





-
星天牛Anoplophora chinensis属鞘翅目Coleptera天牛科Cerambycidae,是在中国分布较广,危害树木种类较多的一种蛀干性害虫,既蛀食大树,也取食幼树,切断树木输送养分和水分的通道,导致树木衰弱或死亡[1]。天牛对树木的危害选择性很强,不同树种对天牛危害或忌避或抗或被喜食[2-4]。在对星天牛行为和习性的观察研究工作中,发现其成虫特别喜欢取食苦楝树皮作为补充营养,而且成虫寿命一般为40~50 d[5]。苦楝Melia azedarach为楝科Meliaceae落叶乔木,抗病虫能力强,其提取物具有广谱性的杀虫作用[6-14]。根据这一特性,笔者在林间对苦楝引诱星天牛成虫的效果进行了较为详细的试验,并且在此基础上,设置室内多树种混合引诱实验,通过将不同树种枝条和成虫放置在特定同一的环境下,排除自然环境以及人为因素的干扰,观察星天牛成虫的取食反应,进一步验证实验结果的准确性和可靠性。实验结果以期为星天牛防治提供新的技术手段。
-
试验地位于浙江省慈溪市沿海防护林,属北亚热带季风气候,四季分明,主栽树种包括杨树Populus sp.,刺槐Robinia pseudoacacia,黄山栾树Koelreuteria integrifoliola,水杉Metasequoia glyptostroboides,柳树Salix sp.,黑松Pinus thunbergii,木麻黄Casuarina equisetifolia,桉树Eucalyptus sp.,红树Mangrove vegetation等,其中木麻黄和黄山栾树等星天牛危害较为严重。
-
林区内选择4块样地,样地基本情况见表 1,分别在林缘和林内栽植2年生苦楝,苦楝为2年生,平均胸径在3~5 cm,平均树高为1~3 m。1号样地和2号样地分别在林缘与林间栽植50株苦楝,3号林间栽植100株苦楝,4号林间栽植50株苦楝,株间距和行间距分别为50 cm和80 cm。
表 1 样地基本情况
Table 1. Basic situation of sampling sites
样地编号 样地基本情况 1 林缘,由西向东依次为苦棟 Melia azedarach,龙柏 Juniperus chinensis ‘Kaizuka',黄山栾树,木麻黄,夹竹桃 Phlox drummondi 2 林间,由西向东依次为龙柏,黄山栾树,苦楝,木麻黄,夹竹 3 林间,由西向东依次为海滨木槿 AHibiscus hamabo,夹竹桃,苦楝,黄山栾树,木麻黄 4 林间,由西向东依次为海滨木槿,黄山栾树,木麻黄,苦楝,龙柏,夹竹桃 -
在星天牛羽化高峰期(6月),连续3 d对4块样地调查并捕杀成虫。调查时间为每天6:30-17:30隔1 h观察1次,详细记录星天牛的取食、交配、停息等活动情况。
-
在慈溪沿海防护林选择产卵孔数量较多的柳树枝条,人工剖开共采集星天牛幼虫300余头,分别装入10 mL离心管中,并放入少量柳树木屑作为星天牛幼虫的饲料,带回实验室饲养。
-
由于星天牛羽化之后存活时间较短,为了保证有足够的供试虫体以确保整体实验的顺利进行,需将带回的星天牛幼虫放置于不同条件下饲养,使成虫羽化时间不同。其中:200头置于温度28 ℃,光照100%,湿度45%,时间持续16 h和温度20 ℃,光照0%,湿度50%,时间持续8 h的条件下提前羽化状态;另100头置于温度25 ℃,光照100%,湿度50%,时间持续14 h和温度18 ℃,光照0%,湿度50%,时间持续10 h的条件下正常羽化状态。
-
用白色记号笔分别在雌虫腹部和雄虫背部标注不同数字,以此标记同期羽化的星天牛成虫5对,雌雄大小一致,无畸形,羽化出孔后饲养7 d,选择苦楝、木麻黄、黄山栾树的枝条,规格为长25 cm直径2 cm的嫩枝,分别放置于玻璃瓶内水培,连同5对经过24 h饥饿处理过的星天牛雌雄成虫放入100 cm×70 cm×50 cm养虫笼内,该养虫笼的12条边是木质材料,6个面用铁网封住。养虫笼置于阴凉处,隔48 h更换1次新鲜枝条,观察成虫取食选择,测量取食面积,每次更换枝条后测量1次,共重复测定4次。
试验数据分析采用的是SAS 软件,数据的SAS分析调用的是方差分析(ANOVA)过程和广义线性模型(GLM)过程。
