




-
GATA转录因子是一类进化上保守的转录调节因子,广泛存在于真核生物中,能特异性识别并结合目标基因启动子区域的共有序列(A/T)GATA(A/G)[1]。植物中第1个GATA基因NTL1是从烟草Nicotiana tabacum中分离得到的,为神经孢子虫Neurospora crassa中NTL2的同源物[2]。植物中大多数GATA转录因子只包含1个CX2CX18CX2C锌指结构域,只有少数成员包含1个CX2CX20CX2C结构域或2个以上的锌指基序[3]。 GATA转录因子可根据其结构域的数量和排序方式进一步分为多个亚家族,例如,在拟南芥Arabidopsis thaliana中共鉴定出30个GATA家族成员,划分为4个亚家族[4]。此外,在水稻Oryza sativa[5]、烟草[6]和花生Arachis hypogaea[7]等植物中也分别鉴定出28、57和47个GATA家族成员,但有关青钱柳Cyclocarya paliurus GATA转录因子的研究还未见报道。
GATA 因子在植物中的生物学功能已得到广泛研究。多项研究表明:GATAs作为调控因子参与植物的多种发育过程,如种子萌发、叶绿体发育、光合响应、碳氮代谢以及次生壁组分生物合成[8−10]。近年来,多个GATA家族成员已被证明在非生物胁迫抗逆性中发挥作用。研究表明:OsGATA16可以通过与OsWRKY45-1的启动子特异性结合,抑制OsWRKY45-1的表达,增强水稻苗期的耐寒性[11]。杨树Populus GATA家族成员PdGNC1在干旱胁迫下通过与PdHKX1启动子结合,调控一氧化氮(NO)和过氧化氢(H2O2)的生成,减弱气孔关闭[12]。过表达IbGATA24可增强激素信号通路相关信号传导和活性氧清除,IbGATA24与IbCOP9-5a相互作用可正调控甘薯Dioscorea esculenta的耐旱性和耐盐性[13]。
青钱柳是中国特有的单属种植物,其叶片中富含黄酮、三萜和多糖等多种生物活性物质,具有降血糖、降血压和抗菌等功效[14]。青钱柳具有重要的经济和生态价值,主要分布在中国亚热带地区的山区溪谷等地,目前的种质资源难以满足市场叶用需求。为推动青钱柳产业的健康发展,将滨海盐碱地等受盐胁迫影响的区域纳入青钱柳人工林的潜在种植范围,已成为可行的解决方案。研究发现:盐胁迫显著抑制青钱柳的光合作用和碳氮代谢,并在分子水平上影响青钱柳的转录和代谢过程[15]。GATA等转录因子调节青钱柳盐胁迫响应的研究尚未见报道,亟待进一步研究为精准提升青钱柳耐盐性提供可能。
本研究利用生物信息学方法对青钱柳GATA基因家族进行成员鉴定,分析其理化性质、基因结构、系统发育和顺式调节元件等,并通过不同浓度的盐胁迫处理解析青钱柳GATA基因家族成员表达的盐胁迫响应模式,旨在为阐明青钱柳在盐胁迫下的分子应答机制和耐盐能力提供理论数据。
-
实验材料为青钱柳JZS30#家系。选取长势均一的当年生实生苗,在南京林业大学白马科研基地温室(温度为25~30 ℃,光照时长为16 h,相对湿度为60%~80%)进行盐胁迫水培处理试验。设置4种氯化钠(NaCl)处理浓度:对照(ck,0 mmol·L−1 NaCl)、低盐浓度(LS,25.7 mmol·L−1NaCl)、中盐浓度(MS,51.3 mmol·L−1NaCl)和高盐浓度(HS,77.0 mmol·L−1NaCl)。水培采用1/2强度的霍格兰营养液(pH 6.0±0.2),气泵连续通气,隔7 d更换1次。每个处理设置3个重复,分别在盐处理15 d (T1)和30 d (T2)采取植株中部完全展开叶片,液氮速冻后−80 ℃储存样本,用于后续转录组测序及实时荧光定量PCR (RT-qPCR)分析。
-
下载美国国家生物技术信息中心(NCBI)数据库中青钱柳二倍体雌先型基因组和注释文件[16],以TAIR数据库中拟南芥GATA基因序列为比对材料。通过BLAST本地搜索和HMMER程序搜索2种方法进行筛选,并比对到Swissport数据库获得青钱柳GATA家族成员和蛋白序列。随后将青钱柳GATA蛋白序列提交到在线工具ProtParam (
https://web.expasy.org/protparam/ ),用以预测氨基酸数目、等电点、不稳定指数和亲水性等理化性质,并通过WolfPsorf在线工具(https://wolfpsort.hgc.jp/ )进行亚细胞定位预测。 -
通过MEGA 11[17]软件对拟南芥、小麦Triticum aestivum、番薯Ipomoea batatas和青钱柳GATA蛋白序列进行比对分析,然后使用相邻连接法和Bootstrap方法进行系统进化树构建。使用在线工具NCBI中的Batch CD-Search功能结合TBtools软件[18]的MEME分析进行青钱柳GATA蛋白序列的保守结构域预测和保守基序鉴定。通过PlantCARE[19]数据库(
https://bioinformatics.psb.ugent.be/webtools/plantcare/html/ )对青钱柳GATA基因序列进行顺式调控元件分析,并通过TBtools软件进行可视化。 -
采用TBtools软件提取青钱柳GATA家族成员在基因组中的位置信息,并结合青钱柳基因组注释文件可视化GATA基因在染色体上的位置图谱。通过TBtools软件中的MCScanX功能进行共线性分析。
-
对样本进行RNA提取、cDNA文库构建和测序(原始数据已上传至NCBI数据库,提取号为PRJNA700136),原始数据通过Fastp (Version 0.18.0,HaploX Biotechnology)过滤,样本映射读数使用StringTie v1.3.1 (约翰斯霍普金斯大学计算生物学中心)组装[20]。StringTie软件(max_memory,30G;seqType,fq;CPU,10;KMER_SIZE,31;min_kmer_cov,9;normalize_reads; normalize_max_read_cov,50)计算FPKM值,并采用TBtools软件可视化青钱柳GATA家族基因在不同盐浓度处理下的表达模式。
-
选取盐胁迫30 d时9个差异表达GATA基因进行RT-qPCR分析。使用MonScript RTIII All-in-One Mix with dsDNase试剂盒(莫纳生物)获得cDNA,使用SYBR Green Realtime PCR Master Mix 试剂盒 (东洋纺株式会社)进行RT-qPCR反应。以18S rRNA作为内参基因[16],引物详细信息见表1。采用2−∆∆Ct 方法计算相对表达量,3次技术重复。
表 1 RT-qPCR引物序列
Table 1. Primer sequences for RT-qPCR
引物 正向引物序列(5′→ 3′) 反向引物序列(5′→ 3′) 18S rRNA AGTATGGTCGCAAGGCTGAAA CAGACAAATCGCTCCACCAA CpaF1st40248 GCTCAGAACACGGAAGTGGA CTGGCCTATACTCTGGCAGC CpaF1st03085 CAGGCCGACGAGGATAGTTC TGGCTGCATCTACGCTGAAA CpaF1st33138 CCGGAACCGGAACTGGTAAA CAAAAACTGGACACCGCAGG CpaF1st12063 CAGGCAGTGCTTCTACTGCT CTGGAGTGTCGGGCTCATTT CpaF1st29899 TGTCGGCACAAGATGAGGTC CCCAGTCTCCCCATTTGCAT CpaF1st24236 CCCAATTCTCCCCATCTCCG TAGGACCCTTGCTTCCCTGT CpaF1st06121 ACAACTGTGGTGGTGCTTCA TTGGTTCGCCTTCTCTGGAC CpaF1st13794 AGCGAGAGCAGAGAGTGAGA GTCCACAAGCCATACGACCA CpaF1st41514 ATGGGGAAGCATGAAGGGTG TTCACCTTGCCTGGATGGAC -
