




-
蜡梅属Chimonanthus是中国的特有属,公认的蜡梅属植物有4个物种,即蜡梅C. praecox、山蜡梅C. nitens、柳叶蜡梅C. salicifolius和西南蜡梅C. campanulatus[1−2]。蜡梅广泛分布于中国东南部和中部的多个省份[3−4];山蜡梅仅在长江以南有较窄的分布[5];柳叶蜡梅主要生长在江西、安徽南部和浙江[6]。蜡梅属植物具有镇咳、抗炎、解热、抗菌和降压作用,花可用于提取精油,根、茎和叶可入药[7−9],其干燥的叶被用于治疗感冒和流感[10−13]。其中,柳叶蜡梅和山蜡梅茎叶中的次生代谢产物,如黄酮类、香豆素类、生物碱类和甾体类被认为是药理作用的有效成分[14−15]。
植物种类不同,其花香物质的种类与含量也不同[16]。蜡梅花香的主要物质为萜类化合物(单萜和倍半萜)和苯环类化合物[17−18],山蜡梅和柳叶蜡梅干花的主要花香物质为α-月桂烯、桉叶素和四甲基环癸二烯甲醇[19]。蜡梅在一天的不同时段内释放的花香物质,即日释放节律存在波动。盛开的磬口蜡梅C. praecox var. grandiflorus在13:00释放的花香物质种类多于乔种蜡梅C. praecox var. intermedius[20]。在不同昼夜节律下,磬口蜡梅释放的主要花香物质种类接近,但其数量在12:00最多[21]。
由于蜡梅属植物不同物种以及不同时间段内的花香物质存在差异,有必要对其进行系统的比较分析,以揭示不同物种间以及不同释放节律下花香物质的异同。本研究采用顶空-固相微萃取(HS-SPME)结合气相色谱-质谱联用技术(GC-MS)对3种蜡梅属植物(蜡梅、柳叶蜡梅和山蜡梅)的花香物质进行鉴定和分析,旨在比较3种蜡梅属植物的花香物质及其白天的释放节律。
-
使用3种蜡梅属植物:蜡梅、柳叶蜡梅和山蜡梅(图1),均栽培于浙江农林大学校园内。研究区属亚热带季风气候,年平均气温为16.4 ℃,年降水量为1 628.6 mm。蜡梅、柳叶蜡梅与山蜡梅的采集时间分别为2023年1月9—11日、2022年11月5—7日与2022年11月8—10日。在晴天的9:00、12:00和15:00分别收集3种蜡梅属植物花的挥发物。从每株植物上选取2朵盛开期的花(约2.0 g)作为样品。每个物种各选择3个植株进行重复试验。

图 1 3种蜡梅属植物的花部形态特征
Figure 1. Flower morphological characteristics of 3 Chimonanthus species
-
使用SPME装置收集挥发物。将新鲜盛开的花密封在顶空萃取瓶中,并在鉴定挥发物前平衡30 min。将SPME纤维(65 μm)插入气相色谱-质谱联用仪入口,在250 ℃下预处理30 min。将SPME纤维插入萃取瓶的顶部并提取挥发物,持续30 min。
-
提取挥发物后,使用带有HP-5MS毛细管柱(30 m×0.25 mm×0.25 μm)的GC/MS系统(5977B GC/MSD)进行GC-MS分析。GC烘箱温度的初始值设定为50 ℃,保持3 min,升温程序先以3 ℃·min−1的速度升至100 ℃,再以5 ℃·min−1的速度升至250 ℃,保持3 min。高纯度氦气(99.999%)被用作载气,流速为0.4 mL·min−1。MS分析:记录质谱仪以70 eV的电子能量在30~550扫描范围内获得的数据。离子源温度为230 ℃。
-
使用Qualitative Analysis 10.0将试验数据与NIST11.L和NIST05a.L 标准谱库数据(
https://webbook.NIST.gov/chemistry/name-ser/ )比较来鉴定花香物质。通过正构烷烃C7~C30计算保留指数,并结合网站资料,参考花香物质保留指数辅助质谱定性分析。用峰面积归一化法计算花香物质的相对含量。统计相对含量大于1%的物质,并使用Origin 2023进行主成分堆积图分析和层次聚类分析。 -
3种蜡梅属植物共鉴定出44种花香物质,蜡梅、柳叶蜡梅和山蜡梅的花分别含有19、23和18种花香物质(表1)。由表2和3可知:蜡梅的花香物质被分为6类,其中,醇类(60.83%~65.33%)和酯类(27.53%~31.71%)化合物含量最高。柳叶蜡梅和山蜡梅各鉴定出6类花香物质,其中,烯类化合物含量最高,分别为85.55%~92.96%、93.98%~98.60%。
表 1 3种蜡梅属植物花香物质的相对含量
Table 1. Relative content of floral substances of 3 Chimonanthus species
物质
类别物质名称 保留
指数相对含量/% 蜡梅 柳叶蜡梅 山蜡梅 烯类 莰烯 Camphene 964 −/−/− 2.31±0.22/2.32±0.08/
2.71±0.331.04±0.25/−/
1.63±0.06(-)-莰烯 L-Camphene 965 −/−/− −/−/− −/0.83±0.22/− β-水芹烯 β-Phellandrene 989 0.40±0.11/−/− 80.65±8.97/77.49±4.52/
79.98±4.5454.96±5.58/42.02±3.59/
55.50±1.46β-蒎烯 β-Pinene 1 006 −/−/− −/2.95±0.72/
6.43±0.625.00±0.77/2.29±0.64/
5.43±0.64月桂烯 β-Myrcene 1 007 0.99±0.15/1.01±0.05/
1.02±0.034.91±1.42/−/− −/−/− 4-蒈烯 (+)-4-Carene 1 027 −/−/− 1.09±0.31/0.70±0.17/
1.22±0.040.67±0.13/1.09±0.26/
1.38±0.16(+)-柠檬烯 D-Limonene 1 039 −/−/− −/−/− 30.67±7.26/47.41±6.08/
31.70±7.57(E)-Β-罗勒烯 trans-β-Ocimene 1 048 −/−/− −/−/0.06±0.01 −/−/− γ-松油烯 γ-Terpinene 1 067 −/−/− 2.49±0.45/1.88±0.08/
2.21±0.071.54±0.12/2.65±0.40/
2.46±0.29别罗勒烯 Allocimene B 1 135 1.26±0.23/1.36±0.06/
1.31±0.03−/−/− −/−/− (-)-α-荜澄茄油烯 α-Cubebene 1 352 −/−/− −/−/− −/−/0.08±0.02 毕澄茄烯 Cubebene 1 392 −/−/− −/−/− −/−/0.11±0.02 β-榄香烯 β-Elemen 1 394 −/0.09±0.03/− −/−/− −/−/− β-石竹烯 Caryophyllene 1 425 0.17±0.02/0.30±0.07/
0.13±0.020.21±0.02/0.12±0.03/
0.11±0.040.10±0.03/−/
0.31±0.03Δ-杜松烯 δ-Cadinene 1 513 −/−/− −/0.09±0.03/
0.24±0.06−/−/− 醇类 苯甲醇 Benzyl alcohol 1 044 10.76±2.02/7.49±0.54/
5.05±0.58−/−/− −/−/− 顺-Α,Α-5-三甲基-5-乙烯基四氢化呋喃-2-甲醇
