




-
森林群落是植物在自然条件下分布的最重要的群落类型[1]。其中亚热带常绿阔叶林是全球亚热带大陆东岸湿润气候和季风气候区的典型植被[2],中国是亚热带常绿阔叶林分布面积最大的国家[3]。亚热带常绿阔叶林具有极高的物种多样性、丰富的生物资源和巨大的环境效益[4],对保护环境、维持全球性碳循环平衡和人类可持续发展等具有重要作用[5]。
群落结构是认识群落组成、变化及发展趋势的基础,能够反映群落对环境的适应、群落的动态和机能,并且会影响生态系统的功能特征[6]。群落的垂直结构和径级结构是群落结构中重要的形态和结构特征[7],其中径级结构指林分内各种直径林木按径阶的分配状态。李其斌等[8]认为径级结构是影响植物多样性分布的主要因子;BOULANGEAT等[9]发现乔木物种多样性指数与其周围树种径级存在直接的联系;刘菊莲等[10]认为径级结构不仅揭示了种群在特定时间内个体在不同生长或发育阶段的分布情况,还反映了种群的发展趋势和潜在变化。因此,研究群落径级结构及其更新机制,对阐明种群生态特性、更新对策乃至群落的形成及其稳定性与演替规律等具有重要意义[11]。
物种多样性一直是生态学研究领域的重要内容,森林植被物种多样性与环境因子相关性研究受到了广泛关注。研究发现:人为干扰[12]、林分密度[13]等生物因子,海拔[14]、地形[15]、气候[16]等非生物因子均会对物种多样性产生影响。而土壤环境因子可以通过调节植物营养资源的供给影响森林群落,一定程度上使得土壤环境因子与物种多样性之间的关系更为复杂[17−18]。朱夏夏等[19]在辽东山地老秃顶子发现土壤电导率和pH与Shannon-Wiener指数呈正相关;张荣等[20]在蒙顶山发现全氮与Shannon-Wiener指数呈正相关;南国卫等[21]在黄土高原地区发现有机碳与Shannon-Wiener指数和Pielou均匀度指数呈负相关。因此土壤相关因子对森林群落多样性影响的研究有助于理解物种多样性和土壤之间相互耦合的过程及作用机制。
台州地区位于浙江东南沿海区域,受海洋性季风气候影响显著,南部雁荡山系和北部括苍山系的亚热带常绿阔叶林是该区域的主要森林植被类型。本研究选择2类山系的亚热带常绿阔叶林为研究对象,对比其群落结构和物种组成的差异及其与土壤环境因子的关系,对进一步认识台州沿海地区亚热带常绿阔叶林特征及其变化具有重要意义。
-
雁荡山系(28°12′~28°32′N,121°09′~121°44′E,海拔为42.36~220.55 m)和括苍山系(28°27′~28°41′N,120°47′~121°20′E,海拔为15.63~663.63 m)地处亚热带季风气候区,冬夏季风交替明显,四季分明,温暖湿润,雨量充沛,光照充足,无霜期长。年均气温为17.3 ℃,最冷月(1月)平均气温为6.7 ℃,最热月(7月)平均气温为28.1 ℃。全年有2个明显的降水期,分别为3—6月和8—9月,年均降水量为
1649.6 mm。地带性植被为亚热带常绿阔叶林。 -
2023年9—11月在雁荡山系和括苍山系的典型亚热带常绿阔叶林分别设置11和15个20 m×20 m样方(表1),将每个样方都划分为16个5 m×5 m的小样方。依据文献[22−23]的研究方法,标定样方中所有胸径(DBH)≥1 cm的乔木个体,并划分为3个等级[24−25],1 cm≤DBH<5 cm为小径级,5 cm≤DBH<20 cm为中等径级,DBH≥20 cm为较大径级。记录每株乔木名称、胸径和高度,同时记录每个样地的土壤生态因子(岩石裸露率、土壤深度、凋落物盖度、凋落物厚度、腐殖质层厚度和土壤电导率)。在每个20 m×20 m样方随机选取5个监测点,用环刀取土,根据LY/T 1215—1999《森林土壤水分-物理性质的测定》方法测定土壤养分相关指标(pH、有机碳、全碳、全氮、全磷、硝态氮、铵态氮)。
表 1 雁荡山系和括苍山系森林群落样地概况
Table 1. General situation of plots in forest communities of Yandang and Kuocang Mountains
样地 森林群落 雁荡山系
1~111. 马尾松Pinus massoniana+檵木Loropetalum chinense;2. 樟树Cinnamomum camphora+枫香Liquidambar formosana;3. 窄基红褐柃Eurya rubiginosa;4. 樟树;5. 樟树;6. 木荷Schima superba;7. 枫香;8. 檵木+隔药柃Eurya muricata ;9. 木荷;10. 青冈栎Quercus glauca+枫香;11. 青冈栎 括苍山系
12~2612. 樟树;13. 檵木;14. 木荷;15. 木荷;16. 枫香;17. 木荷+苦槠 Castanopsis sclerophylla;18. 隔药柃;19. 乌桕Triadica sebifera;20. 榔榆Ulmus parvifolia ;21. 苦槠+枫香;22. 杉木Cunninghamia lanceolata+枫香;23. 杉木;24. 木荷;25. 木荷;26. 木荷+薄叶山矾 Symplocos anomala -
重要值是反映各物种在森林植被中作用的指标,以数量、胸径和高度等3个指标分析。分别统计并比较雁荡山系和括苍山系森林群落各物种重要值的变化。采用Simpson指数、Shannon-Wiener指数、Pielou均匀度指数和Margalef丰富度指数反映2类山系森林群落物种多样性水平。物种重要值和群落多样性指数采用任学敏等[26]和FEEST等[27]的方法进行计算。
采用SPSS 27.0进行独立样本t检验,分析雁荡山系和括苍山系森林群落多样性指数差异。采用R 4.3进行冗余分析(RDA)检验环境因子与物种多样性的相关性。采用SigmaPlot 14.0作图。
-
雁荡山系森林群落共记录乔木67种,隶属31科46属。科分析结果显示:山矾科Symplocaceae的物种最多,有7属7种,蔷薇科Rosaceae和壳斗科Fagaceae次之,分别有3属6种和4属6种。属分析结果显示:山矾属Symplocos的物种最多,有7种,其次为石楠属Photinia,有4种。括苍山系森林群落共记录乔木89种,隶属38科62属。科分析结果显示:杜鹃花科Ericaceae的物种最多,有5属9种,樟科Lauraceae次之,有5属7种。属分析结果显示:山矾属Symplocos的物种最多,有5种,其次为冬青属Ilex、榕属Ficus、山茶属Camellia和越橘属Vaccinium,各有4种。2类山系森林群落中重要值≥1%的共有物种为木荷Schima superba 、枫香Liquidambar formosana 、樟树Cinnamomum camphora、杉木Cunninghamia lanceolata、檵木Loropetalum chinense 、窄基红褐柃Eurya rubiginosa 、青冈栎Quercus glauca、苦槠Castanopsis sclerophylla、水团花Adina pilulifera 、隔药柃Eurya muricata和白花龙Styrax faberi (表2)。括苍山系森林群落中木荷重要值为27.37%,比雁荡山系高16.80%,苦槠和隔药柃的重要值在括苍山系森林群落也明显高于雁荡山系。但括苍山系森林群落中枫香、樟树、檵木和青冈栎的重要值均低于雁荡山系,杉木、窄基红褐柃、水团花和白花龙的重要值在2类山系森林群落中变化不明显。
表 2 雁荡山系和括苍山系森林群落重要值≥1%的优势物种
Table 2. Dominant species with importance value ≥1% in forest communities of Yandang and Kuocang Mountains