-
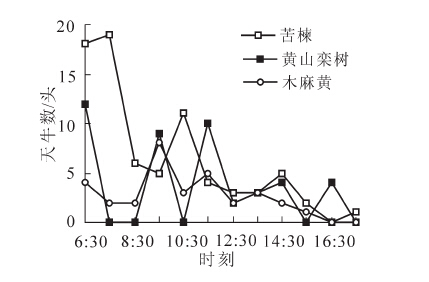
星天牛成虫多在白天活动,夜晚静息。苦楝作为引诱树种在不同时间诱集到的星天牛成虫数见图 1,共捕获77头,引诱率达到50.33%。不同时间不同树种捕捉到的星天牛成虫数量见图 2,共捕获星天牛成虫153头,其中雌虫102头,雄虫51头,正在交尾的8对。由数据统计可看出:星天牛集中活动的时间为6:30-11:30,星天牛选择晴朗无风的天气进行交尾,时间集中在11:00-16:00。

图 1 不同时间苦楝诱虫结果
Figure 1. Attracting insects results of Melia azedarach in different times

图 2 不同时间各树种捕捉成虫结果
Figure 2. Capture adult results of different tree species of different times different times
-
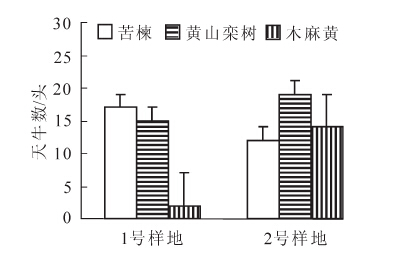
1号样地和2号样地分别是在林缘与林间种植50株苦楝,诱集到的星天牛成虫结果见图 3,样地各树种捕捉到的星天牛成虫结果见图 4。可知,1号样地诱集到的成虫数量是2号样地的1倍以上,1号诱虫率为50.00%,2号诱虫率仅为26.67%。

图 3 不同位置苦楝诱虫结果
Figure 3. Attracting insects results of Melia azedarach at different locations

图 4 不同位置各树种捕捉成虫结果
Figure 4. Capture adult results of different tree species at different locations
-
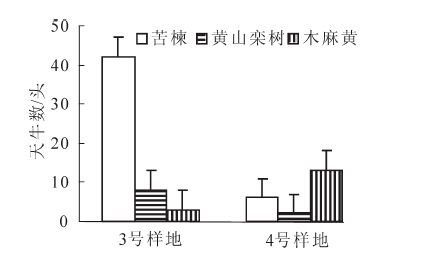
3号样地种植苦楝的数量是4号样地的2倍,诱集到的星天牛成虫结果见图 5。3号样地苦楝诱集成虫数量是4号样地的7倍,3号样地苦楝的诱虫率为79.25%,而4号样地诱虫率仅为28.57%。由此可见:苦楝对星天牛的诱集效果与苦楝数量有直接关系,建议种植适当数量的苦楝作为隔离带对星天牛进行集中诱杀。调查发现,无论是黄山栾树还是木麻黄,离苦楝树距离较近的树种星天牛数量也相对较多,分析可能是星天牛在苦楝上取食之后飞到其他树上继续为害。建议在设置诱饵树时,应考虑风向并且在一定距离内应尽可能离受保护树种林分远一点,以免成虫在苦楝上补充营养后因未及时杀灭使其再飞到其他树种上产卵危害。

图 5 不同数量苦楝诱虫结果
Figure 5. Attracting insects results of different trees of Melia azedarach
-
见表 2。通过4次的取食面积测量,可以发现: 星天牛成虫对苦楝枝条的取食量最大,黄山栾树次之,木麻黄最少。苦楝枝条的平均被取食面积是6.19 cm2。经方差分析,得出各树枝星天牛的取食面积有显著性差异(F=14.92,P=0.001 4)。
表 2 星天牛对各树种枝条的取食面积比较
Table 2. Feeding areas on the branches
样枝编号 取食面积/cm2 黄山栾树 木麻黄 苦楝 1 3.12 3.16 5.27 2 4.11 3.82 6.22 3 3.94 3.08 6.71 4 4.83 4.72 6.54 -
在野外实验过程中发现,9:00之前苦楝的引诱效果较为明显,10:00之后天牛则一般选择生长较为茂盛的黄山栾树和木麻黄进行取食,分析原因可能与树木的生长状况和气温有关。苦楝为2年生植株,平均胸径只有3~5 cm,树冠顶端茂盛程度远不及树龄较大的黄山栾树和木麻黄。9:00之前当地气温基本在24 ℃左右,微风,星天牛选择喜食树种而食,这时苗木的茂盛程度成为次要因素;10:00之后当地气温上升至32 ℃,阳光照射强烈,星天牛则选择了茂盛的黄山栾树和木麻黄进行取食,在调查过程中发现,这时的星天牛一般隐藏在树叶下以躲避阳光,较不易被发现。