2-Furanmethanol, 5-ethenyltetrahydro-α,α,5-trimethyl-, cis-1 096 0.60±0.24/−/
0.53±0.08−/−/− −/−/− 反式-4-(异丙基)-1-甲基环己-2-烯-1-醇
2-Cyclohexen-1-ol, 1-methyl-4-(1-methylethyl)-, trans-1 103 −/−/− 2.48±0.76/−/− −/−/− 芳樟醇 Linalool 1 112 49.14±8.82/55.59±2.81/
59.16±3.64−/0.41±0.12/− −/−/− 苯乙醇 Phenylethyl Alcohol 1 117 −/−/− −/0.55±0.21/
0.78±0.28−/−/− 冰片 endo-Borneol 1 167 −/−/− 0.83±0.20/−/− −/−/− 桃金娘烯醇 Myrtenol 1 174 −/0.06±0.01/− −/−/− −/−/− 2,2,6-三甲基-6-乙烯基四氢-2H-呋喃-3-醇
2H-Pyran-3-ol, 6-ethenyltetrahydro-2,2,6-trimethyl-1 178 −/0.09±0.01/
0.12±0.01−/−/− −/−/− 4-萜烯醇 Terpinen-4-ol 1 179 −/−/− −/−/0.14±0.02 0.17±0.04/−/− α-松油醇 α-Terpineol 1 192 −/−/− 2.06±0.20/7.31±2.76/
1.80±0.254.05±1.87/0.24±0.05/
0.69±0.28橙花醇 cis-Geraniol 1 260 0.07±0.02/0.11±0.01/
0.07±0.02−/−/− −/−/− 2,6-二甲基辛-2,7-二烯-1,6-二醇 2,7-Octadiene-1,6-
diol, 2,6-dimethyl-1 278 −/0.11±0.01/
0.05±0.01−/−/− −/−/− 肉桂醇 Cinnamyl alcohol 1 306 0.26±0.06/0.41±0.04/
0.35±0.03−/−/− −/−/− 酯类 乙酸苄酯 Benzyl acetate 1 172 24.34±1.73/19.33±0.47/
17.67±0.78−/−/− −/−/− 水杨酸甲酯 Methyl salicylate 1 197 7.37±1.77/8.60±0.79/
9.86±1.16−/−/− −/−/− 乙酸异龙脑酯 Isobornyl acetate 1 288 −/−/− −/0.11±0.03/− −/−/− 癸酸甲酯 Decanoic acid, methyl ester 1 328 −/−/− −/0.18±0.05/− −/−/− 二乙二醇丁醚醋酸酯 Ethanol, 2-(2-butoxyethoxy)-, acetate 1 371 −/−/− 0.28±0.04/
0.40±0.12/−0.32±0.13/
0.06±0.02/−2-癸烯酸甲酯 2-Decenoic acid, methyl ester 1 374 −/−/− −/0.33±0.08/− −/−/− 2,4-葵二烯酸甲酯 2,4-Decadienoic acid, methyl ester, (E,Z)- 1 397 −/−/− 0.97±0.33/2.81±0.16/
2.62±0.440.29±0.05/−/− 乙酸桂酯 Cinnamyl acetate 1 447 −/0.04±0.01/− −/−/− −/−/− 酚类 丁香酚 Eugenol 1 360 1.20±0.21/1.26±0.12/
0.99±0.19−/−/− −/−/− 醛类 苯甲醛 Benzaldehyde 977 0.15±0.02/0.12±0.00/
0.11±0.00−/−/− −/−/− 2-(4-甲基-3-环己烯-1-基)丙醛 3-Cyclohexene-1-
acetaldehyde, α,4-dimethyl-1 176 −/−/− −/−/− 0.96±0.19/2.55±0.78/
0.40±0.14杂环类 吲哚 Indole 1 294 3.30±0.23/4.03±0.09/
3.60±0.20−/−/− −/−/− 芳香烃类 对二甲苯 P-Xylene 915 −/−/− 1.00±0.27/1.82±0.54/
1.12±0.32−/0.46±0.12/
0.18±0.05酮类 樟脑 Camphor 1 147 −/−/− −/0.09±0.02/
0.12±0.04−/−/− 其他 (+)-氧化柠檬烯 (+)-Limonene oxide 1 137 −/−/− 0.74±0.13/0.45±0.02/
0.46±0.040.19±0.02/0.41±0.12/
0.12±0.04石竹素 Caryophyllene oxide 1 533 −/−/− −/−/− 0.05±0.01/−/− 总计 44 14/17/15 13/18/15 14/11/13 说明:相对含量的3个数据依次在9:00/12:00/15:00检测,数值为平均值±标准差。−表示未检出。 表 2 不同节律下3种蜡梅属植物花香物质的种类
Table 2. Statistics on the variety of floral substances of 3 Chimonanthus species different rhythms
物质类别 蜡梅 柳叶蜡梅 山蜡梅 物质类别 蜡梅 柳叶蜡梅 山蜡梅 种数 小计 种数 小计 种数 小计 种数 小计 种数 小计 种数 小计 烯类 4/4/3 5 6/7/8 9 7/6/9 10 杂环类 1/1/1 1 −/−/− − −/−/− − 醇类 5/7/7 8 3/3/3 6 2/1/1 2 芳香烃类 −/−/− − 1/1/1 1 −/1/1 1 酯类 2/3/2 3 2/5/1 5 2/1/− 2 酮类 −/−/− − −/1/1 1 −/−/− − 酚类 1/1/1 1 −/−/− − −/−/− − 其他 −/−/− − 1/1/1 1 2/1/1 2 醛类 1/1/1 1 −/−/− − 1/1/1 1 合计 14/17/15 19 13/18/15 23 14/11/13 18 说明:种数的3个数据依次在9:00/12:00/15:00检测。−表示未检出。 表 3 不同节律下3种蜡梅属植物花香物质的相对含量
Table 3. Statistics on the relative content of floral substances of 3 Chimonanthus species different rhythms
类别 相对含量/% 蜡梅 柳叶蜡梅 山蜡梅 烯类 2.82/2.76/2.46 91.66/85.55/92.96 93.98/96.29/98.60 醇类 60.83/63.86/65.33 5.37/8.27/2.72 4.22/0.24/0.69 酯类 31.71/27.97/27.53 1.25/3.83/2.62 0.61/0.06/− 酚类 1.20/1.26/0.99 −/−/− −/−/− 醛类 0.15/0.12/0.11 −/−/− 0.96/2.55/0.40 杂环类 3.30/4.03/3.60 −/−/− −/−/− 芳香烃类 −/−/− 1.00/1.82/1.12 −/0.46/0.18 酮类 −/−/− −/0.09/0.12 −/−/− 其他 −/−/− 0.74/0.45/0.46 0.24/0.41/0.12 说明:相对含量的3个数据依次在9:00/12:00/15:00检测。−表示未检出。 -