基于BLAST和HMM筛选对青钱柳基因组序列进行比对分析,在全基因组水平上发现46个青钱柳GATA家族成员。蛋白性质分析结果(表2)显示:GATA家族成员蛋白分子量为14 358.49~92 724.86 Da,编码蛋白长度为132~827个氨基酸;等电点(pI)为4.85~10.40,其中19个成员呈碱性(pI>7),其余均为酸性(pI<7);亲水性系数为−1.119~−0.368,成员的亲水性值均小于0。此外,亚细胞定位显示:37个GATA基因定位于细胞核,其余9个基因预测分布于叶绿体、细胞质和质膜中。
表 2 青钱柳GATA家族理化性质分析
Table 2. Physical and chemical properties analysis of GATA family in C. paliurus
基因 编码蛋白长度/个氨基酸 分子量/Da 等电点 不稳定指数 脂肪族指数 亲水性 基因位置 亚细胞定位 CpaF1st31008 350 38 871.14 5.42 55.22 58.54 −0.718 Chr5:3275133-3276675 细胞核 CpaF1st04551 333 36 772.69 6.27 59.72 55.98 −0.759 Chr10:5255396-5257097 细胞核 CpaF1st40248 278 31 171.34 8.85 61.17 68.71 −0.711 Chr7:36854287-36855776 细胞核 CpaF1st11493 259 29 016.11 6.52 50.25 54.67 −0.789 Chr12:27234110-27235170 细胞核 CpaF1st03085 399 44 049.62 5.58 59.07 50.63 −0.728 Chr1:39570084-39572566 细胞核 CpaF1st12368 440 48 903.86 6.51 50.23 63.36 −0.672 Chr13:4422154-4428720 细胞核 CpaF1st33138 376 41 285.27 6.53 57.64 55.74 −0.639 Chr5:28272282-28274034 细胞核 CpaF1st15405 304 33 337.68 8.77 59.30 65.72 −0.519 Chr14:14310065-14314628 细胞核 CpaF1st44867 381 41 982.50 5.30 64.62 52.23 −0.708 Chr9:24293467-24295185 细胞核 CpaF1st04260 337 36 733.24 5.96 55.24 65.76 −0.519 Chr10:2743778-2747468 细胞核 CpaF1st10391 827 92 724.86 8.74 43.31 77.56 −0.368 Chr12:16098279-16120607 质膜 CpaF1st33607 162 17 768.52 9.88 49.03 68.09 −0.560 Chr5:34149029-34150449 细胞核 CpaF1st05998 132 14 358.49 9.68 57.91 64.32 −0.765 Chr10:23155264-23156372 细胞核 CpaF1st45364 141 15 497.20 10.40 52.67 68.58 −0.482 Chr9:31189984-31191453 细胞核 CpaF1st05992 151 16 553.39 9.76 48.39 73.71 −0.430 Chr10:23093430-23094145 细胞核 CpaF1st42300 249 27 444.28 8.34 63.80 48.23 −0.743 Chr8:29402271-29403166 细胞核 CpaF1st42347 250 27 563.34 8.06 64.29 47.28 −0.764 Chr8:29941694-29942592 细胞核 CpaF1st06487 252 27 884.50 8.03 60.42 37.22 −0.907 Chr10:29829715-29830640 细胞核 CpaF1st24908 352 39 131.69 8.82 53.42 61.59 −0.726 Chr3:6660741-6662851 细胞核 CpaF1st14436 225 24 929.32 9.19 42.21 39.51 −1.119 Chr14:936552-939513 细胞核 CpaF1st12063 364 40 120.77 4.85 48.65 70.47 −0.665 Chr13:1849772-1855913 细胞核 CpaF1st31471 359 38 841.07 4.97 48.63 67.33 −0.658 Chr5:7431224-7439528 叶绿体 CpaF1st21387 327 36 490.18 9.69 50.48 57.92 −0.869 Chr2:9770484-9772235 细胞核 CpaF1st19401 528 58 511.73 6.11 55.39 67.54 −0.564 Chr16:14302411-14309576 细胞核 CpaF1st18106 258 28 349.77 9.05 56.55 58.64 −0.799 Chr15:20131363-20133090 细胞核 CpaF1st29466 542 60 103.26 5.92 55.85 70.30 −0.580 Chr4:23938169-23945173 细胞核 CpaF1st12064 310 33 115.02 5.12 42.23 73.61 −0.423 Chr13:1856825-1860692 叶绿体 CpaF1st03558 308 32 874.44 4.89 41.13 64.29 −0.542 Chr1:43510795-43514419 叶绿体 CpaF1st43529 305 32 777.21 5.77 32.70 65.28 −0.595 Chr9:5950153-5957707 细胞核 CpaF1st31469 304 32 856.19 5.83 38.43 60.63 −0.670 Chr5:7423788-7429644 细胞核 CpaF1st29899 541 60 796.46 5.96 47.87 74.23 −0.659 Chr4:30906208-30913091 细胞核 CpaF1st30660 336 36 905.18 6.21 44.02 66.19 −0.471 Chr5:498151-499582 叶绿体 CpaF1st42916 345 37 921.05 6.13 38.43 59.39 −0.623 Chr9:535642-537262 细胞质 CpaF1st18811 538 60 555.96 5.97 45.12 70.63 −0.672 Chr16:5904043-5913138 细胞核 CpaF1st24236 373 41 270.78 5.30 41.12 63.86 −0.564 Chr2:43555546-43557671 细胞核 CpaF1st15509 775 84 488.54 6.27 41.70 64.62 −0.815 Chr14:15829121-15836785 叶绿体 CpaF1st31203 373 40 721.49 5.86 45.42 60.46 −0.397 Chr5:4827679-4829481 叶绿体 CpaF1st00992 667 75 052.44 7.08 70.50 61.56 −0.838 Chr1:12126306-12130501 细胞核 CpaF1st06121 654 72 697.62 6.00 60.97 66.36 −0.722 Chr10:24970094-24974866 细胞核 CpaF1st13794 708 78 850.38 7.29 64.92 68.04 −0.702 Chr13:22615455-22620576 细胞核 CpaF1st41514 701 77 486.71 6.69 52.86 70.67 −0.644 Chr8:18653928-18659825 细胞核 -
