






-
森林乡村指的是乡村自然生态风貌保存完好,乡土田园特色突出,森林氛围浓郁,森林功能效益显著,涉林产业发展良好,人居环境整洁,保护管理有效的生态宜居乡村。为贯彻乡村振兴战略,建设生态宜居的人居环境,有效保护森林乡村,2019年12月,国家林业和草原局先后公布了2批国家森林乡村名录,自此,森林乡村的保护上升到国家层面。森林乡村的空间分布会对区域空间整体效应的发挥产生直接影响[1],因此,探索森林乡村的空间分布特征及其影响因素,便于全方位地识别区域内森林乡村的空间关系及布局特征。优化森林乡村的空间布局,正确认识森林乡村的形成因素,对于推进生态文明建设及乡村振兴实践具有重要意义[2]。
目前,关于森林乡村的研究处于初步探索阶段,尚未形成系统的理论体系。从研究主题上看,较多关注与森林乡村相关的乡村绿化[3-5]、森林资源开发[6] 、森林旅游[7]、森林法制建设[8-9]、生态景观价值[10]、土地利用[11]、森林环境收入[12-13]、健康教育[14]等方面;从研究方法上看,多聚焦于定性分析及实证研究;从研究尺度上看,多聚焦于国家级层面宏观尺度上大样本数据的定量分析[15]和村级层面微观尺度上小样本数据的实证分析[16],为后续研究的开展奠定了理论基础。但仍然存在不足,从研究主题上看,以森林乡村空间分布为主题的研究并不多,而探讨森林乡村的空间分布特征及影响因素有助于优化森林乡村的空间布局,对于提高区域辐射效应,分区分类推进乡村振兴战略具有重要意义。从研究方法上看,利用大数据及地理信息系统(GIS)空间分析法定量研究森林乡村的空间格局可以为区域内森林乡村的发展提供科学依据。从研究尺度上看,对省市内部森林乡村的空间分布差异缺乏关注,这在一定程度上限制了对森林乡村分布特征的认识。因此,亟需拓展中微观尺度上森林乡村的空间异质性研究。浙江省作为“绿水青山就是金山银山”理念的发源地及率先实践地,生态文明建设成效显著,示范效应强,是生态文明建设及乡村振兴的典范。本研究从省级尺度分析浙江省森林乡村的空间分布特征及其影响因素,不仅能丰富乡村地理学的研究内容,为森林乡村的全域发展提供空间统计上的依据,而且能为后续森林乡村的深入研究提供方法借鉴,从而为区域内森林乡村的保护及可持续发展提供科学依据。
-
浙江省具有独特的地理环境,森林面积为660.23万 hm2,森林覆盖率为61.15%,森林资源丰富,森林乡村数量众多。自2019年开展森林乡村的评选以来,浙江省共有447个乡村入选森林乡村名录。
选取浙江省447个森林乡村为对象,通过百度应用程序编接口(API)坐标拾取器拾取各个森林乡村的经纬度,用ArcGIS 10.5软件构建浙江省森林乡村的矢量点位。森林乡村的社会经济数据来源于浙江省统计局(
http://tjj.zj.gov.cn/ )、浙江政务网(http://hz.zjzwfw.gov.cn/ )、浙江文物网(http://wwj.zj.gov.cn/ )、浙江省生态环境厅。浙江省的数字高程模型(DEM)、气温等数据来源于中国科学院地理空间云数据中心(http://www.gscloud.cn/ )。将百度API坐标拾取器拾取到的447个森林乡村的经纬度坐标保存到Excel中,用ArcGIS 10.5软件导入Excel中的坐标点,配置在浙江省域底图上,对气温、降水、DEM等矢量图进行边界裁剪处理。 -
地理学中用基尼系数来分析区域空间的离散分布情况[17]。通过空间基尼系数研究浙江省森林乡村在省域尺度上的空间分布情况,公式为:
$$ G_{\rm{{i n i}}}=\frac{-\displaystyle \sum\limits_{i=1}^{N} p_{i} \ln p_{i}}{\ln N} 。 $$ (1) 式(1)中:Gini为基尼系数,pi为第i个区域中森林乡村占总数的比例,N为浙江省下辖地级行政区的数量。Gini取值为0~1,Gini越接近1,集中程度越高。Gini<0.2代表绝对平均,0.2≤Gini<0.3代表比较平均,0.3≤Gini<0.4代表相对合理,0.4≤Gini<0.5代表差距较大, Gini≥0.5代表差距悬殊。
-
采用ArcGIS中的衡量核密度分析点要素的集群分布状态,即研究浙江省森林乡村的空间分布形态,公式为:
$$ D\left(X_{i}\right)=\frac{1}{u r} \sum\limits_{i=1}^{u} k\left(\frac{A_{i}}{r}\right)。 $$ (2) 式(2)中: Ai为估值点Xo到事件Xoi的距离,r>0为搜索半径,u为搜索半径范围内的点数量,k为空间权重函数。D(Xi)越大,表明森林乡村空间分布的密度越高。
-
通过ArcGIS中全局自相关判断森林乡村在省域层面空间分布上的自相关性(I),公式为:
$$ I=\frac{n}{S} \times \frac{\displaystyle \sum\limits_{i=1}^{n} \displaystyle \sum\limits_{j=1}^{n} \mathrm{W}_{i j}\left(X_{i}-\overline{X}\right)\left(X_{j}-\overline{X}\right)}{\displaystyle \sum\limits_{i=1}^{n}\left(X_{i}-\overline{X}\right)^{2}}。 $$ (3) 式(3)中:Xi和Xj表示在第i、j县域单元上森林乡村的数量,
$\overline X$ 为森林乡村数量的均值,Wij为i、j县域单元(90个县城行政区)之间的距离权重,S为距离权重之和,n为浙江省内森林乡村数量的总和。I的取值为[−1, 1]。若I∈[−1,0],表明具有空间负相关性;若I∈[0, 1],则表明具有空间正相关性,0表示不存在空间相关,即空间随机分布。此外,由式(3)可得出Z值,用Z值检验空间自相关的统计显著性,Z为正代表存在空间正相关,Z为负代表空间是分散的。森林乡村在县域上的局域关联指数(Gi):
$$ G_{i}=\frac{\displaystyle \sum\limits_{j=1}^{n} W_{i j} X_{j}-\overline{X} \displaystyle \sum\limits_{j=1}^{n} W_{i j}}{\sqrt[\leftroot{-1}\uproot{-8}{{H}}] {\dfrac{{n\displaystyle\sum\limits_{j = 1}^n {W_{ij}^2} - {{\left( {\displaystyle\sum\limits_{j = 1}^n {{W_{ij}}} } \right)}^2}}}{{n - 1}}} }。 $$ (4) 式(4)中:n为浙江省内森林乡村数量的总和,Wij为i、j县域单元之间的距离权重,H为森林乡村数量的标准差,
