




-
杜鹃群落是以杜鹃属Rhododendron植物为优势种的植被类型,主要分布于中国西南地区海拔较高的山地[1]。全世界杜鹃属植物约960种,中国约650种,是杜鹃属分布中心,也是杜鹃花生物多样性最高的国家,除新疆地区之外,全国其他省(市、自治区)均有分布,其中以西南地区分布最为广泛。贵州省是杜鹃原生种分布区数量最多的省份之一,是杜鹃花中国中心分布区向外扩张的过渡地带,有着丰富的原生种和大量可能的新种和变种[2]。有关学者先后对杜鹃植物开展了种子萌发特性、形态解剖、抗旱评价、引种驯化、繁殖利用、群落特征以及多样性保护及保育研究[3-5]。但对于杜鹃群落生物多样性的研究较少,尤其采用群落数量分类学方法对杜鹃群落生物多样性研究鲜有报道。采用群落数量分类方法结合群丛特性进行分析,能够定量解析群丛特性、组成特点,更加准确地反映杜鹃与其他植物种类间的关系。在对退化杜鹃群落进行恢复中,以群丛为单位进行恢复,提高了其可操作性。本研究选择贵阳市乌当区万亩杜鹃群落作为研究对象,采用典型样地调查、群落数量分类及多样性分析等方法展开研究,旨在筛选出万亩杜鹃群落生物多样性最高的群丛,为杜鹃群落生物多样性保护提供技术指导,奠定理论基础。
-
本研究选择贵阳市乌当区典型杜鹃群落为研究对象。乌当区位于贵阳市东北部,属贵阳城市规划的9个组团之一,地处26°55′~26°33′N,106°30′~107°03′E。乌当区属亚热带季风湿润气候,具有明显的高原性气候的特点,冬无严寒,夏无酷暑,光、热、水同季,垂直气候差异明显,年平均降水量1 179.8~1 271.0 mm。年平均气温14.6 ℃。森林覆盖率达49.36%。由于地形、地貌类型多样,形成多种小气候,宜于农业生产,大部分地区可满足农作物1 a 2熟,蔬菜1 a 3~4熟的需要。主要灾害性天气有干旱、倒春寒、冰雹、秋季连阴雨、秋风、凝冻[6]。区境内河流均属于长江流域乌江水系,河床狭窄,比降大,流速较快。全区土壤为黄壤区,黄棕壤、石灰土、紫色土和冲积土[7]。植被属于黔中山原湿润性灰岩常绿栎林、常绿落叶混交林及马尾松Pinus massoniana林区,主要树种有青栲Cyclobalanopsis glauca,丝栗栲Castanopsis fargesii,细叶青冈栎Cyclobalanopsis gracilis,柞木 Quercus mongolica等,落叶树种有枫香Liquidambar formosana,光皮桦 Betula luminifera等。次生植被有大面积天然牧草和灌丛草坡[8-9]。
-
采用典型样地调查法,对贵阳市乌当区杜鹃群落进行调查。在贵阳市乌当区杜鹃群落分布区域随机布设调查样点15处,分别在每个调查样点区域内随机设置10 m × 10 m典型群落调查样方3个,共计调查样方45个,统计每个调查样方内的植物种类、生活型、密度、高度、盖度、频度等群落特征指标,用于群落数量分类与多样性等指标的计算。
-
贵阳乌当杜鹃群落调查数据统计分析与整理,均采用SPSS 18.0软件和Microsoft Office Excel 2007软件完成。
-
结合《中国植被》和实际生态学意义,采用TWINSPAN分类法,对贵阳乌当杜鹃群落进行群落数量分类。
-
采用重要值(Ⅵ,对贵阳乌当杜鹃群落进行分析,计算公式[10-11]:
$$I{V_{Ii}} = \left( {\frac{{{D_i}}}{{\sum {{D_i}} }} + \frac{{{F_i}}}{{{{\sum F }_i}}} + \frac{{{C_i}}}{{\sum {{C_i}} }}} \right) \div 3 \times 100\% $$ 其中: VIi表示第i种物种的重要值;Di表示第i种物种的密度;Fi表示第i种物种的频度;Ci表示第i种物种的盖度(乔木层中Ci表示第i中物种的显著度)。
-
采用Margalef指数、Simpson指数和Shannon-Wiener指数对贵阳乌当杜鹃群落物种多样性进行分析,计算公式为[10-13]:
$$\begin{array}{l} {\rm{M}}\arg {\rm{alef}}指数R = \frac{{S - 1}}{{\ln N}};\\ {\rm{Simpson}}指数D = \sum\limits_{i = 1}^s {{{({p_i})}^2}} ;\\ {\rm{Simpson}} - {\rm{Wiener}}指数H = - \sum\limits_{i = 1}^s {({p_i}ln{p_i})} \end{array}。$$ 其中:S表示植被群落中物种数目,N表示群丛中全部物种个体总数,Pi为第i种的相对重要值。
-
采用Pielou均匀性指数对贵阳乌当杜鹃群落进行分析,计算公式为[11]:
$${\rm{Pielou}}均匀性指数E = \frac{H}{{\ln S}}。$$ 其中:H 表示Shannon-Wiener多样性指数,S表示植被群落中物种数目。
-
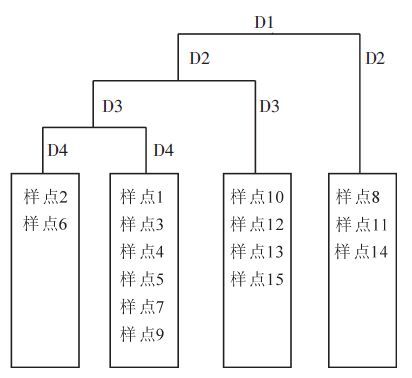
采用TWINSPAN分类法,对贵阳乌当杜鹃群落15个调查样点进行等级分类,经过4次划分,得到为4组。结合《中国植被》和实际生态意义,采用第4级的分类结果,将贵阳乌当杜鹃群落划分为4组(图 1),结合分类将4个组命名为4个植物群丛。群丛Ⅰ:锈叶杜鹃Rhododendron siderophyllum+白栎Quercus fabrei群丛,包括样点2和样点6。群丛Ⅱ:锈叶杜鹃+白栎+麻栎Quercus acutissima群丛,包括样点1,样点4,样点9,样点3,样点5和样点7。群丛Ⅲ:锈叶杜鹃+麻栎群丛,包括样点10,样点12,样点13和样点15号。群丛Ⅳ:锈叶杜鹃+麻栎+藤黄Garcinia hanburyi群丛,包括样点8,样点11和样点14。

图 1 乌当杜鹃群落15个样点TWINSPAN树状分类图
Figure 1. TWINSPAN classification of 15 sampling pointies in the Wudang of the Rhododendron community
表 1 贵阳乌当杜鹃群丛重要值特征
Table 1. Important values trait of the Rhododendron communities in Wudang,Guiyang
科名 种名 生活型 重要值 群丛I 群丛Ⅱ 群丛Ⅲ 群丛Ⅳ 杜鹃花科Ericaceae 镑叶杜鹃Rhododendron siderophyllum S 48.72 29.80 36.23 48.74 杜鹃花科Ericaceae 小果南烛Lyonia ovalifolia S 6.02 5.51 5.26 — 杜鹃花科Ericaceae 映山红Rhododendron S 7.32 8.12 8.27 4.66 壳斗科Dagaceae 麻栋Quercus acutissima T 8.91 28.55 11.34 10.28 壳斗科Dagaceae 栲 Castanopsis fargesii T — 3.55 5.80 0.80 壳斗科Dagaceae 白栋Quercus fabrei S 13.66 11.45 1.90 7.90 禾本科Gramineae 五节芒 Miscanthus floridulus P — 0.17 — — 禾本科Gramineae 茅叶荩草 Arthraxon lanceolatus P — — 0.64 — 禾 本 科 Gramineae 芒 Miscanthus sinensis P — — 0.88 1.04 蕨科 Pteridiaceae 蕨 Pteridium aquilinum P — 1.32 2.13 — 蕨科 Pteridiaceae 密毛蕨 Pteridium reyolutum P — — 0.76 5.07 桑科 Moraceae 小构树 Broussonetia CasinoCi S — — 0.40 — 桑科 Moraceae 專草 Humulus scandens P — — 0.67 — 藤黄科 Guttiferae 藤黄 Farcinia hanburyi T — — — 8.82 藤黄科 Guttiferae 金丝桃 Hypericum