为解析青钱柳GATA基因家族成员之间的发育进化关系,将鉴定到的46个青钱柳GATA基因与拟南芥和甘薯GATA基因进行多序列比对并构建进化树(图1)。青钱柳GATA家族成员被分为4个亚家族,其中Class Ⅱ和Class Ⅲ亚家族分别包含6和16个家族成员,Class Ⅰ和Class Ⅳ亚家族均包含12个成员。基于系统发育分析结果,可以推测同一亚家族内的基因成员在进化上具有较近的亲缘关系,可能承担相似的生物学功能。
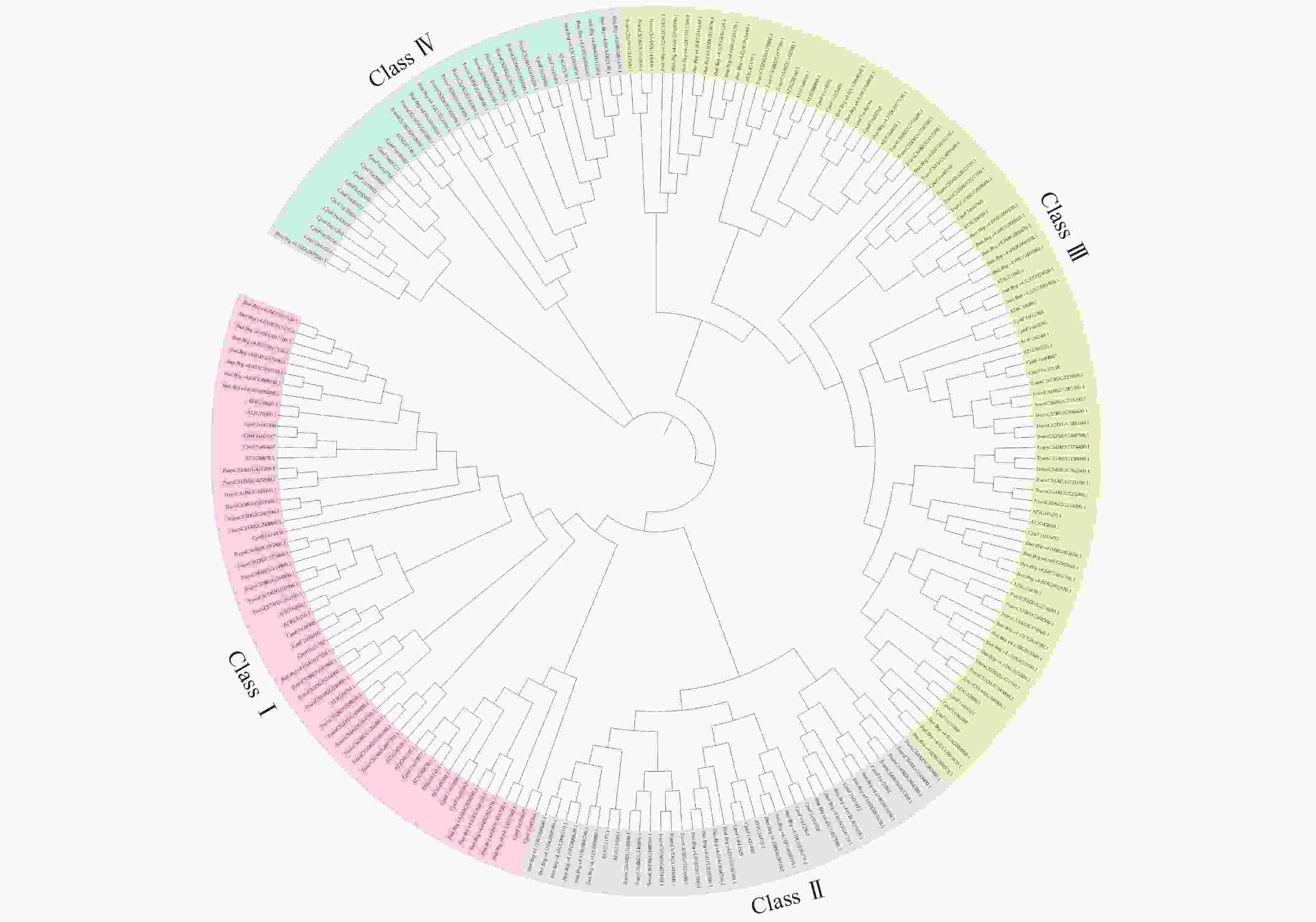
图 1 青钱柳 GATA 基因家族系成员统发育分析
Figure 1. Phylogenetic analysis of GATA gene family in C. paliurus
-
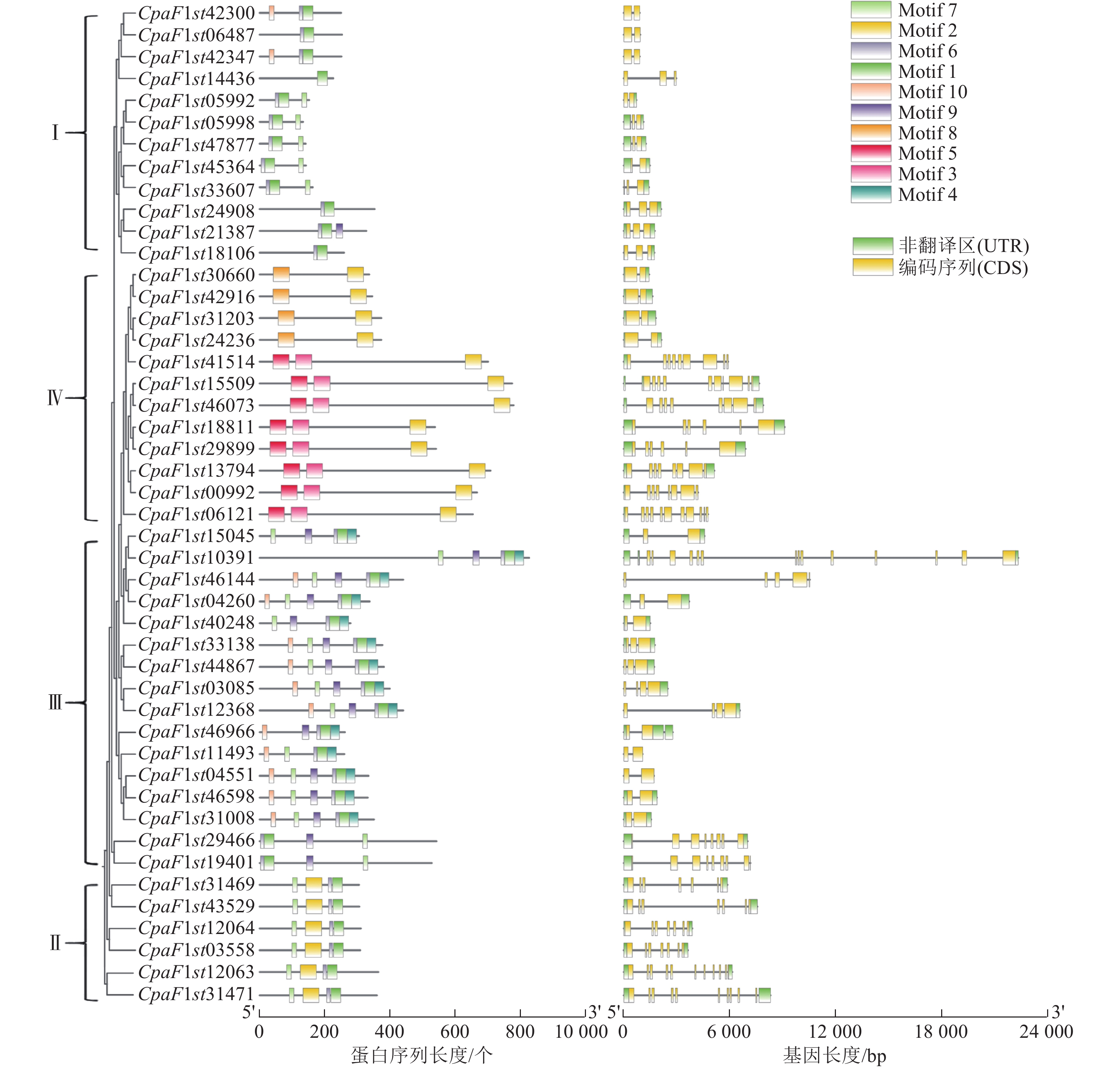
GATA家族成员基因结构的多样性影响外显子和内含子组成的差异性,可能造成基因功能的分化。对青钱柳GATA家族进行基因结构分析发现(图2):Class Ⅰ、Class Ⅱ 及 Class Ⅲ 亚家族内CpGATA基因结构较为类似,而各亚家族间基因结构差异明显。Class Ⅲ 亚家族内CpaF1st10391基因结构特异于其他基因,包含内含子序列最长,而CpaF1st11493基因长度最短,仅包含2个外显子和1个内含子;Class Ⅰ亚家族基因结构相似且长度相对较短,均包含1~2个内含子以及2~3个外显子,Class Ⅱ 和 Class Ⅳ 亚家族基因均包含较多的外显子和内含子数量。
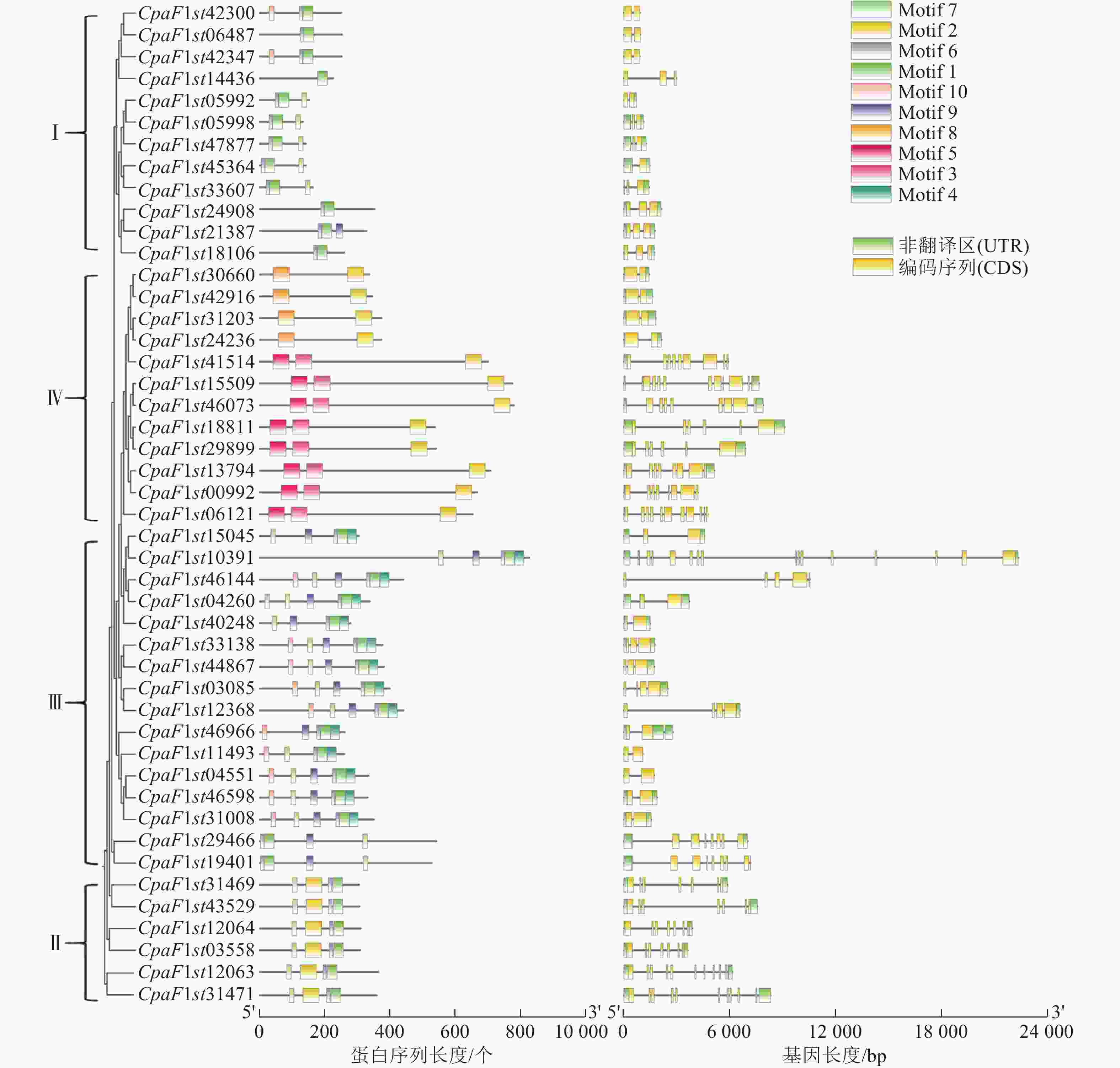
图 2 青钱柳 GATA 家族保守基序分析和基因结构
Figure 2. Motif analysis and gene structure of GATA family in C. paliurus
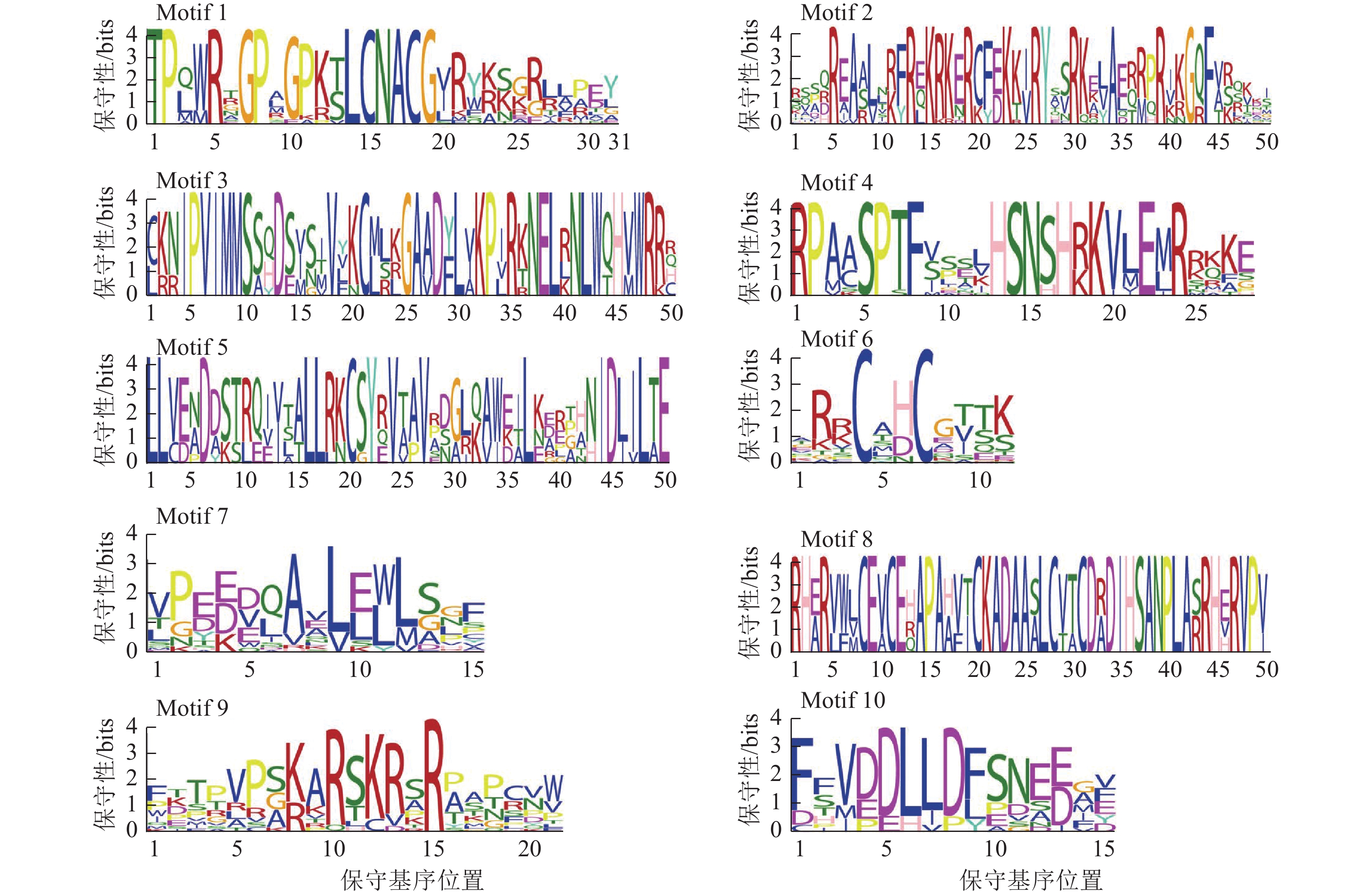
为了深入解析青钱柳GATA家族的蛋白结构及其在转录调控中的潜在功能,本研究采用MEME工具对CpGATA基因进行保守序列预测分析,最终鉴定了10个保守基序(图3)。由图2可知:同一亚家族内成员包含相似的保守基序,Class Ⅲ亚家族成员含有保守基序种类最多,数量为4~6个,均包含Motif 1、Motif 6、Motif 7和Motif 9。Class Ⅱ 亚家族的6个GATA成员包含的保守基序数量和相对位置最为一致,均包含有Motif 1、Motif 2、Motif 6和Motif 7。Class Ⅳ亚家族包含的保守基序数量较少,其中Motif 3和Motif 5为该亚族特有的保守基序。
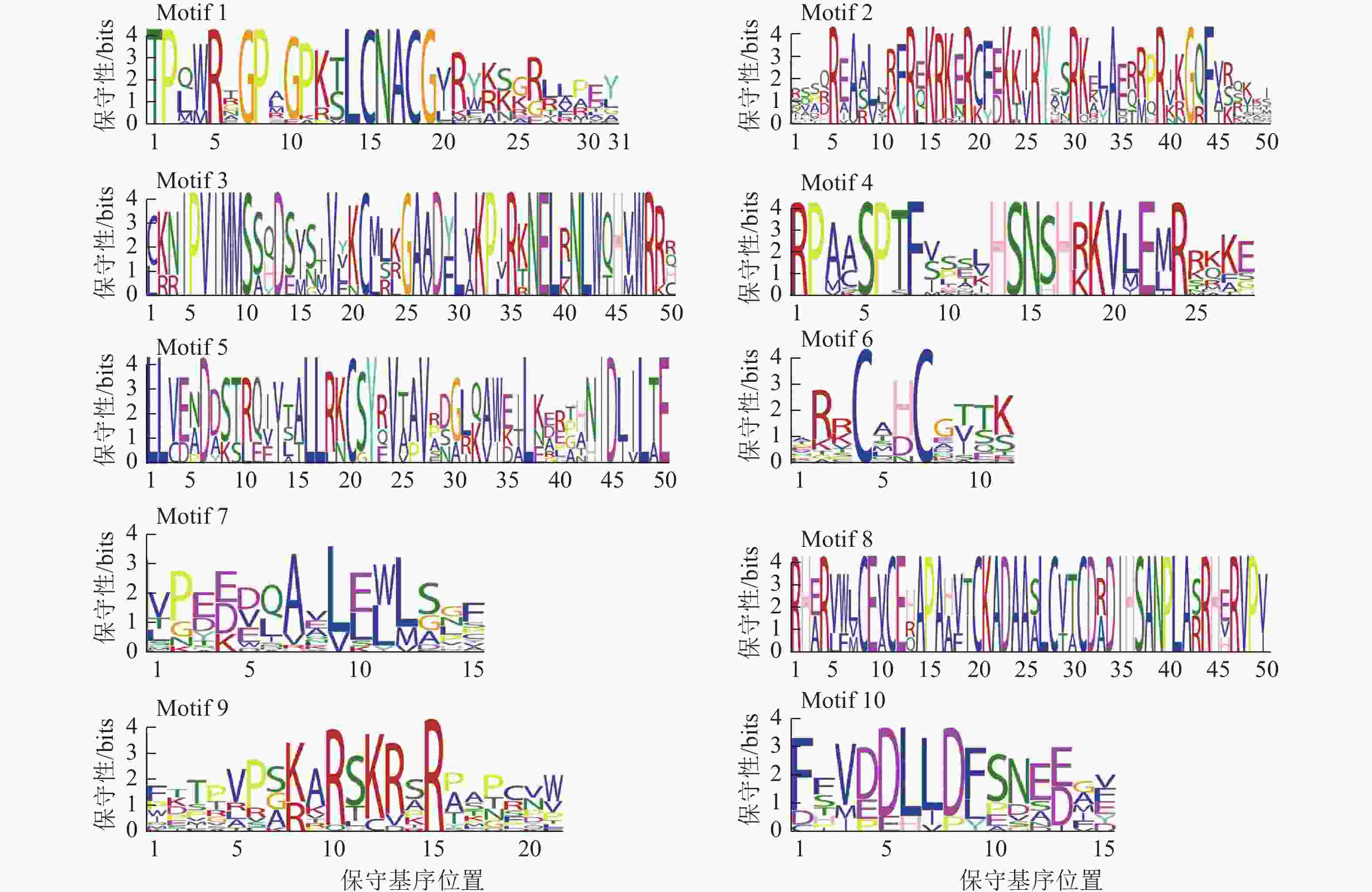
图 3 青钱柳 GATA 家族基因序列
Figure 3. Gene motifs of GATA family in C. paliurus
-
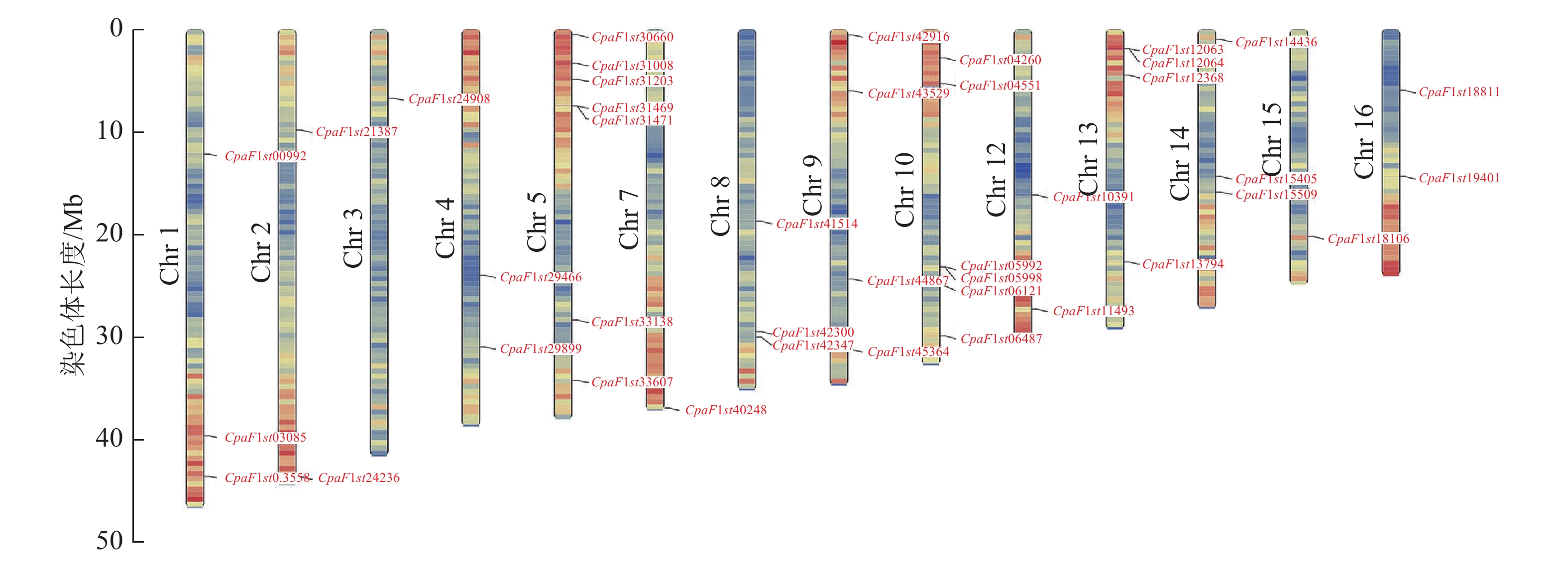
染色体定位分析显示:青钱柳GATA家族基因分布于除Chr 6和Chr 11外的14条染色体上(图4)。Chr 5和Chr 10染色体上分布CpGATA基因数量最多,分别有7和6条;而Chr 2、Chr 3、Chr 8和Chr 15染色体上分布最少,均含有1条CpGATA基因。此外,Chr 1染色体长度最长,而仅含有CpaF1st00992、CpaF1st03085和CpaF1st03558 3条CpGATA基因,表明青钱柳GATA家族基因在染色体上呈不均匀分布状态。
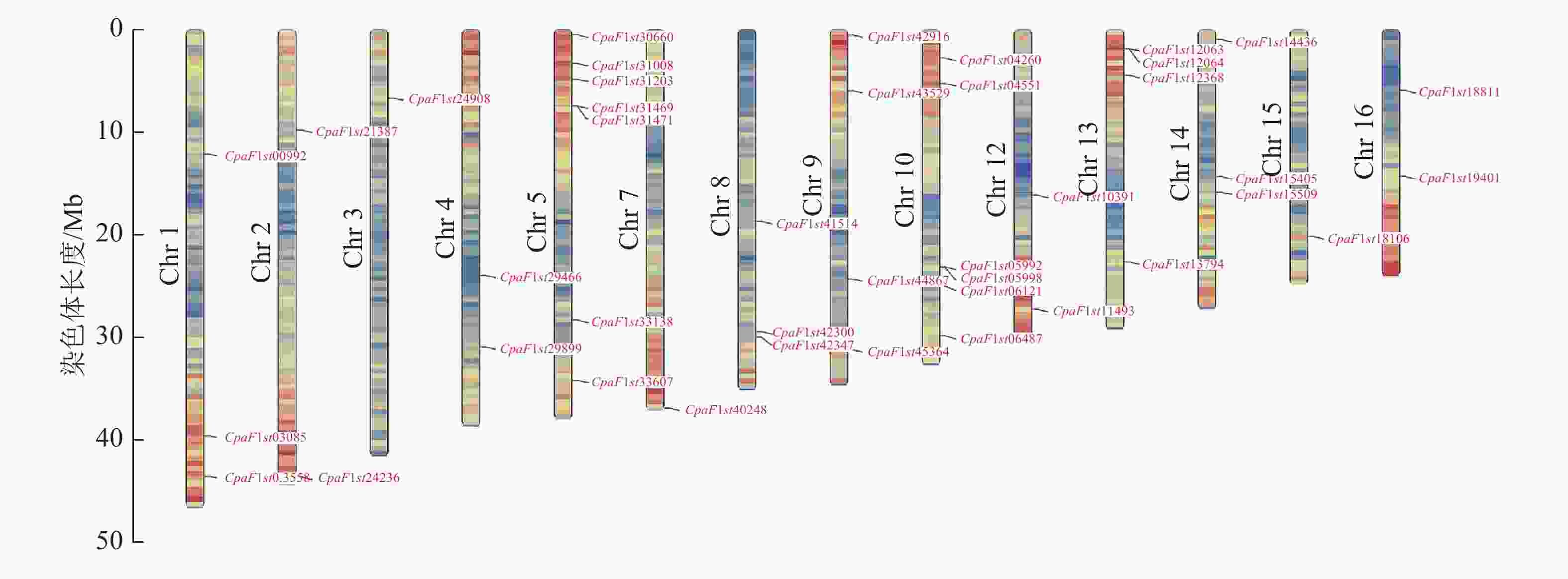
图 4 青钱柳 GATA 家族染色体分布
Figure 4. Chromosome locations of GATA family in C. paliurus
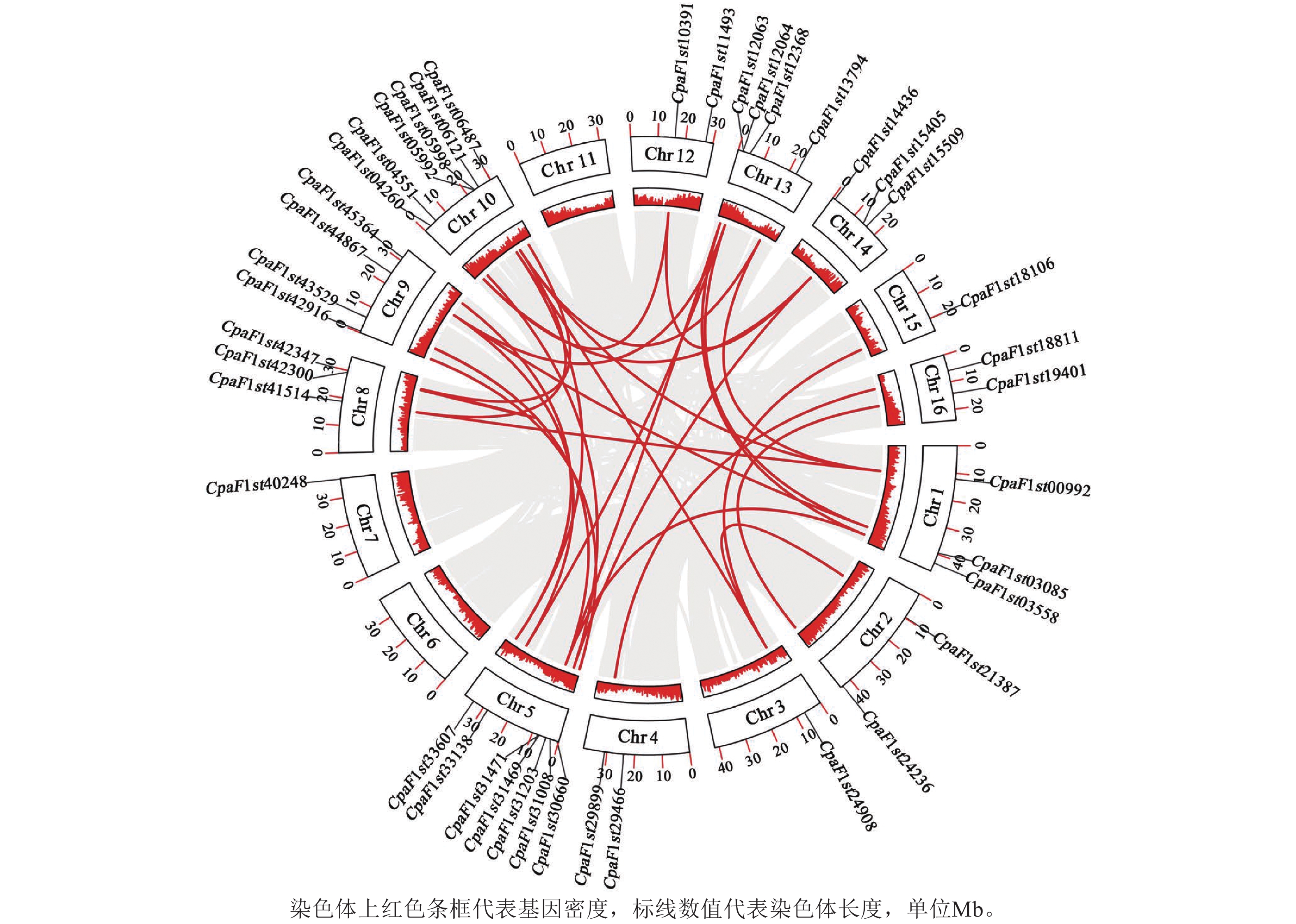
为进一步解析青钱柳GATA基因家族的进化关系,通过MCScanX对家族成员进行共线性分析(图5),共鉴定出33个共线性基因对。其中Chr 5和Chr 10染色体上CpGATA基因相关性较强,各包含10个共线性基因对;而Chr 2、Chr 15和Chr 16染色体上CpGATA共线性基因对较少,共包含5对线性关系。上述结果表明:Chr 5和Chr 10染色体上CpGATA遗传关系更为紧密,在基因功能上可能更为相似。
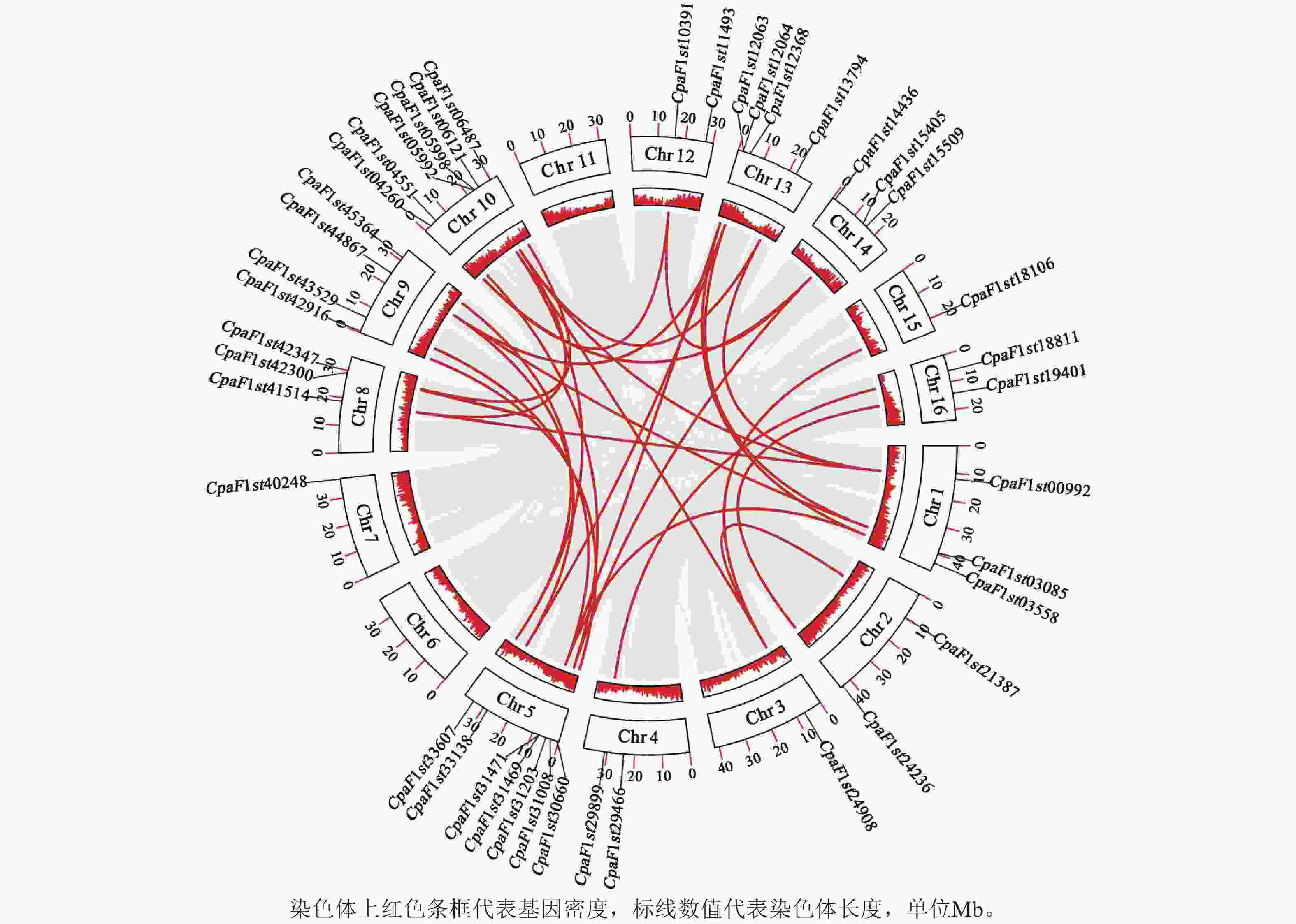
图 5 青钱柳 GATA 基因家族共线性分析
Figure 5. Collinear analysis of GATA gene family in C. paliurus
-
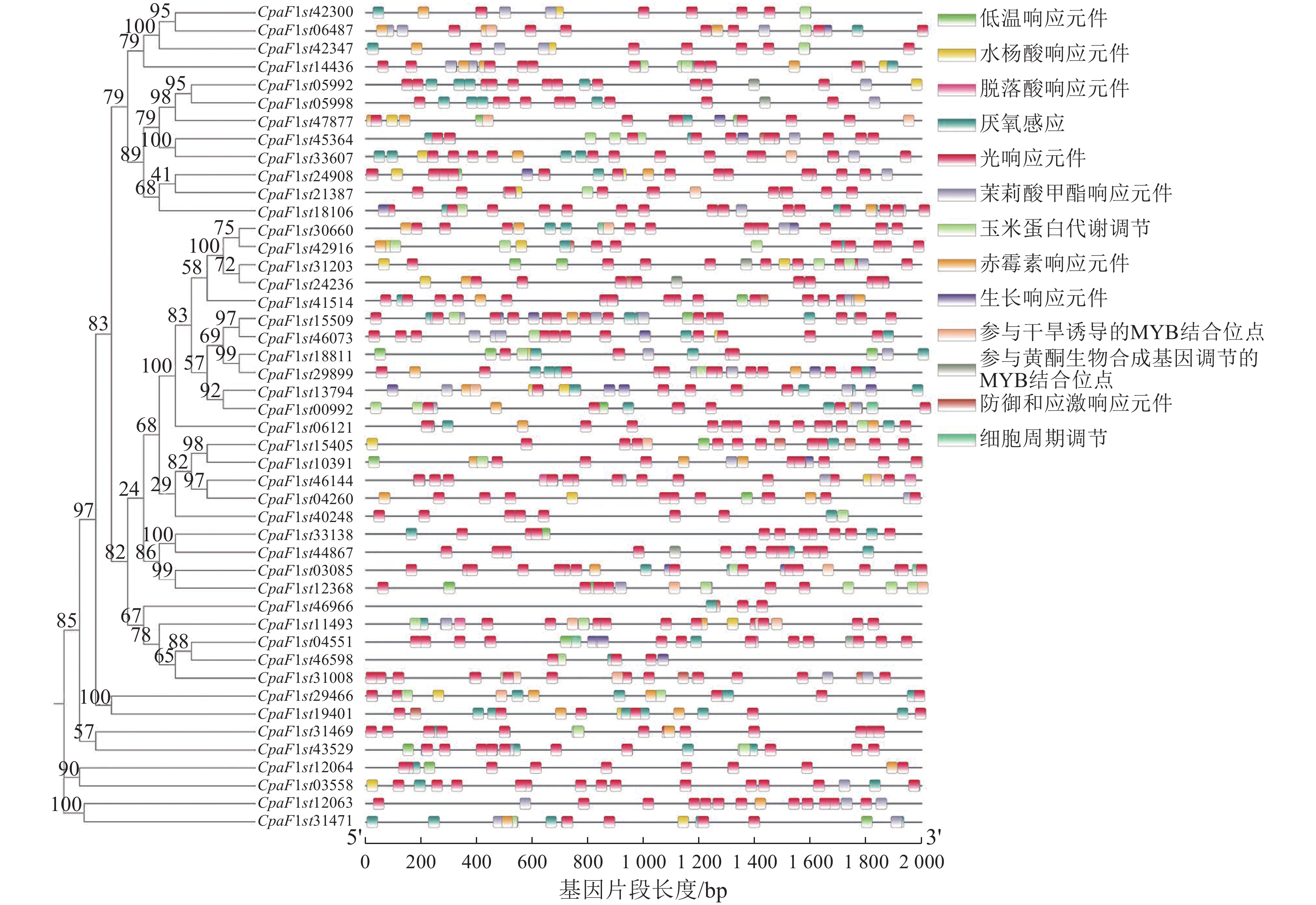
对青钱柳GATA家族成员上游2 000 bp的基因片段进行顺式作用元件分析,在46个青钱柳GATA家族成员中共鉴定出13种响应元件(图6)。大部分GATA家族成员启动子区域包含光响应、脱落酸响应和厌氧感应等多种响应元件。在近半数的CpGATA基因启动子区发现植物激素调节元件,包括赤霉素响应、水杨酸响应和茉莉酸甲酯响应元件。此外,16个成员含有低温响应元件,10个成员含有防御和应激响应元件,6个成员含有参与黄酮生物合成基因调节的MYB结合位点。以上结果表明:青钱柳GATA家族基因可能在各类非生物胁迫响应中发挥关键作用。
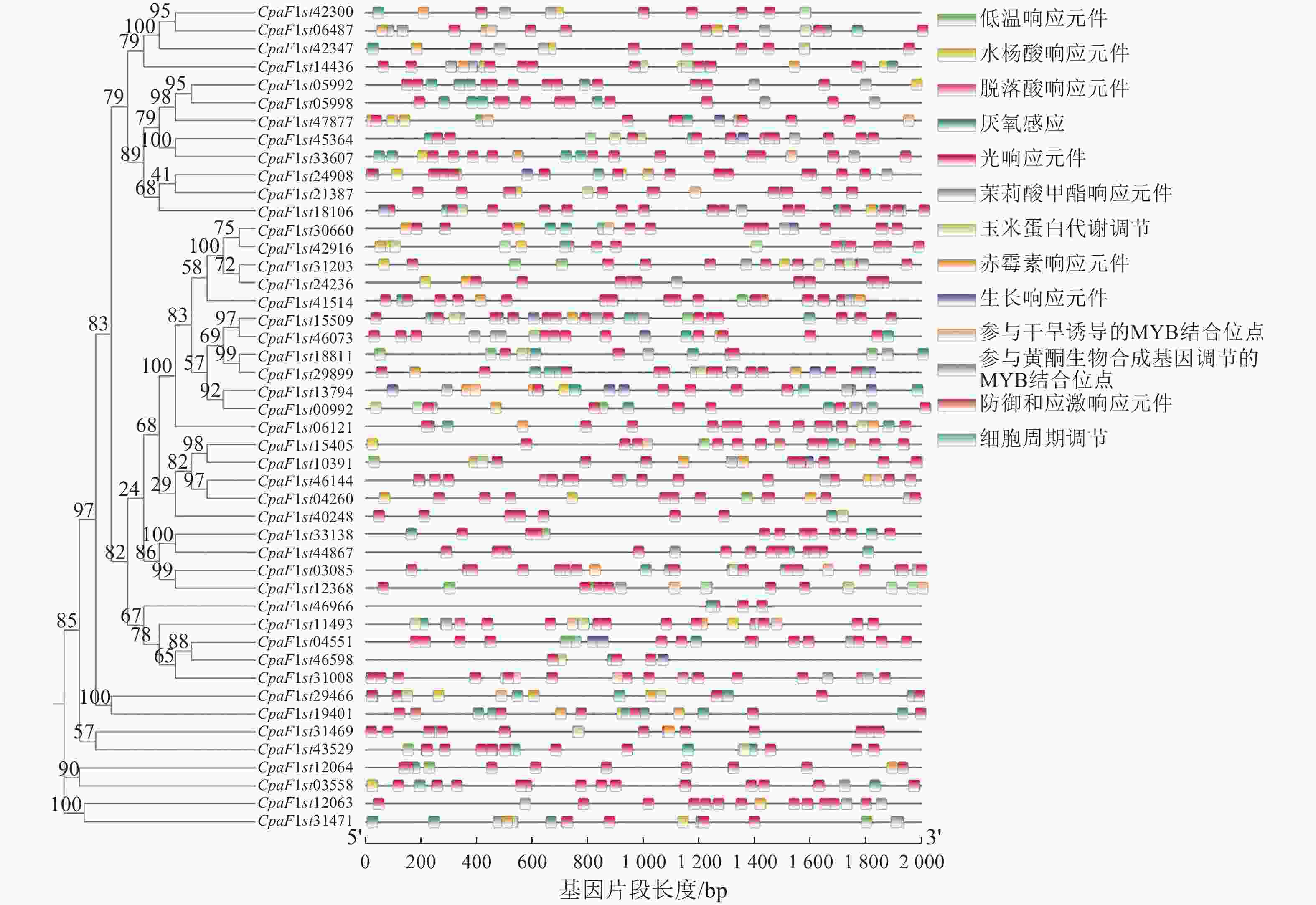
图 6 青钱柳GATA基因家族顺式作用元件分析
Figure 6. Analysis of cis-regulatory elements of GATA gene family in C. paliurus
-
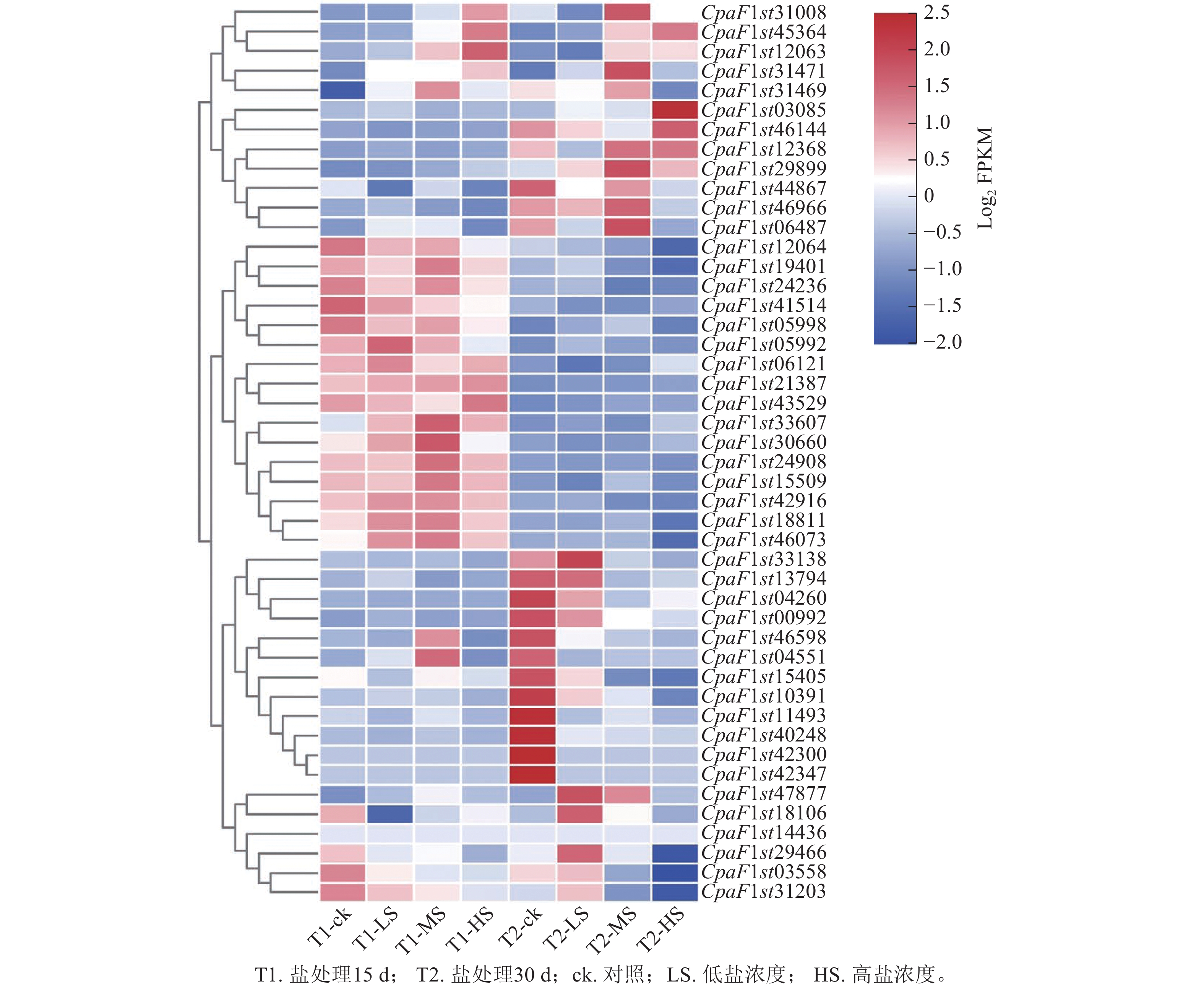
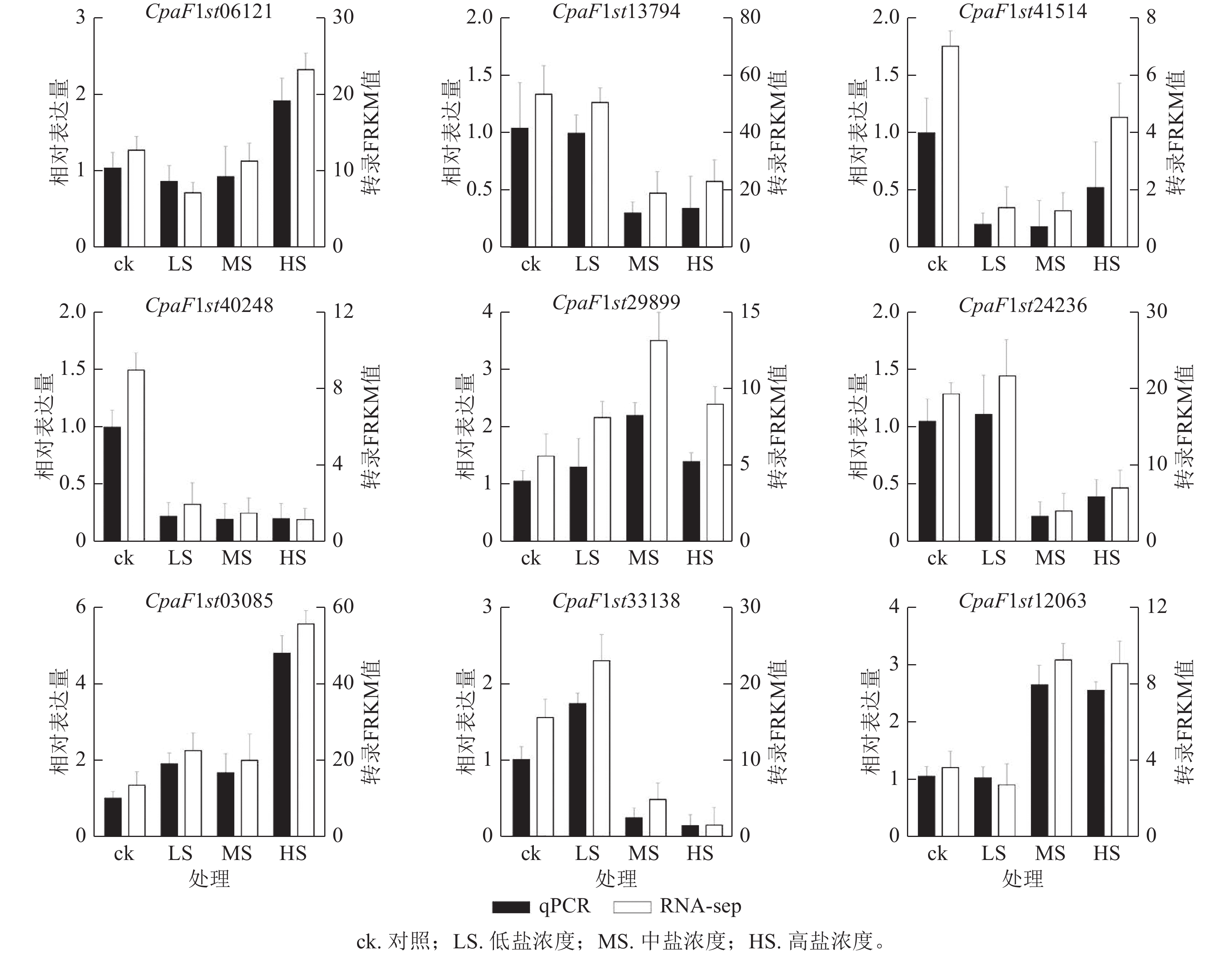
为探究青钱柳GATA基因家族在盐胁迫响应中的调控作用,本研究通过不同盐浓度处理和不同处理时长的转录组数据分析其基因表达模式(图7)。由聚类分析可知:在盐胁迫处理30 d时,CpaF1st31008、CpaF1st45364和CpaF1st12063等12个基因的相对表达量高于15 d,且盐胁迫诱导其表达量上调。在盐处理15 d时,CpaF1st12064、CpaF1st19401和CpaF1st24236等16个基因的相对表达量显著高于30 d时。此外,CpaF1st33138、CpaF1st13794和CpaF1st04260等18个基因表达量受盐胁迫处理抑制而显著下调。上述结果表明:青钱柳GATA家族成员的相对表达量在盐胁迫处理下呈多样化趋势,推测其在青钱柳应对盐胁迫的过程中可能发挥不同的调控功能。此外,通过RT-qPCR发现:青钱柳GATA基因在盐胁迫下表达趋势与转录分析表现相一致,进一步验证了RNA测序结果可信(图8)。
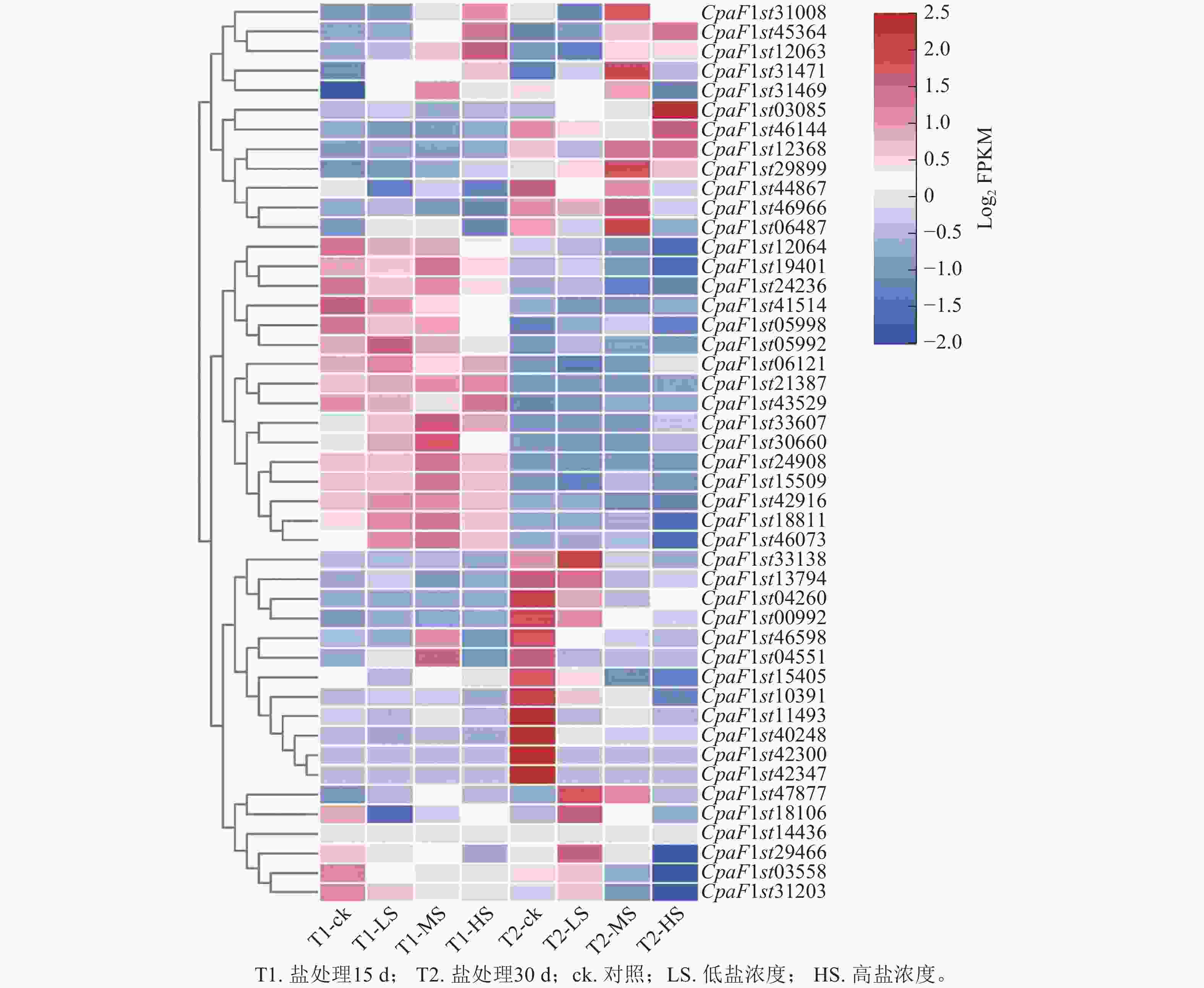
图 7 青钱柳GATA基因家族在盐胁迫下的表达模式
Figure 7. Expression patterns of GATA gene family in C. paliurus under salt stress
