(4): 365−377. DOI: 10.1111/j.1461-0248.2011.01736.x. [13] BULLOCK J M, HILL B C, DALE M P, et al. An experimental study of the effects of sheep grazing on vegetation change in a species-poor grassland and the role of seedling recruitment into gaps[J]. Journal of Applied Ecology, 1994, 31(3): 493−507. DOI: 10.2307/2404445. [14] 史志华, 刘前进, 张含玉, 等. 近十年土壤侵蚀与水土保持研究进展与展望[J]. 土壤学报, 2020, 57(5): 1117−1127. SHI Zhihua, LIU Qianjin, ZHANG Hanyu, et al. Study on soil erosion and conservation in the past 10 years: progress and prospects[J]. Acta Pedologica Sinica, 2020, 57(5): 1117−1127. DOI: 10.11766/trxb202002240070. [15] AKIYAMA T, KAWAMURA K. Grassland degradation in China: methods of monitoring, management and restoration[J]. Grassland Science, 2007, 53(1): 1−17. DOI: 10.1111/j.1744-697x.2007.00073.x. [16] 晏和飘, 李文龙, 梁天刚, 等. 青藏高原退化高寒草地恢复对不同措施响应的Meta分析[J]. 草地学报, 2021, 29(S1): 190−198. YAN Hepiao, LI Wenlong, LIANG Tiangang, et al. Effects of different restoration measures on degraded alpine grasslands of the Tibetan Plateau: a meta-analysis[J]. Acta Agrestia Sinica, 2021, 29(S1): 190−198. DOI: 10.11733/j.issn.1007-0435.2021.Z1.022. [17] SUN J, CHENG G W, LI W P. Meta-analysis of relationships between environmental factors and aboveground biomass in the alpine grassland on the Tibetan Plateau[J]. Biogeosciences, 2013, 10(3): 1707−1715. DOI: 10.5194/bg-10-1707-2013. [18] JING Zhaobin, CHENG Jimin, SU Jishuai, et al. Changes in plant community composition and soil properties under 3-decade grazing exclusion in semiarid grassland[J]. Ecological Engineering, 2014, 64: 171−178. DOI: 10.1016/j.ecoleng.2013.12.023. [19] XIONG Dingpeng, SHI Peili, SUN Yinliang, et al. Effects of grazing exclusion on plant productivity and soil carbon, nitrogen storage in alpine meadows in northern Tibet, China[J]. Chinese Geographical Science, 2014, 24(4): 488−498. DOI: 10.1007/s11769-014-0697-y. [20] SHANG Zhanhuan, CAO Jingjuan, GUO Ruiying, et al. Effect of enclosure on soil carbon, nitrogen and phosphorus of alpine desert rangeland[J]. Land Degradation & Development, 2017, 28(4): 1166−1177. DOI: 10.1002/ldr.2283. [21] PATRICK L B, FRASER L H, KERSHNER M W. Large-scale manipulation of plant litter and fertilizer in a managed successional temperate grassland[J]. Plant Ecology, 2008, 197(2): 183−195. DOI: 10.1007/s11258-007-9369-7. [22] 张艺斐, 王兴, 潘海珠, 等. 中国北方荒漠草原长期围栏封育恢复效应及其环境依赖性[J]. 生态学报, 2025, 45(17): 8529−8543. ZHANG Yifei, WANG Xing, PAN Haizhu, et al. Long-term enclosure restoration effects and their environmental dependencies in desert steppes of northern China[J]. Acta Ecologica Sinica, 2025, 45(17): 8529−8543. DOI: 10.20103/j.stxb.202406061313. [23] 江康威, 张青青, 王亚菲, 等. 放牧干扰下天山北坡中段植物功能群特征及其与土壤环境因子的关系[J]. 