




 下载:
下载:

-
1964年Shinozaki等[1-2]在研究各种植物群落的生产结构图时,首次发现在某一高度之上的累计叶量与该高度的非同化组织密度之间具有明显的线性关系,从而提出单位管道模型(unit pipe model)。该模型把树木以及整个群落视为一个均一的单位管道的集合,管道将叶子和根连接起来,管道既起到机械支撑的作用,又有输导水分的作用,认为树干内单位管道支持着单位数量的叶片。管道模型假说提出至今,在解释树木形态[3-4]、资源分配[5]、森林生产力评估[6]以及通过木质部横截面积预测茎段末端叶面积和干物质量[7-9]等方面发挥了重要的作用。这些研究都说明了这个假说的合理性。然而,通过管道理论研究单株林木的水分运输效率及其调控机制的研究鲜有报道。水力结构理论形成于20世纪70年代[10],它是指植物在特定的环境条件下,为适应生存竞争的需要所形成的不同的形态结构和水分运输供给策略。通常用导水率(Kh),比导率(Ks),叶比导率(Kl)和胡伯尔值(Hv)等参数来描述。本研究拟通过对刺槐Robinia pseudoacacia 不同生长时期水力结构参数的测定探讨管道理论及其适用性。
-
选择正常生长且均匀一致的刺槐苗木(2年生)共20株,于2007年11月移入花盆(直径23 cm×高度26 cm)栽植后正常培养,并于2008年5月开始处理苗木,充分供水(土壤水分保持在80%田间持水量),并测定相关生理参数。
-
苗木枝条水力结构参数测定采用改良的“冲洗法”测定[11],即在一定压力梯度下用特定溶液冲洗被测苗木茎段内部的栓塞,单位时间通过茎段横截面的溶液量则能反映被测茎段内部的栓塞程度,进而计算相应的水力结构参数。测定时,将被测苗木截成3 cm左右长的茎段,迅速用湿毛巾包裹拿回实验室,同时测定各个茎段的直径和长度,直径取上下端直径的平均值,茎末端叶片干质量用烘干称量法(105 ℃,8 h)测定。为避免导水率在实验过程中下降,冲洗溶液用0.1 mol·L-1草酸溶液,压力梯度(dp·dx-1)定义为加载茎段末端的压力(P)与茎段长度(L)之比。由于本次研究材料使用的2年生刺槐苗木尚未形成明显的边材与心材,故以枝条直径代替功能木质部直径。
-
苗木水力结构参数为导水率(Kh),比导率(Ks),叶比导率(Kl)和胡伯尔值(Hv),其值均由“冲洗法”测定的离体茎段水流量F计算得出[11-12]。
导水率(Kh)为通过一个离体茎段的水流量(F,kg·s-1)与该茎段2端压力梯度(dp·dx-1,MPa·m-1)的比值:Kh=F/( dp·dx-1)。比导率(Ks)为导水率与茎段边材横截面积(S,m2)的比值。它标志该茎段孔隙值的大小。Ks=Kh/S。叶比导率(Kl)为导水率(Kh)与茎段末端叶面积(A,m2)或叶干质量(W,g)的比值:Kl =Kh/A(W)。胡伯尔值(Hv)为边材横截面(或茎横截面)与茎末端的叶面积(或叶干质量)的比值:Hv=S/A(W)。
-
所测数据用Excel统计软件来处理。
-
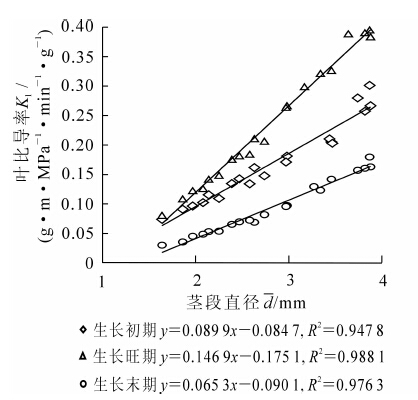
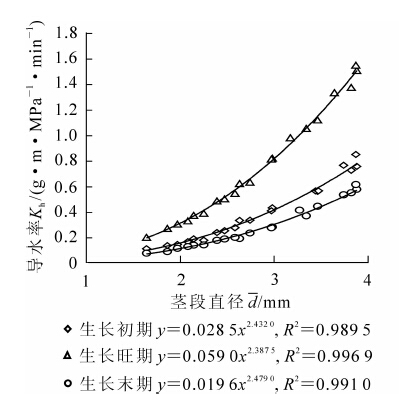
由图 1可知:在正常水分条件下,刺槐在不同生长时期,其木质部导水率均随着茎段直径增大而增加。以生长初期为例,刺槐茎段直径在1.64~3.88 mm范围内,其木质部导水率的变化范围为0.11~0.75 g·m·MPa-1·min-1,这是因为较粗茎段中含有更多的输水组织,因此导水能力较强,两者关系可用指数函数较好拟合。苗木不同生长阶段茎段导水率亦存在差异。其中,生长旺期导水率最大,生长初期次之,生长末期最小。

图 1 茎段直径与苗木导水率的关系
Figure 1. Relationships of segment diameter with Kh
-
