





-
杉木Cunninghamia lanceolata是中国南方特有的速生用材树种,生长快,材质好,用途广,深受群众喜爱,是重要的商品材树种[1]。第7次全国森林资源清查结果显示,中国杉木人工林面积有853.86万hm2,占全国人工林面积的21.35%,蓄积为6.2亿m3,木材产量约占商品材的31.64%[2]。中国木材多以中小径材为主,大径材资源很少,木材供需的结构性矛盾十分突出[3]。近年来,中国各地的实践表明,人工林近自然经营能够实现长期稳定的林木生长和林分发育,是人工培育大径材的有效途径之一[4-5]。培育杉木大径材已为社会所急需,而培育大径材的关键措施在于密度控制[6]。间伐作为一种有效改善林分密度的经营手段,对于提升森林产量与品质有着重要意义。合理间伐能够有效促进林木生长,最佳间伐强度的选择一直是森林经营措施中的关键环节。当前,国内有关间伐对杉木林分和单株的影响已开展了一系列研究,如张水松等[7]研究得出林分间伐可以有效促进杉木个体生长,但不能增加其林分蓄积;孙洪刚等[8]亦得出相似结论,认为间伐不能增加杉木林分断面积。然而上述研究多侧重于杉木间伐后的效果,而有关间伐后杉木生长的动态研究尚少见报道,仅李婷婷等[9]比较了2种强度(47%和61%)间伐后4 a内杉木林分蓄积生长量、单木生长量等指标的动态变化。目前,广西杉木的栽培面积已逾100万hm2,总蓄积量约1.2亿m3,占广西林木蓄积量的20%[10]。本研究以桂西南14年生杉木人工林为对象开展近自然化改造的间伐试验,定期进行生长观测,揭示间伐后杉木林分的生长动态,以期选择合理的间伐强度,为完善杉木人工林近自然经营和大径材培育技术提供科学依据。
-
研究地位于广西凭祥市热林中心伏波实验场(22°03′N, 106°51′E),海拔400 m,属于南亚热带季风气候区,干湿季节明显,年均气温为20.5 ℃,年均降水量为1 500.0 mm,年蒸发量为1 388.0 mm。土壤为花岗岩发育而成的砖红壤。
1993年春季于热林中心伏波实验场采用广西融水种源的1年生裸根苗营造杉木人工纯林,初植密度为2 500株·hm-2,分别于1999年和2003年进行了强度约为20%的透光伐和强度约为30%的抚育性间伐。
-
2007年10月,选择立地条件和杉木生长情况基本一致的地段开展间伐试验。试验采用随机区组设计,设置4个间伐强度(表 1)和1个对照(ck),4次重复,共20个小区,小区面积为1 500 m2。间伐后在杉木林下采用1年生苗均匀套种大叶栎Castanopsis fissa,红椎C. hystrix,格木Erythrophleum fordii,灰木莲Manglietia glauca,铁力木Mesua ferrea,枫香Liquidambar formosana和香梓楠Michelia hedyosperma等阔叶树种,套种密度为450~525株·hm-2。对照为不间伐、不套种。间伐前,根据单木生长竞争特征将林木分为目标树、特殊目标树、干扰树和一般木,分别进行标记、编号。选择目标树做永久性标记,按照试验设计要求,伐除全部干扰树及部分一般木[4]。此工作在2007年底之前全部完成。
表 1 杉木人工林间伐强度的设置
Table 1. Thinning intensity treatment of Chinese fir plantation
