




-
农业生产中化感作用的存在会对种子萌发、幼苗生长、根系发育,甚至作物产量和品质等造成严重影响[1-3]。番茄Lycopersicon esculentum作为世界上普遍栽培的果蔬之一,其化感作用的研究已受到国内外学者的广泛重视。研究表明:番茄植株和种子浸提液、茎叶等不同器官提取物、根系分泌物、地上部挥发物以及植株残体腐解物中普遍存在化感物质[4-9]。番茄秸秆是农业生产系统中重要的园艺类生物资源,但由于化感作用的存在导致很难对其进行直接利用。通过秸秆堆制过程产生的高温分解化感物质,是削减其化感作用的重要途径[10-11]。有研究表明:番茄秸秆单独堆制或与牛粪共堆后均能显著降低其自毒作用[12-13],但堆制后对其他作物化感效应的研究还少见报道。本研究选取常见的蔬菜类型黄瓜Cucumis sativus和大白菜Brassica campestris ssp. pekinensis为受体对象,通过将番茄秸秆与醋槽进行共堆后,分析了其浸提液分别对黄瓜和大白菜种子萌发和幼苗生长的影响,以期初步明确堆制番茄秸秆的化感效应及效应强度,为进一步提高番茄秸秆资源的综合利用提供参考依据。
-
番茄秸秆来源于试验地周边,腐熟醋槽来源于恒顺生物科技有限公司;供试的黄瓜种子品种为‘津优1号’‘Jinyou No. 1’(天津科润农业股份有限公司提供),供试的大白菜种子品种为‘快菜208’‘Kuaicai 208’(南京金盛达种子有限公司提供)。
-
首先,将番茄秸秆原料用粉粹机进行粉碎,粉碎标准为1~4 cm。由于番茄秸秆pH值较高,将它与pH值较低的腐熟醋槽配比(8 : 1)后进行发酵。采用静态通风式堆制,并定期进行翻堆。到发酵后期适时补充水分确保其完全腐熟(腐熟判断标准:堆肥32 d后堆肥温度保持在26 ℃左右没有发生明显变化且利用大白菜种子进行发芽实验发芽率达到85%及以上)。
-
称取10.0 g腐熟秸秆至装有100.0 mL蒸馏水的三角瓶中,利用180 r·min-1的摇床摇120 min后,在常温下用3层纱布浸提48 h后滤纸过滤得到100.0 g·L-1的浸提母液,存入冰箱待用。
-
选取颗粒饱满的黄瓜和大白菜种子,用体积分数为75%的乙醇溶液处理5 s后,再用蒸馏水将种子冲洗干净后供试验用。
-
试验共设置5组处理:用100.0 g·L-1浸提母液梯度稀释成10.0,20.0,30.0,40.0和50.0 g·L-1的浸提液。另外,以清水为对照组(ck)。将预处理过的黄瓜种子(30粒)和大白菜种子(50粒)分别放入垫有2层滤纸、直径为9 cm的培养皿中,分别加入5.0 mL不同质量浓度的基质浸提液和清水,将种子均匀铺好后置于25 ℃恒温培养箱中培养。每组处理和对照组重复3次,间隔1 d定期向培养皿中加入5.0 mL浸提液或清水。每天记录各处理和对照组的发芽情况(以种子芽长达到其种子长度的一半作为黄瓜发芽标准,以种子芽长达0.2 cm及以上作为大白菜发芽标准),最后计算发芽率和发芽指数;培养5 d后随机选取10株植株,利用直尺(规格20 cm)分别测定植株的苗长(即地上部分)和根长,测量植株鲜质量。
-
发芽率(%)=(发芽种子数/供试种子数)×100%;发芽指数(GI)=∑(Gt/Dt)。其中:Gt为第t天的发芽数,Dt为发芽天数。
化感效应指数(IR)的计算参照WILLIAMSON等[14]的方法进行:当T≥C时,IR=1-C/T;当T<C时,IR=T/C-1,其中C为对照值,T为处理值。当IR>0时表示化感促进作用,当IR<0时表示化感抑制作用,其绝对值的大小与作用强度保持一致。
利用Excel 2010整理数据、作图,利用SPSS 19.0软件进行单因素方差分析及组间差异的Duncan氏多重比较。
-
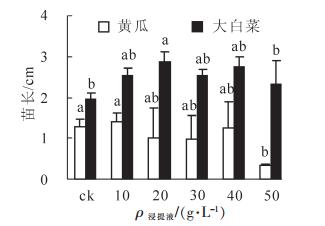