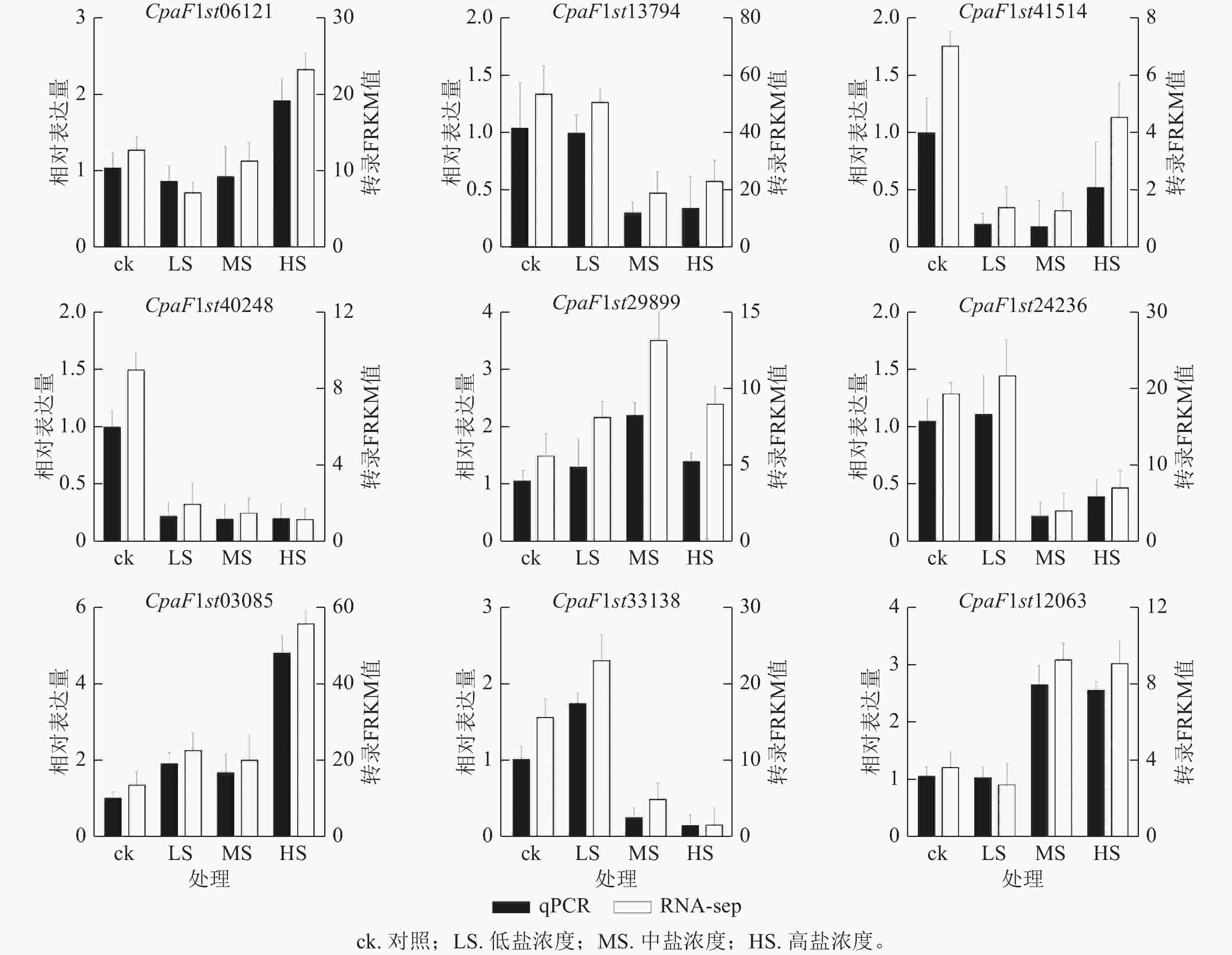
图 8 盐处理30 d青钱柳GATA基因表达的RT-qPCR验证
Figure 8. RT-qPCR validation of GATA genes expression under salt stress for 30 days
-
GATA转录因子作为锌指蛋白转录因子的重要成员之一,不仅参与植株生长发育和碳氮代谢调控等次生代谢过程,在植物的非生物胁迫响应中也发挥重要作用[21]。据研究报道,GATA通过介导植物体内活性氧清除、叶绿素生物合成以及植物激素信号转导等生物过程调节植株的盐胁迫耐受性[22]。目前,已有研究在转录和代谢水平上解析青钱柳对盐胁迫的响应模式[23],但GATA等转录因子在盐胁迫下的响应策略仍有待进一步系统分析。
本研究在全基因组水平对青钱柳GATA基因家族共鉴定出46个GATA家族成员,数量大于水稻(29个) [24]和辣椒Capsicum annuum (24个) [25],但少于烟草(57个) [6]。水稻和青钱柳染色体倍性为二倍体且基因组较小,辣椒基因组较大且富含转座子等重复序列,但基因数量较少,而烟草基因组庞大且为异源四倍体,推测GATA基因数量可能与物种染色体倍性及基因组大小存在正相关关系。亚细胞定位显示:青钱柳82.61%的GATA家族成员定位于细胞核,其余定位于叶绿体和细胞质,表明青钱柳GATA转录因子主要在细胞核内发挥其生物学功能。与此相似,高粱Sorghum bicolor [26]和黑果枸杞Lycium ruthenicum[27]中GATA成员在细胞核中占比分别为80.65%和82.35%。系统进化树分析结果表明:青钱柳GATA家族成员分为4个亚族,这与桑树Morus notabilis [28]的GATA基因的分类结果一致。基因结构分析发现:同一亚家族内GATA成员保守基序数量和分布较为相似,表明各亚家族内基因结构具有相当的保守性,可能具有相似的生物学功能。基因家族功能的多样性与外显子和内含子的数量密切相关,在基因进化过程中,内含子数量的变化对其表达调控具有重要影响[29]。相关研究认为:缺乏内含子或内含子数量较少的基因在植物体内表现出更高的表达敏感性[30],内含子数量可能是调控基因表达效率的关键因素之一。本研究中,Class Ⅰ亚家族基因结构相似且内含子数量最少,均包含1~2个内含子,其中CpaF1st42300、CpaF1st42347和CpaF1st47877等基因表达均在盐胁迫下呈下降趋势,而CpaF1st45364在盐胁迫下表达上调,推测该亚家族成员在参与青钱柳逆境胁迫响应过程中表现出更强积极性。基因复制事件驱动基因家族的扩张,提升了基因丰富度并促进新功能的进化[31]。本研究中,青钱柳GATA家族中共鉴定出33对共线性关系,且部分共线性基因(如CpaF1st41514和CpaF1st00092)在盐胁迫下表现出不同的表达趋势,表明基因复制机制对青钱柳GATA家族成员的多样性形成具有重要影响,这可能提升了对逆境的适应能力。
GATA家族是植物体内广泛存在的锌指蛋白家族成员之一,大部分GATA蛋白含有的特殊锌指结构具有高度保守性[31]。已有研究表明:GATA转录因子不仅参与调节植物的生长发育和次生代谢过程,更是在植物的盐害胁迫响应中发挥重要作用[13]。据报道,盐胁迫下黑果枸杞根部和叶部的LrGATAs基因表达显著上调[27];在甘薯中过表达IbGATA24可与IbCOP9-5相互作用,通过调节抗氧化酶活性与激素信号通路相关基因表达增强植株对盐胁迫的耐受抗性[13]。系统进化树分析发现:Class Ⅳ亚族中与IbGATA24同源性最强的CpaF1st31471和CpaF1st12063在盐胁迫下表达上调,推测其在盐胁迫应对中起正向调控作用。本研究中,不同亚家族的CpGATAs在盐胁迫下表达模式各异,Class Ⅲ亚家族的CpaF1st
12368 、CpaF1st31008和CpaF1st46144在盐胁迫下表达上调,而Class Ⅳ亚家族的CpaF1st00992和CpaF1st13794在盐胁迫下表达下调;此外,不同盐处理时间的CpGATAs表达也存在特异性,来自ClassⅠ亚家族的CpaF1st21387和CpaF1st24908在盐处理15 d的表达量明显高于盐处理30 d的表达量,表明青钱柳GATA家族成员可能具多样性调控机制以应对盐逆境胁迫。结合顺式作用元件分析发现:青钱柳GATA成员包含多种植物激素调节和防御应激响应元件,表明青钱柳GATA因子在非生物胁迫应对中扮演重要角色。 -
本研究基于青钱柳基因组数据,共鉴定出46个GATA基因家族成员,并通过系统进化分析将其划分为4个亚家族,同一亚家族内的基因结构具有高度相似性。此外,从青钱柳GATA成员中检测到33对共线性关系,表明基因复制事件可能在该家族的进化过程中起到了关键作用。顺式作用元件分析发现:青钱柳GATA成员启动子区域含有多个与防御应激响应和激素信号传导相关的作用元件,推测其可能广泛参与青钱柳对非生物胁迫的响应调控过程。结合转录组分析发现:CpaF1st12368、CpaF1st31008和CpaF1st46144在盐胁迫下表达上调,CpaF1st00992和CpaF1st13794在盐胁迫中起到负调控作用。
Identification of the GATA gene family in Cyclocarya paliurus and expression analysis under salt stress
-
摘要:
目的 探究青钱柳Cyclocarya paliurus GATA转录因子在盐胁迫下的调节功能。 方法 基于青钱柳基因组和转录组数据对GATA基因家族进行成员鉴定,全面分析其理化性质、基因结构和系统进化等,并通过4种氯化钠浓度(0、25.7、51.3和77.0 mmol·L−1)的盐胁迫处理,分析GATA基因家族响应盐胁迫的表达模式。 结果 从青钱柳基因组中共鉴定出46个GATA基因家族成员,分布在14条染色体上。GATA家族编码蛋白长度为132~827个氨基酸,蛋白分子量为 14358.49 ~92724.86 Da,亲水系数为−1.119~−0.368;亚细胞定位显示:37个GATA基因定位于细胞核,其余9个基因分布于叶绿体、细胞质和质膜中。启动子顺式作用元件分析预测:GATA基因家族成员中包含生长素响应元件、赤霉素响应元件和低温响应元件等13种响应元件。转录组分析结果表明:CpaF1st31008、CpaF1st45364和CpaF1st12063等12个基因表达量在盐处理30 d时高于15 d,且盐胁迫处理下表达上调,而CpaF1st13794和CpaF1st04260等18个基因在盐胁迫处理下表达下调。结论 青钱柳GATA基因家族成员在转录水平积极参与盐胁迫响应,且表现出多样性的响应模式。