monogynum S — 1.11 — — 百合科Liliaceae 拔葜 Smilax china CS 0.67 8.34 — — 车前科 Plantaginaceae 车 前 草 Plantago asiatica P — — 0.37 — 豆科 Leguminosae 三叶崖豆藤 Millettia unijuga CS — 1.39 — — 金缕梅科 Hamamelidaceae 枫香 Liquidambar formosana T — — — 2.23 菊科 Compositae 艾蒿 Artemisia vulgaris P — — 0.46 — 茜草科Rubiaceae 鸡矢藤 Paederia scandens CS — — 1.19 — 蔷薇科Rosaceae 插田泡Rubus coreanus S — — 1.52 2.88 忍冬科 Caprifoliaceae 珍珠荚莲 liburnum/oetidum S 3.88 — 7.40 — 莎草科 Cyperaceae 莎 草 Cyperus rotundus P — 0.34 — — 山茶科Theaceae 怜木 Juryajaponica S 3.33 — 5.96 4.36 杉科 Taxodiaceae 杉木 Cunninghamia lanceolata T 7.50 — — — 石松科 Lycopodiaceae 石松 Lycopodium clavatum P — — 2.71 — 石竹科 Caryophyllaceae 银柴胡 Fypsophilaoldhamiana P — — 0.37 — 五加科 Araliaceae 檧木 Aralia chinensis S — — 5.75 2.53 虎耳草科 Saxifragaceae 滇鼠刺 Ktea ilicifolia S — 0.36 — — 苋科 Amaranthaceae 牛漆 Achyranthes bidentata P — — — 0.69 说明:共计植物31种,其中群丛Ⅰ9种,群丛Ⅱ13种,群丛Ⅲ21种,群丛Ⅳ13种。“—”表示该样地中未发现该种植物;“T”表示乔木植物(tree);“P”表示多年生植物(perennial plant);“S”表示灌木(shrubbery);“CS”表示攀援灌木(climbing shrub)。 -
群丛Ⅲ中,杜鹃花科Ericaceae,壳斗科Fagaceae分别共出现3种植物,分别占群丛出现种的14.29%;禾本科Gramineae,蕨科Pteridiaceae,桑科Moraceae分别出现2种植物,分别占群丛出现种的9.52%,其他科共出现9种植物,占群丛出现种的42.86%。群丛Ⅳ中,壳斗科出现3种植物,占群丛出现种的23.08%,杜鹃花科出现2种植物,占群丛出现种的15.38%;其他科共出现7种植物,占群丛出现种的53.85%。
对贵阳乌当杜鹃群落植物生活型分析发现,乔木植物5种,占群落出现种的16.13%;灌木植物14种(其中攀援灌木植物3种,占群落出现种的9.68%),占群落出现种的45.16%;草本植物12种(均为多年生草本植物)占群落出现种的38.71%。对贵阳乌当杜鹃群落4种群丛植物组成生活型分析发现,群丛Ⅰ中共出现9种植物,其中乔木2种,占群丛出现植物种的22.22%;灌木植物7种(其中攀援植物1种,占群丛出现种的11.11%),占群丛出现种的77.78%;群丛中未出现草本植物种类。群丛Ⅱ中共出现13类植物,其中乔木2种,占群丛出现植物种的15.38%;灌木植物8种(其中攀援植物2种,占群丛出现种的15.38%),占群丛出现种的61.54%;草本植物3种,占群丛出现种的23.08%。群丛Ⅲ中共出现21种植物,其中乔木2种,占群丛出现植物种的9.52%;灌木植物10种(其中攀援植物1种,占群丛出现种的4.76%),占群丛出现种的47.62%;草本植物9种,占群丛出现种的42.86%。群丛Ⅳ中共出现13种植物,其中乔木4种,占群丛出现植物种的30.77%;灌木植物6种,占群丛出现种的46.15%;草本植物3种,占群丛出现种的23.08%。