植物生态学报, 2024, 48(6): 701−718. JIANG Kangwei, ZHANG Qingqing, WANG Yafei, et al. Characteristics of plant functional groups and the relationships with soil environmental factors in middle part of northern slope of Tianshan Mountains under different grazing intensities[J]. Chinese Journal of Plant Ecology, 2024, 48(6): 701−718. DOI: 10.17521/cjpe.2023.0225. [24] 吴建波, 包晓影, 李洁, 等. 不同围封年限对典型草原群落及大针茅种群特征的影响[J]. 草地学报, 2010, 18(4): 490−495. WU Jianbo, BAO Xiaoying, LI Jie, et al. Influence of fencing duration on community and population of Stipa grandis in a typical steppe[J]. Acta Agrectir Sinica, 2010, 18(4): 490−495. DOI: 10.3969/j.issn.1007-0435.2010.04.003. [25] 彭珍, 肖红, 潘涛涛, 等. 模拟践踏和降雨对天祝高寒区垂穗披碱草有性繁殖的影响[J]. 中国草地学报, 2020, 42(2): 14−21. PENG Zhen, XIAO Hong, PAN Taotao, et al. Effects of simulated trampling and rainfall on sexual reproduction of Elymus nutans in Tianzhu alpine meadow[J]. Chinese Journal of Grassland, 2020, 42(2): 14−21. DOI: 10.16742/j.zgcdxb.20190132. [26] 施颖, 胡廷花, 高红娟, 等. 两种放牧模式下高寒草甸群落植被构成及稳定性特征[J]. 草业学报, 2019, 28(9): 1−10. SHI Ying, HU Tinghua, GAO Hongjuan, et al. The community vegetation composition and stability characteristics of alpine meadow under two grazing modes[J]. Acta Prataculturae Sinica, 2019, 28(9): 1−10. DOI: 10.11686/cyxb2018571. [27] 中华人民共和国农业农村部. NY/T 3647—2020 草食家畜羊单位换算[S]. 北京: 中国农业出版社, 2020. Ministry of Agriculture and Rural Affairs of the People’s Republic of China. Conversion of Herbivore Livestock Unit: NY/T 3647−2020[S]. Beijing: China Agriculture Press, 2020. [28] WHITTAKER R H. Evolution and measurement of species diversity[J]. Taxon, 1972, 21(2/3): 213−251. [29] 马克平. 生物群落多样性的测度方法Ⅰα多样性的测度方法(上)[J]. 生物多样性, 1994, 2(3): 162−168. MA Keping. The measurement method of biological community diversity Ⅰ The measurement method of α diversity (Ⅰ)[J]. Chinese Biodiversity, 1994, 2(3): 162−168. DOI: 10.17520/biods.1994027. [30] 马克平, 刘玉明. 生物群落多样性的测度方法Ⅰα多样性的测度方法(下)[J]. 生物多样性, 1994, 2(4): 231−239. MA Keping, LIU Yuming. The measurement method of biological community diversity Ⅰ The measurement method of α diversity (Ⅱ)[J]. Chinese Biodiversity, 1994, 2(4): 231−239. DOI: 10.17520/biods.1994038. [31] 张钦弟, 卫伟, 陈利顶, 等. 黄土高原草地土壤水分和物种多样性沿降水梯度的分布格局[J]. 自然资源学报, 2018, 33(8): 1351−1362. ZHANG Qindi, WEI Wei, CHEN Liding, et al. Spatial variation of soil moisture and species diversity patterns along a precipitation gradient in the grasslands of the Loess Plateau[J]. Journal of Natural Resources, 2018, 33(8): 1351−1362. DOI: 10.31497/zrzyxb.20170726. [32] MAESTRE F T, QUERO J L, GOTELLI N J, et al. Plant species richness and ecosystem multifunctionality in global drylands[J]. Science, 2012, 335(6065): 214−218. DOI: 10.1126/science.1215442. [33] HOU Dongjie, HE Weiming, LIU Changcheng, et al. Litter accumulation alters the abiotic environment and drives community successional changes in two fenced grasslands in Inner Mongolia[J]. Ecology and Evolution, 2019, 9(16): 9214−9224. DOI: 10.1002/ece3.5469. [34] ZHANG Yiwen, CHANG Xiaofeng, ZHANG Yuwei, et al. Litter accumulation suppresses grass production but facilitates shrub expansion in a long-term fenced grassland[J]. Ecosphere, 2024, 15(4): e4844. DOI: 10.1002/ECS2.4844. [35] 段敏杰, 高清竹, 万运帆, 等. 