$\overline X $ 为森林乡村数量的均值,Xj表示在第j县域单元上森林乡村的数量。若Gi>0,表示森林乡村集聚程度与邻近地区相似,处于高—高集聚或者低—低集聚状态;若Gi<0,则表示该区域森林乡村集聚程度与邻近地区相异,处于低—高集聚或者高—低集聚状态。并用Jenks 自然断裂法将空间分布划分为冷点、次冷点、次热点、热点共4种类型,获得浙江省森林乡村冷热点空间分布图。 -
村庄是特定地区人地关系地域系统中“人”和“地”这2个子系统相互影响、相互作用演化形成的产物,其形成和分布受地理环境及人文社会因素共同影响[18]。与传统乡村相比,森林乡村具有较高的生态价值。自然禀赋是森林乡村形成与分布的基础,影响森林乡村的整体分布,人文底蕴使得区域内森林乡村的分布存在差异[19]。因此,本研究从自然及人文2个方面展开对森林乡村空间分布影响因素的分析[20-24],并选取地形地貌、气候条件、河流水系、森林资源作为自然因素下的二级因素,文化资源、社会经济基础及交通通达性为人文因素下的二级因素展开分析。
-
首先通过SPSS 26.0 的Pearson相关性检验功能,探究相关因素对浙江省森林乡村空间分布的影响程度。公式如下:
$$ R=\frac{\displaystyle \sum\limits_{i=1}^{m}\left(X_{i}-\overline{X}\right)\left(Y_{j}-\overline{Y}\right)}{\sqrt{\displaystyle \sum\limits_{{i}=1}^{{m}}\left(X_{i}-\overline{X}\right)^{2}} \sqrt{\displaystyle \sum\limits_{{i}=1}^{{m}}\left(Y_{i}-\overline{Y}\right)^{2}}} 。 $$ (5) 式(7)中:Xi 表示第i个县域单元上森林乡村的数量,Yi为在i县域单元上所选取影响因素的变量,m为选取变量的数量,
$\overline X_i $ 与$\overline Y_i $ 为2个变量的平均值。当R>0时,代表Xi与Yi正相关;R<0 时负相关性。其绝对值越接近1,则代表Xi与Yi的相关性越强,反之越弱[24]。 -
由于Pearson相关系数忽略了空间分层的异质性问题,不能准确反映数据的空间差异特征,因此,本研究从空间分异性视角,运用地理探测器的统计方法,进一步分析空间分布差异的影响因素。地理探测器的核心是通过空间分异性揭示因变量与自变量空间分布的相似性[25],以此度量影响因子的解释度(q)[26]。
$$ q=\frac{\displaystyle \sum\limits_{h=1}^{L} N_{h} \sigma_{h}^{2}}{N {\rm{\sigma}}^{2}} 。 $$ (6) 式(6)中:h=1
$,\cdots, $ L为因变量或自变量的分区,N和σ2分别为研究区域内观测点的数量(浙江省内森林乡村数量的总和)与总体方差,Nh和σh2即为第h个子区域内森林乡村的数量与总体方差。q的取值为[0,1],q值越大代表自变量对因变量的解释力越强,即某因素对森林乡村的数量影响越强,反之则越弱。 -
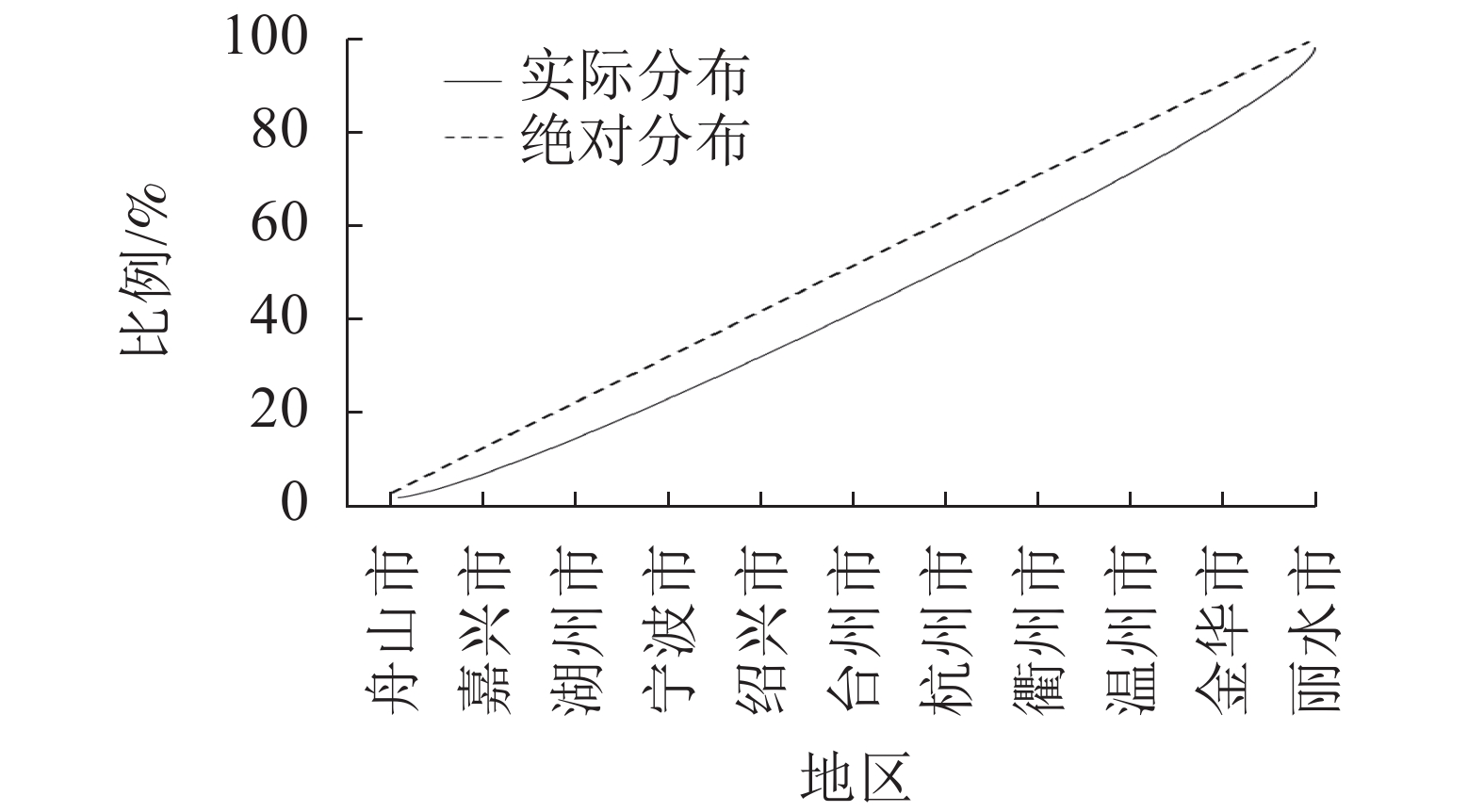
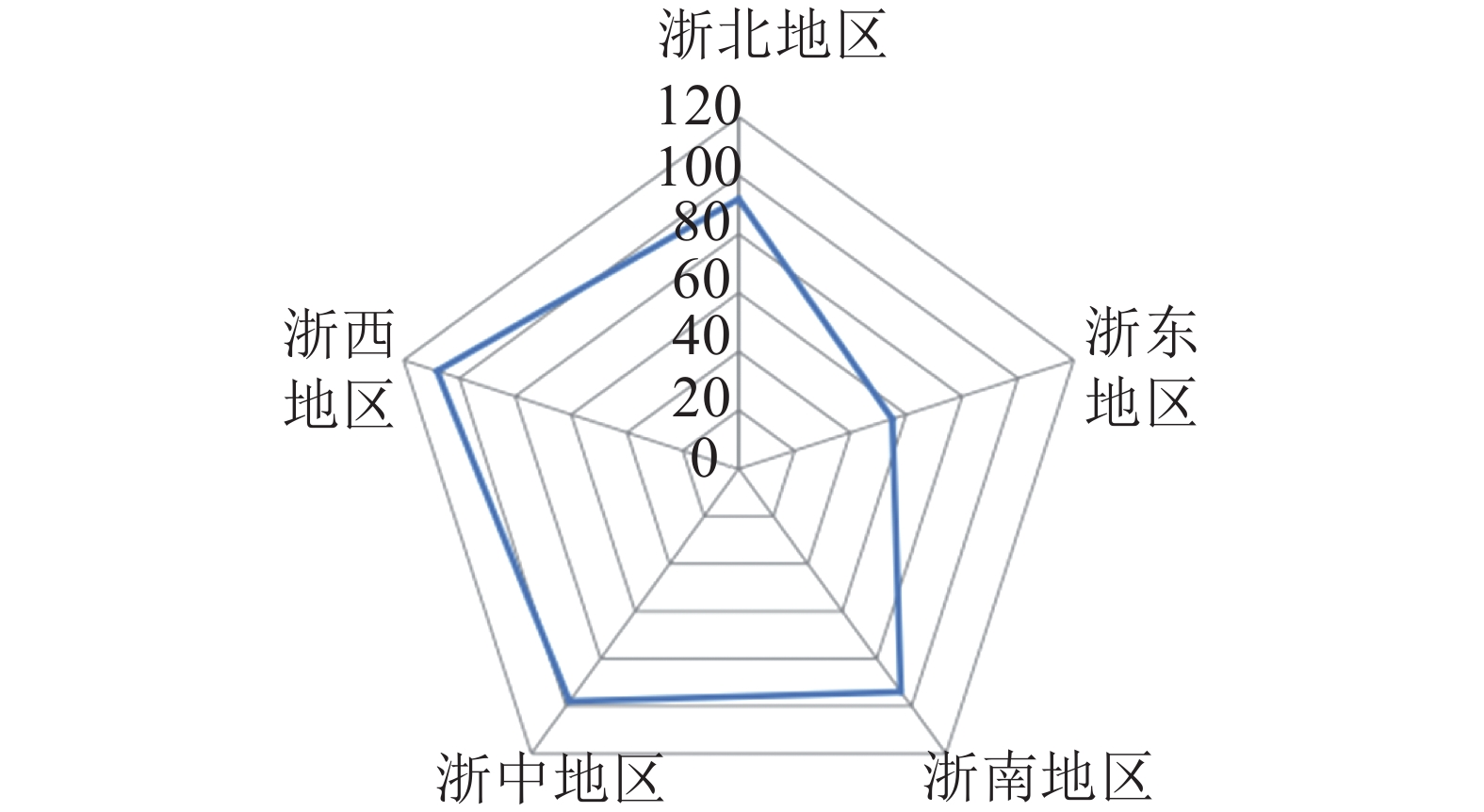
根据式(1)计算得到空间基尼系数(Gini)为0.969,代表森林乡村在浙江省市级行政区域的空间分布差异显著,并呈现明显的集中性。将浙江省划分为5个地区,即浙北(杭州、嘉兴、湖州),浙东(宁波、舟山),浙南(温州、台州),浙中(金华、绍兴),浙西(衢州、丽水)。由图1可知:森林乡村数量最多的是浙西地区,共有108个,占24.16%,其次是浙中、浙南地区,浙东地区数量最少,仅55个,占12.30%。此外,从绘制洛伦兹曲线(图2)可直观地看到弯曲程度较大,进一步验证了浙江省市域层面森林乡村空间分布的不均衡性。浙江省森林乡村的空间分布整体形成了以丽水(柯城)、绍兴(柯桥)、温州(乐清)为中心的多中心分布格局,在空间集聚上呈现团状集聚分布的特征。森林乡村数量分布大致呈现“西多东少”的格局,呈西南—东北向的分布态势,与浙江省自西南向东北呈阶梯状倾斜的基础地理格局十分契合。这代表森林乡村的发展与地形的联系紧密,西部地区以山地为主,森林覆盖率高,为森林乡村的形成及发展提供了丰富的资源。
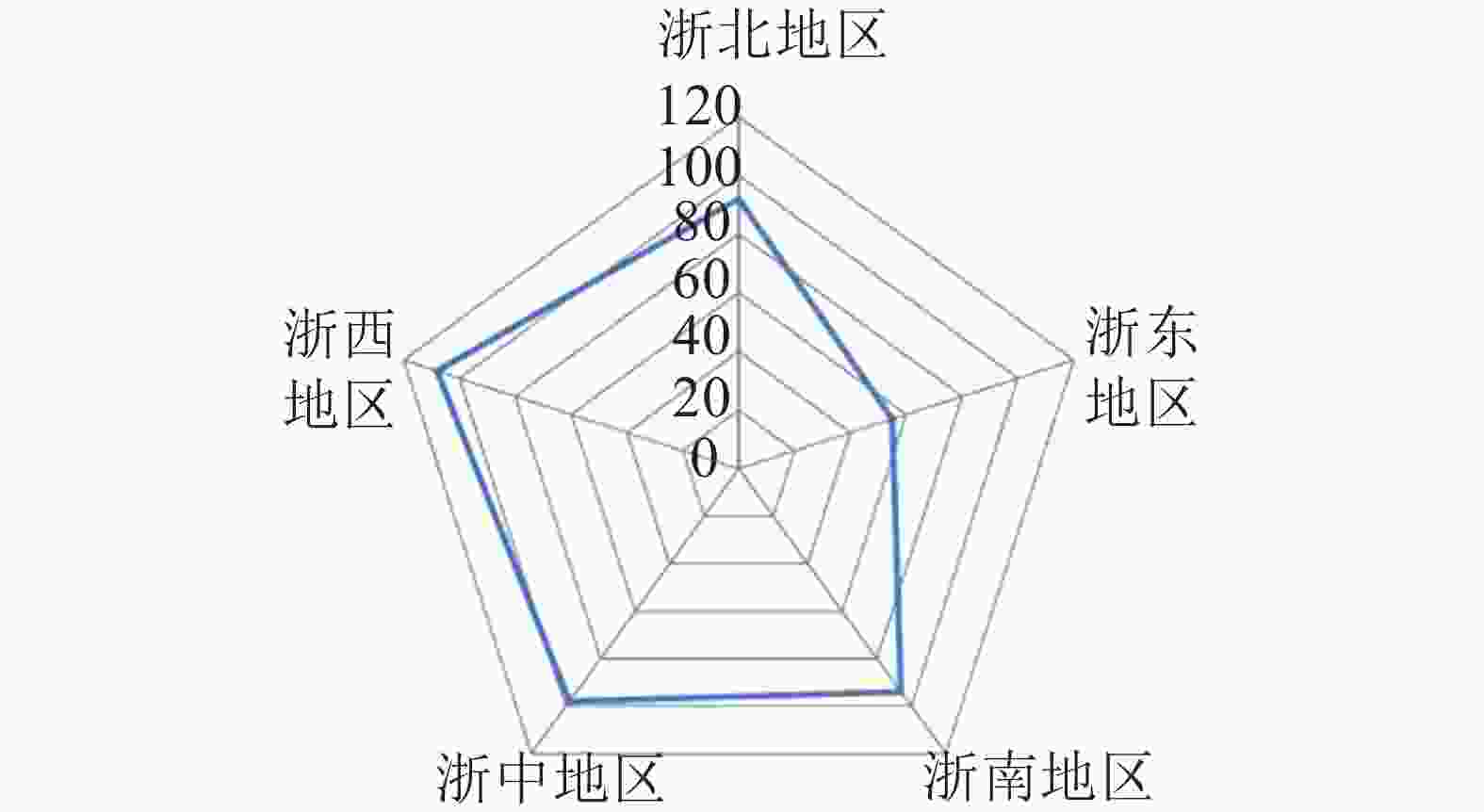
图 1 浙江省5个地区森林乡村数量统计
Figure 1. Number statistics of the forest villages in the five major divisionsin in Zhejiang Province
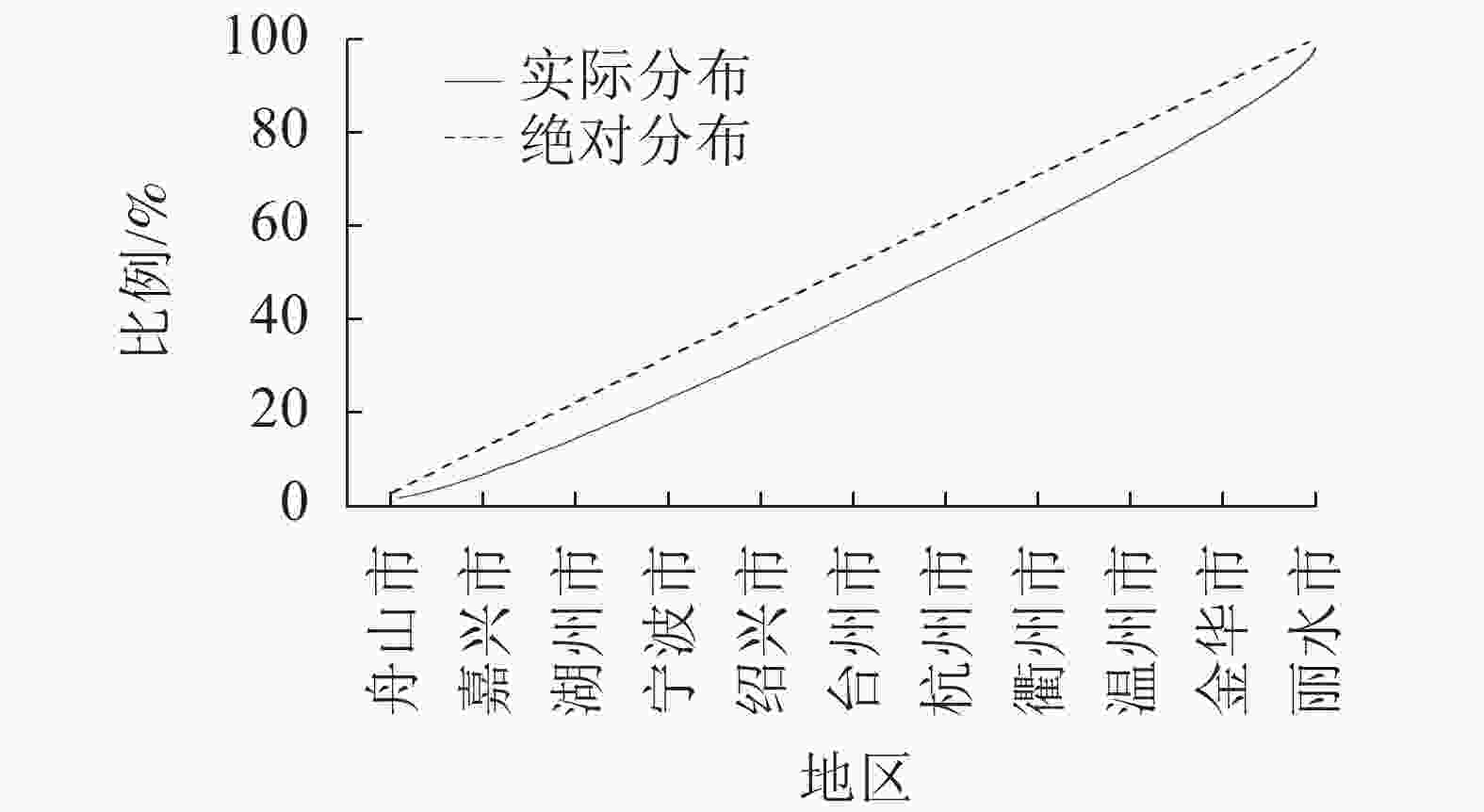
图 2 浙江省市域尺度森林乡村分布洛仑兹曲线
Figure 2. Lorenz curve of distribution of forest villages on the city scale in Zhejiang Province
-
图3显示:①第1批森林乡村呈现5个核心集聚区、4个核心连绵区的空间分布特征。5个集聚中心分别是衢州市中部(柯城区与常山县交界区域)、绍兴市东南部(柯桥、上虞、嵊州及新昌交界区)、宁波市东南部(余姚及海曙区、鄞州及奉化区交界区域)、温州市西部(瑞安、平阳、文成县交界区域)、温州市北部(乐清县),而核心连绵区则围绕核心集聚区成包围状分布。②第2批森林乡村形成2个核心集聚区。2个核心集聚区为金华市南部(金东、武义及永康县交界区域)、温州市北部(乐清、永嘉县)与台州市南部(黄岩、路桥、温岭县的交界区域)。③从2批森林乡村的核密度分布来看,形成了双核心集聚区、双核心连绵区。2个集聚中心分别为绍兴南部(柯桥、嵊州、新昌县交界区域)、温州北部(永嘉、乐清县)与台州市南部(黄岩、路桥、温岭县的交界处区域)。2个核心连绵区分别是以绍兴为中心的东西向带状连绵区,以温州与台州交界区域为核心的团状连绵区。具体而言,森林乡村空间分布地域差异显著,呈现出多个层级明显的高密度区,且多分布在东部,具有“小范围聚集、大范围分散”的多中心组团分布特点。
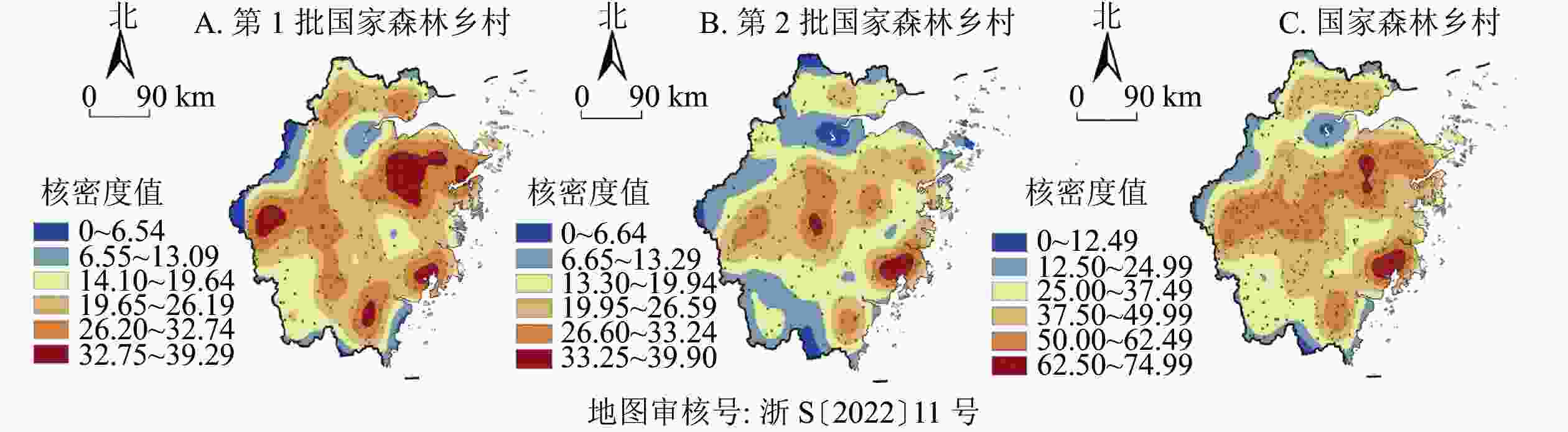
