





-
生物多样性是衡量某一地区生态是否可持续发展的重要指标[1],而昆虫多样性研究在生物多样性的研究中有着举足轻重的作用[2]。昆虫体积小,数量多,繁殖力强,所需生存空间占比小,并且在不同环境中都能找到。昆虫对生存坏境变化敏感的特性也为研究昆虫多样性与环境变化的关系提供了前提,因此,开展昆虫多样性的研究意义重大[3]。近年来,对于昆虫多样性的研究越来越广泛,从农业到森林生态系统,昆虫多样性的研究都取得了重要进展。李浩[4]发现有机果园的昆虫多样性更高,生态系统更稳定。马玲等[5]在对小兴安岭不同林型的昆虫多样性分析中发现不同林型存在昆虫多样性差异。山核桃Carya cathayensis主要分布在以浙江省杭州市临安区为中心的天目山山区[6]。山核桃因具有较高的营养价值颇受人们的喜爱,已成为该地区林农的主要收入来源。随着产业发展,传统经营方式的山核桃林地生态问题日渐严重[7],经营模式的改变迫在眉睫。为此,专家已提出套种作物、设置隔离带、改变施肥方式等多种更具生态化的经营模式[6]。本研究选取临安地区有明显差异的4种经营方式,调查其昆虫多样性的组成和差异,探讨不同经营模式对昆虫多样性的影响,旨在通过昆虫多样性的分析对山核桃的经营模式提出建议。
-
浙江省杭州市临安区位于杭州市西部,浙江省西北部,29°56′~30°23′N,118°51′~119°52′E。现有山核桃林总面积为2.87万hm2,约占全国现有面积的40%[6]。区内山核桃主要分布于岛石、龙岗、昌化、湍口等乡镇[8]。表 1为本研究选取的4种不同经营模式山核桃林分布及概况。选取的山核桃林树龄均为20~30 a,造林密度为300~525株·hm-2。
表 1 不同经营模式山核桃林分布概况
Table 1. Distribution of different management modes of Carya cathayensis forests
模式代号 经营模式 地点 经纬度 海拔/m 生境 Ⅰ 野生的山核桃林 板桥镇孟坞里 30°11′8.29″N, 119°44′57.46″E 100 多年无人管理,呈自由生长状态,林下植被复杂,以灌木、草本植物为主。无施药和施肥。林下覆盖度大于95% Ⅱ 林下种植植物的山核桃林 湍口镇湍口村 30°03′23.88″N, 119°08′29.80″E 182 林下种植多种绿肥作物包括黑麦草、紫花苜蓿、白三叶、菊科植物等;灌木有南天竹;乔木有香榧、黄枫、桂花、银杏、樟树等。以生态调控治理为主,其他治理手段为辅。林下覆盖度大于85% Ⅲ 传统经营的山核桃林 河桥镇寺坞里村 30°04′14.99″N, 119°10′50.02″E 108 施除草剂和杀虫剂,林下植被少且单一,主要为草本植物。林下覆盖度小于10% Ⅳ 参照有机化管理的山核桃林 河桥镇寺坞里村 30°04′12.40″N, 119°10′51.12″E 108 割草机除草,施放有机肥。林下植被少,林间套种经济作物,主要为黄精、香榧苗。林下覆盖度大于65% 说明:黑麦草Lolium multiflorum,紫花苜蓿Medicago sativa,白三叶Trifolium repens,南天竹Nandina domestica,香榧Torreya grandis ‘Merrillii’,黄枫Tri kvartala,桂花Osmanthus fragrans,银杏Ginkgo biloba,樟树Cinnamomum camphora,黄精Polygonatum sibiricum -
标本收集利用马氏网、灯诱和样方调查的方法。①马氏网收集:在4个研究地各随机设置5个马氏网,每个马氏网互相间隔50 m,于2016年5-10月,隔30 d收集并替换收集瓶;②灯诱:诱集鳞翅目昆虫,在4个研究地的中心区域各选取1处空旷地,于2017年7月、8月各进行1次灯诱,每次诱集1晚,时间为19:00-21:00;③样方调查:调查山核桃树表面和钻蛀性的昆虫,在4个研究地各随机选取5株较矮的山核桃树,每株树的相互间隔50 m,于2017年7月、8月各进行1次调查,采集并记录树上的昆虫。最后将收集到的昆虫标本带回实验室鉴定并计数。
-
数据利用Excel 2013和SPSS 19.0软件计算处理。群落特征指数:物种丰富度S用群落中的物种种数表示;多样性指数H′用Shannon-Wiener多样性指数表示,公式为$H^{\prime}=-\sum\limits_{i}^{S} P_{i} \ln P_{i}$;均匀度E用基于Shannon-Wiener多样性指数与理论最大多样性指数的比值来表示,公式为$E=\left(-\sum\limits_{i}^{S} P_{i} \ln P_{i}\right) / \ln S$;个体数N用全部物种的个体数之和表示。优势度指数C用Simpson优势度指数表示,公式为$C=\sum\limits_{i}^{s} P_{i}^{2}$。其中:Pi为第i个物种数量占所有物种总数的比例[15-19]。不同经营模式间采用单因素分析(one-way ANOVA)和Duncan多重比较方法行进多样性差异分析[20]。群落相对稳定性分析用天敌类群物种种数和害虫类物种种数之比(Sn/Sp)及群落物种种数和个体数之比(St/Si)表示,其中Sn/Sp表示食物链的复杂度和相互制约度,而St/Si反映种间数量的制约作用[21]。功能集团分析参考文献[22-24]将所采集的昆虫分为以下5种不同功能集团:捕食性、食叶性、钻蛀性、寄生性和中性昆虫。将归类好的功能集团进行主分量分析和相似性分析。
-
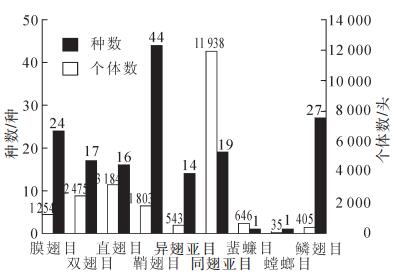
本研究共获得标本22 283号,共计9目84科163种。由图 1可知:从物种数看,鞘翅目Coleoptera昆虫44种,鳞翅目Lepidoptera昆虫27种,膜翅目Hymenoptera昆虫24种,分别占总数的26.99%,16.56%,14.72%,是3类最多的昆虫。从个体数看,同翅亚目Homoptera,直翅目Orthoptera,双翅目Orthoptera数量最多,分别占总个体数的53.54%,14.28%,11.10%。