比导率(Ks)是指单位茎段边材横截面积的导水率。在茎段边材横截面积一定的情况下,Ks越大,该部分输水效率越高,单位面积的输水能力则越强。由图 2可知:在正常水分条件下,刺槐苗木在不同生长时期,比导率均随茎段直径的增加而增加。以生长初期为例,刺槐苗木茎段直径从1.64 mm增大到3.88 mm,而其比导率则从5.09 g·m·MPa-1·min-1·cm-2增加到6.36 g·m·MPa-1·min-1·cm-2。刺槐苗木不同生长阶段其茎段比导率大小顺序为:生长旺期>生长初期>生长末期。这表明随着苗木的生长,生理活性逐渐加强,其木质部导管的输水效率也越来越高,至8月份生长旺期达到最大值,之后随苗木生理代谢活动的减弱,木质部导管输水效率也随之下降。

图 2 茎段直径与苗木比导率的关系
Figure 2. Relationships of segment diameter with Ks
-
叶比导率(Kl)是指示茎段末端叶片供水状况的重要指标。Kl值越大,表明单位叶面积的供水状况越好。如图 3所示:正常水分供应条件下,刺槐苗木叶比导率随茎段直径的增加而增加。生长初期枝条直径由1.64 mm增加到3.88 mm时,其叶比导率则由0.07 g·m·MPa-1·min-1·g-1增加到0.27 g·m·MPa-1·min-1·g-1。表明直径较大的茎段对茎末端叶片的供水能力强于直径较小的茎段。另外,根据导水率公式及蒸腾速率的计算公式可以得出:dp·dx-1=E/Kl。其中E为蒸腾速率。从式中可以看出:在蒸腾速率相同的情况下,苗木茎段的Kl越大,供给茎段末端叶片水分所需压力梯度就越小。例如,生长初期刺槐苗木直径为3.88 mm的茎段其叶比导率为0.27 g·m·MPa-1·min-1·g-1,约为直径1.64 mm茎段的叶比导率(0.07 g·m·MPa-1·min-1·g-1)的3.6倍。这表明在相同蒸腾速率的情况下,直径为3.88 mm的茎段给叶片供水所需压力梯度为直径1.64 mm茎段的1/3.6。如果在干旱条件下,直径较大的茎段末端的叶片会更容易获得水分,从而保证生物量积累较多的重要器官不受干旱胁迫造成的损害。
从苗木不同生长阶段来看,处于生长旺期的刺槐苗木叶比导率达最大值,其次为生长初期,生长末期最小。这表明随着苗木的生长,茎段对其末端叶片的供水效率逐渐增大。此后,在苗木逐渐进入休眠状态的过程中,枝条对其末端叶片的供水效率又恢复到较低水平状态。

图 3 茎段直径与苗木叶比导率的关系
Figure 3. Relationships of segment diameter with Kl
-
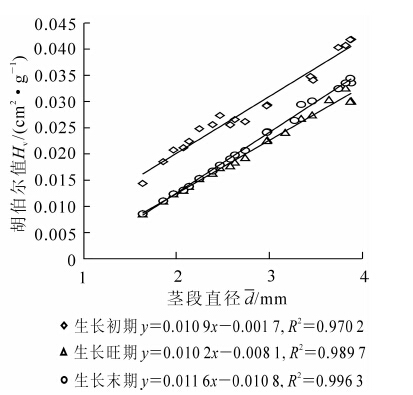
胡伯尔值(Hv)是指维持每单位叶面积或叶干质量所投入的茎段组织的多少。胡伯尔值越大,说明维持单位叶面积水分供给所需的茎干组织越多。不同植物的胡伯尔值存在差异。由上述定义还可得出:Kl=Hv·Ks。由式可知:植物供水情况取决于两部分:①发达的输水组织。即茎段和枝条的直径和长度;②输水组织具有较高的输水效率。由图 4可知:刺槐苗木胡伯尔值随茎段直径的增加而增大。例如,在生长初期,刺槐苗木茎段直径由1.64 mm增加到3.88 mm,其相应的胡伯尔值则由0.01 cm2·g-1增加到0.05 cm2·g-1。这表明随着枝条直径的增加,苗木枝条维持单位叶干质量所投入的组织量也相应增多,而这对于处于逆境中的植物是非常重要的,说明植物更倾向于保护较粗茎段。由图 4还可看出:刺槐苗木生长初期胡伯尔值均大于生长旺期及生长末期,生长旺期和生长末期差异不显著(P>0.05)。这是由于生长初期的苗木刚开始展叶时枝条直径生长相对缓慢且叶量较少所致。

图 4 茎段直径与苗木胡伯尔值的关系
Figure 4. Relationships of segment diameter with Hv
-