图 3 浙江省森林乡村核密度分布示意图
Figure 3. Kernel density estimation of forest villages in Zhejiang Province
-
由表1可见:第1批、第2批森林乡村空间自相关指数I估计值分别是0.398、0.710,正态统计值Z分别为5.447、9.608,均为正,说明2批森林乡村空间分布呈现显著的空间正相关性,不同区域森林乡村在空间上的集聚特征显著。
表 1 浙江森林乡村的全局空间自相关指数
Table 1. Global I index of forest villages in Zhejiang Province
森林乡村批次 I Z P 第2批森林乡村 0.398 3 5.447 0 0.000 0 第1批森林乡村 0.710 3 9.608 2 0.000 0 2批森林乡村 0.786 2 10.617 0 0.000 0 图4显示:①从空间分布形态来看,第1批森林乡村热点区呈现条带状分布特征,热点集聚区呈现“L”形分布形态,位于杭州西南部、衢州西南部及其与丽水北部的交界区域,丽水南部及会稽山脉东南部。第2批森林乡村热点区分布相对分散,呈半环状集聚特征,西部热点区呈现“C”形分布形态,集中分布于杭州西南方、衢州西南方及其与丽水北部的交界区域,东南部则呈现零星分布形态。②从冷热点数量来看,第2批森林乡村热点区有11个县级市,比第1批森林乡村多2个县级市。第2批的森林乡村次冷点区有11个县级市,比第1批森林乡村多6个县级市。热点区和次冷点区增加,说明森林乡村分布的高值、低值集聚性在加强。整体而言,浙江省森林乡村呈现西南部热,东北部冷的空间分布格局。热点区域多分布在历史文化底蕴较为丰富的浙西地区,并呈现条带状分布向条状及零星分布状转变的特征。此外,森林乡村分布高值集聚与低值集聚趋势逐渐明显,空间冷热格局渐进稳定。
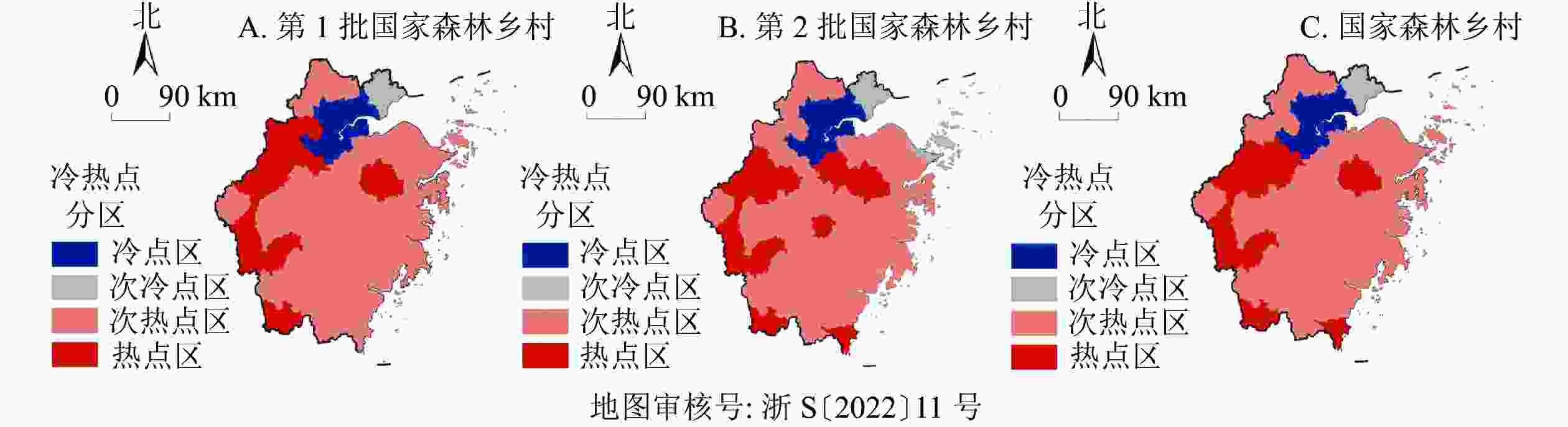
图 4 浙江省森林乡村冷热点空间分析示意图
Figure 4. Cold and hot spot analysis of forest villages in Zhejiang Province
-
地形地貌中海拔因素影响最明显。运用ArcGIS软件将森林乡村分布点与海拔高程地图进行缓冲区分析,结果显示:森林乡村的平均海拔为217 m,低于全省平均海拔307 m。86.35%的森林乡村分布在海拔500 m以下的山间盆地及丘陵地带,主要包括丽水境内的松古盆地,金华、衢州境内的金衢盆地以及这些盆地周边的丘陵地带。12.76%的森林乡村位于海拔500~1 000 m的低山区,主要包括丽水境内的仙霞岭、温州境内的洞宫山、金衢盆地周边的千里岗岭,仅0.89%的森林乡村分布在海拔高于1 000 m的山地。可见,森林乡村倾向于分布在地形起伏度较小的区域,空间分布上具有明显的“盆地丘陵”指向性特点。这与森林乡村的发展定位密不可分,一方面地形起伏度小的乡村便于科学规划,整合周边的森林公园等资源,打造休闲游憩的场所和乡村森林旅游的平台;另一方面,特殊的地形地貌有利于文化的形成[27]。浙西、浙中丘陵及浙南的山间盆地形成了相对封闭的地理环境,为森林乡村的历史文化遗存提供了有利条件,因此,地形地貌对森林乡村的空间分布具有显著影响。
-
将森林乡村分布点与年平均气温及年平均降雨量的地图进行叠加分析,结果显示:浙江省域范围内,森林乡村在年平均气温高且年平均降水量多的浙西地区分布较为密集,如衢州、丽水等地年平均气温较高,年降水量也较丰富,聚集了更多的森林乡村。浙江省年平均气温及年平均降水量的Pearson相关系数依次为0.209、0.134,影响力(q)依次为0.139、0.236,表明气候条件对森林乡村空间分布格局的影响有限,两者弱相关。
-
河流水系具有不同的形态及水体特性,对于增添森林乡村的旅游文化气息,营造舒适的观赏环境具有重要作用。将森林乡村分布点与河流水系地图进行缓冲区分析可知:森林乡村主要沿富春江、瓯江、衢江干支流分布,顺沿河道两侧成散点状分布。2.5 km缓冲区内的森林乡村达112个(25.06%),2.5~10.0 km缓冲区内森林乡村数量呈现减少趋势。浙江省各地水资源总量与森林乡村分布数量的Pearson相关系数为0.486,q为0.306,表明河流水系对森林乡村的空间分布格局具有一定影响。值得注意的是,尽管杭嘉湖平原河网密布,但平原地区多以耕地为主,所以森林资源低于同等水文状况的山地丘陵区。
-
丰富的森林资源能为以森林为题材的休闲、运动、康养等旅游项目提供场地与资源条件,推动森林产业发展,带动当地村民就业,实现脱贫致富。浙江省各县域森林覆盖率与森林乡村分布数量Pearson相关系数为0.732,q为0.555,表明森林覆盖率与森林乡村数量间具有较强的正相关。因此,对于依赖森林资源发展森林产业的森林乡村来说,森林覆盖率具有显著影响。
-
古村落聚集了历史文物、历史建筑等有形文化遗产[28],区域内非物质文化遗产及古村落数量越多,代表区域内文化资源越丰富[29]。对于以发展森林旅游及传播生态文化为目的的森林乡村来说,非物质文化遗产及古村落具有重要价值。本研究将浙江省内1092个古村落及2006—2021年入选的5批国家级非物质文化遗产 (226个)来衡量文化资源与森林乡村分布的关系。浙江省各县域古村落数量与森林乡村数量的Pearson 相关系数为0.764,q为0.770;国家级非物质文化遗产数量与森林乡村数量的Pearson 相关系数为0.529,q为0.413,表明古村落、国家级非物质文化遗产与森林乡村分布的相关性强。由此可见,森林乡村将文化资源赋存作为森林产业发展的重要参考因素,并试图与文化底蕴丰富的古村落建立关联。一方面,统筹推进森林乡村与周边古村落的关系,依托区域内古村落的基底,通过科学规划建设,整合周边资源,挖掘休闲、游览及体验价值,打造乡村森林旅游业新亮点;另一方面,合理利用非物质文化遗产资源举办森林文化活动,设计文创产品,对于传播生态文化知识具有重要意义。
-