样地 物种 个体数/株 平均高度/m 胸高断面积/(m2·hm−2) 重要值/% 雁荡山系1~11 枫香Liquidambar formosana* 80 9.22 2.0656 13.11 木荷 Schima superba* 90 8.18 1.2252 10.53 樟树Cinnamomum camphora* 25 10.95 1.9007 8.95 杉木Cunninghamia lanceolata* 107 5.18 0.6754 8.10 马尾松 Pinus massoniana 62 6.13 1.0626 7.05 檵木 Loropetalum chinense* 134 3.51 0.1017 6.40 窄基红褐柃Eurya rubiginosa* 97 3.87 0.1205 4.99 青冈栎Quercus glauca* 26 10.68 0.7822 4.84 苦槠 Castanopsis sclerophylla* 26 7.53 0.2777 2.64 水团花 Adina pilulifera* 50 3.46 0.0601 2.46 栲树Castanopsis fargesii 31 5.91 0.1308 2.24 羊舌树Symplocos glauca 41 3.41 0.0448 1.99 赤楠Syzygium buxifolium 45 2.79 0.0177 1.93 隔药柃Eurya muricata* 44 2.56 0.0163 1.83 毛八角枫 Alangium kurzii 17 5.9 0.1807 1.60 白花龙 Styrax faberi* 26 4.08 0.0335 1.37 野桐 Mallotus tenuifolius 27 3.50 0.0259 1.31 山茶 Camellia japonica 17 5.57 0.0747 1.21 中华杜英 Elaeocarpus chinensis 16 5.92 0.0594 1.13 括苍山系12~26 木荷 Schima superba* 475 10.12 5.3722 27.37 薄叶山矾 Symplocos anomala 320 4.71 0.2163 7.43 杉木Cunninghamia lanceolata* 104 7.93 1.4826 6.18 枫香 Liquidambar formosana* 68 9.88 1.6550 5.82 窄基红褐柃Eurya rubiginosa* 253 3.15 0.0674 4.79 苦槠 Castanopsis sclerophylla* 106 6.68 0.8548 4.61 隔药柃Eurya muricata* 116 3.88 0.4328 3.28 马银花Rhododendron ovatum 140 4.48 0.0695 3.06 樟树Cinnamomum camphora* 22 11.45 1.0489 2.86 青冈栎Cyclobalanopsis glauca* 62 8.38 0.4343 2.77 水团花 Adina pilulifera* 108 3.86 0.1640 2.53 树参 Dendropanax dentiger 67 6.62 0.2697 2.31 檵木Loropetalum chinense* 89 4.89 0.1484 2.23 麂角杜鹃 Rhododendron latoucheae 92 4.33 0.0979 2.15 鼠刺 Itea chinensis 88 4.23 0.1171 2.07 白花龙 Styrax faberi* 61 4.00 0.0564 1.37 红楠Machilus thunbergii 40 4.21 0.1512 1.16 檫木 Sassafras tzumu 17 8.54 0.2380 1.02 说明:*表示该物种为2类山系森林群落共有物种。 2类山系森林群落多样性(表3)分析显示:雁荡山系和括苍山系森林群落的Simpson指数和Margalef丰富度指数差异不显著,而雁荡山系森林群落Shannon-Wiener指数和Pielou均匀度指数显著高于括苍山系森林群落(P<0.05)。
表 3 雁荡山系和括苍山系森林群落多样性
Table 3. Diversity of forest communities in Yandang and Kuocang Mountains
研究样地 Simpson指数 Shannon-Wiener指数 Pielou均匀度指数 Margalef丰富度指数 雁荡山系森林群落 0.80±0.15 2.19±0.55 0.48±0.11 3.84±1.18 括苍山系森林群落 0.81±0.10 1.60±0.81 0.34±0.17 3.64±1.05 t −0.28 2.24 2.54 0.44 P 0.78 0.04* 0.02* 0.66 说明:数据为平均值±标准差;*表示差异显著(P<0.05)。 -
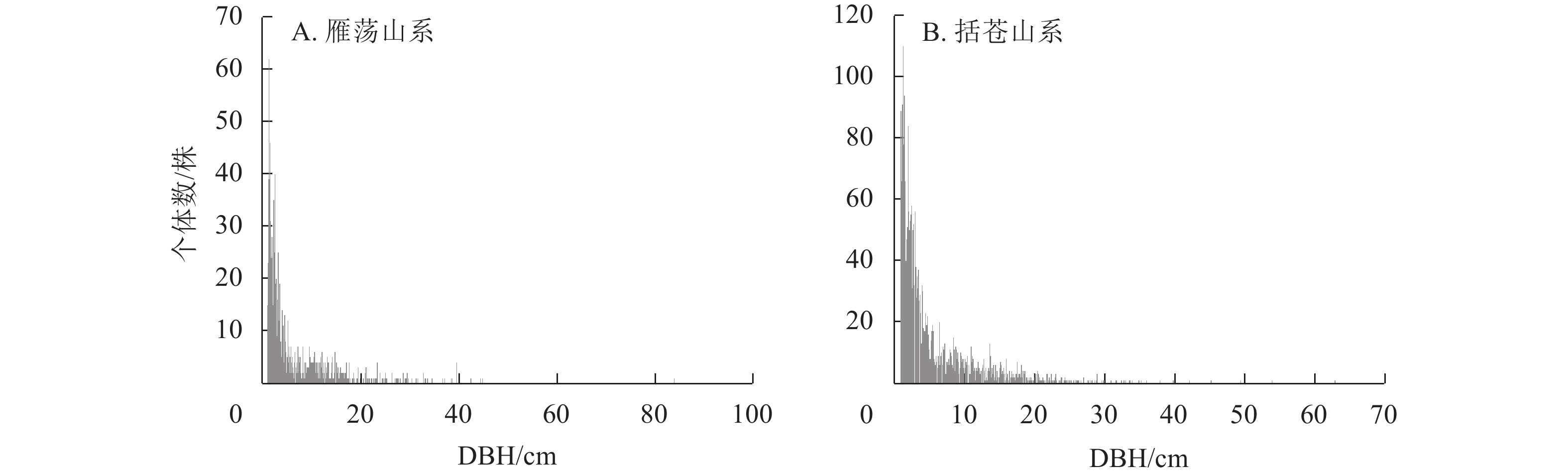
径级结构(图1)分析显示:2类山系森林群落乔木(DBH≥1 cm)总个体径级结构呈倒“J”型分布。在雁荡山系森林群落中,小径级个体有665株,占总个体数的53.7%;中等径级个体有396株,占总个体数的32.0%;较大径级个体仅有76株,占总个体数的6.1%。在括苍山系森林群落中,小径级个体有
1786 株,占总个体数的65.5%;中等径级个体有833株,占总个体数的30.5%;较大径级个体仅有109株,占总个体数的4.0%。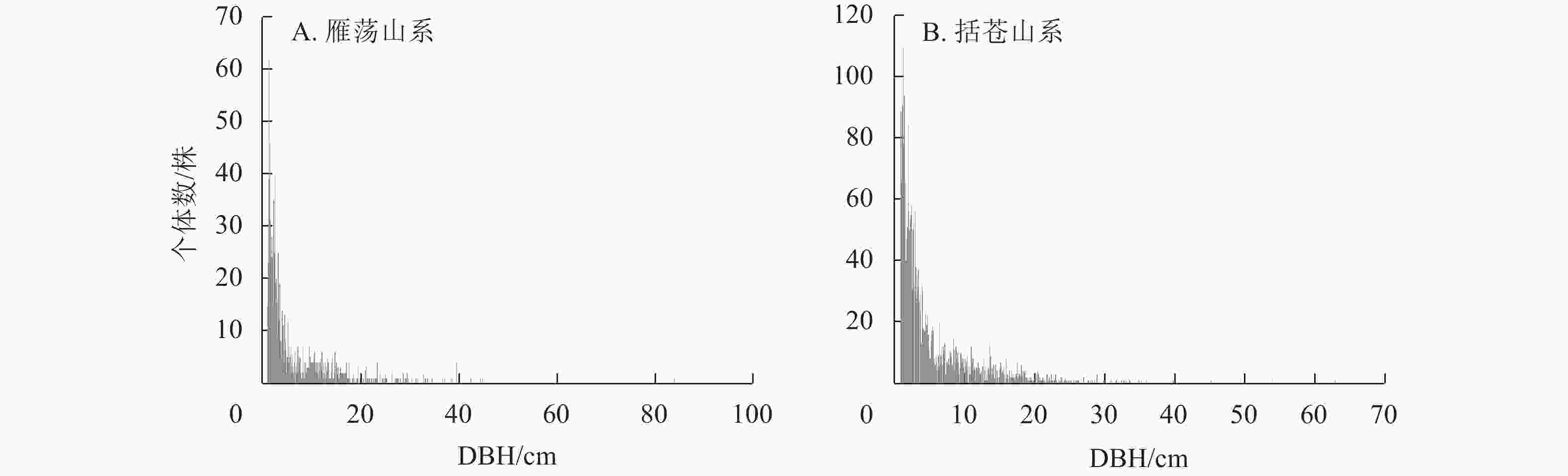
图 1 雁荡山系和括苍山系森林群落胸径≥1 cm乔木径级分布
Figure 1. Diameter distribution of trees (DBH≥1cm) of forest communities in Yandang and Kuocang Mountains
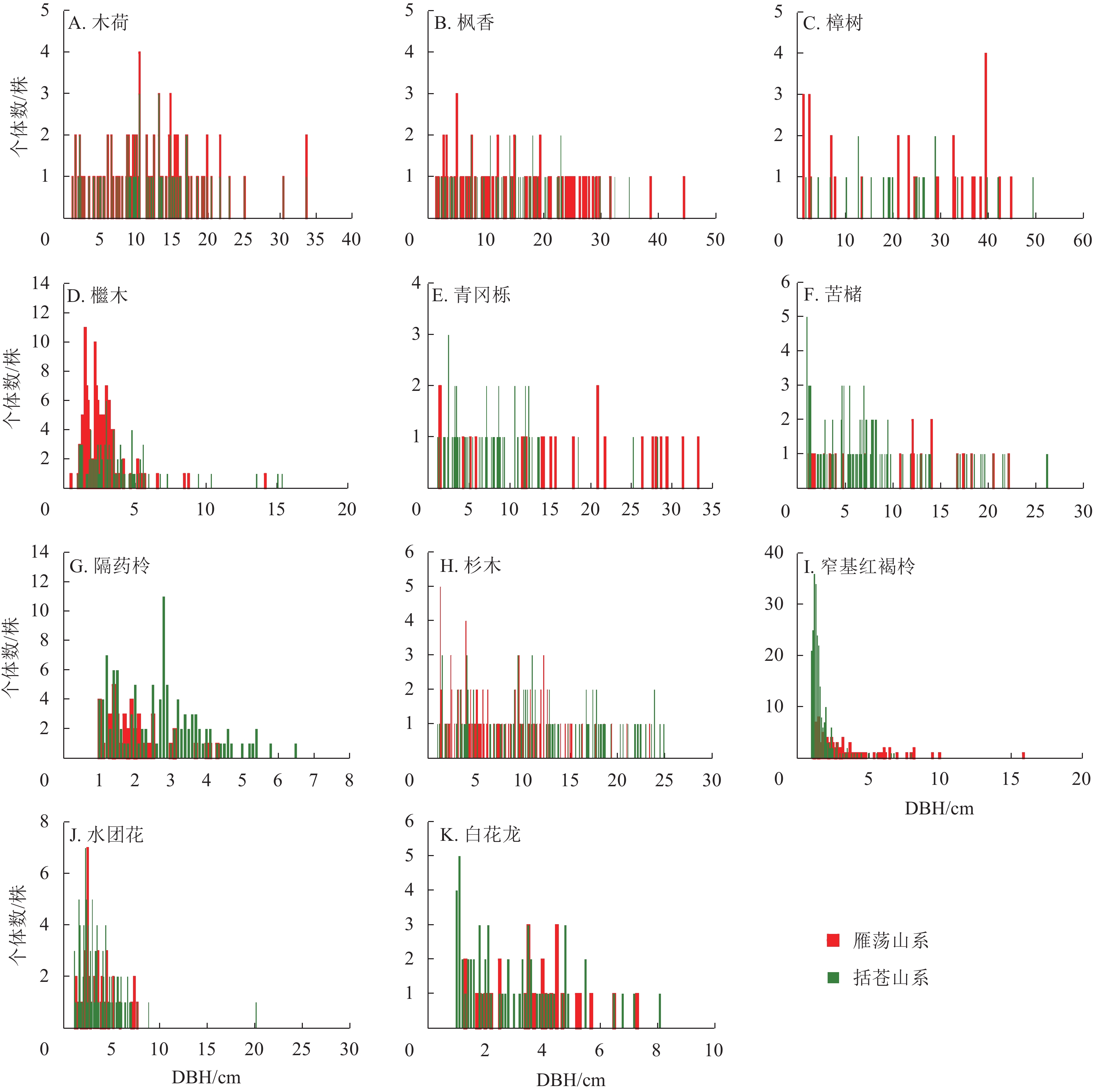
2类山系森林群落重要值≥1%的共有物种为11种(图2)。樟树、檵木和青冈栎在2类森林群落的径级结构差异明显,雁荡山系森林群落的樟树和檵木各径级个体数明显高于括苍山系,括苍山系森林群落的青冈栎个体数大部分属于小径级范围;木荷和隔药柃径级结构呈偏正态分布,木荷个体集中分布在1 cm≤DBH≤10 cm内,2类山系森林群落中隔药柃大径级较少,而且雁荡山系个体数集中在DBH<4 cm内。枫香的径级结构呈平均分布,苦槠、杉木、窄基红褐柃、水团花和白花龙的径级结构呈倒“J”型。

图 2 雁荡山系和括苍山系森林群落共有物种径级分布
Figure 2. Diameter distribution of shared species (DBH≥1 cm) of forest communities in Yandang and Kuocang Mountains
-
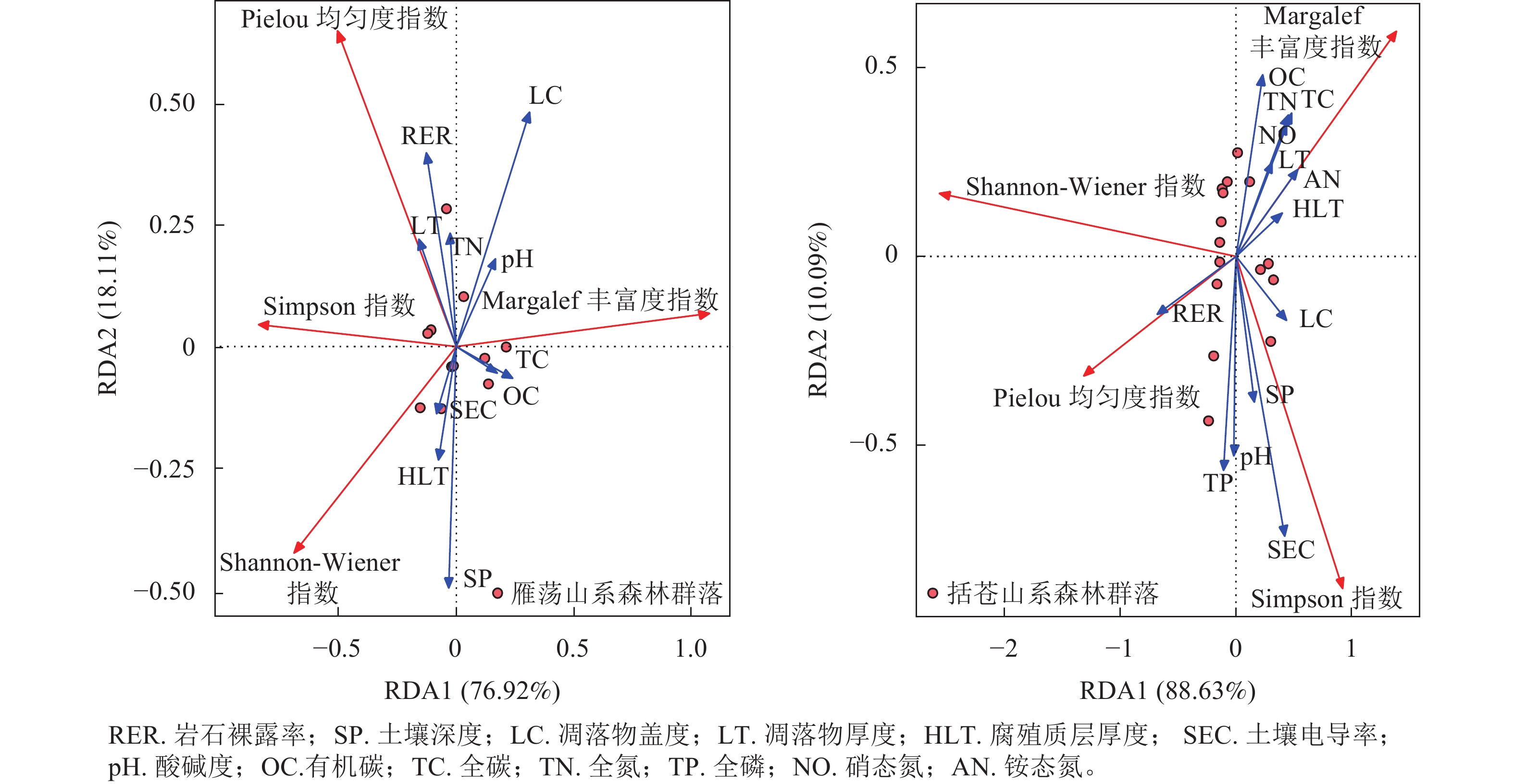
雁荡山系和括苍山系森林群落植物多样性与环境因子的冗余分析(图3)结果显示:雁荡山系森林群落排序轴Ⅰ解释了总变量的76.92%,与有机碳和全碳相关;排序轴Ⅱ解释了总变量的18.11%,与土壤深度、凋落物厚度、土壤电导率、全氮和岩石裸露率相关。括苍山系森林群落排序轴Ⅰ解释了总变量的88.63%,与岩石裸露率和腐殖质层厚度相关;排序轴Ⅱ解释了总变量的10.09%,与土壤全磷、pH、土壤深度、土壤电导率和有机碳相关。冗余分析显示:2类山系森林群落Shannon-Wiener指数均与全氮呈负相关。岩石裸露率与雁荡山系森林群落Shannon-Wiener指数呈负相关,与括苍山系森林群落Shannon-Wiener指数呈正相关。土壤电导率与雁荡山系森林群落Shannon-Wiener指数呈正相关,与括苍山系森林群落Shannon-Wiener指数呈负相关。2类山系森林群落Pielou 均匀度指数均与岩石裸露率和pH呈正相关,均与全碳呈负相关。土壤电导率与雁荡山系森林群落Pielou 均匀度指数呈负相关,与括苍山系森林群落Pielou 均匀度指数呈正相关。全氮与雁荡山系森林群落Pielou 均匀度指数呈正相关,与括苍山系森林群落Pielou 均匀度指数呈负相关。2类山系森林群落Simpson指数与土壤电导率和岩石裸露率呈正相关,而与全碳呈负相关。2类山系森林群落Margalef丰富度指数均与全氮和全碳呈正相关,与土壤电导率呈负相关。