对贵阳乌当杜鹃群丛植物重要值分析发现,群丛Ⅰ中,锈叶杜鹃重要值最高,为48.72%,白栎次之,为13.66%;群丛Ⅱ中,锈叶杜鹃重要值最高,为29.80%,麻栎次之,为28.55%,白栎重要值为11.45%;群丛Ⅲ中,锈叶杜鹃重要值最高,为36.23%,麻栎次之,为11.34%;群丛Ⅳ中,锈叶杜鹃重要值最高,为48.74%,麻栎次之,为10.28%。
综上所述,乌当杜鹃群落是以锈叶杜鹃为主,白栎、麻栎为辅的群落类型。
-
贵阳乌当杜鹃群落Margalef丰富度指数分析如表 2所示。Margalef丰富度指数从高到低的变化趋势为群丛Ⅲ(4.714),群丛Ⅱ(3.306),群丛Ⅳ(3.081),群丛Ⅰ(2.111)。
表 2 贵阳乌当杜鹃群落α多样性与均匀性指数特征
Table 2. Index trait of α diversity and evenness of Rhododendron communities in Wudang,Guiyang
群丛 R D H E Ⅰ 2.111 0.719 1.665 0.758 Ⅱ 3.306 0.798 1.873 0.730 Ⅲ 4.714 0.828 2.270 0.745 Ⅳ 3.081 0.729 1.830 0.714 贵阳乌当杜鹃群落Simpson多样性指数、Shannon-Wiener多样性指数分析如表 2所示。Simpson多样性指数从高到低的变化趋势为群丛Ⅲ(0.828),群丛Ⅱ(0.798),群丛Ⅳ(0.729),群丛Ⅰ(0.719);Shannon-Wiener多样性指数从高到低的变化趋势为群丛Ⅲ(2.270),群丛Ⅱ(1.873),群丛Ⅳ(1.830),群丛Ⅰ(1.665)。
贵阳乌当杜鹃群落Pielou均匀性指数分析如表 2所示。Pielou均匀性指数从高到低的变化趋势为群丛Ⅰ(0.758),群丛Ⅲ(0.745),群丛Ⅱ(0.730),群丛Ⅳ(0.714)。
综上所述,贵阳乌当杜鹃群落Margalef丰富度指数、Simpson多样性指数、Shannon-Wiener多样性指数均呈现出群丛Ⅲ>群丛Ⅱ>群丛Ⅳ>群丛Ⅰ的趋势,而Pielou均匀性指数则呈现出群丛Ⅰ>群丛Ⅲ>群丛Ⅱ>群丛Ⅳ。
-
贵阳乌当杜鹃群落可分为4种植物群丛,依次为群丛Ⅰ:锈叶杜鹃+白栎群丛;群丛Ⅱ:锈叶杜鹃+白栎+麻栎群丛;群丛Ⅲ:锈叶杜鹃+麻栎群丛;群丛Ⅳ:锈叶杜鹃+麻栎+藤黄群丛。杜鹃群落主要以杜鹃科、壳斗科、禾本科植物为主;在贵阳乌当杜鹃群落中锈叶杜鹃种群占绝对优势,麻栎、白栎种群次之,因此,杜鹃群落主要是以锈叶杜鹃为主,麻栎、白栎为辅的植物群落结构类型。贵阳乌当杜鹃群丛Margalef丰富度指数、Simpson多样性指数、Shannon-Wiener多样性指数均呈现出群丛Ⅲ>群丛Ⅱ>群丛Ⅳ>群丛Ⅰ的趋势,而Pielou均匀性指数则呈现出群丛Ⅰ>群丛Ⅲ>群丛Ⅱ>群丛Ⅳ。贵阳乌当杜鹃群落多样性保护应当遵从群丛Ⅲ的植物结构类型进行保护,对群丛Ⅰ,群丛Ⅱ和群丛Ⅳ进行适当修复,使其结构接近群丛Ⅲ的结构类型。
-
本研究针对贵阳乌当万亩杜鹃林群落进行多样性研究,按植物群落的自然演替规律,该群落处于灌木或灌丛群落阶段,正向乔灌群落阶段演替,并非顶级群落及地带性植被类型,因此,处于不稳定阶段。目前,贵阳市乌当区想要打造万亩杜鹃林,作为贵阳城郊以观赏杜鹃花的休闲旅游区,但随着群落演替的进行,杜鹃群落的变化会直接影响旅游观光效果,因此,进一步研究贵阳乌当万亩杜鹃林最佳群落结构、物种组成、物种多样性、人工正向干扰或结构调整,对旅游观光效果及群落的相对稳定性有重要意义。正如黔西的百里杜鹃,目前观光效果很好,在长期人为正向干扰下,其群落结构具相对稳定性,但从物种多样性及演替规律来看,目前的结构是否为最佳,有待于进一步研究。
Analysis of quantity characteristics and α diversity of the Rhododendron communities in Wudang District, Guiyang City
-