处理 保留株数/(株·hm-2) 间伐强度/% Ⅰ 375 69 Ⅱ 488 60 Ⅲ 594 51 Ⅳ 732 40 ck 1 219 0 -
间伐作业前,于杉木纯林内设置4块400 m2的圆形样地进行本底调查,调查样地内所有杉木的胸径、树高、枝下高和冠幅,包括应用围尺进行每木检尺,用皮尺分东、西、南和北4个方向测定冠幅,采用VERTEX超声波测高器测量树高和枝下高。林分平均保留密度为1 219株·hm-2,平均胸径13.71 cm,平均树高11.43 m,平均蓄积量98.68 m3·hm-2。间伐后于各个小区内布设面积为400 m2的圆形固定样地,分别于2008,2010,2012,2014年底和2016年8月进行定株生长观测。
-
冠幅为东西、南北2个方向的平均值。单株材积的计算采用V=gf(h+3),其中:h为全树高(m),g为胸高断面积(m2),f为平均实验形数,杉木取0.429[11]。采用单因素方差分析和Duncan多重比较检验各间伐强度间杉木林分生长表现及其年均增量的差异。应用SPSS 16.0软件进行数据统计分析。由于套种的植株尚未进入主林层,对杉木生长的影响不大,故本研究在分析生长动态时未予考虑。
-
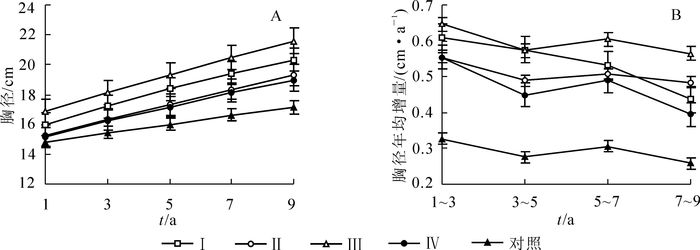
由图 1A可以看出:各处理的胸径生长总体上随着时间的推移而呈直线升高的趋势。间伐后第1~7年,处理Ⅰ和处理Ⅲ胸径差异不显著,而在第7~9年,处理Ⅲ显著高于处理Ⅰ(P<0.05);间伐后第1~9年,处理Ⅰ和处理Ⅲ的胸径显著高于处理Ⅱ和处理Ⅳ(P<0.05),后两者在各时间段的胸径几乎一致。4个间伐处理的胸径在各时间段均显著高于对照(P<0.05),以处理Ⅲ为最高,间伐后第1年,较对照高出13.5%,第9年较对照高26.3%。间伐后第9年,中、大径级比例随着间伐强度的增大呈现出先升高后下降的趋势(表 2)。4种间伐处理未见径级10 cm(含10) 以下的林木,而对照有8.72%;比较12~20 cm径级林木的分布频率,对照和处理Ⅳ较为接近,分别为70.26%和74.36%,其余3个处理为48.42%~65.38%;径阶22 cm以上林木的分布频率,对照和处理Ⅳ亦较为接近,为21.03%和25.64%,而其余处理为34.62%~51.58%。中、大径级林木的比例均以处理Ⅲ为最高。

图 1 不同强度间伐后杉木人工林胸径生长动态
Figure 1. Growth dynamics of diameter at breast height in Chinese fir plantations after thinning with different intensities
表 2 间伐后第9年杉木人工林的径级分布
Table 2. Distribution of diameter grade in Chinese fir plantations 9 years after thinning