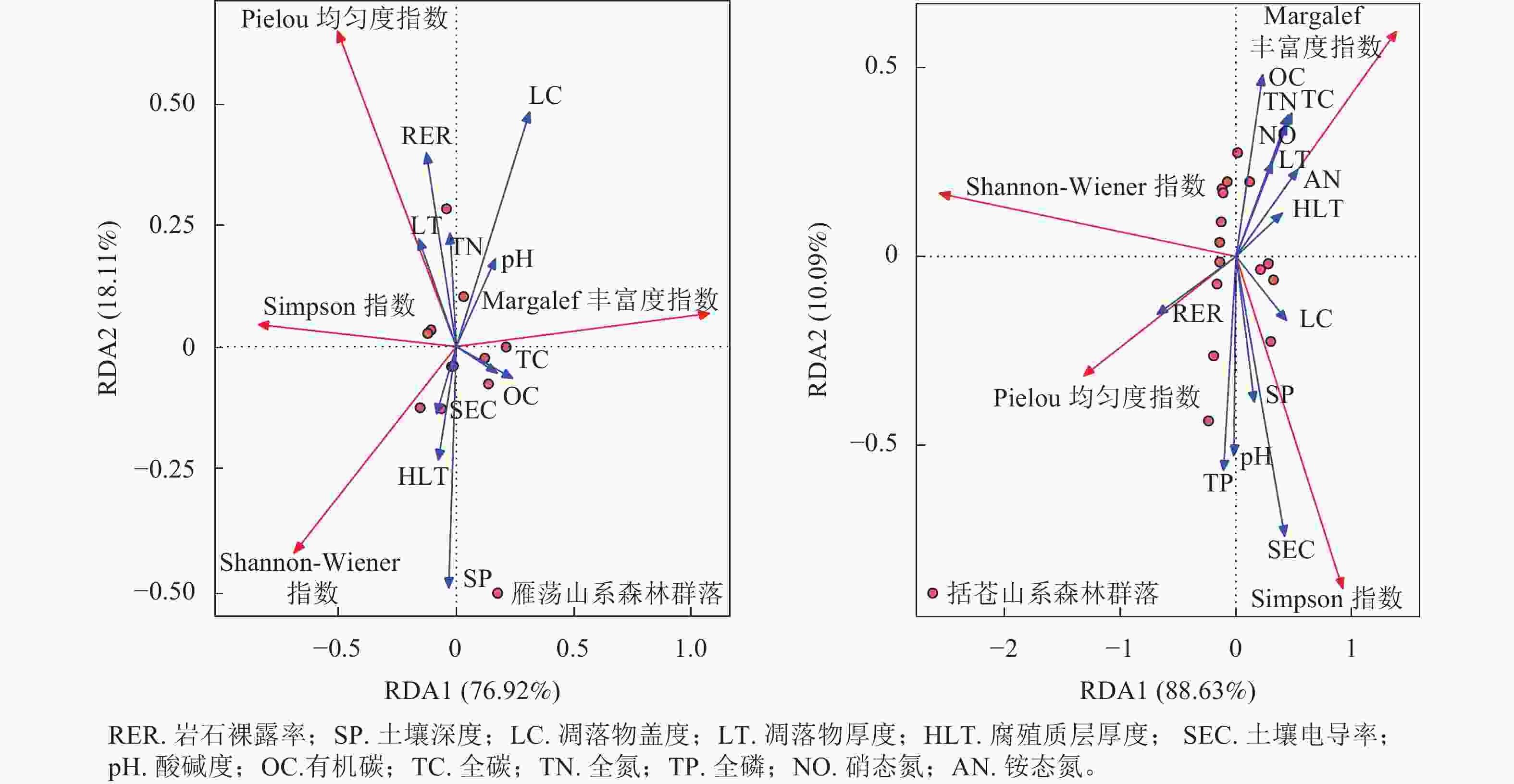
图 3 雁荡山系和括苍山系森林群落植物多样性与环境因子的RDA排序
Figure 3. RDA ranking of plant community diversity and environmental factors of forest communities in Yandang and Kuocang Mountains
-
物种组成是决定群落性质的重要因素,也是了解生态系统维持机制的重要途径[28−29] 。通过研究雁荡山系和括苍山系森林群落的物种组成,发现括苍山系森林群落物种数多于雁荡山系。2类山系森林群落均以山矾科和杜鹃花科物种为主,这与以壳斗科物种为主的古田山常绿阔叶林[30]和以杜鹃花科物种为主的九龙山常绿阔叶林[11]具有一定相似性。2类山系森林群落重要值≥1%的共有物种有11种,如木荷、枫香、樟树等,这些物种在紫金山[31]、午潮山[1]等亚热带常绿阔叶林群落同样为优势物种,表明该地区亚热带常绿阔叶林中木荷、枫香、樟树等为普遍存在的优势种。括苍山系森林群落木荷的重要值高于雁荡山系,木荷是2类山系森林群落重要的优势物种,表明木荷在括苍山系森林群落构建中起到更重要的作用。这可能是2类山系森林群落在海拔上的差异,导致了水热条件的变化,进而造成了凋落物厚度、有机碳和全氮的差异。括苍山系森林群落的凋落物厚度较高,土壤养分含量高,这可能是木荷更具优势的原因。枫香和樟树在2类森林群落重要值较高,表明两者对2类山系森林群落生境都具有较高的适合度,但是雁荡山系森林群落枫香重要值高于括苍山系,樟树则反之,表明枫香更适合低海拔地区,而樟树更适合高海拔地区。雁荡山系森林群落Shannon-Wiener指数与Pielou均匀度指数均高于括苍山系。雁荡山系森林群落物种间数量分布相对均匀,优势物种优势度低,而括苍山系森林群落木荷生长茂盛,郁闭度大,林地凋落物较多,竞争胁迫导致其他物种数较少,因此括苍山系森林群落Shannon-Wiener指数与Pielou均匀度指数较低。表明括苍山系森林群落木荷优势明显,导致其多样性明显低于雁荡山系。
-
雁荡山系和括苍山系森林群落总个体径级结构均呈倒“J”型分布,江西大岗山低海拔常绿阔叶林[32]、鼎湖山南亚热带常绿阔叶林[33]等的研究也有类似的结果。群落个体集中在小径级(1 cm≤DBH≤5 cm),表明幼树储备丰富,自然更新良好,整体呈现稳定生长状态。雁荡山系森林群落的樟树和檵木各径级的个体数均高于括苍山系,表明樟树和檵木更适合生长在低海拔的雁荡山系森林群落。与雁荡山系相比,青冈栎在括苍山系森林群落小径级个体较多,可能由于括苍山系森林群落郁闭度较高,林下光照弱,而青冈栎为阳性树种,幼苗更新生长受到抑制,大径级的植株较少。2类山系森林群落多数物种个体径级结构呈倒“J”型分布,表明这些物种在2类山系森林群落均有良好的更新能力,这与天童山常绿阔叶林[34]的云山青冈Cyclobalanopsis nubium、栲树等优势物种的径级结构相似,表明主要优势物种更新能力强是亚热带常绿阔叶林的特点。木荷的径级分布呈偏正态分布,以中小径级个体为主,充足的幼苗是其种群稳定生长,处于优势地位的原因。由于亚热带常绿阔叶林区域水热光照条件好,木荷等林冠层高大的乔木可以合理利用光照水分,中龄期的个体处于快速生长期,竞争力强,因此个体数较多。而木荷等主要乔木均是阳性树种,冠幅较大造成郁闭度增大,林冠层对光资源截留导致林下的幼苗不易接受充足的阳光和水分,且还要与林下耐阴草本等共同竞争水分、土壤及营养物质,从而影响存活率,致使幼龄期个体数少,使林冠层径级呈偏正态分布[35]。
-
环境因子对植物群落的物种组成、结构和多样性等具有重要影响[36−37]。冗余分析结果显示:全氮增加对2类山系森林群落Shannon-Wiener指数具有抑制作用。SUDING等[38]发现:在氮限制的地区增加氮素会缓解种间竞争,使群落的物种多样性增加。而在氮素水平高的区域,氮素增加可能会加剧种间竞争,导致物种多样性的减少[39]。全氮增加降低了括苍山森林群落的Pielou 均匀度指数,但是增加了雁荡山系森林群落的Pielou均匀度指数,这可能与雁荡山系的植株密度较少有关,整体种间竞争水平较低,全氮的增加促进了多数物种植株的生长,从而提高了Pielou均匀度指数。岩石裸露率与括苍山系森林群落Shannon-Wiener指数和Pielou均匀度指数均呈正相关,这可能与光照有关,括苍山系的裸露岩石区域并没有高大乔木生长,容易形成林窗,导致岩石裸露区域物种和植株数均有显著增加。但是在雁荡山系森林群落,岩石裸露率则与Shannon-Wiener指数呈负相关。雁荡山系森林群落植株密度较少,岩石裸露率高的群落植株密度可能更为稀少,导致多样性指数下降。土壤pH决定了植株生存的适宜度[40−41]。土壤pH与2类山系森林群落Pielou 均匀度指数呈正相关。2类森林群落的土壤pH均值为5.6,土壤性质偏酸性,过低的pH会影响土壤微生物活动,进而影响植物多样性, pH的增加对物种生长起到促进作用,进而增加物种多样性。李其斌等[8]发现长白山土壤 pH与 Pielou 均匀度指数显著负相关,这与该区域土壤pH较高有关,pH增加反而抑制了物种多样性。土壤全碳含量越高,Pielou 均匀度指数越低,这与云顶山有机质与Pielou 均匀度指数呈负相关的结果一致[42]。土壤电导率对2类山系森林群落的Pielou 均匀度指数和Shannon-Wiener指数也存在影响。土壤电导率越低表明土壤中可溶性盐离子含量越低,导致植物吸收的养分越少,进而抑制种子萌发和幼苗生长。
-
综上所述,雁荡山系和括苍山系森林群落具有相似优势物种,括苍山系森林群落优势物种优势度明显高于雁荡山系。雁荡山系森林群落的Shannon-Wiener指数与Pielou均匀度指显著高于括苍山系。2类山系森林群落总个体径级结构呈倒“J”型分布。全氮、岩石裸露率、pH、土壤全碳和土壤电导率是影响2类森林群落物种多样性的主要土壤因子。2类山系森林群落的主要差异是括苍山系森林群落的木荷重要值明显高于雁荡山系。
Analysis of the structure and species composition of the forest communities in Yandang and Kuocang Mountains in Taizhou, Zhejiang Province, China
-
摘要:
目的 以台州地区雁荡山系和括苍山系森林群落为研究对象,研究2类山系森林群落结构和物种组成差异及其成因,为了解台州地区常绿阔叶林变化提供技术支撑。 方法 基于雁荡山系11个和括苍山系15个20 m×20 m森林群落样地调查结果,结合冗余分析(RDA)等方法,分析样地物种组成、径级结构及其与环境因子的关系。 结果 雁荡山系森林群落共记录乔木67种,隶属31科46属,括苍山系森林群落共记录乔木89种,隶属38科62属。2类山系森林群落重要值≥1%的共有物种有木荷Schima superba、枫香Liquidambar formosana、樟树Cinnamomum camphora、杉木Cunninghamia lanceolata、檵木Loropetalum chinense、窄基红褐柃Eurya rubiginosa、青冈栎Quercus glauca、苦槠Castanopsis sclerophylla、水团花Adina pilulifera、隔药柃Eurya muricata和白花龙Styrax faberi。括苍山系森林群落中木荷重要值为27.37%,比雁荡山系高16.80%。雁荡山系森林群落Shannon-Wiener指数、Pielou均匀度指数均显著高于括苍山系(P<0.05)。2类山系森林群落物种总个体径级结构呈倒“J”型。