图 1 山核桃林昆虫群落组成
Figure 1. Insect community composition of Carya cathayensis forests
由表 2可知:4种经营模下的山核桃林中,均为膜翅目、双翅目、鞘翅目、同翅亚目、鳞翅目种数较多。其中,模式Ⅰ和模式Ⅱ种数从大到小的目依次为鞘翅目、鳞翅目、膜翅目、同翅亚目、双翅目;模式Ⅲ和模式Ⅳ种类数从大到小的目依次为鞘翅目、鳞翅目、膜翅目、双翅目、同翅亚目。4种经营模式中均为模式Ⅱ昆虫种数和个体数最多,模式Ⅲ昆虫种数和个体数最少。在4种经营模式中,均为同翅亚目个体数第1位,直翅目第2位。其中,同翅亚目昆虫个体数以模式Ⅳ最多,占62.12%;直翅目昆虫数量以模式Ⅱ最多,占13.52%。并且,在各自经营模式中,同翅亚目昆虫都以蚜科Aphidoidea为主,直翅目昆虫都以蝗科Acrididae为主。
表 2 不同经营模式的总昆虫群落
Table 2. All insect community under different management modes
类别 Ⅰ Ⅱ Ⅲ Ⅳ 种数/种 个体数/头 相对多度/% 种数/种 个体数/头 相对多度/% 种数/种 个体数/头 相对多度/% 种数/种 个体数/头 相对多度/% 膜翅目Hymenoptera 20 343 9.31 24 487 6.94 14 167 3.34 19 257 3.91 双翅目Diptera 16 540 14.66 17 822 11.72 14 464 9.27 16 649 9.86 直翅目Orthoptera 14 689 18.70 16 948 13.52 5 831 16.60 7 716 10.88 鞘翅目Coleoptera 37 414 11.24 44 683 9.74 23 275 5.49 32 431 6.55 异翅亚目Heteroptera 11 159 4.32 14 237 3.38 6 59 1.18 10 88 1.34 同翅亚目Homoptera 17 1 346 36.54 19 3 532 50.36 10 2 972 59.38 12 4 088 62.12 蜚蠊目Blattaria 1 87 2.36 1 169 2.41 1 149 2.98 1 241 3.66 螳螂目Mantodea 1 10 0.27 1 7 0.10 1 5 0.10 1 13 0.20 鳞翅目Lepidoptera 24 96 2.61 27 128 1.83 21 83 1.66 23 98 1.49 合计 141 3 684 100.00 163 7 013 100.00 95 5 005 100.00 121 6 581 100.00 -
从表 3可知:总昆虫种数从高到低的经营模式为模式Ⅱ(163种),模式Ⅰ(141种),模式Ⅳ(121种),模式Ⅲ(95种);个体数从高到低为模式Ⅱ(7 013头),模式Ⅳ(6 581头),模式Ⅲ(5 005头),模式Ⅰ(3 684头)。模式Ⅱ在种数和个体数上均最多;模式Ⅰ在种数上较多,个体数上最少;模式Ⅲ在种数上最少。这是由于模式Ⅱ种植了许多林下植物,并且几乎不使用农药,使得该经营模式下昆虫种类和数量最多;模式Ⅰ为荒废野生状态的山核桃林,林中已基本形成生态平衡,因此该林昆虫种类多,数量少;模式Ⅲ为传统经营模式下的山核桃林,该林使用杀虫剂与除草剂,因此昆虫种类最少,个体数比模式Ⅱ和模式Ⅳ少。4种经营模式下山核桃林总昆虫群落的特征指数从高到低依次为多样性指数模式Ⅰ(3.64),模式Ⅱ(3.12),模式Ⅳ(2.38),模式Ⅲ(2.15),均匀度指数从高到低依次为模式Ⅰ(0.74),模式Ⅱ(0.61),模式Ⅳ(0.50),模式Ⅲ(0.47),优势度指数从高到低依次为模式Ⅲ(0.33),模式Ⅳ(0.31),模式Ⅱ(0.20),模式Ⅰ(0.10)。4种经营模式中,模式Ⅰ和模式Ⅱ的多样性指数较高,物种较丰富。模式Ⅲ和模式Ⅳ优势度较高,存在明显优势种。
表 3 不同经营模式的总昆虫群落特征值
Table 3. Characteristic value of all insect communities under different management modes
经营模式 个体数/头 种数/种 H′ E C Ⅰ 3 684 141 3.64 0.74 0.10 Ⅱ 7 013 163 3.12 0.61 0.20 Ⅲ 5 005 95 2.15 0.47 0.33 Ⅳ 6 581 121 2.38 0.50 0.31 -
由于灯诱与样方调查的次数和昆虫数量的太多或太少,如灯诱共进行2次,无重复组,而样方调查模式Ⅰ的同翅目蚜科数量有1 045个,这都会使显著性分析误差较大。因此,只利用马氏网法收集昆虫群落进行显著性差异分析。由表 4可知:从昆虫物种数上看,模式Ⅱ种数最多,模式Ⅲ最少,4种经营模式的物种数存在显著差异(P<0.05);从昆虫个体数上看,模式Ⅱ个数最多,并显著高于其他,模式Ⅰ和模式Ⅳ个体数差异不显著,模式Ⅰ和模式Ⅲ个体数差异不显著,但模式Ⅳ个体数显著高于模式Ⅲ(P<0.05);从昆虫多样性指数看,模式Ⅱ最高,但和模式Ⅰ差异不显著,模式Ⅰ和模式Ⅱ显著高于模式Ⅲ和模式Ⅳ,模式Ⅲ和模式Ⅳ无显著差异(P<0.05);从均匀度指数看,模式Ⅰ最高,但和模式Ⅱ无显著差异,模式Ⅰ和模式Ⅱ显著高于模式Ⅲ和模式Ⅳ,模式Ⅳ显著高于模式Ⅲ(P<0.05);从优势度指数看,模式Ⅲ最高,显著高于其他,模式Ⅳ显著高于模式Ⅰ和模式Ⅱ,模式Ⅰ和模式Ⅱ无显著差异(P<0.05)。综上可以发现,模式Ⅰ和模式Ⅱ的昆虫群落差异较小,群落稳定性也相较于模式Ⅲ和模式Ⅳ更好。模式Ⅲ的多样性低,且优势度较高,存在明显优势种,群落稳定性较差。
表 4 不同经营模式的马氏网法收集的昆虫群落特征值
Table 4. Characteristic value of insect communities in Malaise trap under different management modes