人口密度、人均国内生产总值(GDP)、森林康养基地数量和第三产业增加值等4个经济社会指标对森林乡村空间分布的影响见表2。首先从呈正向相关的指标来看,各地的第三产业增加值与森林乡村的分布具有一定相关性,反映了森林乡村产业趋向于具有高附加值的森林康养、森林旅游等第三产业,旨在通过森林旅游带动当地经济的增长,取得经济效益与生态效益的双赢。各地的森林康养基地数量与森林乡村的分布相关性不显著,可见,目前的森林乡村周围没有形成一定规模的森林康养产业,森林康养产业发展建设尚处于发展阶段。其次,从异向相关的指标来看,各地的人口密度与森林乡村的分布具有较高的异向相关,表明在人口密度相对较小的地区森林乡村趋于集聚。这是由于人口密度大的地区,对农产品的消费需求也增大,从而驱使林地经营者转变土地利用方式,变林地为耕地,直接导致森林面积的减少[23],进而影响森林乡村的形成。此外,人均GDP与森林乡村具有较低的异向相关,反映了区域经济发展水平较高或较低的地区,森林乡村更为集聚。由于森林乡村在发展过程中,为了促进经济增长,容易过度依赖森林等自然资源,进而加速森林资源的消耗,反之当经济发展水平达到一定程度时,当地居民的环境保护意识及生态服务需求会提高,开始通过整合森林资源,发展高附加值的第三产业而拉动当地经济的增长,从而有利于森林乡村的保存与延续[23]。
表 2 森林乡村与社会经济指标的Pearson相关系数
Table 2. Pearson correlation coefficients between forest villages and socio-economic indicators
指标 Pearson相关系数 q 第三产业增加值 0.356** 0.151 森林康养基地数量 0.237 0.069 人口密度 −0.664 0.485 人均GDP −0.239* 0.143 说明:*P<0.05,**P<0.01 -
交通是人类活动突破地域空间限制的最佳途径[30]。交通可达性好能有效地促进城乡发展要素的流通,为森林乡村产业的发展提供极大便利[31]。由于区位布局的特殊性,公路是连接乡村聚落之间及聚落与外部联系的主要途径[32],因此,选取公路网密度来研究交通条件与森林乡村分布之间的关系。浙江省各县域公路网密度与森林乡村分布数量的Pearson相关系数为0.419,q为0.211,表明交通条件与森林乡村分布的具有一定的正相关性。交通条件一定程度上影响了森林乡村的布局。一方面,交通条件的改善促进了当地居民的对外交往与沟通,公路建设有利于提高农产品的生产效率,赋能当地的农业生产活动;另一方面,交通是森林乡村发展旅游业的先决条件,交通可达性直接影响森林旅游地对游客的吸引力。
-
空间分布特征上,浙江省森林乡村在空间上呈现集聚型分布状态,具有“小范围聚集,大范围分散”的多中心组团分布特点。区域空间分布上,在浙江省5个地区及市域层面上分布不均衡,整体呈现“西多东少”的分布格局。空间分布密度上,地域差异显著,形成了双核心集聚区、双核心连绵区。集聚中心为绍兴南部、温州北部与台州南部的交界处区域;核心连绵区包括以绍兴为中心的东西向带状连绵区、以温州台州交界区域为核心的团状连绵区。空间分布关联上,浙江省森林乡村空间分布呈现显著的空间正相关性,不同区域的森林乡村在空间上集聚特征显著。总体呈现中西南部热,东北部冷的空间分布格局,热点区呈现条带状分布向条状及零星状转变的特征,分布高值集聚与低值集聚趋势逐渐明显,空间冷热格局渐进稳定。
浙江省森林乡村的空间分布格局受自然和人文两大因素的综合影响。地形地貌、气候条件、河流水系、森林覆盖率是影响浙江省森林乡村空间分布的自然因素。地形地貌直接影响河流水系、道路交通、森林资源的布局及文化遗存,进而影响森林乡村的形成;气候并非森林乡村形成的必要条件,但其对森林植被生长的影响会间接影响森林乡村的布局;河流水系是森林乡村形成的重要条件,丰富的水资源是乡村居民生产生活的必要条件,同时为森林旅游产业的发展提供了丰富的景观资源;森林覆盖率是森林乡村评选的基础条件,丰富的森林资源能够为以森林为题材的休闲、运动、康养等产业的开展提供重要条件。文化资源、社会经济基础及交通通达性是影响森林乡村空间分布的人文因素。依托区域内的古村落及非物质文化遗产资源,森林乡村将文化资源赋存作为产业转型升级的关键因素;从社会经济因素来看,森林乡村多分布在人口密度较小、经济水平相对较低的地区,通过发展高附加值的第三产业实现经济效益与生态效益的双赢;交通可达性是森林乡村发展的先决条件,不仅赋予农业生产活动,而且有助于森林乡村聚点成线、整合资源、增强联系,促进全域发展。
-
浙江省开展森林乡村建设起到以点带面的效应,为生态文明建设提供了高质量的浙江样本。为促进森林乡村的发展,基于本研究结论,提出以下建议:①因地制宜,差异发展。森林乡村拥有丰富的森林资源及文化资源,一方面要充分利用森林资源,发展森林康养、森林旅游、林下经济等特色产业,另一方面要充分发挥不同村庄的文化特色,打造生态、文化、产业、经济、景观多功能融合的特色村庄,从而打通“绿水青山”向“金山银山”双向转换的通道,让村民享受到生态文明建设带来的绿色福利和生态红利,实现经济效益与生态效益的双提升。②协同布局,全域发展。目前浙江省森林乡村分布差异显著,因此,要加强不同区域之间的乡村规划引导,着眼于当下森林乡村的空间格局,结合村庄优势,合理调控资源。通过道路交通,增强村与村、县与县、市与市之间的联系,集中连片打造,以此构建全域共建、全域共融、全域共享的森林乡村发展新模式。
限于数据的可获取性,本研究未涉及政策制度等因素的定量化研究,因此,随着未来信息技术的发展及数据可获取性的增强,政策制度、森林生态效益等因素可以作为未来森林乡村空间分布影响因素分析的重点方向,进而完善森林乡村空间格局形成机制的研究,为森林乡村的保护及发展提供更为科学的理论指导与实践引导。
Spatial distribution characteristics and influencing factors of forest villages in Zhejiang Province
-
摘要:
目的 探讨森林乡村的空间分布特征及其影响因素,便于优化森林乡村的空间布局。正确认识森林乡村的形成因素,助推生态文明建设及乡村振兴。 方法 以浙江省447个森林乡村为研究对象,综合运用空间基尼系数、核密度分析、空间自相关等方法分析森林乡村的空间分布特征,并采用皮尔逊(Pearson)相关性分析、地理探测器等方法探究影响其空间分布的自然及人文因素。 结果 ① 从整体空间上看,浙江省森林乡村呈现集聚型的分布状态,具有“小范围聚集,大范围分散”的多中心组团分布特点。②从区域空间上看,在五大分区及市域层面上分布不均衡,整体呈现“西多东少”的分布格局。③从整体空间密度上看,形成了双核心集聚区和双核心连绵区。④从空间分布关联上看,呈现西南部热,东北部冷的空间关联特征,次冷点区域变化较小,热点区域呈现条带状分布向条状及零星分布状转变。⑤浙江省森林乡村的空间分布受地形地貌、气候条件、河流水系、森林资源、文化资源、社会经济基础、交通通达性的综合影响。 结论 由于自然及人文的综合影响,浙江省森林乡村的空间分布差异显著。