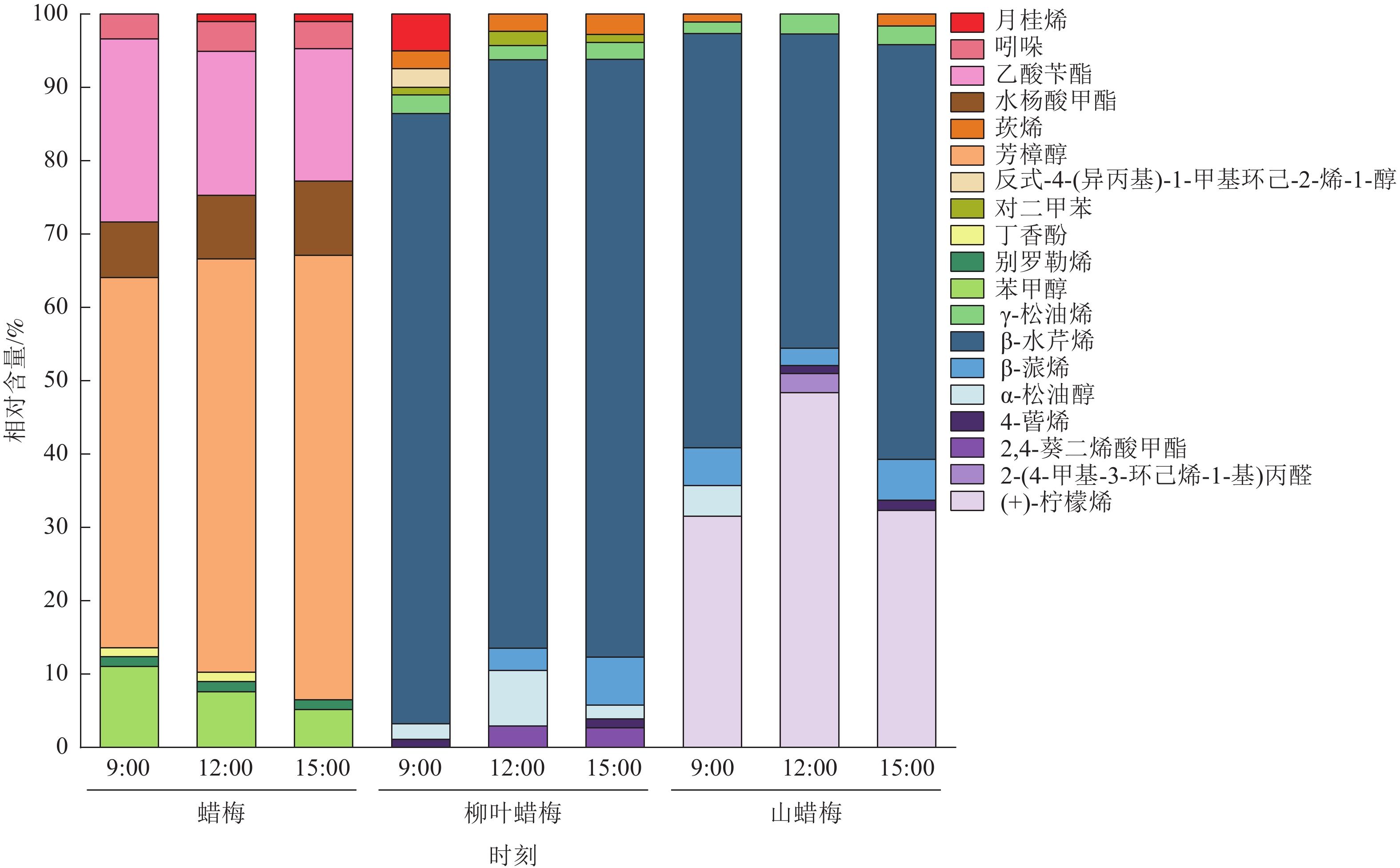
由图2可知:在3种蜡梅属植物中,共有19种物质占花香物质总相对含量的1%以上,主要为烯类(8种)、醇类(4种)和酯类(3种)化合物。
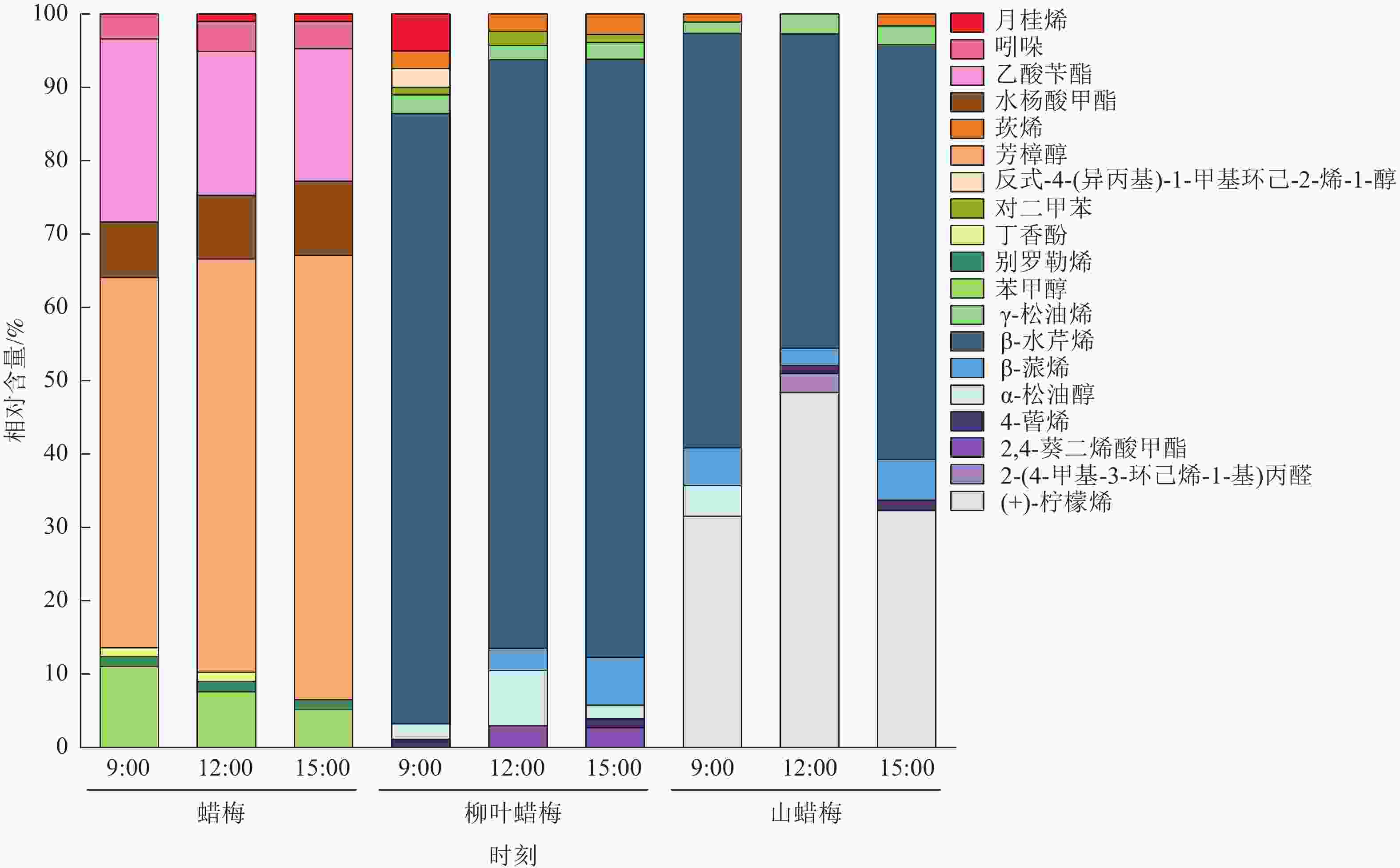
图 2 不同节律下3种蜡梅属植物花香物质的主成分堆积图
Figure 2. PCA stacking diagram of floral substances of 3 Chimonanthus species under different rhythms
蜡梅中有8种物质占花香物质总相对含量的1%以上,主要包括芳樟醇(49.14%~59.16%)、乙酸苄酯(17.67%~24.34%)、水杨酸甲酯(7.37%~9.86%)、苯甲醇(5.05%~10.76%)。芳樟醇和乙酸苄酯在蜡梅花香物质中所占比例较大。
在柳叶蜡梅和山蜡梅花中,分别有10和8种物质占花香物质总相对含量的1%以上。与蜡梅不同的是,柳叶蜡梅花香物质中的主要成分是β-水芹烯(77.49%~80.65%),山蜡梅的主要花香物质是β-水芹烯(42.02%~55.50%)和(+)-柠檬烯(30.67%~47.41%)。
-
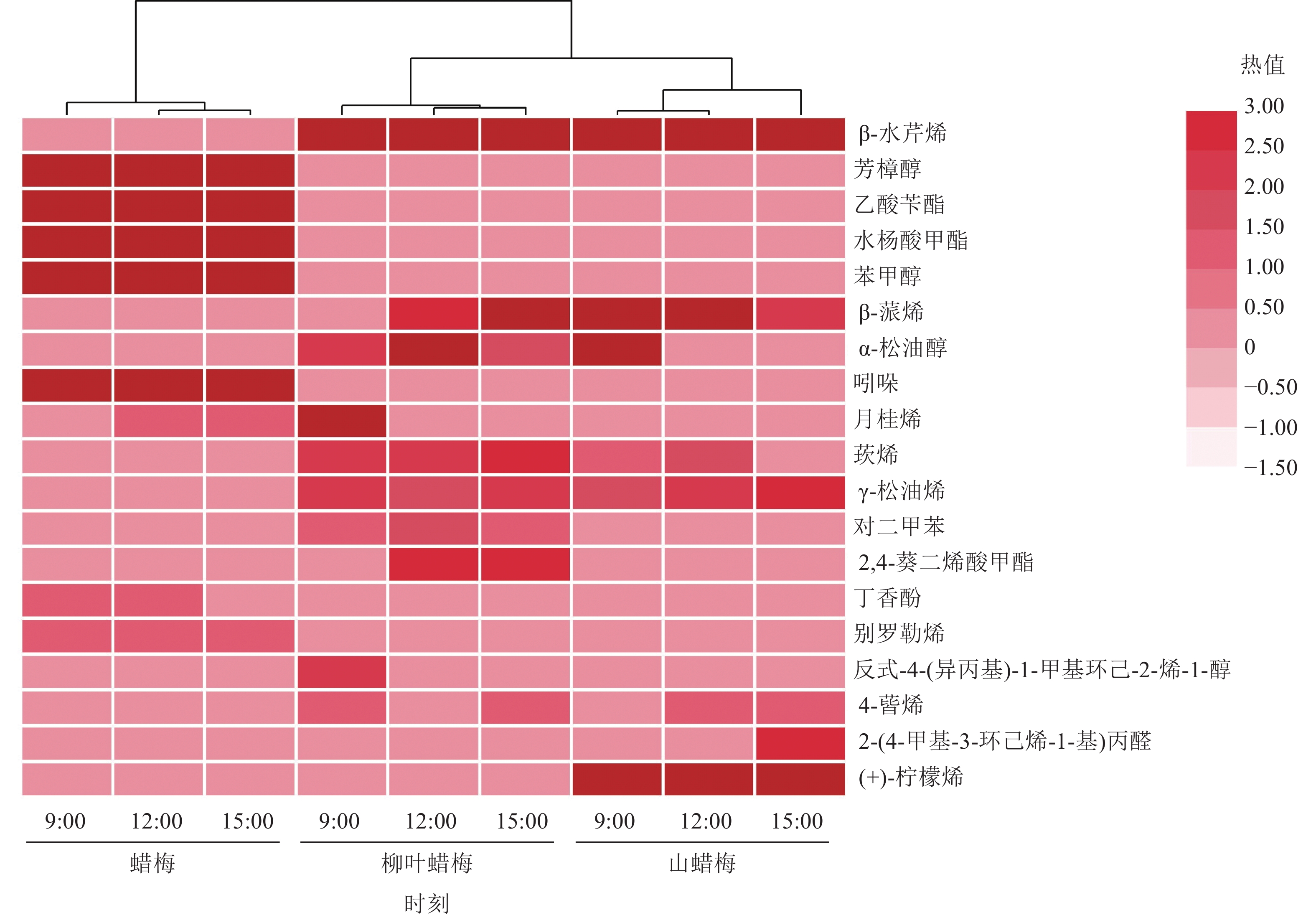
3种蜡梅属植物在上午、中午和下午的花香物质的组成和相对含量存在差异(图3)。蜡梅中含有8种花香物质,其中,芳樟醇和水杨酸甲酯的相对含量随着时间的变化递增,而乙酸苄酯与苯甲醇的相对含量随着时间的变化呈现逐渐减少的趋势,其他物质在上午、中午和下午的相对含量相近。在柳叶蜡梅的花香物质中,占比最大的β-水芹烯的相对含量无明显变化趋势,而β-蒎烯的相对含量随着时间的推移递增,此外,月桂烯仅存在于9:00的柳叶蜡梅花中。山蜡梅主要的花香物质在不同时间呈现不同的比例。β-水芹烯在上午和下午的相对含量比中午高,但(+)-柠檬烯在中午的相对含量最高,α-松油醇仅于9:00的山蜡梅花中被检测到。
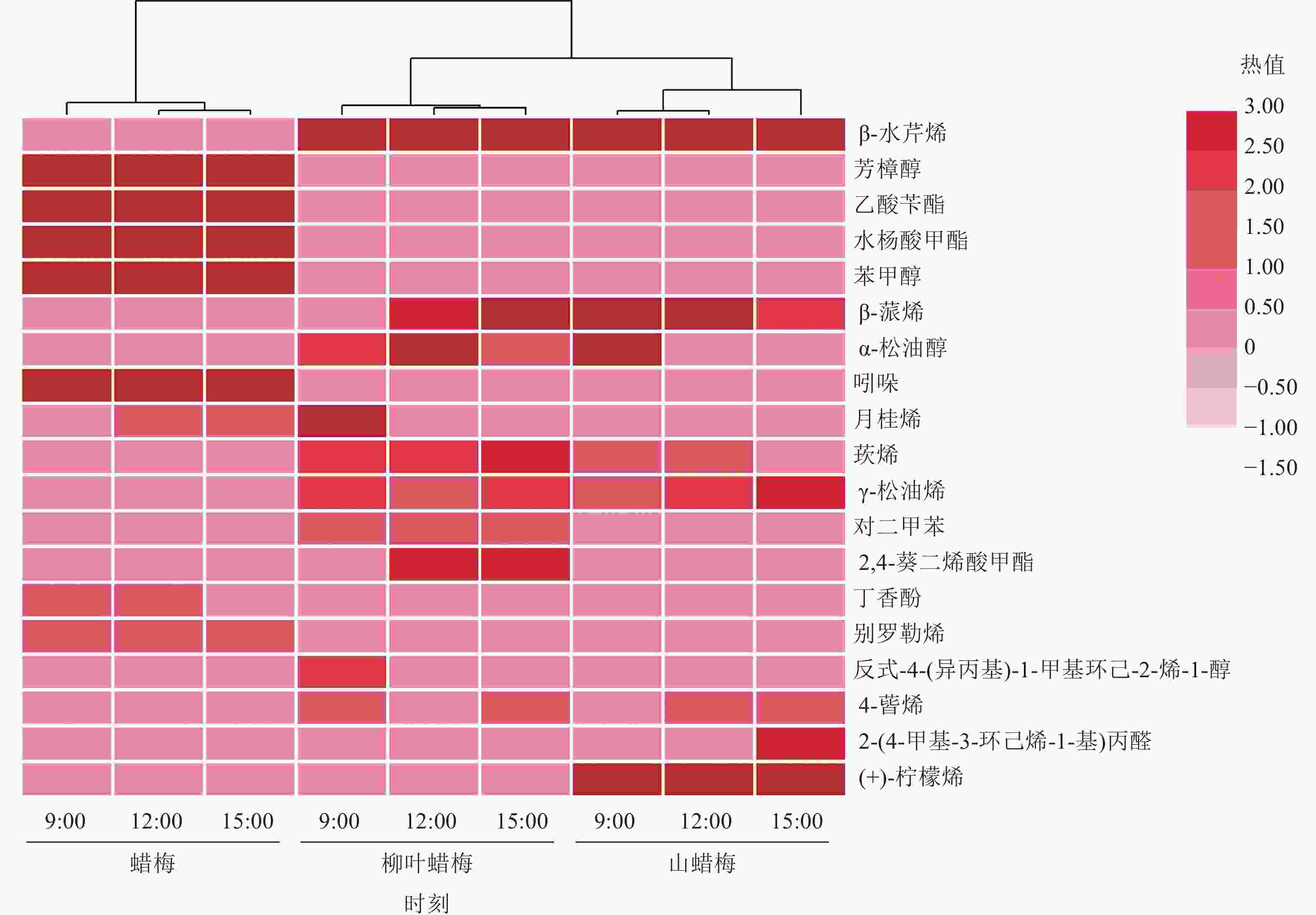
图 3 不同节律下3种蜡梅属植物主要花香物质的热图和层次聚类分析图
Figure 3. Heatmap and hierarchical cluster analysis of the main floral substances of 3 Chimonanthus species under different rhythms
-
本研究在蜡梅、柳叶蜡梅和山蜡梅花中共检测出44种花香物质。其中,从不同节律的蜡梅花中鉴定出19种花香物质,主要为芳樟醇和乙酸苄酯,这2种物质在其他针对蜡梅花香变化的研究中也被证明大量存在[22−24],这进一步验证了其在蜡梅花香中的核心作用。另外,蜡梅的主要花香成分包括烯类、醇类和酯类化合物,这与他人研究结果一致[25]。目前有关柳叶蜡梅和山蜡梅鲜花花香物质的研究较少,只在其干花中鉴定出α-月桂烯[26]。本研究表明柳叶蜡梅花中含量最多的花香物质是β-水芹烯,β-水芹烯和(+)-柠檬烯在山蜡梅的花中也占有很大比例。
-
从蜡梅、柳叶蜡梅和山蜡梅中分别鉴定出19、23和18种花香物质。结果表明3种蜡梅属植物之间的花香物质差异较大。3种蜡梅属植物鉴定出的花香物质中,只有β-水芹烯和β-石竹烯是3个物种共有的,多达30种物质仅在其中一个物种中检测到。仅存在于蜡梅花中的物质(15种)比仅存在于柳叶蜡梅(9种)和山蜡梅(6种)花中的物质多。2种花香物质(芳樟醇和β-月桂烯)在蜡梅和柳叶蜡梅花中存在,而在山蜡梅花中未检测到;10种物质是柳叶蜡梅和山蜡梅花共有的,而在蜡梅花中未检测到。
在9类花香物质中,烯类、醇类和酯类是种类与含量最丰富的类别。物种间的差异性与多样性也表现在这3个类别中。从3种蜡梅属植物中检测到15种烯类化合物,蜡梅中仅检测出5种,而柳叶蜡梅和山蜡梅花中分别检测到9和10种。只有2种烯类(β-水芹烯和β-石竹烯)为3个物种所共有。共检测到13种醇类化合物,蜡梅和柳叶蜡梅花中分别检测到8和6种,而山蜡梅花中仅检测到2种。其中,芳樟醇为蜡梅与柳叶蜡梅共有,4-萜烯醇和α-松油醇为柳叶蜡梅和山蜡梅共有。共检测到8种酯类化合物,其中2种为柳叶蜡梅和山蜡梅共有,3种为蜡梅特有,3种为柳叶蜡梅特有。
柳叶蜡梅和山蜡梅的花香物质含量较为相似,但蜡梅与其他2种植物的花香物质相对含量差异较大。在柳叶蜡梅和山蜡梅中,β-水芹烯是相对含量最丰富的花香物质,而在蜡梅中并未检测到。在蜡梅中,芳樟醇(49.14%~59.16%)与乙酸苄酯(17.67%~24.34%)是相对含量最高的花香物质,而柳叶蜡梅和山蜡梅花中不含芳樟醇与乙酸苄酯。这可能与系统发育关系有关。系统发育分析表明,柳叶蜡梅和山蜡梅的亲缘关系比蜡梅更近[27]。另一方面,作为常见的花香物质,芳樟醇和乙酸苄酯对传粉昆虫具有一定的吸引作用[28−29]。鉴于蜡梅特殊的低温开花环境,可以推测蜡梅的传粉昆虫影响其花香物质的释放。
-
3种蜡梅属植物花香物质的白天释放节律存在差异。柳叶蜡梅和山蜡梅花在上午、中午和下午的花香物质差异较大,而蜡梅的花香物质在3个时间段内变化相对较小。本研究显示:蜡梅在中午检测出的花香物质种类最多,这与先前关于蜡梅花香释放节律的报道结果一致[24]。同样,柳叶蜡梅也在中午释放出种类最为丰富的花香物质。然而,山蜡梅在3个时间段内检测出的花香物质的种类数接近,上午检测到的花香物质种类较多。相关研究揭示了矮牵牛Petunia hybrida、烟草Nicotiana suaveolens等多种园艺植物释放花香物质的昼夜节律模式[30−31]。矮牵牛作为模式物种被用于研究植物花香物质的释放节律[32],其在夜间散发出强烈的香味,这可能与传粉综合症有关。矮牵牛花香物质的释放遵循内源性昼夜节律,大约在黑暗4~6小时后达到峰值[33−34]。由此可见,温度、湿度与光照强度可能对花香物质的释放产生了影响。一般而言, 植物挥发性物质随着温度、光照等强度的增加而增加[35]。
-
从3种蜡梅属植物中共鉴定出44种花香物质,蜡梅、柳叶蜡梅和山蜡梅中分别鉴定出19、23和18种花香物质。这44种物质被划分为9类,其中烯类、醇类和酯类是种类与含量最丰富的类别。通过比较分析可知,柳叶蜡梅和山蜡梅花香物质较为相似,验证了两者在系统发育上的关联性。蜡梅的主要花香物质为芳樟醇和乙酸苄酯,与其他2种植物的花香物质种类差异较大。由于蜡梅特殊的低温开花环境,其传粉昆虫与花香之间的相互作用可能是花香物质产生独特性的原因。本研究揭示了蜡梅属3个物种花香物质的异同,为蜡梅属植物遗传育种、资源利用及开发更深层次的价值提供理论基础。
Floral substances and daytime release rhythms of 3 Chimonanthus species
-
摘要:
目的 花香是观赏植物的重要特性,对植物的繁殖至关重要。探讨蜡梅属Chimonanthus植物花香物质的生物合成和释放机制,为蜡梅属植物资源开发及利用提供理论基础。 方法 采用顶空-固相微萃取(HS-SPME)结合气相色谱-质谱联用技术(GC-MS)对蜡梅C. praecox、柳叶蜡梅C. salicifolius和山蜡梅C. nitens 等3种蜡梅属植物的花香物质进行鉴定和分析。 结果 3种蜡梅属植物共鉴定出44种花香物质,蜡梅、柳叶蜡梅和山蜡梅的花中分别鉴定出19、23和18种物质。蜡梅花中醇类和酯类化合物的相对含量最高,而柳叶蜡梅与山蜡梅的花香物质中烯类化合物的相对含量最高。3种蜡梅属植物花香物质的白天释放节律存在差异。柳叶蜡梅和山蜡梅在上午、中午和下午的花香物质差异较大,但蜡梅的花香物质在不同时段变化较小。系统发育分析表明:柳叶蜡梅和山蜡梅的亲缘关系比蜡梅更近。 结论 柳叶蜡梅和山蜡梅的花香物质较为相似,验证了两者在系统发育上的关联性。鉴于蜡梅特殊的低温开花环境,其传粉昆虫与花香之间的相互作用可能是花香物质产生独特性的原因。图3表3参35 Abstract:Objective Flower fragrance is an important characteristic of ornamental plants, which is very crucial for plants reproduction. This study aims to reveal the biosynthesis and release mechanism of floral substances in Chimonanthus, and to provide a solid theoretical basis for the utilization and development of Chimonanthus resources. Method Headspace solid phase microextraction (HS-SPME) and gas chromatography-mass spectrometry (GC-MS) were used to identify and analyze floral substances of 3 Chimonanthus species, including C. praecox, C. salicifolius and C. nitens. Result A total of 44 floral substances were identified in 3 Chimonanthus species, among which 19, 23 and 18 substances were identified in C. praecox, C. salicifolius and C. nitens respectively. The floral substances of alcohols and esters were the highest in C. praecox, while the content of alkenes was the highest in the floral substances of C. salicifolius and C. nitens. The daytime release rhythms of floral substances in 3 Chimonanthus species were different. The floral substances of C. salicifolius and C. nitens in the morning, noon and afternoon were different, but the floral substances of C. praecox changed little. Phylogenetic analysis showed that C. salicifolius and C. nitens were more closely related than C. praecox. Conclusion The floral substances of C. salicifolius and C. nitens are similar, which caters to the phylogenetic relationship between them. In view of the special low-temperature flowering environment of C. praecox, the interaction between pollination insects and floral fragrance may be the reason for the uniqueness of floral substances. [Ch, 3 fig. 3 tab. 35 ref.] -
Key words:
- Chimonanthus /
- floral substances /
- release rhythms
-
图 1 3种蜡梅属植物的花部形态特征
Figure 1 Flower morphological characteristics of 3 Chimonanthus species
图 2 不同节律下3种蜡梅属植物花香物质的主成分堆积图
Figure 2 PCA stacking diagram of floral substances of 3 Chimonanthus species under different rhythms
图 3 不同节律下3种蜡梅属植物主要花香物质的热图和层次聚类分析图
Figure 3 Heatmap and hierarchical cluster analysis of the main floral substances of 3 Chimonanthus species under different rhythms
表 1 3种蜡梅属植物花香物质的相对含量
Table 1. Relative content of floral substances of 3 Chimonanthus species
物质
类别物质名称 保留
指数相对含量/% 蜡梅 柳叶蜡梅 山蜡梅 烯类 莰烯 Camphene 964 −/−/− 2.31±0.22/2.32±0.08/
2.71±0.331.04±0.25/−/
1.63±0.06(-)-莰烯 L-Camphene 965 −/−/− −/−/− −/0.83±0.22/− β-水芹烯 β-Phellandrene 989 0.40±0.11/−/− 80.65±8.97/77.49±4.52/
79.98±4.5454.96±5.58/42.02±3.59/
55.50±1.46β-蒎烯 β-Pinene 1 006 −/−/− −/2.95±0.72/
6.43±0.625.00±0.77/2.29±0.64/
5.43±0.64月桂烯 β-Myrcene 1 007 0.99±0.15/1.01±0.05/
1.02±0.034.91±1.42/−/− −/−/− 4-蒈烯 (+)-4-Carene 1 027 −/−/− 1.09±0.31/0.70±0.17/
1.22±0.040.67±0.13/1.09±0.26/
1.38±0.16(+)-柠檬烯 D-Limonene 1 039 −/−/− −/−/− 30.67±7.26/47.41±6.08/