自20个世纪70年代初期水力结构理论建立以来[10],随着研究的不断深入,特别是近些年来树木解剖学特征与林木生理生态学的相结合,利用水力结构理论来探索树木对干旱胁迫的适应机制和策略已成为林木生理生态领域的热点问题[13-17]。本次研究通过测定不同生长阶段刺槐苗木枝条的水力结构参数,发现苗木水力结构参数均随枝条直径的增加而增大。这个结果与笔者在樟树Cinnamomum camphora 苗木[18]以及翟洪波等[19]在油松Pinus tabuliformis和侧柏Platycladus orientalis上的研究结果相同。这说明直径较大和生物量积累较多的枝条更容易获得水分,可能是植物在长期进化过程中形成的自我保护机制[20]。在干旱胁迫条件下,植物总是首先牺牲那些直径较小,生物量积累较少的枝条或侧枝,以保护那些生物量积累较大的主枝部分[10]。
从本次研究结果来看,刺槐苗木不同生长时期,其导水率、比导率和叶比导率均为生长旺期最大,生长初期次之,生长末期最小。本研究结构与李晶等[21]在3种锦鸡儿属Caragana植物上的研究结果类似。这说明刺槐苗木水力结构特征与其体内生理活性和器官发育成熟度呈正相关关系,树木个体在整个生长发育期间可以根据外界环境条件来改变其水力结构特征从而影响其导水阻力和水分需求,以适应外界的不利环境条件[22-24]。
根据对刺槐苗木水力结构的研究结果,从以下几个方面探讨管道模型假说存在的问题:①根据管道模型假说,树木木质部是由均一的导管组成,由此推断各个导管对水分的运输效率应该是一样的,然而实验结果并非如此。苗木各部分木质部并不是由均一的单位导管组成,相对细小茎段而言,较粗茎段功能木质部的成熟度和导水效率要高得多。②单位管道模型假说提出树干内单位管道支持着单位数量的叶片,由此可知树木上的所有叶片都具有相同的获得水分的能力,即树木各个部分的Kl应该是相等的,然而实际上不同直径的茎段其Kl值相差较大。③根据本研究的实验结果,刺槐苗木不同部位的胡伯尔值也有较大差异,说明了不同直径的茎段对水分运输所投入的输水组织量并不是均匀的,即树木对较粗的枝条投入了相对较多的输水组织。④在本次研究中,刺槐苗木茎段水力结构参数的季节变化表明,林木可以通过对体内水力结构特征的调整来适应外界环境的变化。有研究显示,树木通过最佳的水力结构来防止木质部空穴和栓塞的发生,从而保证树木在逆境条件下保持木质部水分的正常运输[25]。而管道理论对林木体内水分运输过程及其调控机制解释尚显不足。综上所述,管道模型假说虽然有其存在的合理性,并取得了一定的研究成果,但也有其局限性和适用范围,尤其是在对于林木体内水分的运输过程及其调控机制的解释上,管道模型假说显然过于简单化了,它并无助于人们对林木体内水分运输过程的正确理解。本研究结果同Zhai等[26]在油松上的研究结果较为一致。
-
刺槐苗木水力结构参数值均随枝条直径的增加而增大。在水分胁迫条件下,苗木更倾向于首先保护那些较粗的茎段。刺槐苗木水力结构特征与其体内生理活动强度呈正相关关系,且可根据外界环境条件来调整自己的水力结构特征。管道理论有其存在的合理性和适用范围,在解释林木体内水分运输过程及其调控机制上并不适用。
The pipe model theory based on the hydraulic architecture parameter with Robinia pseudoacacia seedlings
-
摘要: 为了解刺槐Robinia pseudoacacia苗木对体内水分运输过程的调控机制,基于水力结构理论,在充分供水条件下,利用改良的"冲洗法"对2年生刺槐苗木的水力结构参数进行测定,并探讨了管道理论的合理性和适用范围。结果表明:在充分供水条件下,刺槐苗木茎段导水率、比导率、叶比导率和胡伯尔值均随被测茎段直径的增加而增大;苗木不同生长阶段,其导水率、比导率和叶比导率差异显著(P<0.05),且均为生长旺期>生长初期>生长末期,胡伯尔值为生长初期最大,生长旺期和生长末期差异不显著(P>0.05)。