森林乡村的建设与发展中,地形、气候及河流孕育其生态环境,而交通、文化资源及社会经济基础关乎其发展的基础条件、资源优势及经济支撑。今后浙江省在森林乡村建设中,要基于资源优势、基础条件及本地发展政策,坚持科学规划、因地制宜、协同布局等举措以实现全域发展,为生态文明建设提供高质量的浙江样板。图4表2参32 Abstract:Objective This study is to explore the spatial distribution characteristics and influencing factors of forest villages, so as to optimize the spatial layout and understand the forming factors, so as to comprehensively promote the construction of ecological civilization and rural revitalization. Method The spatial distribution characteristics of 447 forest villages in Zhejiang Province were analyzed using spatial Gini coefficient, nuclear density analysis and spatial autocorrelation, and the natural and human factors affecting their spatial distribution were explored by Pearson correlation analysis and geographic detectors. Result (1) The overall spatial distribution of forest villages was in an agglomerated distribution state, with the characteristics of multi-center clusters of “small-scale aggregation and large-scale dispersion”. (2) The regional spatial distribution was uneven at the level of five major districts and cities, displaying a distribution pattern of “more in the west and less in the east”. (3) In terms of overall spatial density, a dual-core agglomeration area and a dual-core continuous area were formed. (4) In terms of spatial distribution correlation, it showed the spatial correlation characteristics of hot in the southwest and cold in the northeast. The change of sub-cold spot area was small, and the hot spot area changed from strip distribution to strip and sporadic distribution. (5) The spatial distribution of forest villages in Zhejiang Province was affected by topography, climate conditions, river systems, forest resources, cultural resources, socio-economic foundation and traffic accessibility. Conclusion Due to the comprehensive influence of nature and humanity, the spatial distribution of forest villages in Zhejiang Province is significantly different. Topography, climate and rivers nurture their ecological environment, while transportation, cultural resources and socio-economic foundations are related to basic conditions, resource advantages and economic support of their development. We should, according to resource advantages, basic conditions and local development policies, adhere to scientific planning, local conditions, coordinated layout and other measures so as to achieve global development, providing high-quality Zhejiang models for the construction of ecological civilization. [Ch, 4 fig. 2 tab. 32 ref.] -
Key words:
- forest village /
- geographic detector /
- spatial distribution /
- influencing factors /
- Zhejiang Province
-
图 1 浙江省5个地区森林乡村数量统计
Figure 1 Number statistics of the forest villages in the five major divisionsin in Zhejiang Province
图 2 浙江省市域尺度森林乡村分布洛仑兹曲线