图8表2参31 Abstract:Objective This study aims to investigate the regulatory function of GATA transcription factors in Cyclocarya paliurus under salt stress. Method Based on the genomic and transcriptomic data of C. paliurus, members of the GATA gene family were identified, and their physicochemical properties, gene structure, and phylogenetic evolution were comprehensively analyzed. The expression patterns of CpGATAs in response to salt stress were investigated through salt stress treatments with 4 sodium chloride concentrations (0, 25.7, 51.3 and 77.0 mmol·L−1). Result The 46 GATA members were identified from the genome, distributed on 14 chromosomes. The encoded protein had a length of 132−827 amino acids, a molecular weight of 14358.49 −92724.86 Da, and a hydrophilicity coefficient of −1.119 − −0.368. Subcellular localization showed that 37 GATA genes were located in the nucleus, while the remaining 9 genes were distributed in chloroplasts, cytoplasm, and plasma membrane. The analysis and prediction of promoter cis acting elements indicated that GATA gene family members included 13 cis-acting promoter elements, including auxin responsive element, gibberellin responsive element and low-temperature responsive element. Transcriptome analysis showed that the expression levels of 12 genes, including CpaF1st31008, CpaF1st45364, and CpaF1st12063, were higher at 30 days of salt treatment than at 15 days, and were upregulated under salt stress. However, 18 genes, including CpaF1st13794 and CpaF1st04260, were downregulated under salt stress.Conclusion Members of the GATA gene family in C. paliurus actively participate in salt stress response at the transcriptional level and exhibit diverse response patterns. [Ch, 8 fig. 2 tab. 31 ref.] -
Key words:
- Cyclocarya paliurus /
- GATA transcription factor /
- salt stress /
- expression analysis
-
图 1 青钱柳 GATA 基因家族系成员统发育分析
Figure 1 Phylogenetic analysis of GATA gene family in C. paliurus
图 2 青钱柳 GATA 家族保守基序分析和基因结构
Figure 2 Motif analysis and gene structure of GATA family in C. paliurus
图 6 青钱柳GATA基因家族顺式作用元件分析
Figure 6 Analysis of cis-regulatory elements of GATA gene family in C. paliurus
图 7 青钱柳GATA基因家族在盐胁迫下的表达模式
Figure 7 Expression patterns of GATA gene family in C. paliurus under salt stress
图 8 盐处理30 d青钱柳GATA基因表达的RT-qPCR验证
Figure 8 RT-qPCR validation of GATA genes expression under salt stress for 30 days
表 1 RT-qPCR引物序列
Table 1. Primer sequences for RT-qPCR
引物 正向引物序列(5′→ 3′) 反向引物序列(5′→ 3′) 18S rRNA AGTATGGTCGCAAGGCTGAAA CAGACAAATCGCTCCACCAA CpaF1st40248 GCTCAGAACACGGAAGTGGA CTGGCCTATACTCTGGCAGC CpaF1st03085 CAGGCCGACGAGGATAGTTC TGGCTGCATCTACGCTGAAA CpaF1st33138 CCGGAACCGGAACTGGTAAA CAAAAACTGGACACCGCAGG CpaF1st12063 CAGGCAGTGCTTCTACTGCT CTGGAGTGTCGGGCTCATTT CpaF1st29899 TGTCGGCACAAGATGAGGTC CCCAGTCTCCCCATTTGCAT CpaF1st24236 CCCAATTCTCCCCATCTCCG TAGGACCCTTGCTTCCCTGT CpaF1st06121 ACAACTGTGGTGGTGCTTCA TTGGTTCGCCTTCTCTGGAC CpaF1st13794 AGCGAGAGCAGAGAGTGAGA GTCCACAAGCCATACGACCA CpaF1st41514 ATGGGGAAGCATGAAGGGTG TTCACCTTGCCTGGATGGAC  下载: 导出CSV
下载: 导出CSV
表 2 青钱柳GATA家族理化性质分析
Table 2. Physical and chemical properties analysis of GATA family in C. paliurus
基因 编码蛋白长度/个氨基酸 分子量/Da 等电点 不稳定指数 脂肪族指数 亲水性 基因位置 亚细胞定位 CpaF1st31008 350 38 871.14 5.42 55.22 58.54 −0.718 Chr5:3275133-3276675 细胞核 CpaF1st04551 333 36 772.69 6.27 59.72 55.98 −0.759 Chr10:5255396-5257097 细胞核 CpaF1st40248 278 31 171.34 8.85 61.17 68.71 −0.711 Chr7:36854287-36855776 细胞核 CpaF1st11493 259 29 016.11 6.52 50.25 54.67 −0.789 Chr12:27234110-27235170 细胞核 CpaF1st03085 399 44 049.62 5.58 59.07 50.63 −0.728 Chr1:39570084-39572566 细胞核 CpaF1st12368 440 48 903.86 6.51 50.23 63.36 −0.672 Chr13:4422154-4428720 细胞核 CpaF1st33138 376 41 285.27 6.53 57.64 55.74 −0.639 Chr5:28272282-28274034 细胞核 CpaF1st15405 304 33 337.68 8.77 59.30 65.72 −0.519 Chr14:14310065-14314628 细胞核 CpaF1st44867 381 41 982.50 5.30 64.62 52.23 −0.708 Chr9:24293467-24295185 细胞核 CpaF1st04260 337 