由此可推断,①在水分胁迫条件下,苗木更倾向于保护那些较粗茎段;②苗木水力结构特征与提体内生理活动呈正相关关系;③管道模型假说虽然有其存在的合理性,但也有其局限性和适用性,在对于水分在林木体内的运输过程以及调控机制的解释,管道模型假说过于简单化了。Abstract: To better understand the pipe model theory and hydraulic architecture characteristics of Robinia pseudoacacia seedlings in different growth stages (early, fast, and last), the improved flushing method was used to measure the hydraulic architecture parameters of two-year-old R.pseudoacacia seedlings with an adequate water supply for three growth stages (early, fast, and last) and there are 20 repeats in every growth stages.Significance tests were done between hydraulic architecture parameters and stem segment diameter and different growth stages by ANOVA.Then the pipe theory was discussed based on hydraulic architecture theory.Results showed that with sufficient water, the hydraulic conductivity (Kh), stem area specific hydraulic conductivity (Ks), leaf specific conductivity (Kl), and Huber value (Hv) of R.pseudoacacia seedlings increased (P<0.05) as the stem segment diameter enlarged.For the three seedling growth stages, Kh, Ks, and Kl were significantly different (P<0.05) with early growth stage> fast growth stage> last growth stage.Hv was greatest (P>0.05) in the early growth stage with no significant differences in fast and last growth stages.Thus, (1) with sufficient water, water transportation efficiency of thicker stems is greater than the thinner stems; (2) hydraulic architecture characteristics of seedlings were different (P<0.05) in three growth stages; and (3) the pipe model theory was not perfect, especially for understanding the inner water transportation processes of a tree and their control mechanism.