31.70±7.57(E)-Β-罗勒烯 trans-β-Ocimene 1 048 −/−/− −/−/0.06±0.01 −/−/− γ-松油烯 γ-Terpinene 1 067 −/−/− 2.49±0.45/1.88±0.08/
2.21±0.071.54±0.12/2.65±0.40/
2.46±0.29别罗勒烯 Allocimene B 1 135 1.26±0.23/1.36±0.06/
1.31±0.03−/−/− −/−/− (-)-α-荜澄茄油烯 α-Cubebene 1 352 −/−/− −/−/− −/−/0.08±0.02 毕澄茄烯 Cubebene 1 392 −/−/− −/−/− −/−/0.11±0.02 β-榄香烯 β-Elemen 1 394 −/0.09±0.03/− −/−/− −/−/− β-石竹烯 Caryophyllene 1 425 0.17±0.02/0.30±0.07/
0.13±0.020.21±0.02/0.12±0.03/
0.11±0.040.10±0.03/−/
0.31±0.03Δ-杜松烯 δ-Cadinene 1 513 −/−/− −/0.09±0.03/
0.24±0.06−/−/− 醇类 苯甲醇 Benzyl alcohol 1 044 10.76±2.02/7.49±0.54/
5.05±0.58−/−/− −/−/− 顺-Α,Α-5-三甲基-5-乙烯基四氢化呋喃-2-甲醇
2-Furanmethanol, 5-ethenyltetrahydro-α,α,5-trimethyl-, cis-1 096 0.60±0.24/−/
0.53±0.08−/−/− −/−/− 反式-4-(异丙基)-1-甲基环己-2-烯-1-醇
2-Cyclohexen-1-ol, 1-methyl-4-(1-methylethyl)-, trans-1 103 −/−/− 2.48±0.76/−/− −/−/− 芳樟醇 Linalool 1 112 49.14±8.82/55.59±2.81/
59.16±3.64−/0.41±0.12/− −/−/− 苯乙醇 Phenylethyl Alcohol 1 117 −/−/− −/0.55±0.21/
0.78±0.28−/−/− 冰片 endo-Borneol 1 167 −/−/− 0.83±0.20/−/− −/−/− 桃金娘烯醇 Myrtenol 1 174 −/0.06±0.01/− −/−/− −/−/− 2,2,6-三甲基-6-乙烯基四氢-2H-呋喃-3-醇
2H-Pyran-3-ol, 6-ethenyltetrahydro-2,2,6-trimethyl-1 178 −/0.09±0.01/
0.12±0.01−/−/− −/−/− 4-萜烯醇 Terpinen-4-ol 1 179 −/−/− −/−/0.14±0.02 0.17±0.04/−/− α-松油醇 α-Terpineol 1 192 −/−/− 2.06±0.20/7.31±2.76/
1.80±0.254.05±1.87/0.24±0.05/
0.69±0.28橙花醇 cis-Geraniol 1 260 0.07±0.02/0.11±0.01/
0.07±0.02−/−/− −/−/− 2,6-二甲基辛-2,7-二烯-1,6-二醇 2,7-Octadiene-1,6-
diol, 2,6-dimethyl-1 278 −/0.11±0.01/
0.05±0.01−/−/− −/−/− 肉桂醇 Cinnamyl alcohol 1 306 0.26±0.06/0.41±0.04/
0.35±0.03−/−/− −/−/− 酯类 乙酸苄酯 Benzyl acetate 1 172 24.34±1.73/19.33±0.47/
17.67±0.78−/−/− −/−/− 水杨酸甲酯 Methyl salicylate 1 197 7.37±1.77/8.60±0.79/
9.86±1.16−/−/− −/−/− 乙酸异龙脑酯 Isobornyl acetate 1 288 −/−/− −/0.11±0.03/− −/−/− 癸酸甲酯 Decanoic acid, methyl ester 1 328 −/−/− −/0.18±0.05/− −/−/− 二乙二醇丁醚醋酸酯 Ethanol, 2-(2-butoxyethoxy)-, acetate 1 371 −/−/− 0.28±0.04/
0.40±0.12/−0.32±0.13/
0.06±0.02/−2-癸烯酸甲酯 2-Decenoic acid, methyl ester 1 374 −/−/− −/0.33±0.08/− −/−/− 2,4-葵二烯酸甲酯 2,4-Decadienoic acid, methyl ester, (E,Z)- 1 397 −/−/− 0.97±0.33/2.81±0.16/
2.62±0.440.29±0.05/−/− 乙酸桂酯 Cinnamyl acetate 1 447 −/0.04±0.01/− −/−/− −/−/− 酚类 丁香酚 Eugenol 1 360 1.20±0.21/1.26±0.12/
0.99±0.19−/−/− −/−/− 醛类 苯甲醛 Benzaldehyde 977 0.15±0.02/0.12±0.00/
0.11±0.00−/−/− −/−/− 2-(4-甲基-3-环己烯-1-基)丙醛 3-Cyclohexene-1-
acetaldehyde, α,4-dimethyl-1 176 −/−/− −/−/− 0.96±0.19/2.55±0.78/
0.40±0.14杂环类 吲哚 Indole 1 294 3.30±0.23/4.03±0.09/
3.60±0.20−/−/− −/−/− 芳香烃类 对二甲苯 P-Xylene 915 −/−/− 1.00±0.27/1.82±0.54/
1.12±0.32−/0.46±0.12/
0.18±0.05酮类 樟脑 Camphor 1 147 −/−/− −/0.09±0.02/
0.12±0.04−/−/− 其他 (+)-氧化柠檬烯 (+)-Limonene oxide 1 137 −/−/− 0.74±0.13/0.45±0.02/
0.46±0.040.19±0.02/0.41±0.12/
0.12±0.04石竹素 Caryophyllene oxide 1 533 −/−/− −/−/− 0.05±0.01/−/− 总计 44 14/17/15 13/18/15 14/11/13 说明:相对含量的3个数据依次在9:00/12:00/15:00检测,数值为平均值±标准差。−表示未检出。  下载: 导出CSV
下载: 导出CSV
表 2 不同节律下3种蜡梅属植物花香物质的种类
Table 2. Statistics on the variety of floral substances of 3 Chimonanthus species different rhythms