由图 1和表 1可知:不同质量浓度番茄秸秆浸提液对黄瓜和大白菜的发芽率、发芽指数的化感效应不尽相同。随着质量浓度的增加,浸提液对黄瓜发芽率表现为低质量浓度促进高浓度抑制的双重作用,浸提液质量浓度为10.0 g·L-1时,黄瓜发芽率为100%,高于对照,化感效应指数为0.022,当浸提液质量浓度增加时发芽率呈现下降趋势且低于对照,表现为抑制作用,但不同质量浓度处理与对照相比差异均不显著。不同质量浓度番茄秸秆浸提液对黄瓜发芽指数均表现为单一的抑制作用,质量浓度为10.0 g·L-1时,黄瓜发芽指数为30.3,化感效应指数为-0.037,随着质量浓度增加,发芽指数逐渐降低,抑制作用越大。但不管对于黄瓜发芽率还是发芽指数,不同浸提液质量浓度处理间的化感效应差异均未达到显著水平。

图 1 不同质量浓度番茄秸秆浸提液对黄瓜和大白菜种子发芽率及发芽指数的影响
Figure 1. Effects of different concentration of the water extractive from tomato residues on germination rate and germination index of cucumber and Chinese cabbage seeds
表 1 不同质量浓度番茄秸秆浸提液对黄瓜和大白菜种子发芽和幼苗生长的化感效应指数的影响
Table 1. Effects of different concentration of the water extractive from tomato residues on allelopathy of the germination and growth of cucumber and Chinese cabbage
ρ/(g·L-1) 对黄瓜的化感效应指数 对大白菜的化感效应指数 发芽率 发芽指数 苗长 根长 鲜质量 发芽率 发芽指数 苗长 根长 鲜质量 10.0 0.022 a -0.037 a 0.078 a -0.192 a -0.008 a 0.021 a 0.027 ab 0.217 a -0.050 a 0.322 a 20.0 0 a -0.042 a -0.230 ab -0.312 a -0.381 b 0.061 a 0.071 a 0.317 a -0.172 a 0.492 a 30.0 -0.011 a -0.043 a -0.203 ab -0.264 a -0.353 b 0.014 a 0.028 b 0.221 a -0.191 a 0.217 a 40.0 -0.011 a -0.056 a -0.087 ab -0.278 a -0.112 ab -0.072 b -0.069 b 0.277 a -0.197 a 0.481 a 50.0 -0.023 a -0.066 a -0.753 b -0.527 b -0.560 b -0.007 ab -0.007 ab 0.127 a -0.251 a 0.142 a 说明:同列中不同字母表示差异显著 (P<0.05) 随着浸提液质量浓度的增加,番茄秸秆浸提液对大白菜发芽率和发芽指数均表现为低促高抑作用;当质量浓度为10.0~30.0 g·L-1,对大白菜发芽率以及发芽指数均表现为促进作用,且均在20.0 g·L-1时达到最大,发芽率以及发芽指数分别为98.7%和52.0,其化感效应指数分别为0.061和0.071,当浸提液质量浓度高于40.0 g·L-1时则开始表现为抑制作用;而且,除了在40.0 g·L-1浸提液处理的发芽率以及20.0 g·L-1浸提液处理的发芽指数与对照差异显著(P<0.05)外,其余均无显著差异。
-
由图 2和表 1可知:不同质量浓度番茄秸秆浸提液对黄瓜和大白菜苗长的影响不同。就黄瓜而言,表现为低促高抑的双重效应,质量浓度为10.0 g·L-1时,苗长较对照高1.29 cm,表现为促进作用,化感效应指数为0.078;当质量浓度为20.0~50.0 g·L-1时,表现为抑制作用,且以50.0 g·L-1的抑制作用最强,对黄瓜苗长的抑制达到显著水平(P<0.05)。对大白菜而言,10.0~50.0 g·L-1各质量浓度浸提液均表现为促进作用,且随浸提液质量浓度的增加,促进作用越明显,但当质量浓度继续增加时其促进作用又有所下降;以20.0 g·L-1处理的苗长较对照增加最多,各质量浓度除了50.0 g·L-1浸提液处理的苗长与对照差异不显著外,其余各质量浓度处理的苗长较对照差异均显著(P<0.05),但不同浸提液质量浓度处理间的化感效应差异均未达到显著水平。

