




-
森林群落内包含着复杂的种间关系,如竞争、寄生、互利共生等[1]。生态位在研究群落物种组成、生物多样性和物种共存等方面有着重要作用[2],它是植物群落中各种群所能利用各种资源的总和及其与相关种群之间的功能关系,体现了种群在群落中的地位、作用和重要性[3],以生态位宽度和生态位重叠指数表征分别体现物种在群落中的地位和种间的联系[4]。种间联结是指不同物种在空间分布上的相互关联性,对生物多样性保护和自然植被的恢复具有一定的指导作用,对揭示群落内的种间关系、演替动态有着重要的理论意义和参考价值[5]。种间关系可分为正联结、无联结和负联结等3种。方庆等[6]认为可以保护与细果秤锤树Sinojackia microcarpa正联结性较高的物种,以期达到保护该濒危物种的目的;丁茂等[7]对落叶阔叶林进行动态监测后发现,种对间正关联数量有所增加,植被群落趋向于稳定的正演替方向发展,说明物种间的正联结性的增加能够提高森林群落的稳定性和物种多样性;杨春玉等[8]发现退化的喀斯特森林从乔灌阶段恢复到乔林阶段,物种种间正联结对数持续增加,直至形成顶级群落,解释了森林群落最终是朝着物种间互利共生和结构稳定的方向发展。但这些研究主要集中于低海拔区域,对于中高海拔地区亚热带森林群落的研究则较少。
浙江巾子峰国家森林公园拥有原生亚热带常绿阔叶林,平均海拔800 m以上,人为活动较少,使得该区域保存有较为丰富且处于不同演替阶段的植被群落,开展该区域森林群落生态位和种间联结的研究,对于中高海拔地区常绿阔叶林的生物多样性保护具有积极意义[9]。对于研究区周边植物群落的研究主要集中在常绿阔叶林种群结构与分布格局[10]、物种组成与群落结构[11]、甜槠Castanopsis eyrei种群特征[12]等,而该地区常绿阔叶林的生态位和种间联结性的研究还未见报道。因此,本研究选取浙江巾子峰国家森林公园境内百丈林区常绿阔叶林作为研究对象,在野外调查的基础上,采用Levins生态位宽度(BL)、Shannon生态位宽度(BS)、Pianka生态位重叠指数(Oik)、Schoener系数(Cik)、方差比率(VR)、卡方($ {\chi}^{2} $)检验、Pearson相关系数和Spearman秩相关系数等方法,研究常绿阔叶林群落的木本植物组成以及优势种的生态位和种间联结性的关系,旨在为亚热带常绿阔叶林生物多样性和植被的保护与恢复提供参考。
-
浙江巾子峰国家森林公园位于浙江省庆元县(27°27′~27°39′N,118°50′~119°06′E),属武夷山系洞宫山脉,总面积为5 752 hm2[13]。年均气温为17.2 ℃,最热月平均气温为26.9 ℃,最冷月平均气温为7.0 ℃,年均降水量为1 689.0 mm,相对湿度为75.8%,年均无霜期为256 d,主要植被类型包括常绿阔叶林、常绿-落叶阔叶混交林、针阔混交林、针叶林、毛竹Phyllostachys edulis林等。样地所在地土壤为黄壤。
-
参照美国热带森林研究中心(CTFS)的方法[14],在浙江巾子峰国家森林公园境内的百丈林区(27°38′N,118°91′E)建立了1个1.4 hm2 (140 m×100 m)和6个0.4 hm2 (20 m×20 m)的常绿阔叶林长期监测样地。对样地内所有胸径(DBH)≥1 cm的木本植物进行每木检尺,并记录种名、胸径、树高、枝下高、空间坐标及生活状态等,样地基本情况如表1。
表 1 研究区常绿阔叶林群落样地基本情况
Table 1. Basic situation of survey sites for broadleaf evergreen forest community survey in the study area
样地号 类型 海拔/m 群落类型 样地面积/m2 坡位 密度/(株·hm−2) 平均胸径/cm 郁闭度 1 天然林 1 163 常绿阔叶林 14 000 上 3 698 7.20 75.8 2 天然林 1 163 常绿阔叶林 400 上 2 750 9.87 93.9 3 天然林 1 146 常绿阔叶林 400 中 4 775 8.39 94.5 4 天然林 1 136 常绿阔叶林 400 中 2 100 12.15 90.7 5 天然林 1 151 常绿阔叶林 400 中 4 150 6.25 88.2 6 天然林 1 121 常绿阔叶林 400 中 1 525 16.86 93.4 7 天然林 1 176 常绿阔叶林 400 上 2 300 11.82 94.0 -
依据马克平等[15]的方法计算物种重要值确定优势种。
-
依据王刚等[16]的方法计算木本植物优势种的Levins生态位宽度指数(BL)和Shannon生态位宽度指数(BS)。依据郭平平等[17]的方法计算Pianka生态位重叠指数(Oik);依据SCHOENER[18]的方法计算Schoener系数(Cik)。
-
计算方法参照SCHLUTER[19]提出的方差比率法(VR)和检验统计量(W)。参照江常春等[20]的方法,卡方($ {\chi}^{2}) $检验采用YATES连续矫正公式;参照吕增伟等[21]的方法计算Spearman秩相关系数和Pearson相关系数。
-
重要值、生态位和种间联结性采用R 4.3.1软件(“spaa”“corrplot”“ggplot”“psych”“vegan”“tidyverse”工具包)[22]和WPS 2021进行数据计算以及绘图。
-
调查样地中木本植物共有113种,隶属于31科62属。由表2可知:在该常绿阔叶林群落木本植物中,筛选出重要值$\geqslant $1%的优势种,共21种,重要值总和为80.32%。在木本植物优势种重要值大小排序中,木荷Schima superba的重要值最大,为14.30%,其次为甜槠和黄山松Pinus taiwanensis,分别为12.85%和12.23%。薄叶山矾Symplocos anomala和华中樱Cerasus conradinae的重要值最小,都为1.02%。这21个优势种的BL和BS平均值分别为17.38和3.03。木荷、甜槠、黄山松、锥栗Castanea henryi和野漆树Toxicodendron succedaneum的生态位宽度相对较大,且BL和BS分别大于20.00和3.30。
表 2 优势植物重要值与生态位宽度
Table 2. Important value and niche breadth of dominant plant species
编号 种名 个体数 分布频度 重要值/% Levins生态位宽度(BL) Shannon生态位宽度(BS) sp1 木荷 Schima superba 984 41 14.30 25.91 3.42 sp2 甜槠 Castanopsis eyrei 682 41 12.85 28.20 3.49 sp3 黄山松 Pinus taiwanensis 245 40 12.23 29.67 3.52 sp4 锥栗 Castanea henryi 261 38 6.02 21.39 3.30 sp5 马银花 Rhododendron ovatum 665 38 5.47 19.43 3.23 sp6 麂角杜鹃 Rhododendron latoucheae 500 35 4.30 18.47 3.19 sp7 大萼黄瑞木Adinandra glischroloma var. macrosepala 284 33 2.83 4.85 2.44 sp8 江南山柳 Clethra delavayi 189 35 2.45 17.07 3.17 sp9 野漆树 Toxicodendron succedaneum 112 33 2.21 24.31 3.32 sp10 浙江新木姜子 Neolitsea chekiangensis 157 28 1.91 11.83 2.79 sp11 微毛柃 Eurya hebeclados 152 27 1.87 