全氮、岩石裸露率、pH、土壤全碳和土壤电导率是影响2类山系群落多样性指数的主要因素。 结论 2类山系森林群落具有相似优势物种,雁荡山系森林群落的Shannon-Wiener指数与Pielou均匀度指数显著高于括苍山系。2类山系森林群落的主要差异是括苍山系森林群落的木荷重要值明显高于雁荡山系。图3表3参42 Abstract:Objective Taking the forest communities of Yandang and Kuocang Mountains in Taizhou as the research objects, this study focuses on the differences of forest community structure and species composition of 2 mountain systems and their causes, which provides technical support for understanding the changes of evergreen broad-leaved forests in Taizhou. Method Based on the survey of 11 plots in Yandang Mountain and 15 plots in Kuocang Mountain (20 m×20 m), species composition, diameter structure and their relationships with the environmental factors were analyzed using methods such as Redundancy Analysis (RDA). Result Forest communities of Yandang Mountain recorded 67 species of trees, belonging to 31 families and 46 genera while those of Kuocang Mountain recorded 89 species of trees, belonging to 38 families and 62 genera. Species having important values ≥1% in the forest communities of Yandang and Kuocang Mountains were Schima superba, Liquidambar formosana, Cinnamomum camphora, Cunninghamia lanceolata, Loropetalum chinense, Eurya rubiginosa, Cyclobalanopsis glauca, Castanopsis sclerophylla, Adina pilulifera, Eurya muricata, Styrax faberi. The important value of Schima superba in the forest communities of Kuocang Mountain was 27.37%, which was 16.80% higher than that in Yandang Mountain. Shannon-Wiener index and Pielou evenness index of the forest communities of Yandang Mountain were significantly higher than those of Kuocang Mountain (P<0.05). The total individual diameter distribution of species in 2 forest communities showed inverted “J” shapes. Total nitrogen, rock exposure rate, pH, soil total carbon and electrical conductivity were the main factors affecting the diversity indexes of forest communities of 2 mountains. Conclusion Shannon-Wiener index and Pielou evenness index of Yandang Mountain forest community were significantly higher than those of Kuocang Mountain forest communities. The main difference between 2 mountain forest communities is that the important value of Schima superba in Kuocang Mountain forest community is significantly higher than that in Yandang Mountain. [Ch, 3 fig. 3 tab. 42 ref.] -
图 1 雁荡山系和括苍山系森林群落胸径≥1 cm乔木径级分布
Figure 1 Diameter distribution of trees (DBH≥1cm) of forest communities in Yandang and Kuocang Mountains
图 2 雁荡山系和括苍山系森林群落共有物种径级分布
Figure 2 Diameter distribution of shared species (DBH≥1 cm) of forest communities in Yandang and Kuocang Mountains
图 3 雁荡山系和括苍山系森林群落植物多样性与环境因子的RDA排序
Figure 3 RDA ranking of plant community diversity and environmental factors of forest communities in Yandang and Kuocang Mountains
表 1 雁荡山系和括苍山系森林群落样地概况
Table 1. General situation of plots in forest communities of Yandang and Kuocang Mountains
样地 森林群落 雁荡山系
1~111. 马尾松Pinus massoniana+檵木Loropetalum chinense;2. 樟树Cinnamomum camphora+枫香Liquidambar formosana;3. 窄基红褐柃Eurya rubiginosa;4. 樟树;5. 樟树;6. 木荷Schima superba;7. 枫香;8. 檵木+隔药柃Eurya muricata ;9. 木荷;10. 青冈栎Quercus glauca+枫香;11. 青冈栎 括苍山系
12~2612. 樟树;13. 檵木;14. 木荷;15. 木荷;16. 枫香;17. 木荷+苦槠 Castanopsis sclerophylla;18. 隔药柃;19. 乌桕Triadica sebifera;20. 榔榆Ulmus parvifolia ;21. 苦槠+枫香;22. 杉木Cunninghamia lanceolata+枫香;23. 杉木;24. 木荷;25. 木荷;26. 木荷+薄叶山矾 Symplocos anomala  下载: 导出CSV
下载: 导出CSV
表 2 雁荡山系和括苍山系森林群落重要值≥1%的优势物种