图 2 不同质量浓度番茄秸秆浸提液对黄瓜和大白菜苗长的影响
Figure 2. Effects of different concentration of the water extractive from tomato residues on shoot length cucumber and Chinese cabbage
-
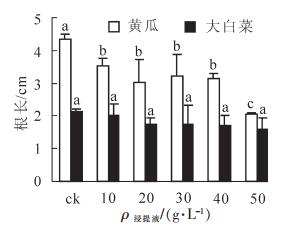
由图 3和表 1可知:不同质量浓度番茄秸秆浸提液对黄瓜和大白菜根长均表现为化感抑制作用,但抑制作用强弱不同。其中,均以50.0 g·L-1质量浓度下根长的化感效应指数绝对值最大(黄瓜和大白菜根长化感效应指数分别为0.527和0.251),说明该浸提液质量浓度的抑制作用最强。与对照相比,10.0~50.0 g·L-1浸提液处理的黄瓜幼苗根长同比对照分别下降了19.3%,31.0%,26.6%,27.8%和52.6%,各处理均较对照达到显著差异水平(P<0.05),但10.0~40.0 g·L-14个质量浓度处理间的差异不显著;而大白菜幼苗根长同比对照分别下降了4.7%,17.4%,18.8%,20.2%和25.8%,各处理的根长与对照差异均不显著。整体来看,不同质量浓度浸提液下黄瓜根长的化感效应指数绝对值(0.192~0.527)均高于相应质量浓度下大白菜的各值(0.050~0.251),说明番茄秸秆浸提液对黄瓜根长的抑制作用大于大白菜。

图 3 不同质量浓度番茄秸秆浸提液对黄瓜和大白菜根长的影响
Figure 3. Effects of different concentration of the water extractive from tomato residues on root length cucumber and Chinese cabbage
-
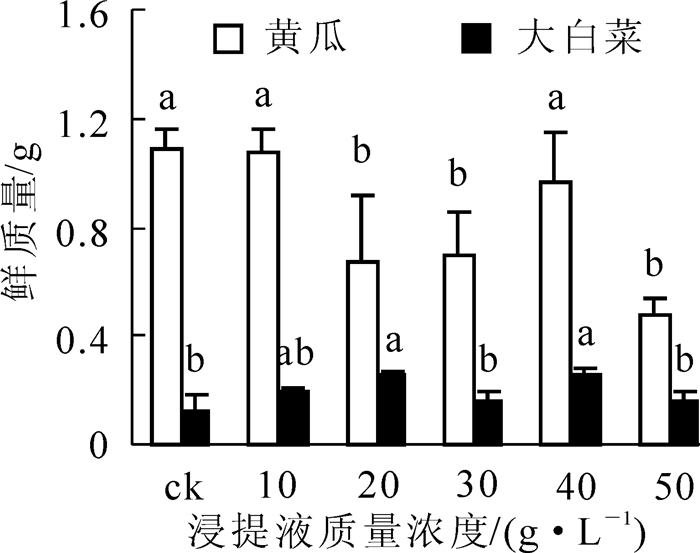
由图 4和表 1可知:不同质量浓度番茄秸秆浸提液对黄瓜和大白菜鲜质量则表现为相反的化感作用,对黄瓜鲜质量均是抑制作用,而对大白菜鲜质量则均是促进作用。与对照相比,20.0,30.0和50.0 g·L-1处理的黄瓜鲜质量显著降低(P<0.05),分别下降了38.6%,36.0%和56.1%;而10.0,20.0和40.0 g·L-1处理的大白菜鲜质量显著增加(P<0.05),分别增加了51.4%,100%和98.2%。对黄瓜鲜质量抑制作用最强的浸提液质量浓度为50.0 g·L-1,最弱的为10.0 g·L-1,其化感效应指数分别为-0.560和-0.008,但不同质量浓度间差异不显著。对大白菜鲜质量促进作用最强的浸提液质量浓度为20.0 g·L-1,其化感效应指数接近0.500,但不同处理间均未表现显著差异。

图 4 不同质量浓度番茄秸秆浸提液对黄瓜和大白菜鲜质量的影响
Figure 4. Effects of different concentration of the water extractive from tomato residues on fresh weight of cucumber and Chinese cabbage
-