物质类别 蜡梅 柳叶蜡梅 山蜡梅 物质类别 蜡梅 柳叶蜡梅 山蜡梅 种数 小计 种数 小计 种数 小计 种数 小计 种数 小计 种数 小计 烯类 4/4/3 5 6/7/8 9 7/6/9 10 杂环类 1/1/1 1 −/−/− − −/−/− − 醇类 5/7/7 8 3/3/3 6 2/1/1 2 芳香烃类 −/−/− − 1/1/1 1 −/1/1 1 酯类 2/3/2 3 2/5/1 5 2/1/− 2 酮类 −/−/− − −/1/1 1 −/−/− − 酚类 1/1/1 1 −/−/− − −/−/− − 其他 −/−/− − 1/1/1 1 2/1/1 2 醛类 1/1/1 1 −/−/− − 1/1/1 1 合计 14/17/15 19 13/18/15 23 14/11/13 18 说明:种数的3个数据依次在9:00/12:00/15:00检测。−表示未检出。
下载: 导出CSV
表 3 不同节律下3种蜡梅属植物花香物质的相对含量
Table 3. Statistics on the relative content of floral substances of 3 Chimonanthus species different rhythms
类别 相对含量/% 蜡梅 柳叶蜡梅 山蜡梅 烯类 2.82/2.76/2.46 91.66/85.55/92.96 93.98/96.29/98.60 醇类 60.83/63.86/65.33 5.37/8.27/2.72 4.22/0.24/0.69 酯类 31.71/27.97/27.53 1.25/3.83/2.62 0.61/0.06/− 酚类 1.20/1.26/0.99 −/−/− −/−/− 醛类 0.15/0.12/0.11 −/−/− 0.96/2.55/0.40 杂环类 3.30/4.03/3.60 −/−/− −/−/− 芳香烃类 −/−/− 1.00/1.82/1.12 −/0.46/0.18 酮类 −/−/− −/0.09/0.12 −/−/− 其他 −/−/− 0.74/0.45/0.46 0.24/0.41/0.12 说明:相对含量的3个数据依次在9:00/12:00/15:00检测。−表示未检出。
下载: 导出CSV
-
[1] 陈龙清, 陈俊愉. 蜡梅属植物的形态、分布、分类及其应用[J]. 中国园林, 1999(1): 76−77. CHEN Longqing, CHEN Junyu. Morphology, distribution, classification and application of Chimonanthus [J]. Chinese Landscape Architecture, 1999(1): 76−77. [2] ZHOU Mingqin, CHEN Longqing, RUAN Rui. Genetic diversity of Chimonanthus nitens Oliv. complex revealed using inter-simple sequence repeat markers [J]. Scientia Horticulturae, 2012, 136: 38−42. [3] 伍碧华, 徐恒伟, 明军, 等. 四川大巴山野生蜡梅资源现状与保护利用[J]. 中国野生植物资源, 2009, 28(5): 33−36. WU Bihua, XU Hengwei, MING Jun, et al. Existence status, protection and utilization of wild wintersweet (Chimonanthus Lindley) in Dabashan Sichuan [J]. Chinese Wild Plant Resources, 2009, 28(5): 33−36. [4] 赵冰, 张启翔. 中国蜡梅属种质资源的分布及其特点[J]. 广西植物, 2007(5): 730−735. ZHAO Bing, ZHANG Qixiang. Distribution and characteristic of Chimonanthus germplasm in China [J]. Guihaia, 2007(5): 730−735. [5] 张川英, 李婷婷, 龚笑飞, 等. 遂昌乌溪江流域山蜡梅生境群落特征与物种多样性[J]. 浙江农林大学学报, 2023, 40(4): 848−858. ZHANG Chuanying, LI Tingting, GONG Xiaofei, et al. Habitat community characteristics and species diversity of Chimonanthus nitens in Wuxi River Basin, Suichang [J]. Journal of Zhejiang A&F University, 2023, 40(4): 848−858. [6] 黎鑫, 杨菲, 陈美霞, 等. 柳叶蜡梅化学成分及生物活性研究进展[J]. 粮食与油脂, 2023, 36(1): 25−28, 33. LI Xin, YANG Fei, CHEN Meixia, et al. Research progress on chemical constituents and biological activities of Chimonanthus salicifolius [J]. Cereals and Oils, 2023, 36(1): 25−28, 33. [7] 涂昆, 邹峥嵘. 蜡梅属植物生物碱的生物活性和合成研究进展[J]. 中草药, 2017, 48(11): 2340−2352. TU Kun, ZOU Zhengrong. Research progress on biological activities and synthesis of alkaloids in plants from Chimonanthus Lindl [J]. Chinese Traditional and Herbal Drugs, 2017, 48(11): 2340−2352. [8] 徐金标, 潘俊杰, 吕群丹, 等. 蜡梅科植物化学成分及其药理活性研究进展[J]. 中国中药杂志, 2018, 43(10): 1957−1968. XU Jinbiao, PAN Junjie, LÜ Qundan, et al. Research advances on chemical constituents from Calycanthaceae plants and their pharmacological activities [J]. China Journal of Chinese Materia Medical, 2018, 43(10): 1957−1968. [9] 李琦, 周君美, 邢丙聪, 等. 蜡梅属药用植物分子生物学研究进展[J]. 中国现代应用药学, 2022, 39(12): 1646−1654. LI Qi, ZHOU Junmei, XING Bingcong, et al. Research progress on the molecular biology of genus Chimonanthus medicinal plants [J]. Chinese Journal of Modern Applied Pharmacy, 2022, 39(12): 1646−1654. [10] ZHOU Xiaojie, YU Yang, CHENG Zhang, et al. Anti-depression effect of Chimonanthus salicifolicus essential oil in chronic stressed rats [J]. Journal of Medicinal Plants Research, 2014, 8: 430−435. [11] WANG Kuiwu, LI Dan, WU Bin, et al. New cytotoxic dimeric and trimeric coumarins from Chimonanthus salicifolius [J]. Phytochemistry Letters, 2016, 16: 115−120. [12] 李帅岚, 邹峥嵘. 