放牧对藏北紫花针茅高寒草原植物群落特征的影响[J]. 生态学报, 2010, 30(14): 3892−3900. DUAN Minjie, GAO Qingzhu, WAN Yunfan, et al. Effect of grazing on community characteristics and species diversity of Stipa purpurea alpine grassland in Northern Tibet[J]. Acta Ecologica Sinica, 2010, 30(14): 3892−3900. DOI: 10.20103/j.stxb.2010.14.027. [36] WANG Chao, TANG Yujia. A global meta-analyses of the response of multi-taxa diversity to grazing intensity in grasslands[J]. Environmental Research Letters, 2019, 14(11): 114003. DOI: 10.1088/1748-9326/ab4932. [37] 王启基, 周兴民. 高寒矮嵩草草甸禾草种群的生长发育节律及环境适应性[J]. 植物生态学与地植物学学报, 1991, 15(2): 168−176. WANG Qiji, ZHOU Xingmin. The growth rhythm of the grasses populations and their adaptability of environment in Kobresia humilis meadow[J]. Chinese Journal of Plant Ecology, 1991, 15(2): 168−176. [38] 汪诗平, 王艳芬, 陈佐忠. 气候变化和放牧活动对糙隐子草种群的影响[J]. 植物生态学报, 2003, 27(3): 337−343. WANG Shiping, WANG Yanfen, CHEN Zuozhong. Effect of climate change and grazing on populations of Cleistogenes squarrosa in Inner Mongolia steppe[J]. Acta Phytoecologica Sinica, 2003, 27(3): 337−343. [39] FOSTER B L, GROSS K L. Partitioning the effects of plant biomass and litter on Andropogon gerardi in old-field vegetation[J]. Ecology, 1997, 78(7): 2091−2104. DOI: 10.2307/2265947. [40] SUDING K N, LAVOREL S, CHAPIN III F S, et al. Scaling environmental change through the community-level: a trait-based response-and-effect framework for plants[J]. Global Change Biology, 2008, 14(5): 1125−1140. DOI: 10.1111/j.1365-2486.2008.01557.x. [41] 徐来仙, 郭秋菊, 姚兰, 等. 凋落物物理阻隔对水杉幼苗出土和早期生长的影响[J]. 浙江农林大学学报, 2022, 39(5): 1018−1027. XU Laixian, GUO Qiuju, YAO Lan, et al. Effect of litter physical barrier on emergence and early growth of Metasequoia glyptostroboides seedlings[J]. Journal of Zhejiang A&F University, 2022, 39(5): 1018−1027. DOI: 10.11833/j.issn.2095-0756.20210704. [42] LETTS B, LAMB E G, MISCHKOLZ J M, et al. Litter accumulation drives grassland plant community composition and functional diversity via leaf traits[J]. Plant Ecology, 2015, 216(3): 357−370. DOI: 10.1007/s11258-014-0436-6. [43] 宋明华, 刘丽萍, 陈锦, 等. 草地生态系统生物和功能多样性及其优化管理[J]. 生态环境学报, 2018, 27(6): 1179−1188. SONG Minghua, LIU Liping, CHEN Jin, et al. Biology, multi-function and optimized management in grassland ecosystem[J]. Ecology and Environment Sciences, 2018, 27(6): 1179−1188. DOI: 10.16258/j.cnki.1674-5906.2018.06.025. [44] 赵菲, 谢应忠, 马红彬, 等. 