小麦Triticum aestivum,棉花Anemone vitifolia,大蒜Allium sativum和玉米Zea mays等作物秸秆及其腐解物质可以通过化感物质的释放对作物生长产生促进或抑制的化感作用[15-17]。周志红等[8]研究结果显示:番茄植株浸提液对黄瓜具有显著的抑制作用。莫云容等[18]的研究证实番茄植株浸提液对大白菜也具有一定的抑制作用。堆制后的番茄秸秆可在一定程度上减轻其化感抑制作用。LI等[10]和WAKJIRA等[11]分别研究了堆制后紫茎泽兰Eupatorium adenophora和外来杂草对其他植物的化感作用,发现经过堆制发酵后其化感物质和对其他植物的影响均显著减少。尽管如此,本研究表明:堆制后的番茄秸后的番茄秸秆仍具有一定的化感抑制作用,同时也表现一定的化感促进作用,但其作用强度受浸提液质量浓度的影响,也与受体对象密切相关。对大白菜种子发芽率和发芽指数,番茄秸秆浸提液表现为低质量浓度(10.0~30.0 g·L-1)促进而高量质浓度(>40.0 g·L-1)抑制的双重效应;对大白菜幼苗苗长和鲜质量、幼苗根长,各质量浓度均表现为单一促进作用或抑制作用。对黄瓜种子的发芽率及幼苗苗长,浸提液也表现为低(10.0~20.0 g·L-1)促高(>20.0 g·L-1)抑的双重效应;而对黄瓜发芽指数、幼苗根长和鲜质量则均表现为单一的抑制作用。这与不同质量浓度浸提液对大白菜的影响有所差异,并且各质量浓度浸提液对黄瓜根长的抑制作用(化感效应指数为-0.192~-0.527)要明显大于相同质量浓度下对大白菜根长的影响(化感效应指数为-0.050~-0.251)。
总体来看,堆制后番茄秸秆浸提液对黄瓜幼苗生长基本表现出一定的抑制作用,对大白菜幼苗生长则表现出一定程度的促进作用,但两者均未达到显著水平,并且对根长的抑制作用在2种受体中均得到了体现。这可能是因为根部作为植物水分和营养物质的吸收器官,直接与环境中的化感物质接触有关。本研究与其他对同类作物的研究结果[6, 8, 18-19]总体相近,但也存在差异。究其原因,可能是由于不同番茄品种的秸秆浸提液具有不同的化感效应所致[17, 20],也有可能与共堆肥调理剂的选择及堆制方式有关[21-22]。
研究表明:化感作用的机制是化感物质首先对细胞膜造成损害,通过细胞膜上的靶位点,将胁迫信号传送到细胞内,从而对离子、水分和激素等的吸收利用产生影响。这些影响会引起植物细胞分裂、伸长和亚显微结构以及光合作用等的变化,影响蛋白质的合成以及基因的表达,从而对种子萌发和幼苗生长产生抑制作用[23-24]。不同化感物质的作用机制也有所差异[25-27],如酚酸类化感物质可以破坏细胞膜的结构和透性,降低根系对离子的吸收能力;肉桂酸、对羟基苯甲酸等可以抑制植物线粒体的新陈代谢;苯甲酸、阿魏酸、香草酸等可以降低植物体内氨基酸的运输和氨基酸向蛋白质的合成速率,阻止DNA的翻译和转录,从而影响蛋白合成和基因表达。有研究证实,番茄植株体内含有一定的酚酸及其他有机酸等化感物质,这些化感物质可通过对细胞膜透性造成损伤以及对细胞膜防御系统产生破坏等对幼苗的生长产生影响[7-8]。而堆制过程中的高温可以分解一部分化感物质[28]从而减轻其化感效应。但有关堆制后番茄秸秆浸提液中化感物质的鉴定分析及更深层次的作用机制还有待进一步研究。
-
堆制番茄秸秆浸提液对黄瓜、大白菜种子萌发以及幼苗生长各指标具有不同的化感作用。
不同质量浓度番茄秸秆浸提液对黄瓜种子的发芽率和苗长均表现为低(10.0~20.0 g·L-1)促高(>20.0 g·L-1)抑的双重质量浓度效应;对黄瓜种子的发芽指数和幼苗鲜质量均表现为抑制作用。
不同质量浓度浸提液对大白菜种子发芽率和发芽指数表现为低(10.0~30.0 g·L-1)促高(>40.0 g·L-1)抑的效应;对幼苗的苗长和鲜质量则为促进效应,且以20.0 g·L-1浸提液浓度的促进作用最为明显。
不同质量浓度番茄秸秆浸提液对黄瓜和大白菜幼苗根的生长均有抑制作用,且对黄瓜根长的抑制作用大于大白菜。
Allelopathic effects of anaqueous extract from composted tomato residues on the growth of cucumber and Chinese cabbage
-