Table 2. Dominant species with importance value ≥1% in forest communities of Yandang and Kuocang Mountains
样地 物种 个体数/株 平均高度/m 胸高断面积/(m2·hm−2) 重要值/% 雁荡山系1~11 枫香Liquidambar formosana* 80 9.22 2.0656 13.11 木荷 Schima superba* 90 8.18 1.2252 10.53 樟树Cinnamomum camphora* 25 10.95 1.9007 8.95 杉木Cunninghamia lanceolata* 107 5.18 0.6754 8.10 马尾松 Pinus massoniana 62 6.13 1.0626 7.05 檵木 Loropetalum chinense* 134 3.51 0.1017 6.40 窄基红褐柃Eurya rubiginosa* 97 3.87 0.1205 4.99 青冈栎Quercus glauca* 26 10.68 0.7822 4.84 苦槠 Castanopsis sclerophylla* 26 7.53 0.2777 2.64 水团花 Adina pilulifera* 50 3.46 0.0601 2.46 栲树Castanopsis fargesii 31 5.91 0.1308 2.24 羊舌树Symplocos glauca 41 3.41 0.0448 1.99 赤楠Syzygium buxifolium 45 2.79 0.0177 1.93 隔药柃Eurya muricata* 44 2.56 0.0163 1.83 毛八角枫 Alangium kurzii 17 5.9 0.1807 1.60 白花龙 Styrax faberi* 26 4.08 0.0335 1.37 野桐 Mallotus tenuifolius 27 3.50 0.0259 1.31 山茶 Camellia japonica 17 5.57 0.0747 1.21 中华杜英 Elaeocarpus chinensis 16 5.92 0.0594 1.13 括苍山系12~26 木荷 Schima superba* 475 10.12 5.3722 27.37 薄叶山矾 Symplocos anomala 320 4.71 0.2163 7.43 杉木Cunninghamia lanceolata* 104 7.93 1.4826 6.18 枫香 Liquidambar formosana* 68 9.88 1.6550 5.82 窄基红褐柃Eurya rubiginosa* 253 3.15 0.0674 4.79 苦槠 Castanopsis sclerophylla* 106 6.68 0.8548 4.61 隔药柃Eurya muricata* 116 3.88 0.4328 3.28 马银花Rhododendron ovatum 140 4.48 0.0695 3.06 樟树Cinnamomum camphora* 22 11.45 1.0489 2.86 青冈栎Cyclobalanopsis glauca* 62 8.38 0.4343 2.77 水团花 Adina pilulifera* 108 3.86 0.1640 2.53 树参 Dendropanax dentiger 67 6.62 0.2697 2.31 檵木Loropetalum chinense* 89 4.89 0.1484 2.23 麂角杜鹃 Rhododendron latoucheae 92 4.33 0.0979 2.15 鼠刺 Itea chinensis 88 4.23 0.1171 2.07 白花龙 Styrax faberi* 61 4.00 0.0564 1.37 红楠Machilus thunbergii 40 4.21 0.1512 1.16 檫木 Sassafras tzumu 17 8.54 0.2380 1.02 说明:*表示该物种为2类山系森林群落共有物种。
下载: 导出CSV
表 3 雁荡山系和括苍山系森林群落多样性
Table 3. Diversity of forest communities in Yandang and Kuocang Mountains
研究样地 Simpson指数 Shannon-Wiener指数 Pielou均匀度指数 Margalef丰富度指数 雁荡山系森林群落 0.80±0.15 2.19±0.55 0.48±0.11 3.84±1.18 括苍山系森林群落 0.81±0.10 1.60±0.81 0.34±0.17 3.64±1.05 t −0.28 2.24 2.54 0.44 P 0.78 0.04* 0.02* 0.66 说明:数据为平均值±标准差;*表示差异显著(P<0.05)。
下载: 导出CSV
-
[1] 田立新, 吴初平, 杨少宗, 等. 浙江杭州午潮山亚热带常绿阔叶林群落结构和物种组成[J]. 应用生态学报, 2020, 31(6): 1909−1915. TIAN Lixin, WU Chuping, YANG Shaozong, et al. Community structure and species composition in a subtropical evergreen broad-leaved forest in Wuchaoshan, Hangzhou, Zhejiang Province, China [J]. Chinese Journal of Applied Ecology, 2020, 31(6): 1909−1915. [2] 吴征镒. 中国植被[M]. 北京: 科学出版社, 1980. WU Zhengyi. Chinese Vegetation [M]. Beijing: Science Press, 1980. [3] 何永涛, 曹敏, 唐勇, 等. 滇中地区常绿阔叶林树种多样性比较研究[J]. 山地学报, 2000, 18(4): 322−328. HE Yongtao, CAO Min, TANG Yong, et al. A comparative study on tree species diversity of evergreen broadleaved forest, central Yunnan [J]. Journal of Mountain Science, 2000, 18(4): 322−328. [4] 丁圣彦, 宋永昌. 常绿阔叶林植被动态研究进展[J]. 生态学报, 2004, 24(8): 1765−1775. DING Shengyan, SONG Yongchang. Research advances in vegetation dynamic of evergreen broad-leaved forest [J]. Acta Ecologica Sinica, 2004, 24(8): 1765−1775. [5] 包维楷, 刘照光, 刘朝禄. 中亚热带湿性常绿阔叶次生林自然恢复15年来群落乔木层的动态变化[J]. 植物生态学报, 2000, 24(6): 702−709. BAO Weikai, LIU Zhaoguang, LIU Zhaolu. Fifteen-year changes of tree layer in secondary Castanopsis-Schima humid evergreen broad-leaved forest in central subtropics of western China [J]. Acta Phytoecologica Sinica, 2000, 24(6): 702−709. [6] 姚兰, 艾训儒, 易咏梅, 等. 鄂西南亚热带常绿落叶阔叶混交林优势种群的结构与动态特征[J]. 