径级/cm 不同处理的林木径级分布频率/% Ⅰ Ⅱ Ⅲ Ⅳ ck 8 0.00 0.00 0.00 0.00 1.54 10 0.00 0.00 0.00 0.00 7.18 12 1.67 2.56 1.05 3.42 10.26 14 6.67 5.13 6.32 9.40 15.90 16 10.00 21.79 13.68 18.80 23.08 18 16.67 12.82 12.63 26.50 10.26 20 25.00 23.08 14.74 16.24 10.77 22 20.00 19.23 16.84 9.40 7.69 24 5.00 11.54 13.68 6.84 5.64 26 6.67 1.28 10.53 5.13 3.59 28 5.00 2.56 6.32 2.56 2.05 30 3.33 0.00 3.16 0.85 2.05 32 0.00 0.00 1.05 0.85 0.00 各处理的胸径年均增量随着时间的推移均呈逐渐降低的趋势(图 1B)。间伐后第1~3年,4个间伐处理间胸径年均增量无明显差异,但均显著高于对照(P<0.01);间伐后第3~9年,处理Ⅲ的胸径年均增量显著高于处理Ⅳ(P<0.05),与处理Ⅰ和处理Ⅱ的处理间差异不显著,4个间伐处理均显著高于对照(P<0.05)。在间伐后的9 a间,处理Ⅲ的胸径年均增量比对照增加了约1.0倍。
-
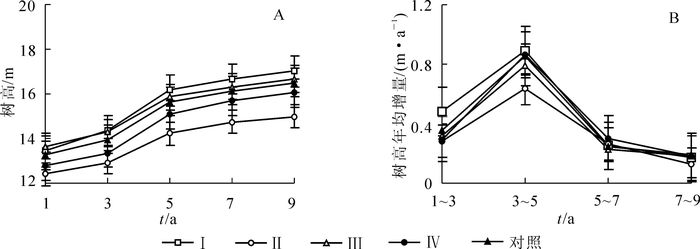
从整体上看,各处理间树高生长较为平稳,而间伐后第3~5年是树高的快速增长期(图 2A和2B)。间伐后第1~9年间,处理Ⅰ和处理Ⅲ和对照的树高显著高于处理Ⅱ和处理Ⅳ(P<0.05)。间伐后第1年,树高以处理Ⅲ为最大,处理Ⅱ为最小,前者较后者高10%;第9年则以处理Ⅰ为最大,较处理Ⅱ高14%。间伐后第1~3年,处理Ⅰ的树高年均增量显著高于处理Ⅱ,处理Ⅲ,处理Ⅳ和对照(P<0.05);第3~9年,5个处理间的树高年均增量无明显差异。

图 2 不同强度间伐后杉木人工林树高生长动态
Figure 2. Height growth dynamics of chinese fir plantations after thinning with different intensities
-
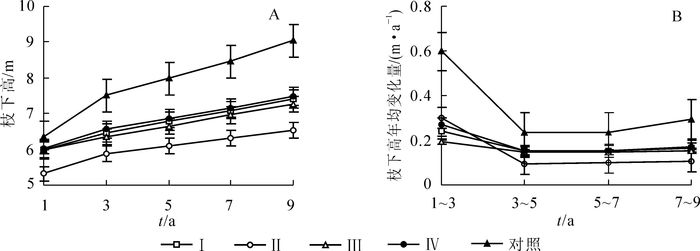
各间伐处理的枝下高变化亦较为平稳,均显著低于对照(图 3A)。处理Ⅰ,处理Ⅲ和处理Ⅳ的枝下高在间伐后各时间段内均非常接近,而处理Ⅱ略微低于此3个处理。对照的枝下高在间伐后第1年高出4个间伐处理0.3~1.0 m;随着时间的推移,差距在不断的拉大,到间伐后第9年,对照的枝下高比处理Ⅱ高1.5 m。

图 3 不同强度间伐后杉木人工林枝下高动态变化
Figure 3. Dynamic changes of crown base height in Chinese fir plantations after thinning with different intensities
由图 3B可知:各间伐处理和对照的枝下高年均增量随着时间的推移先下降而后趋于平稳。对照的枝下高年均增量在间伐后各时间段均显著高于4个间伐处理(P<0.01)。间伐后第1~3年,各处理的枝下高增量出现峰值,以处理Ⅲ为最小,仅为对照的1/3。
-
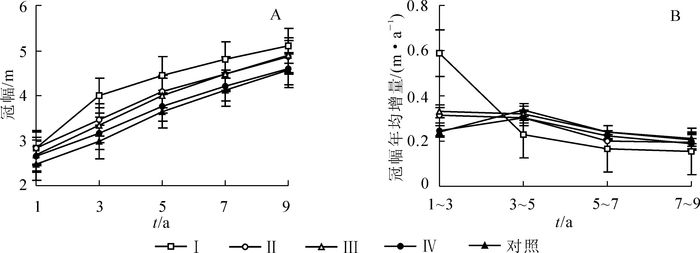
间伐后第1~9年,随着杉木林分保留密度增大,冠幅生长减缓(图 4A)。间伐后第1年、第3年、第5年和第7年,处理Ⅰ、处理Ⅱ和处理Ⅲ的冠幅显著大于处理Ⅳ,而4个间伐处理均显著大于对照(P<0.05)。而间伐后第9年,4个间伐处理的冠幅差异不显著,但仍显著大于对照(P<0.05),以处理Ⅱ为最大,是对照的1.4倍。

图 4 不同强度间伐后杉木人工林冠幅生长动态