12.21 2.89 sp12 短柄枹栎 Quercus glandulifera 115 27 1.86 19.59 3.11 sp13 江南越橘 Vaccinium mandarinorum 135 29 1.75 16.06 3.04 sp14 小叶青冈 Cyclobalanopsis gracilis 90 23 1.62 10.83 2.69 sp15 黄背越橘 Vaccinium iteophyllum 100 27 1.53 19.38 3.11 sp16 树参 Dendropanax dentiger 97 23 1.48 10.04 2.64 sp17 云山青冈 Cyclobalanopsis sessilifolia 72 22 1.37 14.64 2.84 sp18 红楠 Machilus thunbergii 50 22 1.15 13.30 2.82 sp19 吴茱萸五加 Gamblea ciliata var. evodiifolia 47 22 1.09 17.96 2.98 sp20 薄叶山矾 Symplocos anomala 43 20 1.02 14.11 2.81 sp21 华中樱 Cerasus conradinae 38 21 1.02 15.70 2.89 -
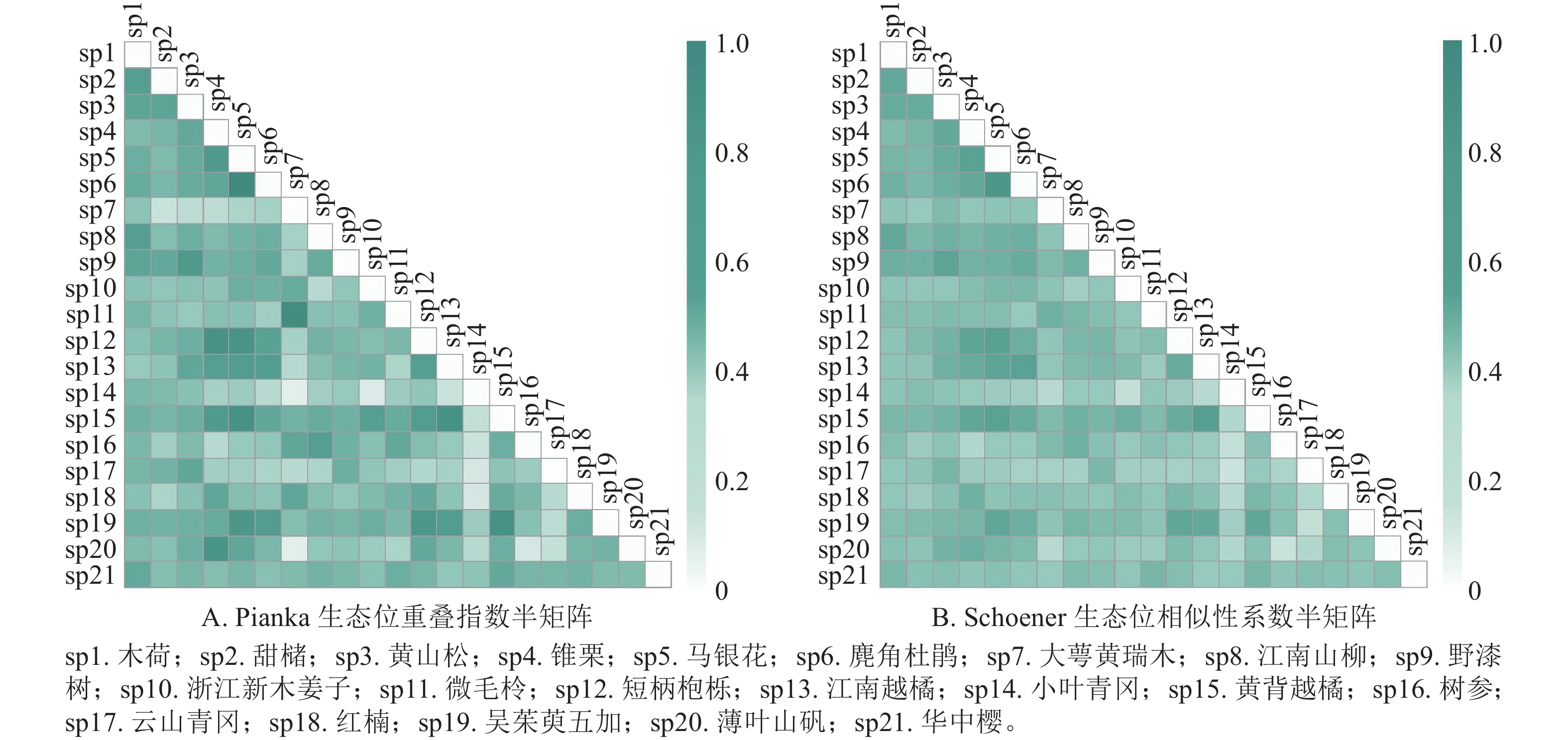
如图1A所示:浙江巾子峰国家森林公园常绿阔叶林木本植物优势种共有210个种对,Oik为0.09~0.84。Oik≥0.5的共有89个种对,占总对数的42.4%,说明这些种对在资源不足时,可能会发生激烈的竞争。Oik<0.5的共有121对,占总对数的57.6%。
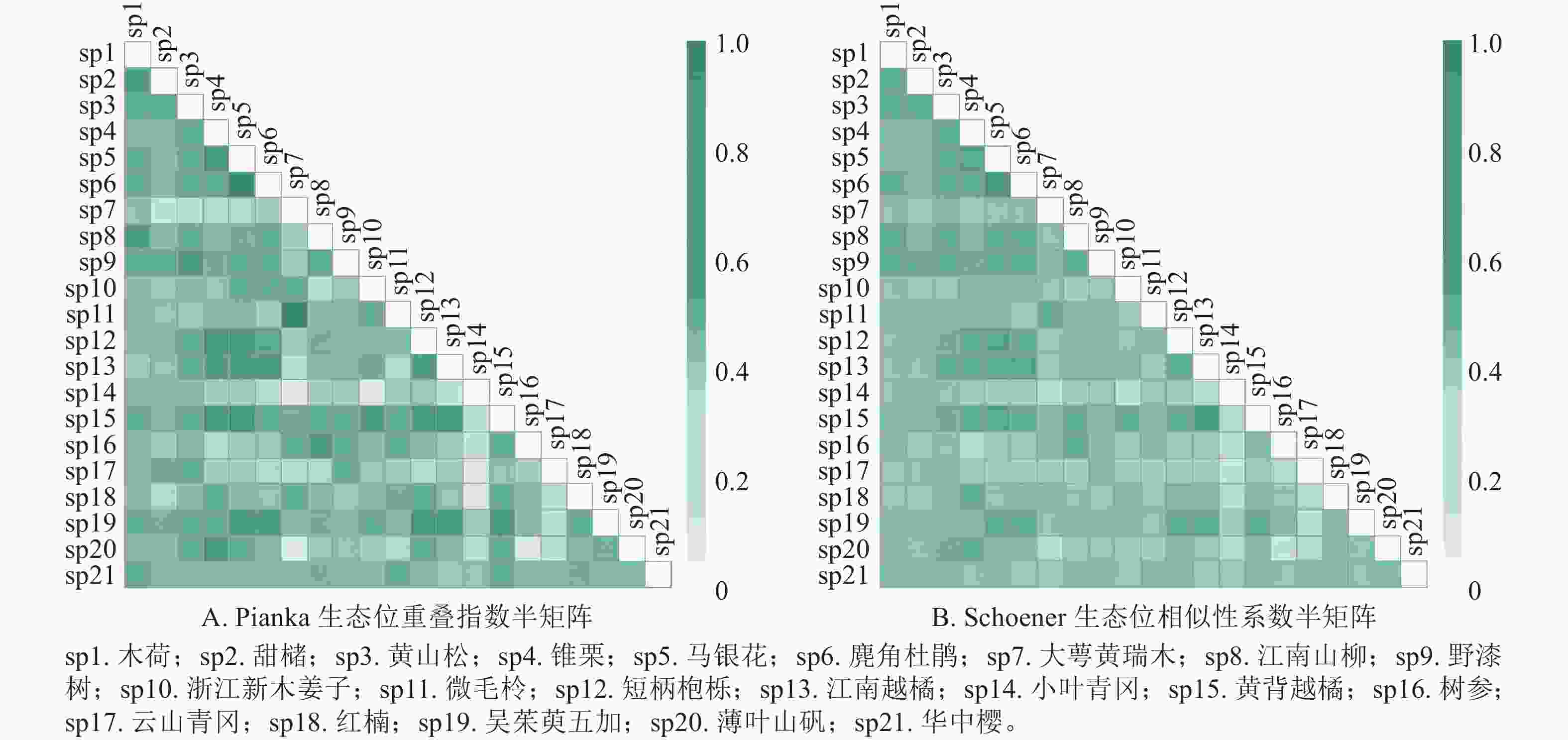
图 1 主要优势树种生态位重叠指数
Figure 1. Niche overlap index of dominant tree species
如图1B所示:群落中木本植物优势种的Cik为0.17~0.74。Cik≥0.5的种对共50对,占总对数的23.8%,这些种对的生态习性较为相似。Cik<0.5的种对共160对,占总对数的76.2%。
-
对木本植物优势种总体关联性分析可知(表3),方差比率(VR)= 2.84>1,表明该常绿阔叶林木本植物优势种之间总体联结呈现正联结。卡方临界值χ2(0.95, 41)= 27.33,χ2(0.05, 41)= 56.94,检验统计量(W)= 116.73,不在临界值之间,表明木本植物优势种之间联结显著(P<0.05)。该群落木本植物优势种种间总体呈现显著正联结(P<0.05),该群落处于稳定阶段。
表 3 优势植物总体关联性
Table 3. Overall associations of dominant plant species
方差比率(VR) 检验统计量(W) 卡方临界值(0.95, 41) 卡方临界值(0.05, 41) 检验结果 2.84 116.73 27.33 56.94 显著正联结 -
通过卡方检验观察到的频数与期望频数之间的差异,结果如表4和表5所示。在21个木本植物优势树种组成的210个种对中,39个种对间无关联,占总对数的18.57%;112个种对间呈不显著正联结(P>0.05),占总对数的53.33%;49个种对间呈不显著负联结(P>0.05),占总对数的23.33%;2个种对间呈极显著正联结(P<0.01);8个种对间呈显著正联结(P<0.05)。
表 4 主要优势树种χ2统计量检验
Table 4. χ2 correlation test of dominant population of dominant tree species
编号 sp1 sp2 sp3 sp4 sp5 sp6 sp7 sp8 sp9 sp10 sp11 sp12 sp13 sp14 sp15 sp16 sp17 sp18 sp19 sp20 sp2 0.00 sp3 0.00 0.00 sp4 0.00 0.00 2.75 sp5 0.00 0.00 2.75 0.42 sp6 0.00 0.00 1.03 3.24 3.24 sp7 0.00 0.00 0.61 0.02 1.92 2.20 sp8 0.00 0.00 1.03 0.01 0.01 4.11 0.13 sp9 0.00 0.00 0.61 0.02 0.02 6.75 0.87 2.20 sp10 0.00 0.00 0.16 0.34 0.34 0.15 0.00 2.30 0.67 sp11 0.00 0.00 0.11 0.36 0.36 1.83 2.16 1.83 0.04 0.00 sp12 0.00 0.00 0.11 3.48 0.36 10.34 5.29 1.83 5.29 0.00 1.43 sp13 0.00 0.00 0.21 0.67 4.57 0.52 3.50 0.06 0.02 0.26 0.19 3.02 sp14 0.00 0.00 0.02 0.05 0.05 0.59 0.00 0.59 0.00 0.02 0.81 0.06 1.50 sp15 0.00 0.00 0.11 0.36 3.48 1.83 2.16 1.83 0.41 2.13 1.43 3.57 6.07 0.06 sp16 0.00 0.00 0.02 0.97 0.05 0.01 0.00 0.01 0.62 0.02 0.06 0.81 0.73 0.79 0.06 sp17 0.00 0.00 0.01 0.02 1.15 1.29 0.03 0.06 0.03 0.10 0.00 0.43 1.78 1.35 0.00 0.53 sp18 0.00 0.00 0.01 0.02 0.02 0.41 0.39 0.06 0.39 0.10 0.00 3.96 1.78 0.28 1.77 0.53 0.04 sp19 0.00 0.00 0.01 0.02 0.02 0.41 2.01 0.06 2.01 0.10 1.77 3.96 0.42 0.01 3.96 1.86 2.10 0.19 sp20 0.00 0.00 0.00 0.00 1.34 1.59 0.10 0.14 0.22 0.01 0.77 0.05 0.86 0.03 0.05 1.17 0.02 0.02 0.02 sp21 0.00 0.00 0.00 0.00 0.00 0.26 0.22 0.26 0.22 0.61 0.20 0.05 1.28 0.21 0.05 0.21 0.60 0.02 0.02 0.03 说明:sp1. 木荷;sp2. 甜槠;sp3. 黄山松;sp4. 锥栗;sp5. 马银花;sp6. 麂角杜鹃;sp7. 大萼黄瑞木;sp8. 江南山柳;sp9. 野漆树;sp10. 浙江新木姜子;sp11. 微毛柃;sp12. 短柄枹栎;sp13. 江南越橘;sp14. 小叶青冈;sp15. 黄背越橘;sp16. 树参;sp17. 云山青冈;sp18. 红楠;sp19. 吴茱萸五加;sp20. 薄叶山矾;sp21. 华中樱。 表 5 主要优势树种的Spearman秩相关系数、Pearson相关系数检验结果比较
Table 5. Comparison of the results of Spearman rank correlation and Pearson rank correlation test of dominant tree species
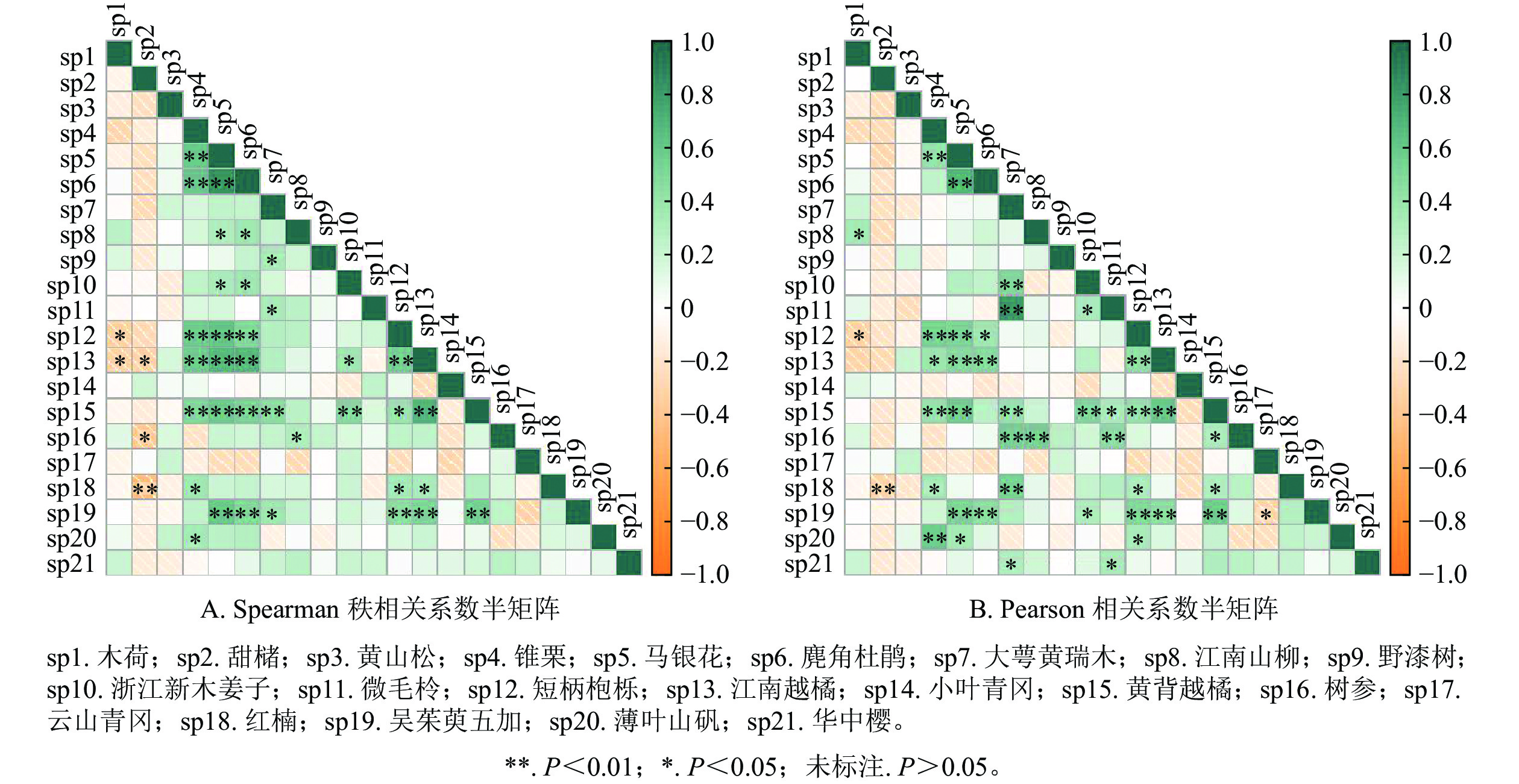