经营模式 种数/种 个体数/头 H′ E C Ⅰ 103.20 ± 5.93 b 506.80 ± 101.59 bc 4.06 ± 0.07 a 0.88 ± 0.015 a 0.038 ± 0.005 c Ⅱ 119.00 ± 3.54 a 758.40 ± 43.90 a 4.16 ± 0.10 a 0.87 ± 0.021 a 0.033 ± 0.008 c Ⅲ 60.40 ± 1.82 d 431.00 ± 31.63 c 3.01 ± 0.07 c 0.74 ± 0.019 c 0.134 ± 0.013 a Ⅳ 78.20 ± 4.60 c 577.00 ± 29.89 b 3.52 ± 0.06 b 0.81 ± 0.004 b 0.066 ± 0.005 b 说明:同列不同小写字母表示差异显著(P<0.05) -
不同经营模式下山核桃林昆虫群落的相对稳定性存在差异。Sn/Sp从高到低依次为模式Ⅳ(0.39),模式Ⅱ(0.33),模式Ⅲ(0.33),模式Ⅰ(0.31)。模式Ⅳ的比值较高,说明该林的天敌昆虫数量所占比例高,群落内部食物网的的复杂程度及相互制约能力较高;模式Ⅰ虽然比值最低,但该林较高的群落多样性指数和均匀度指数,能够明显提高该林抗干扰和抵御外来有害生物的缓冲能力。St/Si从高到低依次为模式Ⅰ(0.04),模式Ⅱ(0.02),模式Ⅲ(0.02),模式Ⅳ(0.02)。模式Ⅰ的比值较高,说明模式Ⅰ的昆虫群落结构较其他更稳定。总体而言,模式Ⅰ的稳定性较好,该种经营模式更能有利于生物多样性发展。
-
由表 5可知:4种经营模式下的山核桃林中均以植食性昆虫数量最多。其中,在个体数与种数上,模式Ⅱ都最多。在个体数与物种数占比上,模式Ⅰ的捕食性昆虫个体数占比8.60%,钻蛀性昆虫个体数占比3.88%,寄生性昆虫个体数占比5.02%,中性昆虫个体数占比13.14%,是4种经营模式下山核桃林最高的。模式Ⅲ植食性及昆虫个体数占比最高(81.80%),模式Ⅰ植食性昆虫物种数占比最高(58.87%),模式Ⅲ中性昆虫物种数占比最高(11.58%),模式Ⅳ捕食性昆虫物种数占比(17.36%),钻蛀性昆虫物种数占比(12.40%)和寄生性昆虫物种数占比(7.44%)都最高。
表 5 不同经营模式下总昆虫群落功能结构
Table 5. Functional structure of all insect communities under different management modes
功能 经营模式 个体数/头 占比/% 种数/种 占比/% 捕食性 Ⅰ 317 8.60 22 15.60 Ⅱ 468 6.67 27 16.56 Ⅲ 249 4.98 14 14.74 Ⅳ 336 5.11 21 17.36 植食性 Ⅰ 2 555 69.35 83 58.87 Ⅱ 5 286 75.37 94 57.67 Ⅲ 4 094 81.80 53 55.79 Ⅳ 5 240 79.62 62 51.24 钻蛀性 Ⅰ 143 3.88 14 9.93 Ⅱ 232 3.31 17 10.43 钻蛀性 Ⅲ 114 2.28 11 11.58 Ⅳ 167 2.54 15 12.40 寄生性 Ⅰ 185 5.02 8 5.67 Ⅱ 285 4.06 10 6.13 Ⅲ 92 1.84 7 7.37 Ⅳ 154 2.34 9 7.44 中性昆虫 Ⅰ 484 13.14 14 9.93 Ⅱ 742 10.58 15 9.20 Ⅲ 456 9.11 11 11.58 Ⅳ 710 10.79 14 11.57 为了更好地了解不同经营模式下不同功能类群昆虫的内部机制和主导因素,分别对上述4种不同经营模式的山核桃林昆虫功能类群进行主分量分析。设定以下变量:捕食性昆虫个体数为X1,种数为X2;植食性昆虫个体数为X3,种数为X4;钻蛀性昆虫个体数为X5,种数为X6;寄生性昆虫个体数为X7,种数为X8;中性昆虫个体数为X9,种数为X10。主分量分析只分析马氏网法收集的昆虫。
由表 6可知:模式Ⅰ的第1主分量代表捕食性个体数和种数、寄生性个体数的综合因子,对昆虫群落的变化起主导作用;第2主分量代表植食性个体数和种数的综合因子。前2个主分量的累计贡献率为90.29%。模式Ⅱ的第1主分量代表捕食性个体数和中性昆虫个体数的综合因子,对昆虫群落的变化起主导作用;第2主分量代表寄生性种数、寄生性个体数及中性昆虫种数的综合因子;第3主分量代表钻蛀性种数和个体数的综合因子。前3个主分量的累计贡献率为95.95%。模式Ⅲ的第1主分量代表中性昆虫种数、钻蛀性种数、植食性物种数及寄生性种数的综合因子,对昆虫群落变化起主导作用;第2主分量代表植食性个体数和钻蛀性个体数的综合因子;第3主分量代表捕食性种数和植食性个体数的综合因子。前3个主分量累计贡献率为92.40%。模式Ⅳ的第1主分量代表寄生性种数和个体数、捕食性个体数及钻蛀性种数和个体数的综合因子,对昆虫群落变化起主导作用;第2主分量代表植食性种数和个体数及中性昆虫个体数的综合因子。前2个主分量累计贡献率为81.92%。
表 6 不同经营模式下马氏网法收集的昆虫群落主分量分析
Table 6. Principal component analysis of insect communities in Malaise trap under different management modes
经营模式 主分量 X1 X2 X3 X4 X5 X6 X7 X8 X9 X10 累计贡献/% Ⅰ 1 0.991 0.935 0.713 0.840 0.852 0.864 0.931 0.889 0.897 0.895 78.09 2 -0.043 -0.142 0.673 0.531 -0.130 -0.310 -0.255 -0.428 0.318 -0.044 90.29 Ⅱ 1 0.938 0.778 0.707 0.829 0.831 0.577 0.866 0.586 0.901 0.712 61.05 2 0.057 -0.516 -0.695 -0.536 0.085 0.201 0.476 0.805 -0.159 0.500 83.43 3 -0.213 0.177 -0.027 -0.082 0.453 0.772 0.005 0.011 -0.398 -0.456 95.95 Ⅲ 1 0.586 0.435 0.228 0.914 0.808 0.946 0.899 0.911 0.827 0.968 62.28 2 -0.715 -0.516 0.837 0.237 0.552 0.163 -0.183 -0.254 0.097 -0.049 82.05 3 0.146 0.676 0.491 -0.100 0.074 0.207 -0.188 0.029 -0.446 -0.150 92.40 Ⅳ 1 0.893 0.506 0.192 0.468 0.804 0.881 0.901 0.926 0.530 0.840 53.87 2 -0.239 -0.767 0.709 0.865 0.346 -0.208 -0.298 -0.342 0.703 0.214 81.92 -