林业科学, 2017, 53(2): 10−18. YAO Lan, AI Xunru, YI Yongmei, et al. Structure and dynamics of dominant populations in the mixed forest of subtropical evergreen and deciduous broad-leaved tree species in the southwest of Hubei Province [J]. Scientia Silvae Sinicae, 2017, 53(2): 10−18. [7] 盘远方, 梁志慧, 李嘉宝, 等. 桂林岩溶石山常绿落叶阔叶混交林群落结构与物种多样性[J]. 生态学报, 2021, 41(6): 2451−2459. PAN Yuanfang, LIANG Zhihui, LI Jiabao, et al. Community structure and species diversity of evergreen deciduous broad-leaved mixed forest in Karst hills of Guilin [J]. Acta Ecologica Sinica, 2021, 41(6): 2451−2459. [8] 李其斌, 张春雨, 赵秀海. 长白山不同演替阶段针阔混交林群落物种多样性及其影响因子[J]. 生态学报, 2022, 42(17): 7147−7155. LI Qibin, ZHANG Chunyu, ZHAO Xiuhai. Species diversity and influencing factors of coniferous and broad-leaved mixed forest communities in different successional stages of Changbai Mountain [J]. Acta Ecologica Sinica, 2022, 42(17): 7147−7155. [9] BOULANGEAT I, GRAVEL D, THUILLER W. Accounting for dispersal and biotic interactions to disentangle the drivers of species distributions and their abundances [J]. Ecology Letters, 2012, 15(6): 584−593. [10] 刘菊莲, 韦博良, 吴雁南, 等. 浙江九龙山常绿阔叶林不同物种的径级结构及空间关联[J]. 浙江农林大学学报, 2023, 40(3): 598−607. LIU Julian, WEI Boliang, WU Yannan, et al. Size structure and spatial association of different species of an evergreen broad-leaved forest in Jiulong Mountain, Zhejiang Province [J]. Journal of Zhejiang A&F University, 2023, 40(3): 598−607. [11] 张志祥, 刘鹏, 刘春生, 等. 浙江九龙山南方铁杉(Tsuga tchekiangensis)群落结构及优势种群更新类型[J]. 生态学报, 2008, 28(9): 4547−4558. ZHANG Zhixiang, LIU Peng, LIU Chunsheng, et al. The structure characteristics and dominant population regeneration types of Tsuga tchekiangensis communities in the Jiulongshan National Natural Reserve of Zhejiang Province [J]. Acta Ecologica Sinica, 2008, 28(9): 4547−4558. [12] 姚俊宇, 齐锦秋, 张柳桦, 等. 人为干扰对碧峰峡山矾次生林群落物种多样性和土壤理化性质的影响[J]. 生态学杂志, 2018, 37(10): 2942−2950. YAO Junyu, QI Jinqiu, ZHANG Liuhua, et al. Effects of anthropogenic disturbance on species diversity and soil physicochemical properties of Symplocos sumuntia secondary forest in Bifengxia [J]. Chinese Journal of Ecology, 2018, 37(10): 2942−2950. [13] 王媚臻, 毕浩杰, 金锁, 等. 林分密度对云顶山柏木人工林林下物种多样性和土壤理化性质的影响[J]. 生态学报, 2019, 39(3): 981−988. WANG Meizhen, BI Haojie, JIN Suo, et al. Effects of stand density on understory species diversity and soil physicochemical properties of a Cupressus funebris plantation in Yunding Mountain [J]. Acta Ecologica Sinica, 2019, 39(3): 981−988. [14] 童跃伟, 屈利利, 符庆响, 等. 大别山南坡森林植物群落物种多样性及其与海拔因子的关系[J]. 生态学报, 2024, 44(12): 1−11. TONG Yuewei, QU Lili, FU Qingxiang, et al. Species diversity of forest plant communities on the southern slope of the Dabie Mountain and its relationship with altitude factors [J]. Acta Ecologica Sinica, 2024, 44(12): 1−11. [15] 温佩颖, 金光泽. 地形对阔叶红松林物种多样性的影响[J]. 生态学报, 2019, 39(3): 945−956. WEN Peiying, JIN Guangze. Effects of topography on species diversity in a typical mixed broadleaved-Korean pine forest [J]. Acta Ecologica Sinica, 2019, 39(3): 945−956. [16] WEIGEL R, GILLES J, KLISZ M, et al. Forest understory vegetation is more related to soil than to climate towards the cold distribution margin of European beech [J]. Journal of Vegetation Science, 2019, 30(4): 746−755. [17] 龚艳宾, 郭建斌, 赵秀梅, 等. 吉林蛟河天然阔叶红松林草本植物多样性及其与土壤因子的关系[J]. 