Figure 4. Growtjh dynamics of crown width in Chinese fir plantations after thinning with different intensities
从图 4B中可以看出:处理Ⅰ在间伐后第1~3年出现冠幅生长高峰,随后其增量逐年减小。处理Ⅱ,处理Ⅲ,处理Ⅳ和对照的冠幅年均增量呈平稳或下降趋势,无显著差异。除了处理Ⅰ的冠幅年均增量在间伐后第1~3年显著高于其他各处理外(P<0.01),其余各时间段内,各间伐处理的冠幅年均增量与对照差异不显著。在间伐后第1~3年,处理Ⅰ的冠幅年均增量为对照2.4倍。
-
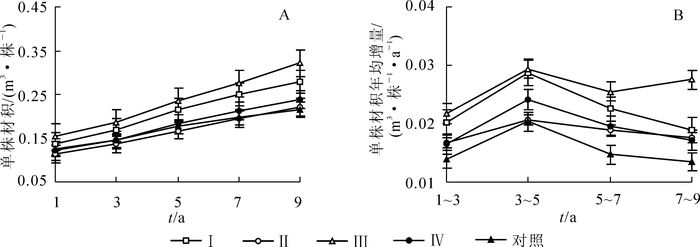
结果显示(图 5A):间伐处理Ⅰ和处理Ⅲ的单株材积在间伐后第1~9年均显著大于处理Ⅱ和处理Ⅳ和对照(P<0.05)。间伐后,处理Ⅲ的单株材积为最高,对照处理的单株材积为最低,随着时间的推移,两者间的差距在逐渐加大。到间伐后第9年,处理Ⅲ的单株材积为对照的1.5倍。

图 5 不同强度间伐后杉木人工林单株材积生长动态
Figure 5. Growth dynamics of individual tree volume in Chinese fir plantations after thinning with different intensities
各处理的单株材积年均增量随时间的推移呈现先增大而后减小或稳定的趋势(图 5B)。间伐后第1~7年,处理Ⅰ和处理Ⅲ的单株材积年均增量显著高于处理Ⅱ,处理Ⅳ以及对照(P<0.05);而在间伐后第7~9年,处理Ⅲ的单株材积年均增量显著高于其他3个间伐处理(P<0.01)。
-
由图 6A可以看出:处理Ⅲ和处理Ⅳ,处理Ⅰ和处理Ⅱ在间伐后各时间段的蓄积量均几乎相等;4个间伐处理的蓄积显著低于对照(P<0.01),处理Ⅲ和处理Ⅳ的蓄积显著高于处理Ⅰ和处理Ⅱ(P<0.05)。在间伐后第5年、第7年、第9年,对照处理的蓄积量与处理Ⅲ和处理Ⅳ的差距在逐渐缩小,间伐后第1年对照的蓄积是处理Ⅲ的1.7倍,第9年为处理Ⅲ的1.5倍。

图 6 不同强度间伐后杉木人工林林分蓄积生长动态
Figure 6. Growth dynamics of stand volume in Chinese fir plantations after thinning with different intensities
各间伐处理的林分蓄积年均增量变化平稳,间伐后第3~5年略微增大;而对照的林分蓄积年均增量在间伐后第1~3年缓慢增长,第3~5年陡然增大,在间伐后第5~7年和第7~9年缓慢降低(图 6B)。间伐后第1~9年,处理Ⅰ和处理Ⅱ的林分蓄积年均增量显著小于对照(P<0.01),而处理Ⅲ和处理Ⅳ与对照无显著差异。间伐后第7~9年,蓄积年均增量最大的间伐处理Ⅲ为对照的83.1%。
-