检验方法 正联结(相关)/对 负联结(相关)/对 无联结/对 极显著 显著 不显著 总数 极显著 显著 不显著 总数 总数 χ2检验 2 8 112 122 0 0 49 49 39 Spearman秩相关系数 21 14 108 142 1 4 63 68 0 Pearson相关系数 25 14 89 128 1 2 79 82 0 说明:极显著P<0.01;显著P<0.05;不显著P>0.05。 Spearman检验结果如图2A和表5所示。呈正联结的种对有142对,占总种对数的67.6%。其中,呈极显著正联结(P<0.01)的种对有21对,显著正联结(P<0.05)的种对有14对。呈负联结的种对有68对,占总种对数的32.4%。其中,呈现显著负联结(P<0.01)的种对有1对,显著负联结(P<0.05)的种对有4对。极显著和显著正、负联结比值为7∶1。Spearman秩相关系数分析仍保持与卡方检验相一致的结果。
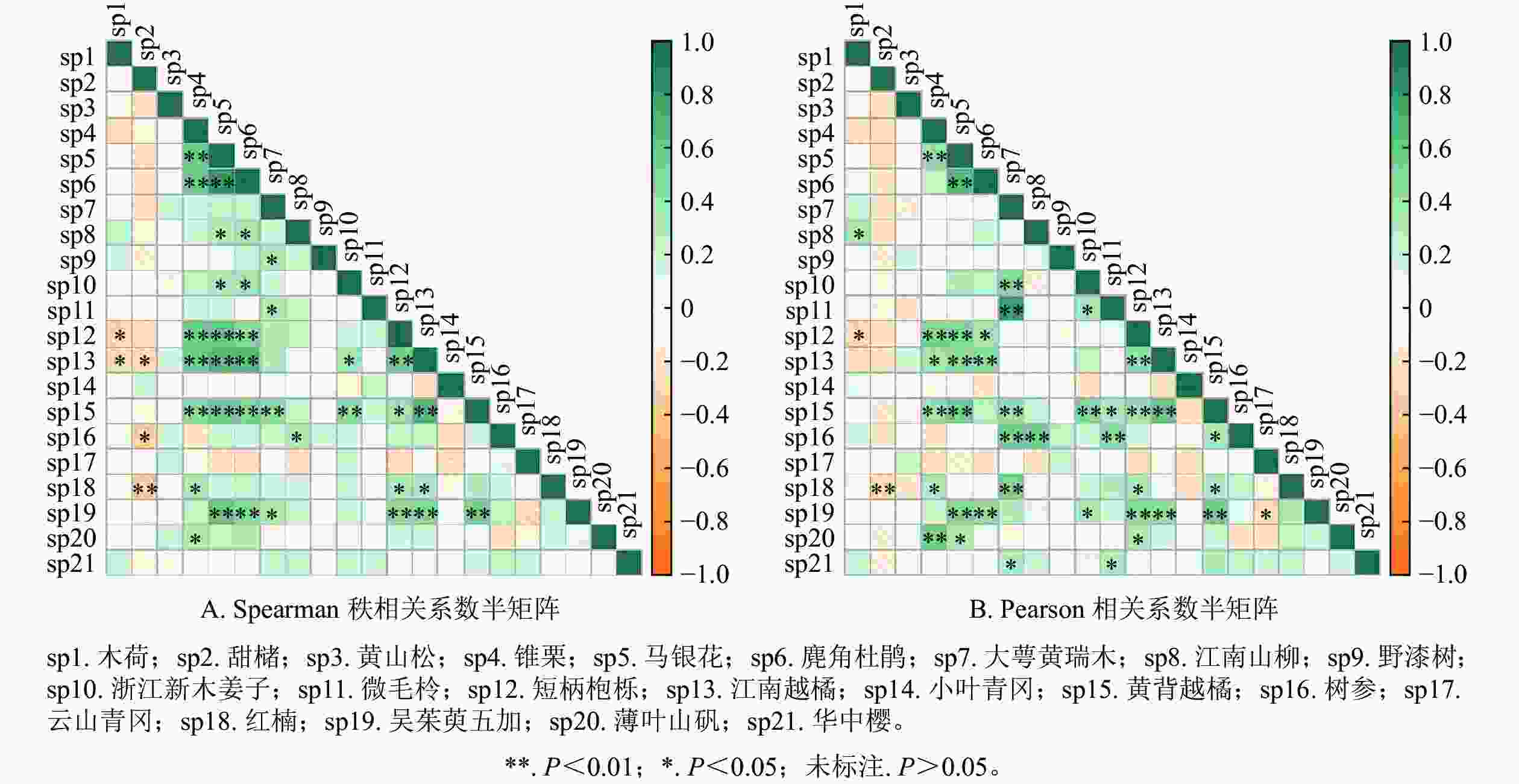
图 2 相关系数半矩阵
Figure 2. Semi-matrix of correlation coefficients
Pearson检验结果如图2B和表5所示。在210个种对中,呈正联结的有128对,占总对数的60.96%;呈负联结的有82对,占总对数的39.04%。在正联结的128个种对中,呈极显著正联结(P<0.01)的有25对,显著正联结(P<0.05)的有14对;在负联结的82个种对中,呈极显著负联结(P<0.01)的有1对,显著负联结(P<0.05)的有2对。极显著和显著正、负联结比值为13∶1。Pearson相关系数分析发现,不显著联结(P>0.05)的种对数仍保持较大一部分,这符合卡方检验结果。
-
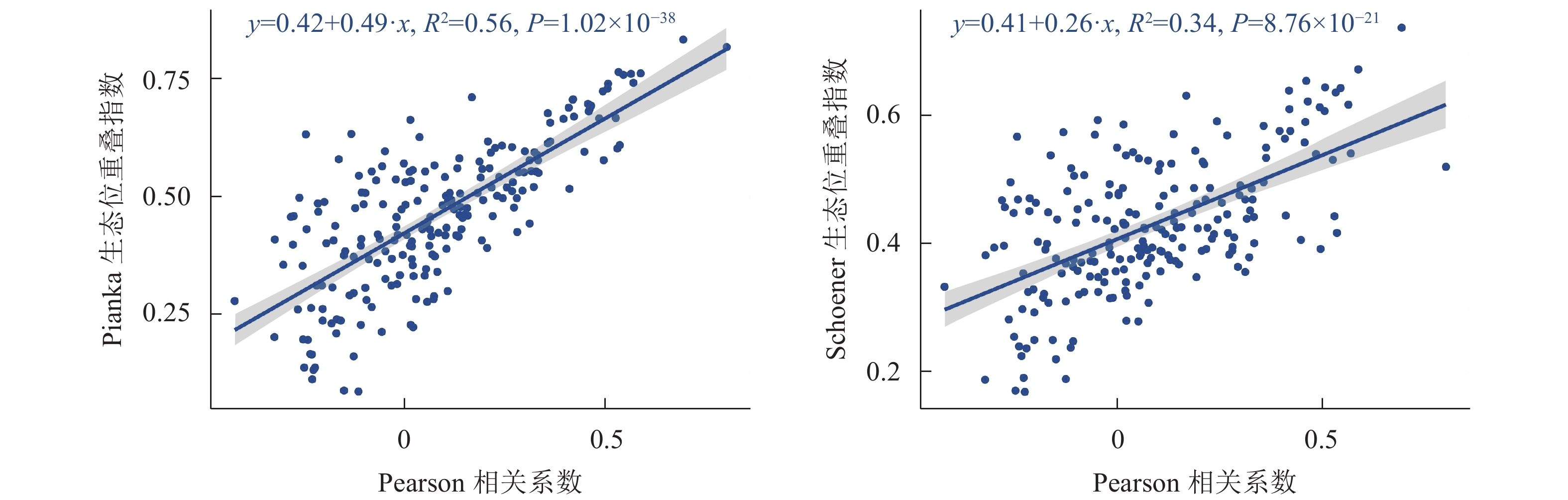
木本植物优势种的Pearson相关系数与生态位重叠指数的回归分析如图3所示,木本植物优势种种间联结与生态位重叠呈极显著正相关(P<0.01)。
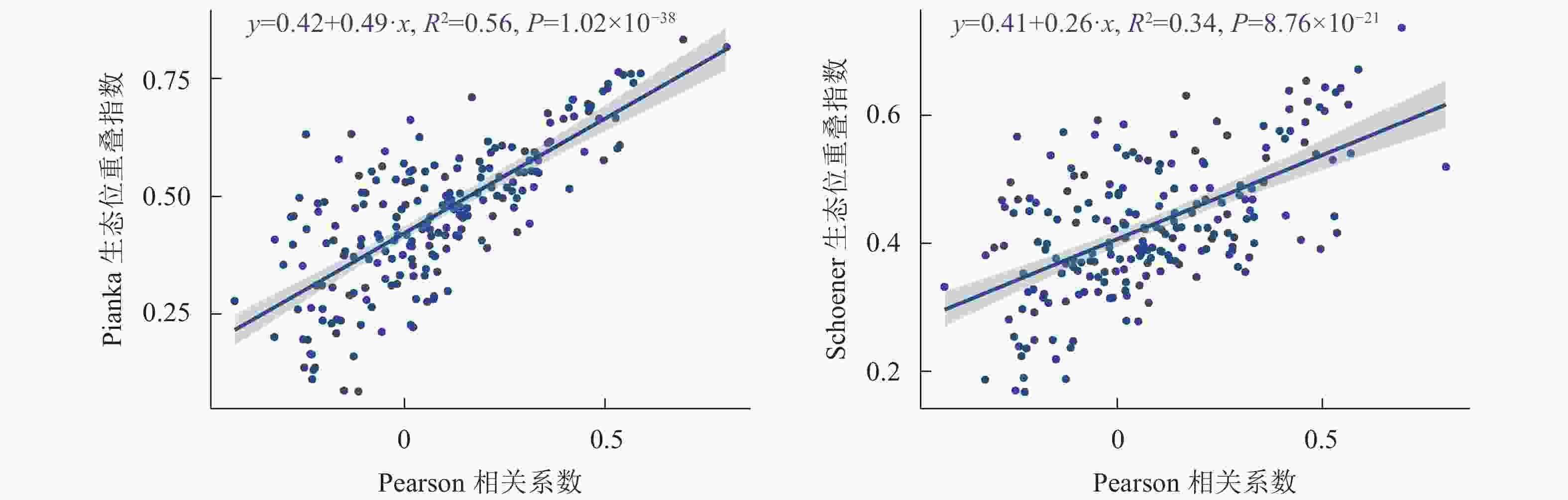
图 3 主要优势树种Pearson与生态位重叠值的回归分析
Figure 3. Regression analysis of Pearson rank correlation with niche overlap of dominant tree species
-
在浙江巾子峰国家森林公园常绿阔叶林中,木荷、甜槠和黄山松的重要值、生态位宽度较大。在一个群落中,某一物种的重要值与生态位宽度呈正相关,即重要值越大,其生态位宽度也越大[23],但叶兴状等[24]研究发现,福建天台山半枫荷Semiliquidambar cathayensis群落中重要值排第2位的半枫荷,其生态位宽度却排第4位。本研究中,黄山松的生态位宽度最大,其次是甜槠和木荷;木荷虽然在该群落中的重要值最大,但其生态位宽度略低于黄山松和甜槠。大萼黄瑞木Adinandra glischroloma var. macrosepala的重要值虽然排在第7位,但其生态位宽度最小,结合实际野外调查,大萼黄瑞木高度聚集分布在同一个样方中,其重要值大小和生态位宽度在一定程度上受到分布频度的影响,这与刘雨婷等[25]的研究结果相似,表明它对该群落环境的适应性和资源的利用能力不全面,竞争性较弱,有可能发展为特化种。
生态位重叠指数和生态位相似比系数可以反映物种对资源的利用程度和环境的适应性[26]。物种的生态位宽度越大,则生态位重叠指数越大,对有共同需求的资源竞争压力也越大[27]。本研究发现,生态位宽度最大的黄山松与甜槠、锥栗、马银花Rhododendron ovatum、麂角杜鹃Rhododendron latoucheae、江南山柳Clethra delavayi、野漆树、短柄枹栎Quercus serrata、江南越橘Vaccinium mandarinorum、黄背越橘Vaccinium iteophyllum和薄叶山矾的生态位重叠指数和生态位相似比系数均超过0.50。吴友贵等[28]发现生态位宽度最大的多脉青冈Cyclobalanopsis multinervis、麂角杜鹃和窄基红褐柃Eurya rubiginosa之间的生态位相似比系数都大于0.7。本研究表明:生态位宽度较小的微毛柃Eurya hebeclados-大萼黄瑞木的生态位重叠值和生态位相似比系数均较大,其值分别为0.82 和0.52,与栾兆平等[29]研究结果相似,即生态位宽度较小的物种,其生态位重叠指数超过了0.90。
种间联结是物种在空间上的不同关系,是物种对环境的适应性和对资源利用相似性的体现[30]。浙江巾子峰国家森林公园常绿阔叶林木本植物优势种总体呈显著正联结。结合卡方检验、Spearman秩相关系数和Pearson相关系数可以较好地判断群落中木本植物优势种种对间的联结性,发现呈正联结的种对数大于负联结的种对数,极显著或显著正联结的种对数也大于极显著或显著负联结的种对数,表明该群落属于演替中期,种间的竞争和协作逐渐平衡。可能与该样地海拔较高,人为活动较少有关[26]。张滋芳等[31]认为物种间有相似环境适应性和资源需求,往往会产生正向联系。本研究马银花与麂角杜鹃、黄背越橘,江南越橘与黄背越橘、麂角杜鹃均呈极显著正联结,因为4种植物均为杜鹃花科灌木,有着相似的生态习性;短柄枹栎与锥栗、吴茱萸五加Gamblea ciliataevodiifolia的生态位宽度在木本植物优势种中占据较高位置,也呈极显著正联结,这与张盟等[4]发现有着紧密正联结的物种往往占据较高生态位宽度相似。而红楠Machilus thunbergii-甜槠呈极显著负联结,可能是甜槠多为大径个体,红楠以小径个体为主,高大繁茂的甜槠限制了树高较小的红楠生长。短柄枹栎-木荷呈显著负联结,因为木荷为常绿树种,短柄枹栎为落叶树种,2个物种对光照、水分等需求不同,这与罗敏贤等[32]认为常绿树种和落叶树种生长所需光性不同,会导致物种间呈显著负联结相一致。
植物之间生态位重叠值与种间联结若呈正联结,表明其存在不同程度的生态位重叠,反之亦然[33]。在本研究发现,浙江巾子峰国家森林公园主要优势树种的Pearson相关系数与生态位重叠的回归分析呈极显著正相关,如马银花-麂角杜鹃种对间呈极显著正联结,种对间的Pianka重叠值为0.84,其Pearson相关系数为0.69,呈正向关系,这与郑俊鸣等[34]认为物种种间正联结越强,其生态位重叠指数也越大的研究结果相似。
-
浙江巾子峰国家森林公园常绿阔叶林属于比较稳定的生态系统,可以选择具有较宽的生态位和适应性较强的树种,如木荷、甜槠、锥栗、马尾松、马银花、麂角杜鹃和短柄枹栎等作为自然植被恢复的优先树种;而对于小叶青冈Cyclobalanopsis gracilis、大萼黄瑞木、薄叶山矾、云山青冈Cyclobalanopsis sessilifolia和红楠等树种,其生态位重叠值和生态位相似系数较低,生态位宽度也较低,应在自然植被恢复和生物多样性保护中进行一定调控。
-
巾子峰国家森林公园常绿阔叶林群落物种多样性较丰富,部分物种对资源的需求有一定的相似性;物种总体呈显著正联结,种间关系较松散,各种对间的关联较弱,该群落逐渐趋于平衡阶段,各物种之间竞争较小。因此,在该地区可以选择有相似的生境需求和较宽生态位的优势乡土树种,调控生态位较小的树种,以期实现该地区群落正向演替。
Niche and interspecific connectivity of dominant species of woody plants in evergreen broad-leaved forest of Jinzifeng National Forest Park
-
摘要:
目的 探究浙江巾子峰国家森林公园常绿阔叶林木本植物优势种的生态位和种间联结性,以期为亚热带常绿阔叶林的生态恢复和生物多样性保护提供参考。 方法 基于样地调查数据,从物种组成、生态位宽度、生态位重叠指数、生态位相似系数、方差比率法、卡方($ {\chi}^{2} $)检验、Pearson相关系数和Spearman秩相关系数等方面研究了21种重要值≥1%的木本植物优势种的生态位和种间联结性。 结果 ①浙江巾子峰国家森林公园常绿阔叶林群落中共有木本植物113种,隶属31科62属,主要有壳斗科Fagaceae、蔷薇科Rosaceae、樟科Lauraceae和杜鹃花科Ericaceae等。②木荷Schima superba、甜槠Castanopsis eyrei和黄山松Pinus taiwanensis的重要值与生态位宽度较大,在该群落中处于建群种,与其他木本植物优势种间具有较高的生态位重叠值和生态位相似性系数。21个木本植物优势种的生态位重叠值和生态位相似性系数集中在0.10~0.50。③浙江巾子峰国家森林公园常绿阔叶林木本植物优势种间总体呈显著正联结(P<0.05)。经$ {\chi}^{2} $检验,在210个种对中,有112个种对呈不显著正联结,49个种对呈不显著负联结,仅有2个种对呈极显著正联结(P<0.01) ,8个种对呈显著正联结(P<0.05),39个种对呈无联结。Spearman秩相关检验和Pearson相关检验结果基本一致,不显著联结的种对分别占81.4%和80.0%,种间联结性较弱,正负联结比系数大于1。④主要优势树种间的Pearson相关系数与生态位重叠值的回归分析呈极显著正联结(P<0.01)。 结论 浙江巾子峰国家森林公园常绿阔叶林群落各物种之间竞争逐渐平衡,处于演替中期,物种丰富。图3表5参34 Abstract:Objective This study explore the ecological niche and interspecific association of the dominant species of woody plants in evergreen broad-leaved forest in Jinzifeng National Forest Park, Zhejiang Province, in order to provide a reference for ecological restoration and biodiversity conservation of subtropical evergreen broad-leaved forest. Method Niche composition, niche width, niche overlap index, niche similarity coefficient, variance ratio method, $ {\chi}^{2} $ test, Pearson correlation coefficient, and Spearman rank correlation coefficient were used to study the niche and interspecific association of 21 dominant species of woody plants with important values ≥1%. Result (1) There were 113 species of woody plants belonging to 31 families and 62 genera in the evergreen broad-leaved forest community in Jinzifeng National Forest Park, including Fagaceae, Rosaceae, Lauraceae, and Ericaceae. (2) The importance values and niche widths of Schima superba, Castanopsis eyrei and Pinus taiwanensis were larger. They were established species in this community, and had higher niche overlap values and niche similarity coefficients with other dominant species of woody plants. The niche overlap values and niche similarity coefficients of 21 dominant species of woody plants were mainly concentrated in the range of 0.10−0.50. (3) The dominant species of evergreen broad-leaved forest woody plants in Jinzifeng National Forest Park showed a significant positive association (P<0.05). Among 210 species pairs, 112 species pairs showed no significant positive association, 49 species pairs showed no significant negative association, only 2 species pairs showed extremely significant positive association (P<0.01) , 8 species pairs showed significant positive association (P<0.05) , 39 species pairs showed no association. Spearman rank correlation test and Pearson correlation test showed no significant association. Species pairs accounted for 81.4% and 80.0%, respectively. The interspecific association was weak, with positive and negative association ratios greater than 1. (4) The regression analysis of Pearson correlation coefficient and niche overlap values of dominant tree species showed a extremely significant positive correlation (P<0.01). Conclusion The evergreen broad-leaved forest community in Jinzifeng National Forest Park is in the middle stage of succession, and the competition among species is gradually balanced. [Ch, 3 fig. 5 fig. 34 ref.] -
图 3 主要优势树种Pearson与生态位重叠值的回归分析
Figure 3 Regression analysis of Pearson rank correlation with niche overlap of dominant tree species
表 1 研究区常绿阔叶林群落样地基本情况
Table 1. Basic situation of survey sites for broadleaf evergreen forest community survey in the study area