对马氏网法收集的昆虫进行聚类分析。由图 2可知:在度量值d为5时,4种不同经营模式的昆虫群落可分为3类:模式Ⅱ和模式Ⅳ归为一类,两者昆虫群落结构较为接近;模式Ⅲ和模式Ⅰ分别为一类。这2种经营方式的群落结构有较大差异。

图 2 不同经营模式下马氏网法收集的昆虫群落聚类分析
Figure 2. Cluster analysis of insect communities in Malaise trap under different management modes
-
本研究共获得昆虫22 283号,共计9目84科163种,在物种数上优势类群均为鞘翅目和膜翅目昆虫,个体数量上优势类群均为同翅目和直翅目。由于山核桃林长久过度开发的管理模式,虽然近年来逐渐发生改变,但林下结构依然很单一[24],因此4种经营模式中优势类群均为相同。但是,经过数据分析,不难看出4种经营模式依然存在差异。在特征指数上,模式Ⅰ的经营方式昆虫多样性最好,模式Ⅱ次之,模式Ⅳ较模式Ⅲ更好。从相对稳定性分析来看,为模式Ⅰ的群落结构相较其他经营模式更稳定,模式Ⅱ次之,模式Ⅳ较模式Ⅲ更稳定。因此,模式Ⅰ的经营模式下生态坏境最稳定,模式Ⅱ次之,模式Ⅲ生态坏境最不稳定。
在4种经营模式中,植食性昆虫的种类和数量均最多,其中,均为山核桃蚜虫数量最多,但蚜虫聚居性、行动能力弱,防治相对简单。蝗科Acrididae昆虫数量均为第2位,蝗科昆虫是山核桃林中的重要害虫之一,因此限制山核桃林内蝗科昆虫的数量极为重要。鞘翅目也为山核桃林中的重要害虫类群,研究中发现,天牛科Cerambycidae昆虫种类较多,是其中重要的危害类群,高效防治山桃林内的天牛科昆虫对山核桃林的管理有着重要意义。
从主分量分析可以发现:不同经营模式下,昆虫群落变化的主导因素差异较大,说明4种经营模式存在一定的差异性。无人干扰的野生状态的山核桃林林内生态环境是4种经营模式中最稳定的,捕食性昆虫和寄生性昆虫主导着该群落的昆虫变化。林下种植植物的山核桃林的植被丰富,因此有更丰富的昆虫种类和数量,捕食性昆虫的食物更多来自中性昆虫,因此捕食性昆虫和中性昆虫比较重要。传统经营模式的山核桃林,因过多使用杀虫剂与除草剂,且生态系统是4种经营模式中最不稳定的,因此对昆虫群落的变化其主导因素的类群较多。模式Ⅳ为参照有机化管理的山核桃林,会对林内的害虫进行生态化控制,因此昆虫群落变化的主导因素为更难控制的几类昆虫。只需相应控制某一类昆虫种类和数量,就能极大改变该经营模式内昆虫群落的结构。这为山核桃林生态调控指明了新的方向。
昆虫群落结构能够在一定程度上反应其生存坏境的状态。在度量值为5时,模式Ⅱ和模式Ⅳ的经营模式较为接近。在度量值为10时,模式Ⅱ,模式Ⅲ和模式Ⅳ的经营模式较为接近。综上可知,模式Ⅰ的生态系统最为稳定,但在实际生产过程中,模式Ⅰ的经营模式不适合被采用,只可作为参考研究。林下种植植物的山核桃林经营方式更适合作为今后林农的经营发展方向,建议林农在保证成本与产量的前提下,参照林下种植植物的经营模式进行种植经营。
Insect community structures of Carya cathayensis forests with different management modes in Zhejiang Province
-
摘要: 通过分析4种不同经营模式下(荒废野生的山核桃Carya cathayensis林,林下种植植物的山核桃林,传统经营的山核桃林,参照有机化管理的山核桃林)昆虫群落结构的差异,对山核桃林经营模式提出意见与建议。采用马氏网、灯诱和样方调查收集昆虫标本,利用Excel 2013,SPSS 19.0软件对昆虫多样性特征参数、群落稳定性、主分量、群落相似性进行计算和分析。共获得标本22 283号,共计9目84科163种,在物种数上优势类群均为鞘翅目Coleoptera和膜翅目Hymenoptera昆虫,个体数量上优势类群均为同翅目Homoptera和直翅目Orthoptera。从特征性指数看,荒废野生的山核桃林昆虫多样性指数为3.64,均匀度指数为0.74,表现都为最好;林下种植植物的山核桃林昆虫多样性指数为3.12,均匀度指数为0.61,表现次之;马氏网法收集到的昆虫群落中,林下种植植物的山核桃林的昆虫物种数和个体数显著高于其他林(P < 0.05),荒废野生和林下种植植物的山核桃林的昆虫多样性指数和均匀度指数差异不显著,但显著高于其他林(P < 0.05),传统经营的山核桃林的昆虫优势度指数最高,且显著高于其他林(P < 0.05)。从群落结构稳定性上看,参照有机化管理的山核桃林的天敌与害虫种数比值(Sn/Sp)为0.39,表现最好,说明昆虫群落内部食物网的复杂程度及相互制约能力较高;荒废野生的山核桃林的群落物种数和个体数比值(St/Si)为0.04,表现最好,说明昆虫种间数量的相互制约能力较强。主分量分析表明:不同经营模式存在一定差异,昆虫群落变化的主导因素各不相同。相似性分析发现,在度量值为5时,林下种植植物的山核桃林和参照有机化管理的山核桃林的昆虫群落结构接近,荒废野生的山核桃林和传统经营的山核桃林的昆虫群落结构存在差异。荒废野生的山核桃林的昆虫生态结构最稳定,林下种植植物的山核桃林次之。但在实际生产过程中,由于无法荒废山核桃林,因此,林下种植植物的山核桃林经营模式更适合农户,也是未来山核桃林经营模式发展的方向。