-
[1] SHINOZAKI K, YODA K, HOZUMI K, et al.A quantitative analysis of plant form the pipe model theory(Ⅰ):basic analysis[J].Jpn J Ecol, 1964, 14(3):97-105. [2] SHINOZAKI K, YODA K, HOZUMI K, et al.A quantitative analysis of plant form the pipe model theory(Ⅱ):further evidence of the theory and its application in forest ecology[J].Jpn J Ecol, 1964, 14(4):133-139. [3] 刘盛,李国伟.基于管道模型理论的树形结构分析[J].东北林业大学学报, 2006,34(6):15-16. LIU Sheng, LI Guowei.Tree structure based on pipe model theory[J].J Northeast For Univ, 2006, 34(6):15-16. [4] 殷晖,关文彬.基于树体管道模型的林冠结构对降水截留的影响[J].科技通报,2012,28(5):89-93,98. YIN Hui, GUAN Wenbin.The effect of rainfall interception by canopy structure based on pipe model[J].Bull Sci Technol, 2012, 28(5):89-93, 98. [5] 刘盛,刘成,张士民.基于管道模型的榆树水分年轮输导模式研究[J].北华大学学报:自然科学版,2002,3(5):438-440. LIU Sheng, LIU Cheng, ZHANG Shimin.On the water transportation along Ulmus pumila L.growth ring based on the pipe model theory[J].J Beihua Univ Nat Sci, 2002, 3(5):438-440. [6] 彭孝飞,王百田,迟璐,等.油松林木地上部分生物量研究[J].广东农业科学,2012,39(1):42-45. PENG Xiaofei, WANG Baitian, CHI Lu, et al.Research on the aboveground biomass of Pinus tabulaeformis[J].Guangdong Agric Sci, 2012, 39(1):42-45. [7] CHIBA Y.Architectural analysis of relationship between biomass and basal area based on pipe model theory[J].Ecol Model, 1998, 108(1):219-225. [8] 刘盛,刘成.长白落叶松水分输导模式及叶生物量估测方法[J].东北林业大学学报, 2005,33(5):35-37. LIU Sheng, LIU Cheng.The model of water transportation and the estimation methods of leave biomass for Larix olgensis[J].J Northeast For Univ, 2005, 33(5):35-37. [9] 杨洪国,李治宇.应用管道模型原理建立柏木桤木叶面积叶量估测模型[J].四川林勘设计,2012,106(1):35-37. YANG Hongguo, LI Zhiyu.Estimation models for leaf area and amount of cedar wood and alder based on the pipeline model[J].Sichuan For Expl Des, 2012, 106(1):35-37. [10] ZIMMERMANN M H.Hydraulic architecture of some diffuse-porous trees[J].Can J Bot, 1978, 56(18):2286-2295. [11] SPERRY J S,DONNELLY J R, TYREE M T.A method for measuring hydraulic conductivity and embolism in xylem[J].Plant Cell&Environ, 1988, 11(1):35-40. [12] ZOTZ G, TYREE M T, PATINO S.Hydraulic architecture and water relations of a flood-tolerant tropical tree, Annona glabra[J].Tree Physiol, 1997, 17(6):359-365. [13] TYREE M T, EWERS F W.The hydraulic architecture of tree and other woody plants[J].New Phytol, 1991, 119(3):345-360. [14] 孙青,郭锐,沈繁宜,等.木本植物木质部栓塞修复机制的探讨[J].北京林业大学学报,2007,29(5):94-98. SUN Qing, GUO Rui, SHEN Fanyi, et al.Discussion on the embolism repairing in xylem of woody plants[J].J Beijing For Univ, 2007, 29(5):94-98. [15] KLEIN T, COHEN S, YAKIR D, et al.Hydraulic adjustments underlying drought resistance of Pinus halepensis[J].Tree Physiol, 2011, 31(6):637-648. [16] ROSSI L,SEBASTIANI L,TOGNETTI R, et al.Tree-ring wood anatomy and stable isotopes show structural and functional adjustments in olive trees under different water availability[J].Plant Soil, 2013,372(11):567-579. [17] 樊大勇,谢宗强.