样地号 类型 海拔/m 群落类型 样地面积/m2 坡位 密度/(株·hm−2) 平均胸径/cm 郁闭度 1 天然林 1 163 常绿阔叶林 14 000 上 3 698 7.20 75.8 2 天然林 1 163 常绿阔叶林 400 上 2 750 9.87 93.9 3 天然林 1 146 常绿阔叶林 400 中 4 775 8.39 94.5 4 天然林 1 136 常绿阔叶林 400 中 2 100 12.15 90.7 5 天然林 1 151 常绿阔叶林 400 中 4 150 6.25 88.2 6 天然林 1 121 常绿阔叶林 400 中 1 525 16.86 93.4 7 天然林 1 176 常绿阔叶林 400 上 2 300 11.82 94.0  下载: 导出CSV
下载: 导出CSV
表 2 优势植物重要值与生态位宽度
Table 2. Important value and niche breadth of dominant plant species
编号 种名 个体数 分布频度 重要值/% Levins生态位宽度(BL) Shannon生态位宽度(BS) sp1 木荷 Schima superba 984 41 14.30 25.91 3.42 sp2 甜槠 Castanopsis eyrei 682 41 12.85 28.20 3.49 sp3 黄山松 Pinus taiwanensis 245 40 12.23 29.67 3.52 sp4 锥栗 Castanea henryi 261 38 6.02 21.39 3.30 sp5 马银花 Rhododendron ovatum 665 38 5.47 19.43 3.23 sp6 麂角杜鹃 Rhododendron latoucheae 500 35 4.30 18.47 3.19 sp7 大萼黄瑞木Adinandra glischroloma var. macrosepala 284 33 2.83 4.85 2.44 sp8 江南山柳 Clethra delavayi 189 35 2.45 17.07 3.17 sp9 野漆树 Toxicodendron succedaneum 112 33 2.21 24.31 3.32 sp10 浙江新木姜子 Neolitsea chekiangensis 157 28 1.91 11.83 2.79 sp11 微毛柃 Eurya hebeclados 152 27 1.87 12.21 2.89 sp12 短柄枹栎 Quercus glandulifera 115 27 1.86 19.59 3.11 sp13 江南越橘 Vaccinium mandarinorum 135 29 1.75 16.06 3.04 sp14 小叶青冈 Cyclobalanopsis gracilis 90 23 1.62 10.83 2.69 sp15 黄背越橘 Vaccinium iteophyllum 100 27 1.53 19.38 3.11 sp16 树参 Dendropanax dentiger 97 23 1.48 10.04 2.64 sp17 云山青冈 Cyclobalanopsis sessilifolia 72 22 1.37 14.64 2.84 sp18 红楠 Machilus thunbergii 50 22 1.15 13.30 2.82 sp19 吴茱萸五加 Gamblea ciliata var. evodiifolia 47 22 1.09 17.96 2.98 sp20 薄叶山矾 Symplocos anomala 43 20 1.02 14.11 2.81 sp21 华中樱 Cerasus conradinae 38 21 1.02 15.70 2.89
下载: 导出CSV
表 3 优势植物总体关联性
Table 3. Overall associations of dominant plant species
方差比率(VR) 检验统计量(W) 卡方临界值(0.95, 41) 卡方临界值(0.05, 41) 检验结果 2.84 116.73 27.33 56.94 显著正联结
下载: 导出CSV
表 4 主要优势树种χ2统计量检验
Table 4. χ2 correlation test of dominant population of dominant tree species
编号 sp1 sp2 sp3 sp4 sp5 sp6 sp7 sp8 sp9 sp10 sp11 sp12 sp13 sp14 sp15 sp16 sp17 sp18 sp19 sp20 sp2 0.00 sp3 0.00 0.00 sp4 0.00 0.00 2.75 sp5 0.00 0.00 2.75 0.42 sp6 0.00 0.00 1.03 3.24 3.24 sp7 0.00 0.00 0.61 0.02 1.92 2.20 sp8 0.00 0.00 1.03 0.01 0.01 4.11 0.13 sp9 0.00 0.00 0.61 0.02 0.02 6.75 0.87 2.20 sp10 0.00 0.00 0.16 0.34 0.34 0.15 0.00 2.30 0.67 sp11 0.00 0.00 0.11 0.36 0.36 1.83 2.16 1.83 0.04 0.00 sp12 0.00 0.00 0.11 3.48 0.36 10.34 5.29 1.83 5.29 0.00 1.43 sp13 0.00 0.00 0.21 0.67 4.57 0.52 3.50 0.06 0.02 0.26 0.19 3.02 sp14 0.00 0.00 0.02 0.05 0.05 0.59 0.00 0.59 0.00 0.02 0.81 0.06 1.50 sp15 0.00 0.00 0.11 0.36 3.48 1.83 2.16 1.83 0.41 2.13 1.43 3.57 6.07 0.06 sp16 0.00 0.00 0.02 0.97 0.05 0.01 0.00 0.01 0.62 0.02 0.06 0.81 0.73 0.79 0.06 sp17 0.00 0.00 0.01 0.02 1.15 1.29 0.03 0.06 0.03 0.10 0.00 0.43 1.78 1.35 0.00 0.53 sp18 0.00 0.00 0.01 0.02 0.02 0.41 0.39 0.06 0.39 0.10 0.00 3.96 1.78 0.28 1.77 0.53 0.04 sp19 0.00 0.00 0.01 0.02 0.02 0.41 2.01 0.06 2.01 0.10 1.77 3.96 0.42 0.01 3.96 1.86 2.10 0.19 sp20 0.00 0.00 0.00 0.00 1.34 1.59 0.10 0.14 0.22 0.01 0.77 0.05 0.86 0.03 0.05 1.17 0.02 0.02 0.02 sp21 0.00 0.00 0.00 0.00 0.00 0.26 0.22 0.26 0.22 0.61 0.20 0.05 1.28 0.21 0.05 0.21 0.60 0.02 0.02 0.03 说明:sp1. 木荷;sp2. 甜槠;sp3. 黄山松;sp4. 锥栗;sp5. 马银花;sp6. 麂角杜鹃;sp7. 大萼黄瑞木;sp8. 江南山柳;sp9. 野漆树;sp10. 浙江新木姜子;sp11. 微毛柃;sp12. 短柄枹栎;sp13. 江南越橘;sp14. 小叶青冈;sp15. 黄背越橘;sp16. 树参;sp17. 云山青冈;sp18. 红楠;sp19. 吴茱萸五加;sp20. 薄叶山矾;sp21. 华中樱。
下载: 导出CSV
表 5 主要优势树种的Spearman秩相关系数、Pearson相关系数检验结果比较
Table 5. Comparison of the results of Spearman rank correlation and Pearson rank correlation test of dominant tree species