选择最佳间伐强度调整林分结构,对于林木生长极为重要[12-13]。本研究中14年生杉木的胸径和冠幅在间伐后呈现快速生长,不论是从林分的胸径均值还是年均增长量来看,4个间伐处理均显著高于对照,且间伐强度越大,其胸径、冠幅生长越快。间伐对杉木生长的促进作用无明显滞后现象,间伐后第1~3年即是其胸径和冠幅的快速增长期,随后6 a(至最后1次观测)其胸径增加量仍显著高于对照。CRECENTE-CAMPO等[14]研究得出:强度间伐(32%~46%)和超强度间伐(51%~57%)能使50年生欧洲赤松Pinus sylvestris的胸径、冠幅增量在短期(4 a)内显著增加。MISSON等[15]对挪威云杉Picea abies间伐(强度为40%~80%)后25 a连续观测研究发现:强度间伐对林木径向生长的促进作用能够持续约4~10 a。而17年生湿地松Pinus elliottii间伐后对胸径和冠幅生长的促进作用存在2~4 a的滞后现象[16]。由此可见,间伐对林木生长促进作用的起始时间、持续时间因树种、林龄而异;杉木生长对间伐响应快速,间伐对其保留木生长的促进作用持续时间长。其可能的原因在于:其一、杉木为中性偏荫树种,对间伐后光环境变化的适应能力强,因而间伐对其生长促进作用无滞后现象;其二、杉木属窄冠型树种,其冠幅小、冠长大,而且其冠幅生长相对较慢,因而间伐对杉木生长的促进作用持续时间长。
本研究中杉木各间伐处理与对照间的树高年均增量相差不大。此与张水松等[7]对10年生杉木人工林强、中、弱度间伐后20 a的生长观测结果一致。然而,周成军等[17]对33年生杉木人工林进行了强度为12.9%~61.5%的择伐试验,发现中度(23.5%)择伐更有利于树高生长。这些研究得出不一致的结果,可能与其杉木年龄、立地条件、生长势等差异有关。不过,大多数学者认为间伐对树高生长影响较小,如,MEDHURST等[18]对不同年龄亮果桉Eucalyptus nitens人工林进行强度为14%~72%的间伐研究,亦得出间伐不影响树高生长的结论。4个间伐处理的杉木枝下高年均增量显著低于对照,而4个间伐处理间差异不显著。究其原因,可从自然整枝与林分竞争方面得以解释:枝下高的变化直接取决于自然整枝强度,而自然整枝强度与林分竞争有关;对照林分密度大,生长空间竞争激烈,会加剧其自然整枝[19-20],而4个处理的间伐强度为40%~69%,其空间竞争差异可能尚不明显。
间伐是增加林木收获量和大径材比例最常见的干预措施[21]。林分单株材积及其年均增长量方面,处理Ⅲ均显著高于对照林分。从林分蓄积来看,处理Ⅰ和处理Ⅱ的蓄积及其年均增量均显著低于对照,而处理Ⅲ和处理Ⅳ的蓄积年均增量与对照无显著差异。随着时间的推移,4个间伐处理的蓄积年均增量与对照的差距在缩小。由此可见,间伐后林木单株材积的增量可弥补因保留木数量减少而造成的林分蓄积增量的降低。国内一些学者研究亦表明,间伐有利于杉木大径级林木的生长,虽然间伐使林木株数减少,但到主伐年龄时其蓄积增量与不间伐相差不大[22-23]。从间伐后第9年林木径级分布还可以看出,间伐处理的中、大径级林木的比例远高于对照,尤其以处理Ⅲ的大径级林木比例最高,有利于大径材生产,其经济效益亦会得到显著提升。
杉木林分的胸径、冠幅和单株材积的增量随间伐强度的增大而增大,而枝下高和林分蓄积则呈相反的变化趋势。杉木人工林近自然化改造不同于传统的人工林经营,其间伐强度高,旨在快速培育大、中径材,且在其间伐后套种珍贵乡土阔叶树种,营造针阔异龄混交林。根据不同间伐处理下杉木单株和林分生长表现,建议选择50%的间伐强度为宜。本研究是在未考虑套种阔叶树影响的情况下进行的,在今后的长期试验中,还应考虑到林下阔叶树逐步进入到主林层后林分的空间结构与环境变化、林木枯损、种间关系等因素对杉木生长的影响,此亦为今后研究的重点。
-
承蒙中国林业科学研究院热带林业研究所曾杰研究员审阅修改论文。特致谢忱!