苦楝对星天牛成虫具有较强的引诱作用,最好栽植在林缘、林间未郁闭的宽阔道路和虫源树旁,数量较多的苦楝引诱效果则更为明显,但苦楝作为星天牛喜食树种不宜过量种植,如未及时灭杀成虫易造成星天牛种群数量的增加,因此要合理规划造林,同时在星天牛羽化时及时对其成虫进行灭杀。
成虫在苦楝树上时,一般较频繁地变换取食部位,有研究表明星天牛成虫在苦楝树上取食、停歇及交配时间一般超过2 h[15],这使得有足够的时间施药灭杀成虫,星天牛成虫羽化后飞至苦楝树上补充营养,建议在苦楝上喷施农药,使其爬触或取食时死亡。
室内多树种混合引诱试验得到的数据经方差分析得出的结果说明: 星天牛成虫对于不同树种的取食具有较为明显的选择偏好性,苦楝被取食量明显大于黄山栾树和木麻黄的被取食量。由标准差分析得到的数据说明:星天牛成虫分别对这3个不同树种枝条的取食量均处于平稳水平,波动幅度不大。这说明星天牛成虫在不同的时间段对这3个不同树种枝条各自的取食量没有明显的差异性。
通过进行室内多树种混合引诱实验发现在特定同一的环境下,苦楝的被取食量远大于黄山栾树和木麻黄,进一步确定了苦楝对星天牛成虫具有较强的引诱作用,这与野外调查得到的结果是一致的。通过野外的调查和室内的实验说明:苦楝对星天牛的引诱效果十分明显,能够引诱到绝大多数的星天牛成虫,并且成虫主要是以取食的形式对苦楝树皮造成破坏,在苦楝植株上极少发现有星天牛产卵行为,这为集中喷药防治星天牛成虫提供理论依据,并且这一结果对于沿海防护林栽植苦楝作为引诱树种具有十分重要的意义。
星天牛飞到苦楝树上取食,应当是苦楝树上某些挥发性的化学物质使其感到食物的存在,能够引起星天牛远距离取食的苦楝挥发性化学物质到底是什么,是否能够从苦楝树中分析出这些化学物质,再人工合成对星天牛进行诱杀,这些问题有待进一步研究解决。
Attractiveness of Melia azedarach to Anoplophora chinensis
-
摘要: 通过设置室内多树种混合引诱实验和林间引诱实验对苦楝Melia azedarach引诱星天牛Anoplophora chinensis成虫的效果进行了详细比较。结果表明:苦楝对星天牛成虫具有较强的引诱作用,并且数量较多的苦楝引诱效果更为明显,100株苦楝的诱虫率为79.25%,而50株苦楝诱虫率仅为28.57%。在同一的环境下,苦楝枝条的平均被取食面积为6.19 cm2,远大于黄山栾树Koelreuteria integrifoliola和木麻黄Casuarina equiselifolia。实验结果以期为星天牛防治提供新的理论基础和技术手段。Abstract: We studied the attractiveness of Melia azedarach to Anoplophora chinensis by both bioassay in the laboratory and field trapping.The results indicated that Melia azedarach had strong attractiveness to Anoplophora chinensis.The increase in the number of Melia azedarach would result in increased attractiveness.The trapping rate of 100 Melia azedarach trees was 79.25 per cent while that of 50 Melia azedarach trees was only 28.57 per cent.In the same environment, the average feeding area of Melia azedarach branch was 6.19 cm2, larger than that of Koelreuteria integrifoliola and Casuarina equisetifolia.The research findings would provide new theoretical foundation and technique for the control and prevention of Anoplophora chinensis.