Abstract: To provide opinions and suggestions for management of Carya cathayensis forests, this research analyzed differences in insect communities using four different management modes:Type Ⅰ-a wild C. cathayensis forest, Type Ⅱ-vegetation planted under a C. cathayensis forest, Type Ⅲ-a traditional C. cathayensis forest, and Type Ⅳ-an organically managed C. cathayensis forest. Methods of collecting insect specimens included Malaise trap, light lure, and quadrat survey. Characteristic indexes of diversity, stability of insect community, principal components, and similarity were calculated and analyzed using Excel 2013 and SPSS 19.0 software. Results showed a total of 22 283 insect specimens being collected belonging to 163 species, 84 families, and 9 orders. Coleoptera and Hymenoptera were dominant groups for species numbers; whereas, Homoptera and Orthoptera were dominant groups for individual numbers. For the entire insect community the best performance was with management Type Ⅰ, the wild C. cathayensis forest, with a diversity index of 3.64 and an evenness index of 0.74. For Type Ⅱ, the diversity index was 3.12, and the evenness index was 0.61; this forest management type had good performance. For the insect communities in Malaise traps, the individual number and species number with Type Ⅱ management were significantly higher than the other management types (P < 0.05). The diversity index and the evenness index for Type Ⅰ and Type Ⅱ management were not significantly different but were significantly higher than other management types (P < 0.05). The dominance index with Type Ⅲ management was significantly higher than other management types (P < 0.05). For community structure, the Sn/Sp ratio for Type Ⅳ management was best with 0.39 meaning that the complexity and mutual restriction ability of the food network in the community was high. The St/Si ratio of the wild C. cathayensis forest was best with 0.04 meaning the mutual restriction of species number was stronger. The principal component analysis showed differences for different management modes. Also the dominant factors for changes in the insect communities were different. A similarity analysis found that when the measure was five, insect community structures were similar for Type Ⅱ and Type Ⅳ management. Overall, the ecological structure of the wild C. cathayensis forest (Type Ⅰ management) was most stable followed by Type Ⅱ, and because inefficient use of a C. cathayensis forest was forbidden in the actual production process, the management mode of vegetation planted under a C. cathayensis forest (Type Ⅱ) was recommended for farmers.