摘要: 以黄瓜Cucumis sativus和大白菜Brassica campestris ssp.pekinensis等2种蔬菜作为受体,研究堆制后不同质量浓度(10.0,20.0,30.0,40.0和50.0 g·L-1)番茄Lycopersicon esculentum秸秆浸提液对种子萌发和幼苗生长的影响,探讨堆制番茄秸秆的化感效应。结果表明:堆制后的番茄秸秆浸提液对黄瓜和大白菜种子的萌发和幼苗生长存在不同程度的化感作用。低质量浓度(10.0~20.0 g·L-1)浸提液可提高黄瓜和大白菜种子发芽率,促进苗长增长和鲜质量增加,而高质量浓度(40.0~50.0 g·L-1)则对黄瓜和大白菜的发芽率、发芽指数以及黄瓜幼苗苗长和鲜质量具有一定的抑制作用。但不论低质量浓度或高质量浓度浸提液对黄瓜和大白菜幼苗根的生长均表现为抑制作用,尤其对黄瓜根长的抑制作用达到显著水平(P < 0.05)。Abstract: To understand the allelopathic effects of composted tomato residues on cucumber and Chinese cabbage vegetables, five concentrations (10.0, 20.0, 30.0, 40.0, and 50.0 g·L-1) of extracts from composted tomato residues and a control (ck) (0.0 g·L-1) were applied to germinating seeds and seedlings during growth and then employed seed germination bioassay method to measure the germination rate, germination rate, seedling length, and fresh weight of cucumber and Chinese cabbage and finally evaluated. Results showed that the germination rate, seedling length, and fresh weight of cucumber and Chinese cabbage were promoted at low aqueous extract concentrations (10.0-20.0 g·L-1). Also, the germination rate and germination index of cucumber and Chinese cabbage as well as the seedling length and fresh weight of cucumber were inhibited at a high concentration (>40.0 g·L-1). Root growth of cucumber and Chinese cabbage for all concentrations was inhibited. Also, the root length of cucumber treated with extracts was significantly lower (P < 0.05) than ck. Overall, the extracts from composted tomato residues showed some inhibitory effect on cucumber seedling growth and a certain role in promoting on the growth of Chinese cabbage seedlings, but both of them did not reach the significant level. Thus, to a certain extent, the tomato residues could alleviate its inhibition effect after composting.