36 733.24 5.96 55.24 65.76 −0.519 Chr10:2743778-2747468 细胞核 CpaF1st10391 827 92 724.86 8.74 43.31 77.56 −0.368 Chr12:16098279-16120607 质膜 CpaF1st33607 162 17 768.52 9.88 49.03 68.09 −0.560 Chr5:34149029-34150449 细胞核 CpaF1st05998 132 14 358.49 9.68 57.91 64.32 −0.765 Chr10:23155264-23156372 细胞核 CpaF1st45364 141 15 497.20 10.40 52.67 68.58 −0.482 Chr9:31189984-31191453 细胞核 CpaF1st05992 151 16 553.39 9.76 48.39 73.71 −0.430 Chr10:23093430-23094145 细胞核 CpaF1st42300 249 27 444.28 8.34 63.80 48.23 −0.743 Chr8:29402271-29403166 细胞核 CpaF1st42347 250 27 563.34 8.06 64.29 47.28 −0.764 Chr8:29941694-29942592 细胞核 CpaF1st06487 252 27 884.50 8.03 60.42 37.22 −0.907 Chr10:29829715-29830640 细胞核 CpaF1st24908 352 39 131.69 8.82 53.42 61.59 −0.726 Chr3:6660741-6662851 细胞核 CpaF1st14436 225 24 929.32 9.19 42.21 39.51 −1.119 Chr14:936552-939513 细胞核 CpaF1st12063 364 40 120.77 4.85 48.65 70.47 −0.665 Chr13:1849772-1855913 细胞核 CpaF1st31471 359 38 841.07 4.97 48.63 67.33 −0.658 Chr5:7431224-7439528 叶绿体 CpaF1st21387 327 36 490.18 9.69 50.48 57.92 −0.869 Chr2:9770484-9772235 细胞核 CpaF1st19401 528 58 511.73 6.11 55.39 67.54 −0.564 Chr16:14302411-14309576 细胞核 CpaF1st18106 258 28 349.77 9.05 56.55 58.64 −0.799 Chr15:20131363-20133090 细胞核 CpaF1st29466 542 60 103.26 5.92 55.85 70.30 −0.580 Chr4:23938169-23945173 细胞核 CpaF1st12064 310 33 115.02 5.12 42.23 73.61 −0.423 Chr13:1856825-1860692 叶绿体 CpaF1st03558 308 32 874.44 4.89 41.13 64.29 −0.542 Chr1:43510795-43514419 叶绿体 CpaF1st43529 305 32 777.21 5.77 32.70 65.28 −0.595 Chr9:5950153-5957707 细胞核 CpaF1st31469 304 32 856.19 5.83 38.43 60.63 −0.670 Chr5:7423788-7429644 细胞核 CpaF1st29899 541 60 796.46 5.96 47.87 74.23 −0.659 Chr4:30906208-30913091 细胞核 CpaF1st30660 336 36 905.18 6.21 44.02 66.19 −0.471 Chr5:498151-499582 叶绿体 CpaF1st42916 345 37 921.05 6.13 38.43 59.39 −0.623 Chr9:535642-537262 细胞质 CpaF1st18811 538 60 555.96 5.97 45.12 70.63 −0.672 Chr16:5904043-5913138 细胞核 CpaF1st24236 373 41 270.78 5.30 41.12 63.86 −0.564 Chr2:43555546-43557671 细胞核 CpaF1st15509 775 84 488.54 6.27 41.70 64.62 −0.815 Chr14:15829121-15836785 叶绿体 CpaF1st31203 373 40 721.49 5.86 45.42 60.46 −0.397 Chr5:4827679-4829481 叶绿体 CpaF1st00992 667 75 052.44 7.08 70.50 61.56 −0.838 Chr1:12126306-12130501 细胞核 CpaF1st06121 654 72 697.62 6.00 60.97 66.36 −0.722 Chr10:24970094-24974866 细胞核 CpaF1st13794 708 78 850.38 7.29 64.92 68.04 −0.702 Chr13:22615455-22620576 细胞核 CpaF1st41514 701 77 486.71 6.69 52.86 70.67 −0.644 Chr8:18653928-18659825 细胞核
下载: 导出CSV
-
[1] LOWRY J A, ATCHLEY W R. Molecular evolution of the GATA family of transcription factors: conservation within the DNA-binding domain [J]. Journal of Molecular Evolution, 2000, 50(2): 103−115. [2] DANIEL-VEDELE F, CABOCHE M. A tobacco cDNA clone encoding a GATA-1 zinc finger protein homologous to regulators of nitrogen metabolism in fungi [J]. Molecular & General Genetics, 1993, 240(3): 365−373. [3] BI Yongmei, ZHANG Yu, SIGNORELLI T, et al. Genetic analysis of Arabidopsis GATA transcription factor gene family reveals a nitrate-inducible member important for chlorophyll synthesis and glucose sensitivity [J]. The Plant Journal, 2005, 44(4): 680−692. [4] BEHRINGER C, SCHWECHHEIMER C. B-GATA transcription factors-insights into their structure, regulation, and role in plant development[J/OL]. Frontiers in Plant Science, 2015, 6: 90[2025-03-22]. DOI: 10.3389/fpls.2015.00090. [5] REYES J C, MURO-PASTOR M I, FLORENCIO F J. The GATA family of transcription factors in Arabidopsis and rice [J]. Plant Physiology, 2004, 134(4): 1718−1732. [6] 杨玄松, 谢雯榕, 顾钢, 等. 