Figure 2 Lorenz curve of distribution of forest villages on the city scale in Zhejiang Province
图 3 浙江省森林乡村核密度分布示意图
Figure 3 Kernel density estimation of forest villages in Zhejiang Province
图 4 浙江省森林乡村冷热点空间分析示意图
Figure 4 Cold and hot spot analysis of forest villages in Zhejiang Province
表 1 浙江森林乡村的全局空间自相关指数
Table 1. Global I index of forest villages in Zhejiang Province
森林乡村批次 I Z P 第2批森林乡村 0.398 3 5.447 0 0.000 0 第1批森林乡村 0.710 3 9.608 2 0.000 0 2批森林乡村 0.786 2 10.617 0 0.000 0  下载: 导出CSV
下载: 导出CSV
表 2 森林乡村与社会经济指标的Pearson相关系数
Table 2. Pearson correlation coefficients between forest villages and socio-economic indicators
指标 Pearson相关系数 q 第三产业增加值 0.356** 0.151 森林康养基地数量 0.237 0.069 人口密度 −0.664 0.485 人均GDP −0.239* 0.143 说明:*P<0.05,**P<0.01
下载: 导出CSV
-
[1] 罗文斌, 蒋理, 雷洁琼, 等. 湖南省国家森林乡村分布格局及影响因素研究[J/OL]. 中国农业资源与区划, 2021[2021-08-01]. https://kns.cnki.net/kcms/detail/11.3513.S.20210607.1046.008.html. LUO Wenbin, JIANG Li, LEI Jieqiong, et al. Research on the distribution pattern and influencing factors of national forest villages in Hunan Province[J/OL]. China Agric Resour Reg, 2021. [2021-08-01]. https://kns.cnki.net/kcms/detail/11.3513.S.20210607.1046.008.html. [2] 陈刚, 吴清, 杨俭波, 等. 中国国家森林乡村的空间分布特征与影响因素[J]. 经济地理, 2021, 41(6): 196 − 204. CHEN Gang, WU Qing, YANG Jianbo. Spatial distribution characteristics and influencing factors of China national forest villages [J]. Econ Geogr, 2021, 41(6): 196 − 204. [3] DÉMURGER S, YANG Weiyong. Economic changes and afforestation incentives in rural China [J]. Environ Dev Econ, 2006, 11(5): 629 − 649. [4] STEFANIE D, VINCENT U, DEIRDRE O, et al. Factors influencing Irish farmers’ afforestation intention [J]. For Policy Econ, 2014, 39(C): 13 − 20. [5] O’LEARY T N, MCCORMACK A G, CLINCH J P. Afforestation in Ireland: regional differences in attitude [J]. Land Use Policy, 2000, 17(1): 39 − 48. [6] 李霞, 余荣卓, 杨金林. 乡村旅游中森林元素的挖掘与开发利用——以南平市乡村旅游为例[J]. 武夷学院学报, 2019, 38(10): 42 − 46. LI Xia, YU Rongzhuo, YANG Jinlin. Exploitation and utilization of forest elements in rural tourism:taking nanping rural tourism as an example [J]. J Wuyi Univ, 2019, 38(10): 42 − 46. [7] 丁宇, 刘正刚, 王卫江, 等. 森林旅游区乡村旅游对社区居民生活质量的影响——基于乌鲁木齐南山林区方家庄的实证分析[J]. 中南林业科技大学学报(社会科学版), 2016, 10(4): 53 − 58. DING Yu, LIU Zhenggang, WANG Weijiang, et al. Rural tourist influence on the living quality of forest tourism area residents: taking fang village of urumqi nanshan forest zone as an example [J]. J Cent South Univ For Technol Soc Sci Ed, 2016, 10(4): 53 − 58. [8] KASSIOUMIS K, PAPAGEORGIOU K, CHRISTODOULOU A, et al. Rural development by afforestation in predominantly agricultural areas: issues and challenges from two areas in Greece [J]. For Policy Econ, 2004, 6(5): 483 − 496. [9] APPIAH D O, BUGRI J T, FORKUO E K, et al. Agricultural and forest land use potential for REDD+ among smallholder land users in rural Ghana [J]. Int J For