Growth response of a Cunninghamia lanceolata plantation to thinning in southwestern Guangxi, China
-
摘要: 以广西凭祥14年生杉木Cunninghamia lanceolata人工林(保存密度为1 219株·hm-2)为对象,设置69%(处理Ⅰ),60%(处理Ⅱ),51%(处理Ⅲ),40%(处理Ⅳ)4个间伐强度处理和不间伐对照,定期观测间伐后9 a内胸径、树高、枝下高和冠幅等林分生长指标,并计算单株材积和林分蓄积,从而揭示林分生长对间伐的动态响应,筛选适宜的间伐强度,为杉木人工林近自然化改造提供技术支撑。结果表明:间伐强度显著影响杉木林分胸径、枝下高、单株材积和蓄积生长(P < 0.05),对树高和冠幅的影响不显著(P>0.05);胸径和单株材积以处理Ⅲ为最高,枝下高和蓄积则以对照为最高。胸径、冠幅和枝下高年均增量在间伐后第1~3年最大,处理间的差异随着时间的推移逐渐缩小。树高、单株材积和林分蓄积年均增量高峰则出现在间伐后第3~5年。根据上述试验结果,为了快速培育杉木大径材,建议在杉木近自然化改造中选择间伐强度约50%(处理Ⅲ)为宜。Abstract: To reveal the dynamic response of stand growth to thinning treatments and to select suitable thinning intensities in Cunninghamia lanceolata (Chinese fir) plantations, a thinning trial was conducted in a 14-year-old plantation of C. lanceolata at Pingxiang City, Guangxi. Stand density was about 1 219 trees·hm-2, and the experiment was arranged in a randomized complete block design with five treatments: four thinning treatments of 69% (Ⅰ), 60% (Ⅱ), 51% (Ⅲ), and 40% (Ⅳ) and a control without thinning and four replicates for each treatment. Diameter at breast height (DBH), tree height, crown base height (CBH), crown width (CW), individual tree volume (ITV), and stand volume (SV) were determined with a circle samping plot of 400 m2 for each replicateperiodically in 9 years after thinning. Results from one-way analysis of variance and Duncan's multiple range tests showed that thinning intensity significantly increased DBH and ITV, decreased CBH and SV (P < 0.05). The highest DBH and ITV were observed in Treatment Ⅲ; whereas, the highest CBH and SV were in the control. The mean annual increments for DBH, CW, and CBH were highest 1 to 3 years after thinning with differences between treatments gradually declining as forest age increased; however, peaks for tree height, ITV, and SV growth occurred 3 to 5 years after thinning. Based on the above findings, a thinning intensity of about 50% (Treatment Ⅲ) should be applied for close-to-natural forest management so as to producelarge-sized timber of C. lanceolata more rapidly.
-
图 1 不同强度间伐后杉木人工林胸径生长动态
Figure 1 Growth dynamics of diameter at breast height in Chinese fir plantations after thinning with different intensities
图 2 不同强度间伐后杉木人工林树高生长动态
Figure 2 Height growth dynamics of chinese fir plantations after thinning with different intensities
图 3 不同强度间伐后杉木人工林枝下高动态变化
Figure 3 Dynamic changes of crown base height in Chinese fir plantations after thinning with different intensities
图 4 不同强度间伐后杉木人工林冠幅生长动态
Figure 4 Growtjh dynamics of crown width in Chinese fir plantations after thinning with different intensities
图 5 不同强度间伐后杉木人工林单株材积生长动态
Figure 5 Growth dynamics of individual tree volume in Chinese fir plantations after thinning with different intensities
图 6 不同强度间伐后杉木人工林林分蓄积生长动态