普通烟草GATA转录因子家族鉴定及基因表达分析[J]. 烟草科技, 2023, 56(1): 11−23. YANG Xuansong, XIE Wenrong, GU Gang, et al. Identification and gene expression analysis of GATA transcription factor family in Nicotiana tabacum [J]. Tobacco Science & Technology, 2023, 56(1): 11−23. [7] 王伟杰, 雷亚柯, 张建航, 等. 花生GATA基因家族鉴定和表达分析[J]. 山西农业大学学报(自然科学版), 2024, 44(2): 10−22. WANG Weijie, LEI Yake, ZHANG Jianhang, et al. Identification and expression analysis of peanut GATA gene family [J]. Journal of Shanxi Agricultural University (Natural Science Edition), 2024, 44(2): 10−22. [8] MARA C D, IRISH V F. Two GATA transcription factors are downstream effectors of floral homeotic gene action in Arabidopsis [J]. Plant Physiology, 2008, 147(2): 707−718. [9] AN Yi, HAN Xiao, TANG Sha, et al. Poplar GATA transcription factor PdGNC is capable of regulating chloroplast ultrastructure, photosynthesis, and vegetative growth in Arabidopsis under varying nitrogen levels [J]. Plant Cell, Tissue and Organ Culture (PCTOC), 2014, 119(2): 313−327. [10] REN Mengxuan, ZHANG Yang, LIU Cong, et al. Characterization of a high hierarchical regulator, PtrGATA12, functioning in differentially regulating secondary wall component biosynthesis in Populus trichocarpa[J/OL]. Frontiers in Plant Science, 2021, 12: 657787[2025-03-22]. DOI: 10.3389/fpls.2021.657787. [11] ZHANG Hongjia, WU Tao, LI Zhao, et al. OsGATA16, a GATA transcription factor, confers cold tolerance by repressing OsWRKY45-1 at the seedling stage in rice[J/OL]. Rice, 2021, 14(1): 42[2025-03-22]. DOI: 10.1186/s12284-021-00485-w. [12] SHEN Chao, ZHANG Yue, LI Qing, et al. PdGNC confers drought tolerance by mediating stomatal closure resulting from NO and H2O2 production via the direct regulation of PdHXK1 expression in Populus [J]. New Phytologist, 2021, 230(5): 1868−1882. [13] ZHU Hong, ZHAI Hong, HE Shaozhen, et al. A novel sweetpotato GATA transcription factor, IbGATA24, interacting with IbCOP9-5a positively regulates drought and salt tolerance[J/OL]. Environmental and Experimental Botany, 2022, 194: 104735[2025-03-22]. DOI: 10.1016/j.envexpbot.2021.104735. [14] 刘洋, 陈培, 周明明, 等. 光质对不同家系青钱柳生物量和叶三萜类化合物积累的影响[J]. 浙江农林大学学报, 2020, 37(6): 1097−1104. LIU Yang, CHEN Pei, ZHOU Mingming, et al. Effects of light quality on biomass and triterpenoid accumulation in leaves of Cyclocarya paliurus from different families [J]. Journal of Zhejiang A&F University, 2020, 37(6): 1097−1104. [15] ZHANG Lei, ZHANG Zijie, FANG Shengzuo, et al. Metabolome and transcriptome analyses unravel the molecular regulatory mechanisms involved in photosynthesis of Cyclocarya paliurus under salt stress[J/OL]. International Journal of Molecular Sciences, 2022, 23(3): 1161[2025-03-22]. DOI: 10.3390/ijms23031161. [16] QU Yinquan, SHANG Xulan, ZENG Ziyan, et al. Whole-genome duplication reshaped adaptive evolution in a relict plant species, Cyclocarya paliurus[J]. Genomics, Proteomics & Bioinformatics, 2023, 21(3): 455−469. [17] TAMURA K, STECHER G, KUMAR S. MEGA11: molecular evolutionary genetics analysis version 11 [J]. Molecular Biology and Evolution, 2021, 38(7): 3022−3027. [18] CHEN Chengjie, CHEN Hao, ZHANG Yi, et al. TBtools: an integrative toolkit developed for interactive analyses of big biological data [J]. Molecular Plant, 2020, 13(8): 1194−1202. [19] LESCOT