-
Key words:
- botany /
- tomato residues /
- compost /
- aqueous extract /
- allelopathy
-
图 1 不同质量浓度番茄秸秆浸提液对黄瓜和大白菜种子发芽率及发芽指数的影响
Figure 1 Effects of different concentration of the water extractive from tomato residues on germination rate and germination index of cucumber and Chinese cabbage seeds
图 2 不同质量浓度番茄秸秆浸提液对黄瓜和大白菜苗长的影响
Figure 2 Effects of different concentration of the water extractive from tomato residues on shoot length cucumber and Chinese cabbage
图 3 不同质量浓度番茄秸秆浸提液对黄瓜和大白菜根长的影响
Figure 3 Effects of different concentration of the water extractive from tomato residues on root length cucumber and Chinese cabbage
图 4 不同质量浓度番茄秸秆浸提液对黄瓜和大白菜鲜质量的影响
Figure 4 Effects of different concentration of the water extractive from tomato residues on fresh weight of cucumber and Chinese cabbage
表 1 不同质量浓度番茄秸秆浸提液对黄瓜和大白菜种子发芽和幼苗生长的化感效应指数的影响
Table 1. Effects of different concentration of the water extractive from tomato residues on allelopathy of the germination and growth of cucumber and Chinese cabbage
ρ/(g·L-1) 对黄瓜的化感效应指数 对大白菜的化感效应指数 发芽率 发芽指数 苗长 根长 鲜质量 发芽率 发芽指数 苗长 根长 鲜质量 10.0 0.022 a -0.037 a 0.078 a -0.192 a -0.008 a 0.021 a 0.027 ab 0.217 a -0.050 a 0.322 a 20.0 0 a -0.042 a -0.230 ab -0.312 a -0.381 b 0.061 a 0.071 a 0.317 a -0.172 a 0.492 a 30.0 -0.011 a -0.043 a -0.203 ab -0.264 a -0.353 b 0.014 a 0.028 b 0.221 a -0.191 a 0.217 a 40.0 -0.011 a -0.056 a -0.087 ab -0.278 a -0.112 ab -0.072 b -0.069 b 0.277 a -0.197 a 0.481 a 50.0 -0.023 a -0.066 a -0.753 b -0.527 b -0.560 b -0.007 ab -0.007 ab 0.127 a -0.251 a 0.142 a 说明:同列中不同字母表示差异显著 (P<0.05)  下载: 导出CSV
下载: 导出CSV
-
[1] KIMBER R W L. Phytotoxicity from plant residues (Ⅱ) the effect of time of rotting of straw from some grasses and legumes on the growth of wheat seedlings[J]. Plant Soil, 1973, 38(2):347-361. [2] 李玉文.化学生态学研究现状和进展 (Ⅱ) 植物他感作用和矿质养分化学生态[J].东北林业大学学报, 1999, 27(1):56-59. LI Yuwen. The situation and development of chemical ecology (Ⅱ) allelopathy and ecology of the mineral nutrients[J]. J Northeast For Univ, 1999, 27(1):56-59. [3] 王建花, 陈婷, 林文雄.植物化感作用类型及其在农业中的应用[J].中国生态农业学报, 2013, 21(10):1173-1183. WANG Jianhua, CHEN Ting, LIN Wenxiong. Plant allelopathy types and their application in agriculture[J]. Chin J Eco-Agric, 2013, 21(10):1173-1183. [4] 康亚龙, 刘彦荣, 刘建国, 等.加工番茄植株浸提液对受体作物幼苗生长及酶活性的影响[J].