封育对典型草原植物群落物种多样性及土壤有机质的影响[J]. 草业科学, 2011, 28(6): 887−891. ZHAO Fei, XIE Yingzhong, MA Hongbin, et al. Effects of enclosure on species diversity and soil organic matter of typical steppe[J]. Pratacultural Science, 2011, 28(6): 887−891. DOI: 10.3969/j.issn.1001-0629.2011.06.003. [45] LOREAU M, NAEEM S, LNCHAUSTI P. Biodiversity and Ecosystem Functioning: Synthesis and Perspectives[M]. Oxford: Oxford University Press, 2002. DOI:10.1093/oso/9780198515708.001.0001. [46] XIONG Shaojun, NILSSON C. The effects of plant litter on vegetation: a meta-analysis[J]. Journal of Ecology, 1999, 87(6): 984−994. DOI: 10.1046/j.1365-2745.1999.00414.x. [47] BAGCHI S, RITCHIE M E. Herbivore effects on above- and belowground plant production and soil nitrogen availability in the Trans-Himalayan shrub-steppes[J]. Oecologia, 2010, 164(4): 1075−1082. DOI: 10.1007/s00442-010-1690-5. [48] SIEPIELSKI A M, MCPEEK M A. On the evidence for species coexistence: a critique of the coexistence program[J]. Ecology, 2010, 91(11): 3153−3164. DOI: 10.1890/10-0154.1. [49] 贺聪, 马永才, 字洪标. 不同围封年限对高寒草地植物群落的影响[J]. 草业学报, 2025, 34(11): 17−30. HE Cong, MA Yongcai, ZI Hongbiao. Effects of the duration of grazing exclusion on plant communities in alpine grassland[J]. Acta Prataculturae Sinica, 2025, 34(11): 17−30. DOI: 10.11686/cyxb2024523. [50] WU Xinwei, WANG Yichen, SUN Shucun. Long-term fencing decreases plant diversity and soil organic carbon concentration of the Zoige alpine meadows on the eastern Tibetan Plateau[J]. Plant and Soil, 2021, 458(1): 191−200. DOI: 10.1007/s11104-019-04373-7. [51] 杨丽丽, 龚吉蕊, 王忆慧, 等. 内蒙古温带草原不同放牧强度和围栏封育对凋落物分解的影响[J]. 植物生态学报, 2016, 40(8): 748−759. YANG Lili, GONG Jirui, WANG Yihui, et al. Effects of grazing intensity and grazing exclusion on litter decomposition in the temperate steppe of Nei Mongol, China[J]. Chinese Journal of Plant Ecology, 2016, 40(8): 748−759. DOI: 10.17521/cjpe.2016.0051. [52] 陈亮, 王绪高. 生物多样性与森林生态系统健康的几个关键问题[J]. 生态学杂志, 2008, 27(5): 816−820. CHEN Liang, WANG Xugao. Biodiversity and forest ecosystem health: some key questions[J]. Chinese Journal of Ecology, 2008, 27(5): 816−820. DOI: 10.13292/j.1000-4890.2008.0178. [53] CHAI Qinglin, MA Zhanying, CHANG Xiaofeng, et al. Optimizing management to conserve plant diversity and soil carbon stock of semi-arid grasslands on the Loess Plateau[J]. CATENA, 2019, 172: 781−788. DOI: 10.1016/j.catena.2018.09.034. [54] YUAN Z Y, JIAO F, LI Y H, et al. Anthropogenic disturbances are key to maintaining the biodiversity of grasslands[J]. Scientific Reports, 2016, 6: 22132. DOI: 10.1038/srep22132. -

-
链接本文:
https://zlxb.zafu.edu.cn/article/doi/10.11833/j.issn.2095-0756.20260181
 点击查看大图
点击查看大图
计量
- 文章访问数: 195
- HTML全文浏览量: 79
- PDF下载量: 41
- 被引次数: 0