摘要: 选择贵阳市乌当区典型杜鹃Rhododendron群落作为研究对象,采用标准样地调查法、群落数量分析法等方法,对其不同杜鹃群丛的植物组成、结构、重要值、α多样性分析进行分析。结果表明:①贵阳乌当杜鹃群落分为4种植物群丛,分别为Ⅰ:锈叶杜鹃Rhododendron siderophyllum+白栎Quercus fabrei群丛;Ⅱ:锈叶杜鹃+白栎+麻栎Quercus acutissima群丛;Ⅲ:锈叶杜鹃+麻栎群丛;Ⅳ:锈叶杜鹃+麻栎+藤黄Garcinia hanburyi群丛。②杜鹃群落主要以锈叶杜鹃为主,麻栎、白栎为辅的植物群落结构类型。③杜鹃群丛Margalef丰富度指数、Simpson多样性指数、Shannon-Wiener多样性指数均呈现出群丛Ⅲ> 群丛Ⅱ> 群丛Ⅳ> 群丛Ⅰ的趋势,而Pielou均匀性指数则呈现出群丛Ⅰ> 群丛Ⅲ> 群丛Ⅱ >群丛Ⅳ的趋势。④杜鹃群落多样性保护应当遵从群丛Ⅲ的植物结构类型进行保护,对群丛Ⅰ,群丛Ⅱ和群丛Ⅳ群丛进行适当修复,使其结构接近群丛Ⅲ的结构类型。Abstract: The Rhododendron communities in Wudang District of Guiyang City was studied by sample plot and quantitative analysis to analyze the plant components, structures, important values and α diversities of different Rhododendron communities. The research had the following findings.(1)There were four Rhododendron communities in Wudang District of Guiyang City,namely,Rhododendron siderophyllum and Quercus fabrei (Community Ⅰ),Rhododendron siderophyllum, Quercus fabrei and Quercus acutissima(Community Ⅱ),Rhododendron siderophyllum and Quercus acutissima (Community Ⅲ),Rhododendron siderophyllum, Quercus acutissima and Garcinia hanburyi(Community Ⅳ). (2) With Rhododendron siderophyllum as the dominant plants,the Rhododendron communities also had plants such as Quercus acutissima and Quercus fabrei. In terms of Margalef index for richness, Simpson index for diversity,Shannon-Wiener index for diversity,the ranking of four Rhododendron communities was as follows:Community Ⅲ> Community Ⅱ> Community Ⅳ> Community Ⅰ;the ranking for Pielou Evenness index was Community Ⅰ> Community Ⅲ> Community Ⅱ> Community Ⅳ. (4) When preserving the diversity of the Rhododendron communities,the plants structures of Community Ⅲ should be used as reference. Community Ⅰ,Community Ⅱ and Community Ⅳ should be restored to make their plants structures close to that of Community Ⅲ.