M, DÉHAIS P, THIJS G, et al. PlantCARE, a database of plant Cis-acting regulatory elements and a portal to tools for in silico analysis of promoter sequences [J]. Nucleic Acids Research, 2002, 30(1): 325−327. [20] PERTEA M, KIM D, PERTEA G M, et al. Transcript-level expression analysis of RNA-seq experiments with HISAT, StringTie and Ballgown [J]. Nature Protocols, 2016, 11(9): 1650−1667. [21] ZHAO Kai, NAN Siyuan, LI Yajing, et al. Comprehensive analysis and characterization of the GATA gene family, with emphasis on the GATA6 transcription factor in poplar[J/OL]. International Journal of Molecular Sciences, 2023, 24(18): 14118[2025-03-22]. DOI: 10.3390/ijms241814118. [22] NUTAN K K, SINGLA-PAREEK S L, PAREEK A. The Saltol QTL-localized transcription factor OsGATA8 plays an important role in stress tolerance and seed development in Arabidopsis and rice [J]. Journal of Experimental Botany, 2020, 71(2): 684−698. [23] ZHANG Lei, ZHANG Zijie, FANG Shengzuo, et al. Integrative analysis of metabolome and transcriptome reveals molecular regulatory mechanism of flavonoid biosynthesis in Cyclocarya paliurus under salt stress[J/OL]. Industrial Crops and Products, 2021, 170: 113823[2025-03-22]. DOI: 10.1016/j.indcrop.2021.113823. [24] 骆鹰, 王有成, 王伟平, 等. 水稻GATA 基因家族生物信息学分析[J]. 分子植物育种, 2018, 16(17): 5514−5522. LUO Ying, WANG Youcheng, WANG Weiping, et al. Bioinformatics analysis of GATA gene family in rice [J]. Molecular Plant Breeding, 2018, 16(17): 5514−5522. [25] 袁岐, 张春利, 赵婷婷, 等. 辣椒GATA转录因子的生物信息学分析[J]. 中国农学通报, 2017, 33(17): 24−31. YUAN Qi, ZHANG Chunli, ZHAO Tingting, et al. Bioinformatics analysis of GATA transcription factor in pepper [J]. Chinese Agricultural Science Bulletin, 2017, 33(17): 24−31. [26] 姚茂星, 周光怡, 丁延庆, 等. 高粱GATA转录因子家族的鉴定和表达模式分析[J]. 分子植物育种, 2022, 20(10): 3178−3187. YAO Maoxing, ZHOU Guangyi, DING Yanqing, et al. Identification and expression pattern analysis of GATA transcription factor family in Sorghum (L. ) [J]. Molecular Plant Breeding, 2022, 20(10): 3178−3187. [27] 曾雨, 祝建刚, 杜雨, 等. 黑果枸杞GATA转录因子家族鉴定与生物信息学分析[J]. 草地学报, 2025, 33(6): 1724−1735. ZENG Yu, ZHU Jiangang, DU Yu, et al. Identification and bioinformatics analysis of GATA transcription factor family in Lycium ruthenicum [J]. Acta Agrestia Sinica, 2025, 33(6): 1724−1735. [28] 刘江, 刘刚, 曾益春, 等. 桑树GATA 基因家族的全基因组鉴定与分析[J]. 西南农业学报, 2022, 35(12): 2724−2731. LIU Jiang, LIU Gang, ZENG Yichun, et al. Genome-wide identification and analysis of mulberry (Morus notabilis) GATA gene family [J]. Southwest China Journal of Agricultural Sciences, 2022, 35(12): 2724−2731. [29] XU Bin, SATHITSUKSANOH N, TANG Yuhong, et al. Overexpression of AtLOV1 in Switchgrass alters plant architecture, lignin content, and flowering time[J/OL]. PLoS One, 2012, 7(12): e47399[2025-03-22]. DOI: 10.1371/journal.pone.0047399. [30] CHUNG B Y W, SIMONS C, FIRTH A E, et al. Effect of 5’UTR introns on gene expression in Arabidopsis thaliana[J/OL]. BMC Genomics, 2006, 7: 120[2025-03-22]. DOI: 10.1186/1471-2164-7-120. [31] 敖涛, 廖晓佳, 徐伟, 等. 蓖麻GATA 基因家族的鉴定和特征分析[J]. 植物分类与资源学报, 2015, 37(4): 453−462. AO Tao, LIAO Xiaojia, XU Wei, et al. Identification and characterization of GATA gene family in Castor bean (Ricinus communis) [J]. Plant Diversity and Resources, 2015, 37(4): 453−462. -

-
链接本文:
https://zlxb.zafu.edu.cn/article/doi/10.11833/j.issn.2095-0756.20250263
 点击查看大图
点击查看大图
计量
- 文章访问数: 1128
- HTML全文浏览量: 373
- PDF下载量: 86
- 被引次数: 0