浙江农林大学学报, 2016, 33(4): 620−628. GONG Yanbin, GUO Jianbin, ZHAO Xiumei, et al. Herbaceous species diversity as related to soil factors in a Korean pine broadleaved forest of Jiaohe, Jilin, Province [J]. Journal of Zhejiang A&F University, 2016, 33(4): 620−628. [18] YAMAMOTO N, YOKOYAMA J, KAWATA M. Relative resource abundance explains butterfly biodiversity in island communities [J]. Proceedings of the National Academy of Sciences, 2007, 104(25): 10524−10529. [19] 朱夏夏, 张华, 朱岩, 等. 老秃顶子石河冰缘地貌森林群落物种多样性及其影响因素[J]. 植物科学学报, 2016, 34(1): 67−77. ZHU Xiaxia, ZHANG Hua, ZHU Yan, et al. Forest community species diversity and the influencing factors in the rock stream periglacial landforms of Mt. Laotudingzi [J]. Plant Science Journal, 2016, 34(1): 67−77. [20] 张荣, 李婷婷, 金锁, 等. 人为干扰对蒙顶山木荷次生林物种多样性及土壤理化性质的影响[J]. 浙江农林大学学报, 2020, 37(5): 867−875. ZHANG Rong, LI Tingting, JIN Suo, et al. Effects of human disturbance on species diversity and soil physiochemical properties of Schima superba community in Mengding Mountain [J]. Journal of Zhejiang A&F University, 2020, 37(5): 867−875. [21] 南国卫, 王静慧, 秦淑莹, 等. 不同恢复年限刺槐林林下草本层的物种多样性[J]. 浙江农林大学学报, 2024, 41 (5): 986−993. NAN Guowei, WANG Jinghui, QIN Shuying, et al. Diversity characteristics of herbaceous species under Robinia pseudoacacia forest in different years [J]. Journal of Zhejiang A&F University, 2024, 41 (5): 986−993. [22] 方精云, 王襄平, 沈泽昊, 等. 植物群落清查的主要内容、方法和技术规范[J]. 生物多样性, 2009, 17(6): 533−548. FANG Jingyun, WANG Xiangping, SHEN Zehao, et al. Methods and protocols for plant community inventory [J]. Biodiversity Science, 2009, 17(6): 533−548. [23] 侯红亚, 王立海. 小兴安岭阔叶红松林物种组成及主要种群的空间分布格局[J]. 应用生态学报, 2013, 24(11): 3043−3049. HOU Hongya, WANG Lihai. Species composition and main populations spatial distribution pattern in Korean pine broadleaved forest in Xiaoxing’an Mountain of Northeast China [J]. Chinese Journal of Applied Ecology, 2013, 24(11): 3043−3049. [24] MICHEA E, DASMANN F. Conservation biology: the science of scarcity and diversity [J]. Trends in Ecology and Evolution, 1987, 2: 169−170. [25] KANG Di, GUO Yaoxin, REN Chengjie, et al. Population structure and spatial pattern of main tree species in secondary Betual platyphylla forest in Ziwuling Mountain, China [J/OL]. Scientific Reports, 2014, 4 : 6873[2024-06-27]. DOI: 10.1038/srep06873. [26] 任学敏, 杨改河, 王得祥, 等. 环境因子对巴山冷杉-糙皮桦混交林物种分布及多样性的影响[J]. 生态学报, 2012, 32(2): 605−613. REN Xuemin, YANG Gaihe, WANG Dexiang, et al. Effects of environmental factors on species distribution and diversity in an Abies fargesii-Betula utilis mixed forest. [J]. Acta Ecologica Sinica, 2012, 32(2): 605−613. [27] FEEST A, ALDRED T, JEDAMZIK K. Biodiversity quality: a paradigm for biodiversity [J]. Nature Precedings, 2009, 10(6): 1077−1082. [28] GAMFELDT L, SNӒLL T, BAGCHI R, et al. Higher levels of multiple ecosystem services are found in forests with more tree species [J/OL]. Nature Communications, 2013, 4 (1): 1340[2024-06-27]. DOI: 10.1038/ncomms2328. [29] 于世勇, 张信杰, 刘波, 等. 我国寒温带典型森林群落特征及生物多样性研究[J]. 温带林业研究, 2023, 6(1): 1−6. YU Shiyong, ZHANG Xinjie, LIU Bo, et al. Research on the characteristics and biodiversity of typical cold-temperate forest communities in China [J]. Journal of Temperate Forestry Research, 2023, 6(1): 1−6. [30] 叶铎, 董瑞瑞, 米湘成, 等. 古田山常绿阔叶林萌生特征及其与群落物种多样性的关系[J]. 生物多样性, 2017, 25(4): 393−400. YE Duo, DONG Ruirui, MI Xiangcheng, et al. Characteristics and effects of sprouting on species diversity in a subtropical evergreen broad-leaved forest in Gutianshan, East China [J]. Biodiversity Science, 2017, 25(4): 393−400. [31] 邹朋峻, 关庆伟, 袁在翔, 等. 紫金山南麓枫香种群结构与动态特征[J]. 