Res, 2016(2): 1 − 7. [10] TAHVANAINEN L, TYRVÄINEN L, NOUSIAINEN I. Effect of afforestation on the scenic value of rural landscape [J]. Scand J For Res, 1996, 11(1): 397 − 405. [11] MÜLLER D, MUNROE D K. Changing rural landscapes in Albania: cropland abandonment and forest clearing in the postsocialist transition [J]. Ann Assoc Am Geogr, 2008, 98(4): 855 − 876. [12] BEDRU B, BART M, FREDU N, et al. The economic contribution of forest resource use to rural livelihoods in Tigray, Northern Ethiopia [J]. For Policy Econ, 2009, 11(2): 122 − 131. [13] VEDELD P, ANGELSEN A, BOJÖ J, et al. Forest environmental incomes and the rural poor [J]. For Policy Econ, 2007, 9(7): 869 − 879. [14] CALLERY J J, SANANN N, TRIPURA R, et al. Engaging ethnic minority communities through performance and arts: health education in Cambodian forest villages [J]. Int Health, 2021, 13(2): 188 − 195. [15] 杨燕, 胡静, 李亚娟, 等. 基于GIS的中国森林乡村空间结构及影响因素研究[J]. 干旱区资源与环境, 2021, 35(6): 182 − 191. YANG Yan, HU Jing, LI Yajuan, et al. Spatial structure of forest villages in china and influencing factors [J]. Arid Land Resour Environ, 2021, 35(6): 182 − 191. [16] 张典铨. 福鼎市国家森林乡村森林绿地建设现状调查[J]. 热带农业工程, 2020, 44(6): 75 − 77. ZHANG Dianquan. Investigation on the current situation of rural forest greenspace construction of national forest in fuding city [J]. Trop Agric Eng, 2020, 44(6): 75 − 77. [17] 王洪桥, 袁家冬, 孟祥君. 东北地区A级旅游景区空间分布特征及影响因素[J]. 地理科学, 2017, 37(6): 895 − 903. WANG Hongqiao, YUAN Jiadong, MENG Xiangjun. Spatial distribution and its influencing factors of level: a scenic spots in Northeast China [J]. Sci Geogr Sin, 2017, 37(6): 895 − 903. [18] 周扬, 黄晗, 刘彦随. 中国村庄空间分布规律及其影响因素[J]. 地理学报, 2020, 75(10): 2206 − 2223. ZHOU Yang, HUANG Han, LIU Yansui. The spatial distribution characteristics and influencing factors of chinese villages [J]. Acta Geogr Sin, 2020, 75(10): 2206 − 2223. [19] 国家林业和草原局. 国家森林乡村评定办法(暂行)[EB/OL]. 2019-09-05[2021-07-10]. http://www. zjly. gov. cn/art/2019/9/6/art_1346870_385. html. State Forestry and Grassland Administration. National Forestry and Rural Assessment Measures (Interim) [EB/OL]. 2019-09-05[2021-07-10]. http://www. zjly. gov. cn/art/2019/9/6/art_1346870_385. html. [20] 国志兴, 王宗明, 宋开山, 等. 1982—2003年东北林区森林植被NDVI与水热条件的相关分析[J]. 生态学杂志, 2007, 26(12): 1930 − 1936. GUO Zhixing, WANG Zongming, SONG Kaishan, et al. Correlations between forest vegetation NDVI and water/thermal condition in northeast China forest regions in 1982−2003 [J]. Chin J Ecol, 2007, 26(12): 1930 − 1936. [21] 陈国磊, 罗静, 曾菊新, 等. 中国少数民族特色村寨空间结构识别及影响机理研究[J]. 地理科学, 2018, 38(9): 1422 − 1429. CHEN Guolei, LUO Jing, ZENG Juxin, et al. Spatial structure identification and influence mechanism of ethnic villages in China [J]. Sci Geogr Sin, 2018, 38(9): 1422 − 1429. [22] 俞飞, 李智勇. 天目山森林文化价值的游客感知分析[J]. 