蜡梅属植物中黄酮和香豆素类成分及药理活性研究进展[J]. 中草药, 2018, 49(14): 3425−3431. LI Shuailan, ZOU Zhengrong. Research progress on flavonoids and coumarins from Chimonanthus plants and its pharmacological activities [J]. Chinese Traditional and Herbal Drugs, 2018, 49(14): 3425−3431. [13] WANG Ning, CHEN Hui, XIONG Lei, et al. Phytochemical profile of ethanolic extracts of Chimonanthus salicifolius S. Y. Hu. leaves and its antimicrobial and antibiotic-mediating activity [J]. Industrial Crops & Products, 2018, 125: 328−334. [14] 熊义权, 肖纯, 龙秀娟. 蜡梅属植物叶、花化学成分及药理性质研究进展[J]. 中国野生植物资源, 2008(1): 8−10, 15. XIONG Yiquan, XIAO Chun, LONG Xiujuan. Research progress on chemical constituents and pharmacological properties of leaves and flowers of Chimonanthus [J]. Chinese Wild Plant Resources, 2008(1): 8−10, 15. [15] 董瑞霞, 潘俊杰, 金叶, 等. 两种蜡梅属药食兼用茶化学成分及药理活性研究进展[J]. 食品工业科技, 2021, 42(16): 429−437. DONG Ruixia, PAN Junjie, JIN Ye, et al. Research advances on chemical constituents and their pharmacological activities of two kinds of medicinal and edible tea from Chimonanthus Lindl [J]. Science and Technology of Food Industry, 2021, 42(16): 429−437. [16] 杨钰, 王艺光, 董彬, 等. 不同梅花品种花香成分鉴定与分析[J]. 浙江农林大学学报, 2024, 41(2): 262−274. YANG Yu, WANG Yiguang, DONG Bin, et al. Identification and analysis of floral scent compounds of Prunus mume cultivars [J]. Journal of Zhejiang A&F University, 2024, 41(2): 262−274. [17] SHANG Junzhong, TIAN Jingpu, CHENG Huihui, et al. The chromosome-level wintersweet (Chimonanthus praecox) genome provides insights into floral scent biosynthesis and flowering in winter [J/OL]. Genome Biology, 2020, 21(1): 200[2023-03-20]. DOI: 10.1186/s13059-020-02088-y. [18] TIAN Jingpu, MA Zhiyao, ZHAO Kaige, et al. Transcriptomic and proteomic approaches to explore the differences in monoterpene and benzenoid biosynthesis between scented and unscented genotypes of wintersweet [J]. Physiologia Plantarum, 2019, 166: 478−493. [19] 徐萌, 张经纬, 吴令上, 等. HS-SPME-GC-MS联用测定蜡梅属植物花的挥发性成分[J]. 林业科学, 2016, 52(12): 58−65. XU Meng, ZHANG Jingwei, WU Lingshang, et al. Determination of volatile components from Chimonanthus flowers by HS-SPME-GC-MS [J]. Scientia Silvae Sinicae, 2016, 52(12): 58−65. [20] LI Yue, JIA Weijia, WANG Qiong, et al. Comparative analysis of floral scent profiles between two Chimonanthus praecox plants under different rhythms and blooming stages [J/OL]. Scientia Horticulturae, 2022, 301 : 111129[2023-03-20]. DOI: 10.1016/j.scienta.2022.111129. [21] 蒙柳贝. 三个品种蜡梅花香成分的差异及动态变化研究[D]. 昆明: 西南林业大学, 2024. MENG Liubei. Study on the Differences and Dynamic Chances of Floral Fragrance Compounds of Three Varieties of Chimonanthus praecox [D]. Kunming: Southwest Forestry University, 2024. [22] 周继荣, 倪德江. 蜡梅不同品种和花期香气变化及其花茶适制性[J]. 园艺学报, 2010, 37(10): 1621−1628. ZHOU Jirong, NI Dejiang. Changes in flower aroma compounds of cultivars of Chimonanthus praecox (L. ) Link and at different stages relative to Chimonanthus tea quality [J]. Acta Horticulturae Sinica, 2010, 37(10): 1621−1628. [23] 陆安霞, 周心如, 叶玉龙, 等. 