西北农业学报, 2014, 23(7):120-130. KANG Yalong, LIU Yanrong, LIU Jianguo, et al. Effects of different organ aqueous of processing tomato on crop seeding growth and enzyme activities[J]. Acta Agric Boreal-Occident Sin, 2014, 23(7):120-130. [5] YU Jingquan, LEE K S, MATSUI Y. Effect of the addition of activated charcoal to the nutrient solution on the growth of tomato in hydroponic culture[J]. Soil Sci Plant Nut, 1993, 39(1):13-22. [6] 王广印, 韩世栋, 张建伟, 等.番茄种子及其萌发期化感作用分析[J].植物资源与环境学报, 2013, 22(1):94-101. WANG Guangyin, HAN Shidong, ZHANG Jianwei, et al. Analysis on allelopathy of seed and its germination stage of Lycopersicon esculentum[J]. J Plant Resour Environ, 2013, 22(1):94-101. [7] KIM Y S, KIL B S. Identification and growth inhibition of phytotoxic substances from tomato plant[J]. Korean J Bot, 1989, 32(1):41-50. [8] 周志红, 骆世明, 牟子平.番茄植株中几种化学成分的化感效应[J].华南农业大学学报, 1998, 19(3):56-60. ZHOU Zhihong, LUO Shiming, MOU Ziping. Study on allelopathic potentials of several chemical compounds of tomato (Lycopersicon)[J]. J South China Agric Univ, 1998, 19(3):56-60. [9] LATTO J, WRIGHT H. Allelopathy in seeds[J]. J Biol Educ, 1995, 29(2):123-128. [10] LI Pei, CHANG Qing, WANG Chen, et al. Composting of aerial parts of crofton weed (Eupatorium adenophorum Spreng), the top invasive plant in southwest China[J]. Compos Sci Util, 2014, 22(3):132-137. [11] WAKJIRA M, BERECHA G, TULU S. Allelopathic effects of an invasive alien weed Parthenium hysterophorus L. compost on lettuce germination and growth[J]. Afr J Agric Res, 2009, 4(11):1325-1330. [12] BONANOMI G, DEL SORBO G, MAZZOLENI S, et al. Autotoxicity of decaying tomato residues affects susceptibility of tomato to Fusarium wilt[J]. J Plant Pathol, 2007, 89(2):219-226. [13] 耿凤展. 番茄秸秆高温堆肥基质对番茄生长影响的研究[D]. 杨凌: 西北农林科技大学, 2015. GENG Fengzhan. Study on the Effects of the Aerobic Compost of Tomato Residues on the Growth of Tomatoes[D]. Yangling:Northwest Agriculture and Forestry University, 2015. [14] WILLIAMSON G B, RICHARDSON D. Bioassays for allelopathy:measuring treatment responses with independent controls[J]. J Chem Ecol, 1988, 14(1):181-187. [15] 吴会芹, 董林林, 王倩.玉米、小麦秸秆水浸提液对蔬菜种子的化感作用[J].华北农学报, 2009, 24(增刊2):140-143. WU Huiqin, DONG Linlin, WANG Qian. Allelopathy of corn and wheat straw aqueous extracts on vegetable seeds[J]. Acta Agric Boreal-Sin, 2009, 24(supp 2):140-143. [16] 张国伟, 刘瑞显, 杨长琴, 等.棉花秸秆浸提液对小麦种子萌发及幼苗生长的化感效应[J].麦类作物学报, 2015, 35(4):555-562. ZHANG Guowei, LIU Ruixian, YANG Changqin, et al. Allelopathic effects of cotton straw extract on seed germination and seeding growth of wheat[J]. J Triticeae Crop, 2015, 35(4):555-562. [17] 魏玲, 程智慧, 张亮.不同品种大蒜秸秆水浸液对番茄的化感效应[J].