浙江农林大学学报, 2020, 37(2): 366 − 373. YU Fei, LI Zhiyong. Analysis of tourists’ perception of forest cultural value in Mount Tianmu [J]. J Zhejiang A&F Univ, 2020, 37(2): 366 − 373. [23] 李凌超, 邓丹丹, 张鼎文, 等. 京津冀森林破碎化的社会经济影响因素分析[J]. 林业经济, 2021, 43(4): 5 − 16. LI Lingchao, DENG Dandan, ZHANG Dingwen, et al. Analysis on socio-economic determinants of forest fragmentation in Beijing-Tianjin-Hebei region [J]. For Econ, 2021, 43(4): 5 − 16. [24] 葛扬, 张建国. 浙江省森林特色小镇空间分布特征及影响因素分析[J]. 浙江农林大学学报, 2020, 37(2): 374 − 381. GE Yang, ZHANG Jianguo. Spatial distribution features and influence factors of forest towns in Zhejiang Province [J]. J Zhejiang A&F Univ, 2020, 37(2): 374 − 381. [25] 姜磊, 陈星宇, 朱竑. 中国城市养老院的空间分布特征及其分异成因[J]. 地理学报, 2021, 76(8): 1951 − 1964. JIANG Lei, CHEN Xingyu, ZHU Hong. The spatial heterogeneity distribution of chinese urban nursing homes and socio-economic driving factors [J]. Acta Geogr Sin, 2021, 76(8): 1951 − 1964. [26] 王劲峰, 徐成东. 地理探测器: 原理与展望[J]. 地理学报, 2017, 72(1): 116 − 134. WANG Jinfeng, XU Chengdong. Geodetector: principles and prospects [J]. Acta Geogr Sin, 2017, 72(1): 116 − 134. [27] 徐柏翠, 潘竟虎. 中国国家级非物质文化遗产的空间分布特征及影响因素[J]. 经济地理, 2018, 38(5): 188 − 196. XU Baicui, PAN Jinghu. Spatial distribution characteristics of the intangible Cultural Heritage in China [J]. Econ Geogr, 2018, 38(5): 188 − 196. [28] 龚胜生, 李孜沫, 胡娟, 等. 山西省古村落的空间分布与演化研究[J]. 地理科学, 2017, 37(3): 416 − 425. GONG Shengsheng, LI Zimo, HU Juan, et al. The spatial distribution and evolution of ancient villages in Shanxi Province [J]. Sci Geogr Sin, 2017, 37(3): 416 − 425. [29] 顾康康, 储金龙, 汪勇政, 等. 黄山市古村落综合品质空间分异——基于101个古村落的实证调研[J]. 地理研究, 2014, 33(11): 2034 − 2042. GU Kangkang, CHU Jinlong, WANG Yongzheng, et al. Spatial differentiation of comprehensive quality for ancient villages in Huangshan city: based on empirical investigationof 101 ancient villages [J]. Geogr Res, 2014, 33(11): 2034 − 2042. [30] 李久林, 储金龙, 李瑶. 古徽州传统村落空间分布格局及保护发展研究[J]. 中国农业资源与区划, 2019, 40(10): 101 − 109. LI Jiulin, CHU Jinlong, LI Yao. Research on the spatial distribution pattern and protection and development of ancient Huizhou traditional villages [J]. China Agric Resour Reg, 2019, 40(10): 101 − 109. [31] 焦胜, 郑志明, 徐峰, 等. 传统村落分布的“边缘化”特征——以湖南省为例[J]. 地理研究, 2016, 35(8): 1525 − 1534. JIAO Sheng, ZHENG Zhiming, XU Feng, et al. The marginalization tendency of the traditional village distribution: the case study of Hunan Province [J]. Geogr Res, 2016, 35(8): 1525 − 1534. [32] 黄洋, 卢海霞, 苟锐. 长江经济带全国森林康养示范基地空间分异[J]. 江苏农业科学, 2021, 49(14): 242 −248. HUANG Yang, LU Haixia, GOU Rui. Spatial differentiation of national forest health demonstration bases in the Yangtze River Economic zone[J]. Jiangsu Agric Sci, 2021, 49(14): 242 −248. -

-
链接本文:
https://zlxb.zafu.edu.cn/article/doi/10.11833/j.issn.2095-0756.20210558
 点击查看大图
点击查看大图
计量
- 文章访问数: 1773
- HTML全文浏览量: 425
- PDF下载量: 22
- 被引次数: 0