检验方法 正联结(相关)/对 负联结(相关)/对 无联结/对 极显著 显著 不显著 总数 极显著 显著 不显著 总数 总数 χ2检验 2 8 112 122 0 0 49 49 39 Spearman秩相关系数 21 14 108 142 1 4 63 68 0 Pearson相关系数 25 14 89 128 1 2 79 82 0 说明:极显著P<0.01;显著P<0.05;不显著P>0.05。
下载: 导出CSV
-
[1] GU Li, GONG Zhiwen, LI Weizhong. Niches and interspecific associations of dominant populations in three changed stages of natural secondary forests on Loess Plateau, P. R. China [J/OL]. Scientific Reports, 2017, 7 (1): 6604[2024-04-10]. DOI: 10.1038/s41598-017-06689-9. [2] KIRCHHEIMER B, WESSELY J, GATTRINGER A, et al. Reconstructing geographical parthenogenesis: effects of niche differentiation and reproductive mode on holocene range expansion of an alpine plant [J]. Ecology Letters, 2018, 21(3): 392−401. [3] 刘润红, 陈乐, 涂洪润, 等. 桂林岩溶石山青冈群落灌木层主要物种生态位与种间联结[J]. 生态学报, 2020, 40(6): 2057−2071. LIU Runhong, CHEN Le, TU Hongrun, et al. Niche and interspecific association of main species in shrub layer of Cyclobalanopsis glauca community in karst hills of Guilin, Southwest China [J]. Acta Ecologica Sinica, 2020, 40(6): 2057−2071. [4] 张盟, 郑俊鸣, 万佳艺, 等. 福建省东门屿优势木本植物的生态位与种间联结[J]. 森林与环境学报, 2022, 42(1): 11−19. ZHANG Meng, ZHENG Junming, WAN Jiayi, et al. Niche and interspecific associations of dominant woody plants in Dongmen Island, Fujian Province [J]. Journal of Forest and Environment, 2022, 42(1): 11−19. [5] ALVAREZ-YÉPIZ J C, BÚRQUEZ A, DOVCIAK M. Ontogenetic shifts in plant-plant interactions in a rare cycad within angiosperm communities [J]. Oecologia, 2014, 175(2): 725−735. [6] 方庆, 谭菊荣, 许惠春, 等. 珍稀濒危植物细果秤锤树群落物种组成与生态位分析[J]. 浙江农林大学学报, 2022, 39(5): 931−939. FANG Qing, TAN Jurong, XU Huichun, et al. Species composition and niche of Sinojackia microcarpa, a rare and endangered plant [J]. Journal of Zhejiang A&F University, 2022, 39(5): 931−939. [7] 丁茂, 汪宇坤, 何煜然, 等. 安徽鹞落坪国家级自然保护区落叶阔叶林树种多样性、种间联结及群落稳定性变化[J]. 生态学报, 2023, 43(7): 2818−2830. DING Mao, WANG Yukun, HE Yuran, et al. Changes of species diversity, interspecific associations and community stability for the deciduous broad-leaved forest in Yaoluoping National Nature Reserve, Anhui Province [J]. Acta Ecologica Sinica, 2023, 43(7): 2818−2830. [8] 杨春玉, 刘绍飞, 喻理飞. 喀斯特森林恢复过程中优势乔木树种种间联结性分析[J]. 浙江林学院学报, 2010, 27(1): 44−50. YANG Chunyu, LIU Shaofei, YU Lifei. Interspecific associations of dominant tree species with restoration of a karst forest [J]. Journal of Zhejiang Forestry College, 2010, 27(1): 44−50. [9] 刘啸林, 吴友贵, 张敏华, 等. 浙江百山祖25 ha亚热带森林动态监测样地群落组成与结构特征[J]. 生物多样性, 2024, 32(2): 21−31. LIU Xiaolin, WU Yougui, ZHANG Minhua, et al. Community composition and structure of a 25-ha forest dynamics plot of subtropical forest in Baishanzu, Zhejiang Province [J]. Biodiversity Science, 2024, 32(2): 21−31. [10] 陈小荣, 李乐, 夏家天, 等. 百山祖亮叶水青冈种群结构和分布格局[J]. 浙江农林大学学报, 2012, 29(5): 647−654. CHEN Xiaorong, LI Le, XIA Jiatian, et al. Population structure and distribution of Fagus lucida in the Baishanzu forest [J]. Journal of Zhejiang A&F University, 2012, 29(5): 647−654. [11] 徐敏, 骆争荣, 于明坚, 等. 百山祖北坡中山常绿阔叶林的物种组成和群落结构[J]. 浙江大学学报(农业与生命科学版), 2007, 33(4): 450−457. XU Min, LUO Zhengrong, YU Mingjian, et al. Floristic composition and community structure of mid-montane evergreen broad-leaved forest in north slope of Baishanzu Mountain [J]. Journal of Zhejiang University (Agriculture & Life Sciences), 2007, 33(4): 450−457. [12] 蔡焕满, 吴素美, 吴逸卿, 等. 