-
图 1 不同时间苦楝诱虫结果
Figure 1 Attracting insects results of Melia azedarach in different times
图 2 不同时间各树种捕捉成虫结果
Figure 2 Capture adult results of different tree species of different times different times
图 3 不同位置苦楝诱虫结果
Figure 3 Attracting insects results of Melia azedarach at different locations
图 4 不同位置各树种捕捉成虫结果
Figure 4 Capture adult results of different tree species at different locations
图 5 不同数量苦楝诱虫结果
Figure 5 Attracting insects results of different trees of Melia azedarach
表 1 样地基本情况
Table 1. Basic situation of sampling sites
样地编号 样地基本情况 1 林缘,由西向东依次为苦棟 Melia azedarach,龙柏 Juniperus chinensis ‘Kaizuka',黄山栾树,木麻黄,夹竹桃 Phlox drummondi 2 林间,由西向东依次为龙柏,黄山栾树,苦楝,木麻黄,夹竹 3 林间,由西向东依次为海滨木槿 AHibiscus hamabo,夹竹桃,苦楝,黄山栾树,木麻黄 4 林间,由西向东依次为海滨木槿,黄山栾树,木麻黄,苦楝,龙柏,夹竹桃  下载: 导出CSV
下载: 导出CSV
表 2 星天牛对各树种枝条的取食面积比较
Table 2. Feeding areas on the branches
样枝编号 取食面积/cm2 黄山栾树 木麻黄 苦楝 1 3.12 3.16 5.27 2 4.11 3.82 6.22 3 3.94 3.08 6.71 4 4.83 4.72 6.54
下载: 导出CSV
-
[1] 魏建荣, 赵文霞, 张永安.星天牛研究进展[J].植物检疫, 2011,25(5):81-85. WEI Jianrong,ZHAO Wenxia,ZHANG Yong'an.Research progress on Anoplophora chinensis[J].Plant Quar,2011,25(5):81-85. [2] 钦俊德.昆虫与植物的关系[M].北京:科学出版社,1987. [3] 张凌健.星天牛喜食银糖槭[J].植物保护,1988,14(3):54. ZHANG Lingjian.Silver Anoplophora eating sugar maple[J].Plant Prot,1988,14(3):54. [4] 赵一宇, 李金宁, 徐德昌,等.几种忌避树种的提取物对天牛驱避作用的初步研究[J].宁夏农学院学报,1992,13(4):19-24. ZHAO Yiyu,LI Jinning,XU Dechang,et al.Anthelmintic effect of the extractions from several tree species on capricorn beetle[J].J Ningxia Agric Coll,1992,13(4):19-24. [5] 黄金水,何学友,高美玲,等.利用苦楝树诱杀木麻黄星天牛成虫的可行性研究:木麻黄防护林主要虫害控制技术研究之一[J].防护林科技,1998(2):9-11. HUANG Jinshui,HE Xueyou,GAO Meiling,et al.The use of booby traps China berry tree Casuarina feasibility study of adult Anoplophora[J].Prot For Sci Technol, 1998(2):9-11. [6] 黄金水,何学友,高美玲,等.苦谏树林间引诱星天牛试验研究[J].防护林科技,2000(专刊):7-13. Huang Jinshui,HE Xueyou,GAO Meiling,et al.Experiment of alluring Anoplophora chinensis (forster) with Mella azedarach Linn.in plantations[J].Prot For Sci Technol,2000(supp):7-13.. [7] 赵善欢.几种楝科植物种核油对稻褐虱的拒食作用试验[J].