-
图 1 乌当杜鹃群落15个样点TWINSPAN树状分类图
Figure 1 TWINSPAN classification of 15 sampling pointies in the Wudang of the Rhododendron community
表 1 贵阳乌当杜鹃群丛重要值特征
Table 1. Important values trait of the Rhododendron communities in Wudang,Guiyang
科名 种名 生活型 重要值 群丛I 群丛Ⅱ 群丛Ⅲ 群丛Ⅳ 杜鹃花科Ericaceae 镑叶杜鹃Rhododendron siderophyllum S 48.72 29.80 36.23 48.74 杜鹃花科Ericaceae 小果南烛Lyonia ovalifolia S 6.02 5.51 5.26 — 杜鹃花科Ericaceae 映山红Rhododendron S 7.32 8.12 8.27 4.66 壳斗科Dagaceae 麻栋Quercus acutissima T 8.91 28.55 11.34 10.28 壳斗科Dagaceae 栲 Castanopsis fargesii T — 3.55 5.80 0.80 壳斗科Dagaceae 白栋Quercus fabrei S 13.66 11.45 1.90 7.90 禾本科Gramineae 五节芒 Miscanthus floridulus P — 0.17 — — 禾本科Gramineae 茅叶荩草 Arthraxon lanceolatus P — — 0.64 — 禾 本 科 Gramineae 芒 Miscanthus sinensis P — — 0.88 1.04 蕨科 Pteridiaceae 蕨 Pteridium aquilinum P — 1.32 2.13 — 蕨科 Pteridiaceae 密毛蕨 Pteridium reyolutum P — — 0.76 5.07 桑科 Moraceae 小构树 Broussonetia CasinoCi S — — 0.40 — 桑科 Moraceae 專草 Humulus scandens P — — 0.67 — 藤黄科 Guttiferae 藤黄 Farcinia hanburyi T — — — 8.82 藤黄科 Guttiferae 金丝桃 Hypericum monogynum S — 1.11 — — 百合科Liliaceae 拔葜 Smilax china CS 0.67 8.34 — — 车前科 Plantaginaceae 车 前 草 Plantago asiatica P — — 0.37 — 豆科 Leguminosae 三叶崖豆藤 Millettia unijuga CS — 1.39 — — 金缕梅科 Hamamelidaceae 枫香 Liquidambar formosana T — — — 2.23 菊科 Compositae 艾蒿 Artemisia vulgaris P — — 0.46 — 茜草科Rubiaceae 鸡矢藤 Paederia scandens CS — — 1.19 — 蔷薇科Rosaceae 插田泡Rubus coreanus S — — 1.52 2.88 忍冬科 Caprifoliaceae 珍珠荚莲 liburnum/oetidum S 3.88 — 7.40 — 莎草科 Cyperaceae 莎 草 Cyperus rotundus P — 0.34 — — 山茶科Theaceae 怜木 Juryajaponica S 3.33 — 5.96 4.36 杉科 Taxodiaceae 杉木 Cunninghamia lanceolata T 7.50 — — — 石松科 Lycopodiaceae 石松 Lycopodium clavatum P — — 2.71 — 石竹科 Caryophyllaceae 银柴胡 Fypsophilaoldhamiana P — — 0.37 — 五加科 Araliaceae 檧木 Aralia chinensis S — — 5.75 2.53 虎耳草科 Saxifragaceae 滇鼠刺 Ktea ilicifolia S — 0.36 — — 苋科 Amaranthaceae 牛漆 Achyranthes bidentata P — — — 0.69 说明:共计植物31种,其中群丛Ⅰ9种,群丛Ⅱ13种,群丛Ⅲ21种,群丛Ⅳ13种。“—”表示该样地中未发现该种植物;“T”表示乔木植物(tree);“P”表示多年生植物(perennial plant);“S”表示灌木(shrubbery);“CS”表示攀援灌木(climbing shrub)。  下载: 导出CSV
下载: 导出CSV
表 2 贵阳乌当杜鹃群落α多样性与均匀性指数特征
Table 2. Index trait of α diversity and evenness of Rhododendron communities in Wudang,Guiyang
群丛 R D H E Ⅰ 2.111 0.719 1.665 0.758 Ⅱ 3.306 0.798 1.873 0.730 Ⅲ 4.714 0.828 2.270 0.745 Ⅳ 3.081 0.729 1.830 0.714
下载: 导出CSV
-