百山祖国家公园五岭坑常绿阔叶林甜槠的种群特征[J]. 浙江林业科技, 2024, 44(1): 1−7. CAI Huanman, WU Sumei, WU Yiqing, et al. The population characteristics of Castanopsis eyrei in evergreen broad-leaved forest in Wulingkeng of Baishanzu National Park [J]. Journal of Zhejiang Forestry Science and Technology, 2024, 44(1): 1−7. [13] 李伟成, 郑彦超, 盛海燕, 等. 浙江庆元巾子峰国家森林公园植被群落的数量分类与排序[J]. 浙江农林大学学报, 2021, 38(3): 523−533. LI Weicheng, ZHENG Yanchao, SHENG Haiyan, et al. Numerical classification and ordination of vegetation communities in Jinzifeng National Forest Park, Qingyuan, Zhejiang [J]. Journal of Zhejiang A&F University, 2021, 38(3): 523−533. [14] CONDIT R. Tropical Forest Census Plots: Methods and Results from Barro Colorado Island, Panama and a Comparison with Other Plots [M]. Berlin, Heidelberg: Springer Science & Business Media, 1998. [15] 马克平, 刘玉明. 生物群落多样性的测度方法: Ⅰα多样性的测度方法(下)[J]. 生物多样性, 1994, 2(4): 231−239. MA Keping, LIU Yuming. Measuring methods of biological community diversity: Ⅰα diversity measurement method (Part 2) [J]. Chinese Biodiversity, 1994, 2(4): 231−239. [16] 王刚, 赵松岭, 张鹏云, 等. 关于生态位定义的探讨及生态位重叠计测公式改进的研究[J]. 生态学报, 1984, 4(2): 119−127. WANG Gang, ZHAO Songlin, ZHANG Pengyun. Discussion on the definition of niche and study on the improvement of the measurement formula of niche overlap [J]. Acta Ecologica Sinica, 1984, 4(2): 119−127. [17] 郭平平, 税伟, 江聪, 等. 退化天坑倒石坡林下优势物种生态位特征[J]. 应用生态学报, 2019, 30(11): 3635−3645. GUO Pingping, SHUI Wei, JIANG Cong, et al. Niche characteristics of understory dominant species of talus slope in degraded Tiankeng [J]. Chinese Journal of Applied Ecology, 2019, 30(11): 3635−3645. [18] SCHOENER T W. Resource partitioning in ecological communities: research on how similar species divide resources helps reveal the natural regulation of species diversity [J]. Science, 1974, 185(4145): 27−39. [19] SCHLUTER D. A variance test for detecting species associations, with some example applications [J]. Ecology, 1984, 65(3): 998−1005. [20] 江常春, 梁希妮, 田磊, 等. 浙江省常绿阔叶林优势树种种间联结分析[J]. 浙江师范大学学报(自然科学版), 2022, 45(4): 437−445. JIANG Changchun, LIANG Xini, TIAN Lei, et al. Analysis on interspecific association of dominant species in evergreen broad-leaved forests in Zhejiang Province [J]. Journal of Zhejiang Normal University (Natural Sciences), 2022, 45(4): 437−445. [21] 吕增伟, 蒋天雨, 徐绣琴, 等. 六盘山白桦群落乔木层主要物种生态位与种间联结[J]. 福建农林大学学报(自然科学版), 2023, 52(5): 660−668. LÜ Zengwei, JIANG Tianyu, XU Xiuqin, et al. Niche and interspecific association of main species in arbor layer of Betula platyphylla community in Liupan Mountain, Ningxia [J]. Journal of Fujian Agriculture and Forestry University (Natural Science Edition), 2023, 52(5): 660−668. [22] VILLANUEVA R A M, CHEN Z J. Ggplot2: elegant graphics for data analysis [J]. Measurement: Interdisciplinary Research and Perspectives, 2019, 17(3): 160−167. [23] 陈林, 辛佳宁, 苏莹, 等. 异质生境对荒漠草原植物群落组成和种群生态位的影响[J]. 生态学报, 2019, 39(17): 6187−6205. CHEN Lin, XIN Jianing, SU Ying, et al. Effects of heterogeneous habitats on community composition and niche characteristics of different plant populations in the desert steppe of China [J]. Acta Ecologica Sinica, 2019, 39(17): 6187−6205. [24] 叶兴状, 王妙青, 程诺, 等. 福建天台山半枫荷天然群落的物种组成、生态位和种间关系[J]. 植物资源与环境学报, 2021, 30(6): 19−28. YE Xingzhuang, WANG Miaoqing, CHENG Nuo, et al. Species composition, niche, and interspecific relationships of Semiliquidambar cathayensis natural community in Tiantai Mountain of Fujian Province [J]. Journal of Plant Resources and Environment, 2021, 30(6): 19−28. [25] 刘雨婷, 侯满福, 贺露炎, 等. 