-
Key words:
- insect ecology /
- insect community /
- diversity /
- Carya cathayensis forests /
- management mode /
- Zhejiang
-
图 2 不同经营模式下马氏网法收集的昆虫群落聚类分析
Figure 2 Cluster analysis of insect communities in Malaise trap under different management modes
表 1 不同经营模式山核桃林分布概况
Table 1. Distribution of different management modes of Carya cathayensis forests
模式代号 经营模式 地点 经纬度 海拔/m 生境 Ⅰ 野生的山核桃林 板桥镇孟坞里 30°11′8.29″N, 119°44′57.46″E 100 多年无人管理,呈自由生长状态,林下植被复杂,以灌木、草本植物为主。无施药和施肥。林下覆盖度大于95% Ⅱ 林下种植植物的山核桃林 湍口镇湍口村 30°03′23.88″N, 119°08′29.80″E 182 林下种植多种绿肥作物包括黑麦草、紫花苜蓿、白三叶、菊科植物等;灌木有南天竹;乔木有香榧、黄枫、桂花、银杏、樟树等。以生态调控治理为主,其他治理手段为辅。林下覆盖度大于85% Ⅲ 传统经营的山核桃林 河桥镇寺坞里村 30°04′14.99″N, 119°10′50.02″E 108 施除草剂和杀虫剂,林下植被少且单一,主要为草本植物。林下覆盖度小于10% Ⅳ 参照有机化管理的山核桃林 河桥镇寺坞里村 30°04′12.40″N, 119°10′51.12″E 108 割草机除草,施放有机肥。林下植被少,林间套种经济作物,主要为黄精、香榧苗。林下覆盖度大于65% 说明:黑麦草Lolium multiflorum,紫花苜蓿Medicago sativa,白三叶Trifolium repens,南天竹Nandina domestica,香榧Torreya grandis ‘Merrillii’,黄枫Tri kvartala,桂花Osmanthus fragrans,银杏Ginkgo biloba,樟树Cinnamomum camphora,黄精Polygonatum sibiricum  下载: 导出CSV
下载: 导出CSV
表 2 不同经营模式的总昆虫群落
Table 2. All insect community under different management modes
类别 Ⅰ Ⅱ Ⅲ Ⅳ 种数/种 个体数/头 相对多度/% 种数/种 个体数/头 相对多度/% 种数/种 个体数/头 相对多度/% 种数/种 个体数/头 相对多度/% 膜翅目Hymenoptera 20 343 9.31 24 487 6.94 14 167 3.34 19 257 3.91 双翅目Diptera 16 540 14.66 17 822 11.72 14 464 9.27 16 649 9.86 直翅目Orthoptera 14 689 18.70 16 948 13.52 5 831 16.60 7 716 10.88 鞘翅目Coleoptera 37 414 11.24 44 683 9.74 23 275 5.49 32 431 6.55 异翅亚目Heteroptera 11 159 4.32 14 237 3.38 6 59 1.18 10 88 1.34 同翅亚目Homoptera 17 1 346 36.54 19 3 532 50.36 10 2 972 59.38 12 4 088 62.12 蜚蠊目Blattaria 1 87 2.36 1 169 2.41 1 149 2.98 1 241 3.66 螳螂目Mantodea 1 10 0.27 1 7 0.10 1 5 0.10 1 13 0.20 鳞翅目Lepidoptera 24 96 2.61 27 128 1.83 21 83 1.66 23 98 1.49 合计 141 3 684 100.00 163 7 013 100.00 95 5 005 100.00 121 6 581 100.00
下载: 导出CSV
表 3 不同经营模式的总昆虫群落特征值
Table 3. Characteristic value of all insect communities under different management modes
经营模式 个体数/头 种数/种 H′ E C Ⅰ 3 684 141 3.64 0.74 0.10 Ⅱ 7 013 163 3.12 0.61 0.20 Ⅲ 5 005 95 2.15 0.47 0.33 Ⅳ 6 581 121 2.38 0.50 0.31
下载: 导出CSV
表 4 不同经营模式的马氏网法收集的昆虫群落特征值
Table 4. Characteristic value of insect communities in Malaise trap under different management modes
经营模式 种数/种 个体数/头 H′ E C Ⅰ 103.20 ± 5.93 b 506.80 ± 101.59 bc 4.06 ± 0.07 a 0.88 ± 0.015 a 0.038 ± 0.005 c Ⅱ 119.00 ± 3.54 a 758.40 ± 43.90 a 4.16 ± 0.10 a 0.87 ± 0.021 a 0.033 ± 0.008 c Ⅲ 60.40 ± 1.82 d 431.00 ± 31.63 c 3.01 ± 0.07 c 0.74 ± 0.019 c 0.134 ± 0.013 a Ⅳ 78.20 ± 4.60 c 577.00 ± 29.89 b 3.52 ± 0.06 b 0.81 ± 0.004 b 0.066 ± 0.005 b 说明:同列不同小写字母表示差异显著(P<0.05)
下载: 导出CSV
表 5 不同经营模式下总昆虫群落功能结构
Table 5. Functional structure of all insect communities under different management modes