Figure 6 Growth dynamics of stand volume in Chinese fir plantations after thinning with different intensities
表 1 杉木人工林间伐强度的设置
Table 1. Thinning intensity treatment of Chinese fir plantation
处理 保留株数/(株·hm-2) 间伐强度/% Ⅰ 375 69 Ⅱ 488 60 Ⅲ 594 51 Ⅳ 732 40 ck 1 219 0  下载: 导出CSV
下载: 导出CSV
表 2 间伐后第9年杉木人工林的径级分布
Table 2. Distribution of diameter grade in Chinese fir plantations 9 years after thinning
径级/cm 不同处理的林木径级分布频率/% Ⅰ Ⅱ Ⅲ Ⅳ ck 8 0.00 0.00 0.00 0.00 1.54 10 0.00 0.00 0.00 0.00 7.18 12 1.67 2.56 1.05 3.42 10.26 14 6.67 5.13 6.32 9.40 15.90 16 10.00 21.79 13.68 18.80 23.08 18 16.67 12.82 12.63 26.50 10.26 20 25.00 23.08 14.74 16.24 10.77 22 20.00 19.23 16.84 9.40 7.69 24 5.00 11.54 13.68 6.84 5.64 26 6.67 1.28 10.53 5.13 3.59 28 5.00 2.56 6.32 2.56 2.05 30 3.33 0.00 3.16 0.85 2.05 32 0.00 0.00 1.05 0.85 0.00
下载: 导出CSV
-
[1] 盛炜彤, 范少辉.杉木人工林长期生产力保持机制研究[M].北京:科学出版社, 2005. [2] 国家林业局森林资源管理司.第7次全国森林资源清查及森林资源状况[J].林业资源管理, 2010(1): 1-8. Department of Forest Resources Management of State Forestry Administration. Seventh national forest inventory and forest resources [J]. For Resour Manage, 2010(1): 1-8. [3] 石春娜, 王立群.我国森林资源质量变化及现状分析[J].林业科学, 2009, 45(11): 90-97. SHI Chunna, WANG Liqun. Status quo and changes analysis of forest resources quality in China [J]. Sci Silv Sin, 2009, 45(11): 90-97. [4] 陆元昌.近自然森林经营理论与实践[M].北京:科学出版社, 2006. [5] 高云昌, 张文辉, 何景峰, 等.黄龙山油松人工林间伐效果的综合评价[J].应用生态学报, 2013, 24(5): 1313-1319. GAO Yunchang, ZHANG Wenhui, HE Jingfeng, et al. Effects of thinning intensity on Pinus tabulaeformis plantation in Huanglong Mountain, Northwest China: a comprehensive evaluation [J]. J Appl Ecol, 2013, 24(5): 1313-1319. [6] 何齐发, 杨馥宁, 郑小贤.杉木大径材作业级水平经营模式研究[J].林业资源管理, 2008(6): 55-58. HE Qifa, YANG Funing, ZHENG Xiaoxian. Study on sustainable management pattern of Chinese fir for big-diameter timber [J]. For Resour Manage, 2008(6): 55-58. [7] 张水松, 陈长发, 吴克选, 等.杉木林间伐强度试验20年生长效应的研究[J].林业科学, 2005, 41(5): 56-65. ZHANG Shuisong, CHEN Changfa, WU Kexuan, et al. Growth effect of intermediate cuttings intensity experiment for twenty years in Cunninghamia lanceolata [J]. Sci Silv Sin, 2005, 41(5): 56-65. [8] 孙洪刚, 张建国, 段爱国, 等.杉木密度间伐试验林林分断面积生长效应[J].林业科学研究, 2010, 23(1): 6-12. SUN Honggang, ZHANG Jianguo, DUAN Aiguo, et al. Long-term effects of thinning on stand basal area increment of Cunninghamia lanceolata [J]. For Res, 2010, 23(1): 6-12. [9] 李婷婷, 陆元昌, 庞丽峰, 等.杉木人工林近自然经营的初步效果[J].林业科学, 2014, 50(5): 90-100. LI Tingting, LU Yuanchang, PANG Lifeng, et al. Initial effect of close-to-nature management of Chinese fir plantation[J]. Sci Silv Sin, 2014, 50(5): 90-100. [10] 陈代喜, 陈琴, 蒙跃环, 等.杉木大径材高效培育技术探讨[J].南方农业学报, 2015, 46(2): 293-298. CHEN Daixi, CHEN Qin, MENG Yuehuan, et al. High-efficient management technology of big-diameter timber of Cunninghamia laceolata [J]. J Southern Agric, 2015, 46(2): 293-298. [11] 孟宪宇.