西北农林科技大学学报 (自然科学版), 2008, 36(10):139-145. WEI Ling, CHENG Zhihui, ZHANG Liang. Allelopathy of straw aqueous extracts of different garlic varieties on tomato (Lycopersicon esculentum)[J]. J Northwest A & F Univ Nat Sci Ed, 2008, 36(10):139-145. [18] 莫云容, 赵凯, 邓明华.番茄植株水浸提液对生菜和大白菜化感作用的研究[J].湖南生态科学学报, 2015, 2(1):1-5. MO Yunrong, ZHAO Kai, DENG Minghua. Allelopathy of aqueous extract from tomato plants on lettuce and celegy cabbage[J]. J Hunan Ecol Sci, 2015, 2(1):1-5. [19] 邓天福, 王建华, 高扬帆, 等.番茄化感物质对几种蔬菜幼苗生长的影响[J].贵州农业科学, 2010, 38(8):43-44. DENG Tianfu, WANG Jianhua, GAO Yangfan, et al. Effects of allelochemicals of tomato on seeding growth of some vegetables[J]. Guizhou Agric Sci, 2010, 38(8):43-44. [20] LOCKERMAN R H, PUTNAM A R. Growth inhibitors in cucumber plants and seeds (Autotoxicity, allelopathy)[J]. J Am Soc Hortic Sci, 1981, 106:418-422. [21] 张军, 雷梅, 高定, 等.堆肥调理剂研究进展[J].生态环境, 2007, 16(1):239-247. ZHANG Jun, LEI Mei, GAO Ding, et al. Application of amendments in composting:a review[J]. Ecol Environ, 2007, 16(1):239-247. [22] KULCU R, YALDIZ O. Composting of goat manure and wheat straw using pine cones as a bulking agent[J]. Bioresour Technol, 2007, 98(14):2700-2704. [23] RICE E L. Allelopathy[M]. 2nd. New York:Academic Press Inc, 1984:309-315. [24] 李寿田, 周健民, 王火焰, 等.植物化感作用机理的研究进展[J].农村生态环境, 2001, 17(4):52-55. LI Shoutian, ZHOU Jianmin, WANG Huoyan, et al. Allelopathic mechanism of plants[J]. Rural Eco-Environ, 2001, 17(4):52-55. [25] 陈立新, 李少博, 乔璐, 等.凋落物叶和土壤浸提液对红松种子萌发及幼苗生长的影响[J].南京林业大学学报 (自然科学版), 2016, 40(2):81-87. CHEN Lixin, LI Shaobo, QIAO Lu, et al. Influence of leaf litter and soil leaching liquor on seed germination and seeding growth of Pinus koraiensis[J]. J Nanjing For Univ Nat Sci Ed, 2016, 40(2):81-87. [26] 刘彦荣. 加工番茄化感作用机理的研究[D]. 石河子: 石河子大学, 2014: 6-8. LIU Yanrong. The Study on Allelopathic Mechanism of Processing Tomato[D]. Shihezi:Shihezi University, 2014:6-8. [27] 谢星光, 陈晏, 卜元卿, 等.酚酸类物质的化感作用研究进展[J].生态学报, 2014, 34(22):6417-6428. XIE Xingguang, CHEN Yan, BU Yuanqing, et al. A review of allelopathic researches on phenolic acids[J]. Acta Ecol Sin, 2014, 34(22):6417-6428. [28] RAJBANSHI S S, INUBUSHI K. Chemical and biochemical changes during laboratory-scale composting of allelopathic plant leaves (Eupatorium adenophorum and Lantana camara)[J]. Biol Fert Soil, 1997, 26(1):66-71. -

-
链接本文:
https://zlxb.zafu.edu.cn/article/doi/10.11833/j.issn.2095-0756.2017.02.011
 点击查看大图
点击查看大图

计量
- 文章访问数: 3313
- HTML全文浏览量: 732
- PDF下载量: 418
- 被引次数: 0