南京林业大学学报(自然科学版), 2023, 47(3): 157−163. ZOU Pengjun, GUAN Qingwei, YUAN Zaixiang, et al. The population structure and dynamics of Liquidambar formosana on the southeast foothill of Zijin Mountain, Nanjing City [J]. Journal of Nanjing Forestry University (Natural Science Edition), 2023, 47(3): 157−163. [32] 宋庆丰, 王兵, 牛香, 等. 江西大岗山低海拔常绿阔叶林物种组成与群落结构特征[J]. 生态学杂志, 2020, 39(2): 384−393. SONG Qingfeng, WANG Bing, NIU Xiang, et al. The characteristic of species composition and community structure of low-altitude evergreen broad-leaved forest in Dagang Mountain, Jiangxi Province [J]. Chinese Journal of Ecology, 2020, 39(2): 384−393. [33] 叶万辉, 曹洪麟, 黄忠良, 等. 鼎湖山南亚热带常绿阔叶林20公顷样地群落特征研究[J]. 植物生态学报, 2008, 32(2): 274−286. YE Wanhui, CAO Honglin, HUANG Zhongliang, et al. Community structure of a 20 hm2 lower subtropical evergreen broadleaved forest plot in Dinghushan, China [J]. Chinese Journal of Plant Ecology, 2008, 32(2): 274−286. [34] 达良俊, 杨永川, 宋永昌. 浙江天童国家森林公园常绿阔叶林主要组成种的种群结构及更新类型[J]. 植物生态学报, 2004, 28(3): 376−384. DA Liangjun, YANG Yongchuan, SONG Yongchang. Population structure and regeneration types of dominant species in an evergreen broadleaved forest in Tiantong National Forest Park, Zhejiang Province, Eastern China [J]. Acta Phytoecologica Sinica, 2004, 28(3): 376−384. [35] 王云泉, 田磊, 仲磊, 等. 东白山自然保护区木荷-马尾松群落结构及物种多样性分析[J]. 浙江大学学报(理学版), 2015, 42(1): 38−46. WANG Yunquan, TIAN Lei, ZHONG Lei, et al. Community structure and species diversity of Schima superba-Pinus massoniana communities in Dongbaishan Nature Reserve [J]. Journal of Zhejiang University (Science Edition), 2015, 42(1): 38−46. [36] 韩铭, 李华, 蔡体久. 黑龙江太平沟国家级自然保护区森林群落植物多样性特征[J]. 森林工程, 2023, 39(5): 40−47. HAN Ming, LI Hua, CAI Tijiu. Plant diversity of forest community in Taipinggou National Nature Reserve, Heilongjiang Province [J]. Forest Engineering, 2023, 39(5): 40−47. [37] 李善家, 王兴敏, 刘海峰, 等. 河西走廊荒漠植物多样性及其对环境因子的响应[J]. 生态环境学报, 2023, 32(3): 429−438. LI Shanjia, WANG Xingmin, LIU Haifeng, et al. Diversity of desert plants in Hexi Corridor and its response to environmental factors [J]. Ecology and Environmental Sciences, 2023, 32(3): 429−438. [38] SUDING K N, GROSS K L, HOUSEMAN G R. Alternative states and positive feedbacks in restoration ecology [J]. Trends in Ecology & Evolution, 2004, 19(1): 46−53. [39] VITOUSEK P M, HOWARTH R W. Nitrogen limitation on land and in the sea: how can it occur? [J]. Biogeochemistry, 1991, 13: 87−115. [40] 陈晓熹, 杨新东, 曾献兴, 等. 环境因子对青云山自然保护区森林群落物种分布的影响[J]. 生态学杂志, 2019, 38(12): 3642−3650. CHEN Xiaoxi, YANG Xindong, ZENG Xianxing, et al. Influences of environmental factors on species distribution in forest community in Wengyuan Qingyunshan Nature Reserve, Guangdong [J]. Chinese Journal of Ecology, 2019, 38(12): 3642−3650. [41] 姜小蕾, 孙振元, 郝青, 等. 崂山次生林群落β多样性格局及其组分的驱动因素[J]. 生态学杂志, 2020, 39(10): 3211−3220. JIANG Xiaolei, SUN Zhenyuan, HAO Qing, et al. Interpretation of environmental factors affecting beta diversity and its components of secondary forest in Lao Mountain [J]. Chinese Journal of Ecology, 2020, 39(10): 3211−3220. [42] 李婷婷, 唐永彬, 周润惠, 等. 云顶山不同人工林林下植物多样性及其与土壤理化性质的关系[J]. 生态学报, 2021, 41(3): 1168−1177. LI Tingting, TANG Yongbin, ZHOU Runhui, et al. Understory plant diversity and its relationship with soil physicochemical properties in different plantations in Yunding Mountain [J]. Acta Ecologica Sinica, 2021, 41(3): 1168−1177. -

-
链接本文:
https://zlxb.zafu.edu.cn/article/doi/10.11833/j.issn.2095-0756.20240416
 点击查看大图
点击查看大图
计量
- 文章访问数: 1578
- HTML全文浏览量: 474
- PDF下载量: 39
- 被引次数: 0