测树学[M]. 2版.北京:中国林业出版社, 1996. [12] 李春明, 杜纪山, 张会儒.抚育间伐对森林生长的影响及其模型研究[J].林业科学研究, 2003, 16(5): 636-641. LI Chunming, DU Jishan, ZHANG Huiru. The effects of thinning on forest growth and model study [J]. For Res, 2003, 16(5): 636-641. [13] WARD J S. Intensity of precommercial crop tree release increases diameter growth and survival of upland oaks [J].Can J For Res, 2009, 39(1): 118-130. [14] CRECENTE-CAMPO F, POMMERENING A, RODRÍGUEZ-SOALLEIRO R. Impacts of thinning on structure, growth and risk of crown fire in a Pinus sylvestris, L. plantation in northern Spain [J]. For Ecol Manage, 2009, 257(9): 1945-1954. [15] MISSON L, VINCKE C, DEVILLEZ F. Frequency responses of radial growth series after different thinning intensities in Norway spruce (Picea abies (L.) Karst.) stands [J]. For Ecol Manage, 2003, 177(1/3): 51-63. [16] SCHEXNAYDER B, CAMILLE J. Growth of a Slash Pine Spacing Study Five Years after Thinning [D]. Baton Rouge: Louisiana State University, 2005. [17] 周成军, 巫志龙, 周新年, 等.山地杉木人工林不同强度择伐后生长动态仿真[J].山地学报, 2012, 30(6): 669-674. ZHOU Chengjun, WU Zhilong, ZHOU Xinnian, et al. The stand growth dynamics simulation of the mountain Chinese fir plantation under different selective cutting intensity [J]. J Mount Sci, 2012, 30(6): 669-674. [18] MEDHURST J L, BEADLE C L, NEILSEN W A. Early-age and later-age thinning affects growth, dominance, and intras [J]. Can J For Res, 2001, 31(2):187-197. [19] 张宁, 张怀清, 林辉, 等.基于竞争指数的杉木林分生长可视化模拟研究[J].林业科学研究, 2013, 26(6): 692-697. ZHANG Ning, ZHANG Huaiqing, LIN Hui, et al. Visual simulation of growth process in Cunninghamia lanceolata based on competition index [J]. For Res, 2013, 26(6): 692-697. [20] FU Liyong, SUN Hua, SHARMA R P, et al. Nonlinear mixed-effects crown width models for individual trees of Chinese fir (Cunninghamia lanceolata) in south-central China [J]. For Ecol Manage, 2013, 302(6): 210-220. [21] HUONG V D, MENDHAM D S, CLOSE D C. Growth and physiological responses to intensity and timing of thinning in short rotation tropical Acacia hybrid plantations in South Vietnam [J]. For Ecol Manage, 2016, 380(8): 232-241. [22] 吴建强, 王懿祥, 杨一, 等.干扰树间伐对杉木人工林林分生长和林分结构的影响[J].应用生态学报, 2015, 26(2): 340-348. WU Jianqiang, WANG Yixiang, YANG Yi, et al. Effects of crop tree release on stand growth and stand structure of Cunninghamia lanceolata plantation [J]. Chin J Appl Ecol, 2015, 26(2): 340-348. [23] 叶功富, 涂育合, 林瑞荣, 等.杉木人工林不同密度管理定向培育大径材[J].北华大学学报(自然科学版), 2005, 6(6): 544-549. YE Gongfu, TU Yuhe, LIN Ruirong, et al. On big-diameter-oriented cultivation techniques of Cunninghamia lanceolata of different density measures [J]. J Beihua Univ Nat Sci, 2005, 6(6): 544-549. -

-
链接本文:
https://zlxb.zafu.edu.cn/article/doi/10.11833/j.issn.2095-0756.2017.05.010
 点击查看大图
点击查看大图
计量
- 文章访问数: 3483
- HTML全文浏览量: 641
- PDF下载量: 313
- 被引次数: 0