蜡梅花离体摊放过程中香气感官评价和挥发性物质分析[J]. 园艺学报, 2020, 47(1): 73−84. LU Anxia, ZHOU Xinru, YE Yulong, et al. Changes of sensory characteristic and volatiles of harvested flowers of Chimonanthus praecox during spreading process [J]. Acta Horticulturae Sinica, 2020, 47(1): 73−84. [24] 余莉. 蜡梅花挥发性组分与花色色素分析[D]. 武汉: 华中农业大学, 2013. YU Li. The Study of Volatile Components and Flower Pigments in Chimonanthus praecox [D]. Wuhan: Huazhong Agricultural University, 2013. [25] 冯楠. 蜡梅花香挥发物测定及2个萜烯合酶基因功能初步研究[D]. 武汉: 华中农业大学, 2017. FENG Nan. Determination of Floral Volatile Components and Preliminary Study on Function of Two Terpene Synthases in Wintersweet [D]. Wuhan: Huazhong Agricultural University, 2017. [26] 林霞. 蜡梅属植物中挥发性成分的研究进展[J]. 福建农业科技, 2019, 50(7): 57−64. LIN Xia. Research progress on volatile components of Chimonanthus Lindl. [J]. Fujian Agricultural Science and Technology, 2019, 50(7): 57−64. [27] ZHOU Shiliang, RENNER S S, WEN Jun. Molecular phylogeny and intra- and intercontinental biogeography of Calycanthaceae [J]. Molecular Phylogenetics and Evolution, 2006, 39(1): 1−15. [28] YANG Xiulian, YUE Yuanzheng, LI Haiyan, et al. The chromosome-level quality genome provides insights into the evolution of the biosynthesis genes for aroma compounds of Osmanthus fragrans [J]. Horticulture Research, 2018, 5 : 72[2023-03-20]. DOI: 10.1038/s41438-018-0108-0. [29] ZHANG Liangsheng, CHEN Fei, ZHANG Xingtan, et al. The water lily genome and the early evolution of flowering plants [J]. Nature, 2020, 577(7788): 79−84. [30] ROEDER S, HARTMANN A, EFFMERT U, et al. Regulation of simultaneous synthesis of floral scent terpenoids by the 1, 8-cineole synthase of Nicotiana suaveolens [J]. Plant Molecular Biology, 2007, 65: 107−124. [31] CHENG Sihua, FU Xiumin, MEI Xin, et al. Regulation of biosynthesis and emission of volatile phenylpropanoids/benzenoids in petunia×hybrida flowers by multi-factors of circadian clock, light, and temperature [J]. Plant Physiology and Biochemistry, 2016, 107: 1−8. [32] FENSKE M P, IMAIZUMI T. Circadian rhythms in floral scent emission [J/OL]. Frontiers in Plant Science, 2016, 7 : 462[2023-03-20]. DOI: 10.3389/fpls.2016.00462. [33] UNDERWOOD B A, TIEMAN D M, SHIBUYA K, et al. Ethylene-regulated floral volatile synthesis in petunia corollas [J]. Plant Physiology, 2005, 138: 255−266. [34] FENSKE M P, HEWETT HAZELTON K D, HEMPTON A K, et al. Circadian clock gene LATE ELONGATED HYPOCOTYL directly regulates the timing of floral scent emission in Petunia [J]. Proceedings of the National Academy of Sciences of the United States of America, 2015, 112: 9775−9780. [35] 蒋冬月, 李永红, 沈鑫. 芸香叶片和花瓣释放挥发性有机物成分及其变化规律[J]. 浙江农林大学学报, 2018, 35(3): 572−580. JIANG Dongyue, LI Yonghong, SHEN Xin. Components and variations of volatile organic compounds released from leaves and flowers of Ruta graveolens [J]. Journal of Zhejiang A&F University, 2018, 35(3): 572−580. -

-
链接本文:
https://zlxb.zafu.edu.cn/article/doi/10.11833/j.issn.2095-0756.20240266
 点击查看大图
点击查看大图

计量
- 文章访问数: 1708
- HTML全文浏览量: 469
- PDF下载量: 31
- 被引次数: 0