昆虫学报,1983,26(1):1-9. ZHAO Shanhuan.Experiments on the use of seed oils of some meliaceous plants as antifeedants in brown planthopper control[J].Acta Entomol Sin,1983,26(1):1-9. [8] 赵善欢,黄炳球,胡美英.楝科植物油及种核抽提物对稻瘿蚊产卵忌避作用的防治试验[J].昆虫学报,1986,29(2):221-224. ZHAO Shanhuan,HUANG Bingqiu,HU Meiying.The oviposition deterring effects of some Meliaceous plant extracts against the rice gall midge orseol/a oryzaewood-mason[J].Acta Entomol Sin,1986,29(2):221-224. [9] 赵博光.6种植物提取物对马尾松毛虫的抑食作用及毒性[J].南京林业大学学报,1994,18(2):73-78. ZHAO Boguang.Feeding inhibition and toxicity of the extracts from six species of plants on caterpillars of Dendrolimus puctatus walker[J].J Nanjing For Univ,1994,18(2):73-78. [10] 汪文陆,赵善欢,韩玖,等.苦楝中几种杀虫有效成分对菜青虫和亚洲玉米螟的生物活性[J].植物保护学报,1992,19(4):395-364. WANG Wenlu,ZHAO Shanhuan,HAN Jiu,et al.Effects of several insecticidal principles from China berry,Melia azedarach on the imported cabbage worm,Pieris rapae and the asiatic corn borer,Ostrinia furnacalis[J].J Plant Prot,1992,19(4):395-364. [11] 韩玖,林文翰,徐任生,等.苦楝化学成分的研究[J].药学学报,1991,26(6):426-429. HAN Jiu,LIN Wenhan,XU Rensheng,et al.Studies on the chemical constitutions of Melia azedarach L.[J].Acta Pharm Sin,1991,26(6):426-429. [12] 唐英,谭世语,张进.楝属植物中药用成分的研究开发[J].化工进展,2002,21(5):334-3371. TANG Ying,TAN Shiyu,ZHANG Jin.Situation of research and development of Melia plant as Chinese medicine[J].Chem Ind Eng Prog,2002,21(5):334-3371. [13] 朱虹,单淑芳,李增智,等.苦楝内生真菌及其代谢产物的杀虫活性[J].中国生物防治,2010,26(1):47-52. ZHU Hong,SHAN Shufang,LI Zengzhi,et al.Insecticidal activity of endophytic fungi from Melia azedarach L.[J].Chin J Biol Cont,2010,26(1):47-52. [14] OELRICHS P B,HILL M W,VALLELY P J,et al.Toxic tetranortriterpenes of the fruits of Melia azedarach[J].Phytochemistry,1983,22(2):531-534. [15] 何学友,黄金水,叶剑雄,等.星天牛行为及控制技术研究(Ⅱ)星天牛控制技术的研究[J].福建林业科技,2003,30(2):2-13. HE Xueyou,HUANG Jinshui,YE Jianxiong,et al.Studies on the Anoplophora chinensis behavior and control technique(Ⅱ) study on the A.chinensis control technique[J].J Fujian For Sci Technol,2003,30(2):2-13. -

-
链接本文:
https://zlxb.zafu.edu.cn/article/doi/10.11833/j.issn.2095-0756.2014.03.017
 点击查看大图
点击查看大图

计量
- 文章访问数: 4221
- HTML全文浏览量: 797
- PDF下载量: 629
- 被引次数: 0