滇东菌子山喀斯特森林群落乔木优势树种生态位和种间联结[J]. 应用生态学报, 2023, 34(7): 1771−1778. LIU Yuting, HOU Manfu, HE Luyan, et al. Niche and interspecific association of dominant tree species in karst forest of Junzi Mountain, Eastern Yunnan, China [J]. Chinese Journal of Applied Ecology, 2023, 34(7): 1771−1778. [26] 何江, 徐来仙, 艾训儒. 湖北七姊妹山亚热带常绿落叶阔叶混交林主要木本植物的生态位与种间联结[J]. 林业科学研究, 2023, 36(3): 138−148. HE Jiang, XU Laixian, AI Xunru. Niche and interspecific association of main woody plants in the subtropical evergreen-deciduous broadleaved mixed forest in Qizimei Mountain, Hubei Province [J]. Forest Research, 2023, 36(3): 138−148. [27] 张东梅, 赵文智, 罗维成. 荒漠草原带盐碱地优势植物生态位与种间联结[J]. 生态学杂志, 2018, 37(5): 1307−1315. ZHANG Dongmei, ZHAO Wenzhi, LUO Weicheng. Niche and interspecific association of dominant plant species in saline-alkaline soils of desert steppe zone [J]. Chinese Journal of Ecology, 2018, 37(5): 1307−1315. [28] 吴友贵, 叶珍林, 周荣飞, 等. 百山祖常绿阔叶林优势种群的生态位[J]. 广西植物, 2016, 36(2): 186−192. WU Yougui, YE Zhenlin, ZHOU Rongfei, et al. Niche of dominant species populations in an evergreen broad-leaved forest in Baishanzu [J]. Guihaia, 2016, 36(2): 186−192. [29] 栾兆平, 郭滨德, 吴金卓. 长白山地区黄檗阔叶混交林主要乔木种群生态位特征[J]. 森林工程, 2023, 39(4): 78−85, 92. LUAN Zhaoping, GUO Binde, WU Jinzhuo. Niche characteristics of main tree populations in broad-leaved mixed forest of Phellodendron amurense in Changbai mountain area [J]. Forest Engineering, 2023, 39(4): 78−85, 92. [30] 涂洪润, 农娟丽, 朱军, 等. 桂林岩溶石山密花树群落主要物种的种间关联及群落稳定性[J]. 生态学报, 2022, 42(9): 3688−3705. TU Hongrun, NONG Juanli, ZHU Jun, et al. Interspecific associations of main species and community stability of Myrsine seguinii community in karst hills of Guilin, southwestern China [J]. Acta Ecologica Sinica, 2022, 42(9): 3688−3705. [31] 张滋芳, 毕润成, 张钦弟, 等. 珍稀濒危植物矮牡丹生存群落优势种种间联结性及群落稳定性[J]. 应用与环境生物学报, 2019, 25(2): 291−299. ZHANG Zifang, BI Runcheng, ZHANG Qindi, et al. Community stability and inter-specific associations between the rare plant Paeonia suffruticosa Andr. var. spontanea Rehd and dominant species [J]. Chinese Journal of Applied & Environmental Biology, 2019, 25(2): 291−299. [32] 罗敏贤, 林碧华, 陈绪辉, 等. 福建龟山伞花木所在群落乔灌层优势种的种间联结性[J]. 植物资源与环境学报, 2022, 31(6): 63−72. LUO Minxian, LIN Bihua, CHEN Xuhui, et al. Interspecific associations of dominant species in arbor and shrub layers of Eurycorymbus cavaleriei located community in Guishan Mountain of Fujian Province [J]. Journal of Plant Resources and Environment, 2022, 31(6): 63−72. [33] 沈雪梨, 吴远密, 朱艺璇, 等. 濒危植物百花山葡萄群落物种生态位特征与种间联结研究[J]. 植物科学学报, 2020, 38(2): 195−204. SHEN Xueli, WU Yuanmi, ZHU Yixuan, et al. Community niche characteristics and interspecific associations of critically endangered species, Vitis baihuashanensis M. S. Kang et D. Z. Lu [J]. Plant Science Journal, 2020, 38(2): 195−204. [34] 郑俊鸣, 李敏, 张盟, 等. 滨海防护林木麻黄和花吊丝竹混交林群落的种间关联和生态位分析[J]. 热带亚热带植物学报, 2021, 29(5): 465−473. ZHENG Junming, LI Min, ZHANG Meng, et al. Interspecific association and niche of mixed forest communities of Casuarina equisetifolia and Dendrocalamus minor var. amoenus in the Windbreak of sandy coast [J]. Journal of Tropical and Subtropical Botany, 2021, 29(5): 465−473. -

-
链接本文:
https://zlxb.zafu.edu.cn/article/doi/10.11833/j.issn.2095-0756.20240307
 点击查看大图
点击查看大图
计量
- 文章访问数: 1786
- HTML全文浏览量: 582
- PDF下载量: 44
- 被引次数: 0