木质部导管空穴化研究中的几个热点问题[J].植物生态学报,2004,28(1):126-132. FAN Dayong, XIE Zongqiang.Several controversial viewpoints in studying the cavitation of xylem vessels[J].China J Plant Ecol, 2004, 28(1):126-132. [18] 王丁,姚健,薛建辉.土壤干旱胁迫对樟树Cinnamomum camphora (L.) Presl苗木水力结构特征的影响[J].生态学报,2009,29(5):2725-2731. WANG Ding, YAO Jian, XUE Jianhui.Effects of soil drought stress on hydraulic architecture characteristics of camphor seedlings[J].Acta Ecol Sin, 2009, 29(5):2725-2731. [19] 翟洪波,李吉跃,姜金璞,等.油松、侧柏苗木水力结构特征的对比研究[J].生态学报,2002,22(11):1890-1895. ZHAI Hongbo, LI Jiyue, JIANG Jinpu, et al.Comparative study on hydraulic architecture characteristic of Pinus tabulaeformis and Platycladus orientalis seedlings[J].Acta Ecol Sin, 2002, 22(11):1890-1895. [20] 刘晓燕,李吉跃,翟洪波.从树木水力结构特征探讨植物耐旱性[J].北京林业大学学报,2003,25(3):48-54. LIU Xiaoyan, LI Jiyue, ZHAI Hongbo, et al.Discussion on drought resistance through hydraulic architecture of trees[J].J Beijing For Univ, 2003, 25(3):48-54. [21] 李晶,高玉葆,郑志荣,等.内蒙古高原不同生境3种锦鸡儿属植物的水力结构特征及其对环境因子的响应[J].生态学报,2007,27(3):837-845. LI Jing, GAO Yubao, ZHENG Zhirong, et al.The hydraulic architecture of three Caragana species and its relationship with environmental factors in different habitats of the Inner Mongolia Plateau[J].Acta Ecol Sin, 2007, 27(3):837-845. [22] YANG Qiliang, ZHANG Fuang, LI Fusheng, et al.Hydraulic conductivity and water-use efficiency of young pear tree under alternate drip irrigation[J].Agric Water Manage, 2013, 119(3):80-88. [23] 魏亚冉,张侦珍,李清芳,等.珍珠梅和榆叶梅水力结构特征比较研究[J].天津师范大学学报:自然科学版,2011,31(1):71-74. WEI Yaran, ZHANG Zhenzhen, LI Qingfang, et al.Comparative study on hydraulic architecture parameters of Sorbaria sorbifolia A.Br and Prunus triloba Lindl.[J].J Tianjin Norm Univ Nat Sci Ed, 2011, 31(1):71-74. [24] IOGNA P A, BUCCI S J, SCHOLZ F G, et al.Water relations and hydraulic architecture of two patagonian steppe shrubs:effect of slope orientation and microclimate[J].J Arid Environ, 2011, 75(9):763-772. [25] 刘娟娟,李吉跃,张建国.干旱胁迫对油松和侧柏水分运输安全性和有效性的影响[J].生态学报,2010,30(9):2507-2514. LIU Juanjuan, LI Jiyue,ZHANG Jianguo.Influences of drought stress on hydraulic safety and efficiency in the saplings of Pinus tabulaeformis and Platycladus orientalis[J].Acta Ecol Sin, 2010, 30(9):2507-2514. [26] ZHAI Hongbo, LI Jiyue, NIE Lishui.Discussion on pipe model through hydraulic architecture of Pinus tabulaeformis seedling[J].Chin For Sci Technol, 2004, 3(1):53-58. -

-
链接本文:
https://zlxb.zafu.edu.cn/article/doi/10.11833/j.issn.2095-0756.2014.03.024
 点击查看大图
点击查看大图
图(4)
计量
- 文章访问数: 3309
- HTML全文浏览量: 658
- PDF下载量: 479
- 被引次数: 0