[1] 李朝阳,杜凡,姚莹,等. 轿子山自然保护区杜鹃群落植物多样性研究[J]. 西南林学院学报,2010,30(3):34-37. LI Zhaoyang,DU Fan,YAO Ying,et al. Study on plant diversity of Rhododendron community in Jiaozishan Nature Reserve[J]. J Southwest For Univ,2010,30(3):34-37. [2] 欧静,陈训. 贵州省常绿杜鹃亚属资源及园林应用前景分析[J]. 江苏农业科学,2012,40(8):200-203. OU Jing,CHEN Xun. Analysis on subgenus Rhododendron subgenus resources and landscape application prospect of Guizhou Province[J]. Jiangsu Agric Sci,2012,40(8):200-203. [3] 刘永翔,龙毅,刘作易,等. 贵州几种杜鹃花属植物的花粉形态[J]. 贵州农业科学,2009,37(3):24-26. LIU Yongxiang,LONG Yi,LIU Zuoyi,et al. Pollen Morphology of Rhododendrons in Guizhou[J]. Guizhou Agric Sci,2009, 37(3):24-26. [4] 黄承玲,陈训,高贵龙. 3种高山杜鹃对持续干旱的生理响应及抗旱性评价[J]. 林业科学,2011,47(6):48-55. HUANG Chengling,CHEN Xun,GAO Guilong. Physiological response of seedlings of three Azalea species of drought stress and evaluation of drought resistance[J]. Sci Silv Sin,2011,47(6):48-55. [5] 刘纪岗,谢元贵,杨小庆. 贵州百里杜鹃大草原景区迷人杜鹃群落特征研究[J]. 广东农业科学,2012,39(9):46-48. LIU Jigang,XIE Yuangui,YANG Xiaoqing. Study on community characteristics of Rhododendron agastum in the prairie scenic area of Baili Rhododendron protection zone in Guizhou[J]. Guangdong Agric Sci,2012,39(9):46-48. [6] 耿品富,梅素娟,肖兴跃,等. 乌当区休闲农业与乡村旅游管理经营模式探索[J]. 贵州农业科学,2012,40(5):205-209,213. GENG Pinfu,MEI Sujuan,XIAO Xingyue,et al. Exploration of management business model of leisure agriculture and rural tourism in Wudang District[J]. Guizhou Agric Sci,2012,40(5):205-209,213. [7] 李苇洁,易朝阳,陈志华,等. 贵阳二环林带不同林地土壤养分状况研究[J]. 贵州科学,2007,25(3):28-32. LI Weijie,YI Chaoyang,CHEN Zhihua,et al. Soil nutrition status of different stands in the second ring forest-belt of Guiyang[J]. Guizhou Sci,2007,25(3):28-32. [8] 翁明武,陈训,曾亚军,等. 贵阳二环林带植物群落的主要类型及其保护[J]. 贵州科学,2007,25(3):9-19. WENG Mingwu,CHEN Xun,ZENG Yajun,et al. The main type of the plant community and its protection in the second ring forest-belt of Guiyang[J]. Guizhou Sci,2007,25(3):9-19. [9] 石登红,曾亚军,彭惠蓉,等. 贵阳二环林带生物多样性现状及展望[J]. 贵州科学,2007,25(3):64-68. SHI Denghong,ZENG Yajun,PENG Huirong,et al. Present state and prospect of biodiversity in the second ring forest-belt of Guiyang[J]. Guizhou Sci,2007,25(3):64-68. [10] 张金屯. 数量生态学[M]. 2版. 北京:科学出版社,2011:101-103. [11] 宋永昌. 植被生态学[M]. 上海:华东师范大学出版社,2001:53-54. [12] 马姜明,吴蒙,占婷婷,等. 漓江流域岩溶区檵木群落不同恢复阶段物种组成及多样性变化[J]. 生态环境学报,2013,22(1):66-71. MA Jiangming,WU Meng,ZHAN Tingting,et al. Changes of species composition and diversity among restoration stages of Loropetalum chinense communities in karst area of Lijiang River valley[J]. Ecol Environ Sci,2013,22(1):66-71. [13] ANNE E M. 生物多样性测度[M]. 张峰,译. 北京:科学出版社,2011:93-94. -

-
链接本文:
https://zlxb.zafu.edu.cn/article/doi/10.11833/j.issn.2095-0756.2014.02.007
 点击查看大图
点击查看大图
计量
- 文章访问数: 3386
- HTML全文浏览量: 627
- PDF下载量: 623
- 被引次数: 0