功能 经营模式 个体数/头 占比/% 种数/种 占比/% 捕食性 Ⅰ 317 8.60 22 15.60 Ⅱ 468 6.67 27 16.56 Ⅲ 249 4.98 14 14.74 Ⅳ 336 5.11 21 17.36 植食性 Ⅰ 2 555 69.35 83 58.87 Ⅱ 5 286 75.37 94 57.67 Ⅲ 4 094 81.80 53 55.79 Ⅳ 5 240 79.62 62 51.24 钻蛀性 Ⅰ 143 3.88 14 9.93 Ⅱ 232 3.31 17 10.43 钻蛀性 Ⅲ 114 2.28 11 11.58 Ⅳ 167 2.54 15 12.40 寄生性 Ⅰ 185 5.02 8 5.67 Ⅱ 285 4.06 10 6.13 Ⅲ 92 1.84 7 7.37 Ⅳ 154 2.34 9 7.44 中性昆虫 Ⅰ 484 13.14 14 9.93 Ⅱ 742 10.58 15 9.20 Ⅲ 456 9.11 11 11.58 Ⅳ 710 10.79 14 11.57
下载: 导出CSV
表 6 不同经营模式下马氏网法收集的昆虫群落主分量分析
Table 6. Principal component analysis of insect communities in Malaise trap under different management modes
经营模式 主分量 X1 X2 X3 X4 X5 X6 X7 X8 X9 X10 累计贡献/% Ⅰ 1 0.991 0.935 0.713 0.840 0.852 0.864 0.931 0.889 0.897 0.895 78.09 2 -0.043 -0.142 0.673 0.531 -0.130 -0.310 -0.255 -0.428 0.318 -0.044 90.29 Ⅱ 1 0.938 0.778 0.707 0.829 0.831 0.577 0.866 0.586 0.901 0.712 61.05 2 0.057 -0.516 -0.695 -0.536 0.085 0.201 0.476 0.805 -0.159 0.500 83.43 3 -0.213 0.177 -0.027 -0.082 0.453 0.772 0.005 0.011 -0.398 -0.456 95.95 Ⅲ 1 0.586 0.435 0.228 0.914 0.808 0.946 0.899 0.911 0.827 0.968 62.28 2 -0.715 -0.516 0.837 0.237 0.552 0.163 -0.183 -0.254 0.097 -0.049 82.05 3 0.146 0.676 0.491 -0.100 0.074 0.207 -0.188 0.029 -0.446 -0.150 92.40 Ⅳ 1 0.893 0.506 0.192 0.468 0.804 0.881 0.901 0.926 0.530 0.840 53.87 2 -0.239 -0.767 0.709 0.865 0.346 -0.208 -0.298 -0.342 0.703 0.214 81.92
下载: 导出CSV
-
[1] CHRISTENSEN N L, BARTUSK A M, BROWN J H, et al. The report of the Ecological Society of America Committee on the scientific basis for ecosystem management[J]. Ecol Appl, 1996, 6(3):665-691. [2] 国家环境保护局.中国环境保护21世纪议程[M].北京:中国环境科学出版社, 1995. [3] EYRE M D, LOTT D A, GARSID A. Assessing the potential for environment monitoring using ground beetles(Coleoptera:Carabidae) with riverside and Scottish data[J]. Ann Zool Fenn, 1996, 33(1):157-163. [4] 李浩.有机果园与非有机果园节肢动物群落结构及多样性比较[D].天津: 天津师范大学, 2010. LI Hao. Comparison of the Structure and Diversity of Arthropod Communities in Organic Orchard and Non Organic Orchard[D]. Tianjin: Tianjin Normal University, 2010. [5] 马玲, 问荣荣, 焦玥, 等.小兴安岭不同林型的昆虫多样性[J].林业科学, 2016, 52(2):82-90. MA Ling, WEN Rongrong, JIAO Yue, et al. Insect diversity of different forest types in Xiaoxing'anling Mountains[J]. Sci Silv Sin, 2016, 52(2):82-90. [6] 徐燕, 沈月琴, 黄坚钦, 等.农户对山核桃生态化经营模式的意愿分析[J].浙江林学院学报, 2010, 27(5):750-756. XU Yan, SHEN Yueqin, HUANG Jianqin, et al. Farmers' willingness to adopt ecological management model for Carya cathayensis[J]. J Zhejiang For Coll, 2010, 27(5):750-756. [7] 吕秋菊, 沈月琴, 高宇列, 等.山核桃产业的发展过程、动因及展望[J].浙江农林大学学报, 2012, 29(1):97-103. LÜ Qiuju, SHEN Yueqin, GAO Yulie, et al. Development process, agents and prospect of hickory industry[J]. J Zhejiang A & F Univ, 2012, 29(1):97-103 [8] 吕惠进.浙江临安山核桃立地环境研究[J].森林工程, 2005, 21(1):1-6. LÜ Huijin. Research on site environment of Carya cathayensis in Lin'an Zhejiang[J]. For Eng, 2005, 21(1):1-6. [9] 徐公天, 杨志华.中国园林害虫[M].北京:中国林业出版社, 2007. [10] 钱皆兵.宁波林业害虫原色图谱[M].北京:中国农业科学技术出版社, 2012. [日]株式会社学习研究社.青少年百科知识图鉴: 昆虫[M].杨定, 译.郑州: 河南科学技术出版社, 2004. [12] 王直诚.东北天牛志[M].长春:吉林科学技术出版社, 2003. [13] 胡国良, 俞彩珠.山核桃病虫害防治彩色图谱[M].北京:中国农业出版社, 2005. [14] 张巍巍, 李元胜.中国昆虫生态大图鉴[M].重庆:重庆大学出版社, 2011. [15] 顾伟, 马玲, 丁新华, 等.扎龙湿地不同生境的昆虫多样性[J].应用生态学报, 2011, 22(9):2405-2412. GU Wei, MA Ling, DING Xinhua, et al. Insect diversity in different habitats of Zhalong Wetland[J]. Chin J Appl Ecol, 2011, 22(9):2405-2412. [16] 马玲, 顾伟, 丁新华, 等.扎龙湿地昆虫群落结构及动态[J].生态学报, 2011, 31(5):1371-1377. MA Ling, GU Wei, DING Xinhua, et al. Structure and dynamics of insect communities in Zhalong Wetland[J]. Acta Ecol Sin, 2011, 31(5):1371-1377. [17] 麻应太, 李春宁, 崔俊, 等.秦岭牛背梁保护区地面昆虫的多样性[J].西北林学院学报, 2013, 28(3):142-148. MA Yingtai, LI Chunning, CUI Jun, et al. Diversity of ground insects in Qinlin Niubeiliang Reserve[J]. J Northwest For Univ, 2013, 28(3):142-148. [18] 仲雨霞, 付必谦.北京白河湿地夏季昆虫群落的多样性及空间分布格局[J].首都师范大学学报, 2013, 34(5):18-26. ZHONG Yuxia, FU Biqian. The diversity and spatial distribution pattern of the insect communities in the Beijing Baihe wetland in summer[J]. J Cap Norm Univ, 2013, 34(5):18-26. [19] MAGURRA A E. Measuring Biological Diversity[M]. Oxford:Blackwell Science, 2004. [20] 冯琦雅, 陈超凡, 覃林, 等.不同经营模式对蒙古栎天然次生林林分结构和植物多样性的影响[J].林业科学, 2018, 54(1):12-21. FENG Qiya, CHEN Chaofan, QIN Lin, et al. Effects of different management models on stand structure and plant diversity of natural secondary forests of Quercus mongolica[J]. Sci Silv Sin, 2018, 54(1):12-21. [21] 高宝嘉, 张执中, 李镇宇, 等.封山育林与昆虫群落及多样性稳定性影响的研究[J].生态学报, 1992, 12(1):1-7. GAO Baojia, ZHANG Zhizhong, LI Zhenyu, et al. Study on the effect of closing afforestation and insect communities diversity and stability[J]. Acta Ecol Sin, 1992, 12(1):1-7. [22] 高宝嘉, 申曙光, 王正文, 等.园林昆虫群落时间结构及动态研究[J].生态学报, 1998, 18(2):193-197. GAO Baojia, SHEN Shuguang, WANG Zhengwen, et al. Study on time structure and dynamics of garden insect communities[J]. Acta Ecol Sin, 1998, 18(2):193-197. [23] 郝树广, 张孝羲, 程遐年, 等.稻田节肢动物群落营养层及优势功能集团的组成与多样性动态[J].昆虫学报, 1998, 41(4):343-353. HAO Shuguang, ZHANG Xiaoxi, CHEN Xianian, et al. The dynamics of biodiversity and the composition of nutrition classes and dominant guilds of arthropoda community in paddy field[J]. Acta Entomol Sin, 1998, 41(4):343-353. [24] 张锋, 陈志杰, 张淑莲, 等.纸房沟流域植被恢复区乔木林昆虫群落结构及动态分析[J].环境昆虫学报, 2010, 32(4):445-452. ZHANG Feng, CHEN Zhijie, ZHANG Shulian, et al. Structure and dynamic analysis of arbor forest insect communities in the vegetation restoration area of the Zhifang Valley[J]. J Environ Entomol, 2010, 32(4):445-452. -

-
链接本文:
https://zlxb.zafu.edu.cn/article/doi/10.11833/j.issn.2095-0756.2019.03.001
 点击查看大图
点击查看大图
计量
- 文章访问数: 4330
- HTML全文浏览量: 1012
- PDF下载量: 224
- 被引次数: 0
