




-
雷达探测木材内部异常结构主要通过分析发射的电磁波的反射信息来实现。当木材内部存在孔洞时,因孔洞与木材本身具有不同的介电性能,进入木材的电磁波在穿过内部孔洞时会在两者的界面上发生反射。分析反射回波波形资料,可以推断木材内部孔洞情况。利用雷达对木材进行扫描可以快速获知木材内部是否存在孔洞。雷达波在不同含水率不同树种等条件下具有不同的传播速度,其检测图像若采取统一的方法进行分析会存在一定误差。大量研究表明:木材孔洞的探测精度与被测对象的介电常数密切相关,而木材的含水率、纤维密度和纤维方向等均会对介电常数产生一定影响[1],其中木材含水率与介电常数密切相关[2],因此分析木材含水率与介电常数的关系对开展雷达检测木材内部缺陷研究非常重要[3]。由此也可逆向通过雷达回波信号进行木材含水率的预估[4]。在实际应用过程中,针对雷达探测木材内部缺陷存在解析困难、探测精度不高、可视化程度低等不足,国内外学者展开了广泛研究。可行性及可靠性方面,LORENZO等[5]研究了雷达波对树干以及树根检测的可行性;邸向辉等[6]指出:电磁波信号解释分析的困难性是树木无损探测的主要制约因素;JEZOVA等[7]开展了树木内部残损的雷达图像特征研究;WU等[8]评估了探地雷达技术在探测不同树种内部损伤方面的可靠性。成像精度及算法方面,DEVARU等[9]阐述了探地雷达在树木无损检测方面的优点,并设计试验研究了探地雷达对树干内部缺陷的自动检测算法;BUTNOR等[10]采用阈值法对针叶树木内部缺陷进行了探测,并获得了较好的效果;李伟林等[11]对颐和园界河桥旁6株古柳Salix spp.进行雷达探测,并采用匹配滤波器算法计算回波时延以提高检测精度;肖夏阳等[12]通过比较阈值法、匹配滤波器法和希尔伯特积算法在树木内部层位探测的准确性,得出希尔伯特积算法对树木内缺陷区域的检测更加准确。陈勇平等[13]研究也发现:木材含水率等对雷达检测孔洞面积的影响较大,并开展树种、含水率等对雷达预估孔洞面积的初步分析。本研究旨在探究基于雷达检测的木材内部孔洞面积预估修正方法,提高雷达检测精度,从而为木材内部残损检测提供理论和数据支撑。
-
马尾松Pinus massoniana木材采集于湖南省攸县,造林年度为1966年,树龄50 a,胸径约51 cm,树高约22 m。杉木Cunninghamia lanceolata木材采集于湖南省攸县,造林年度为1982年,树龄34 a,胸径约46 cm,树高约25 m。
-
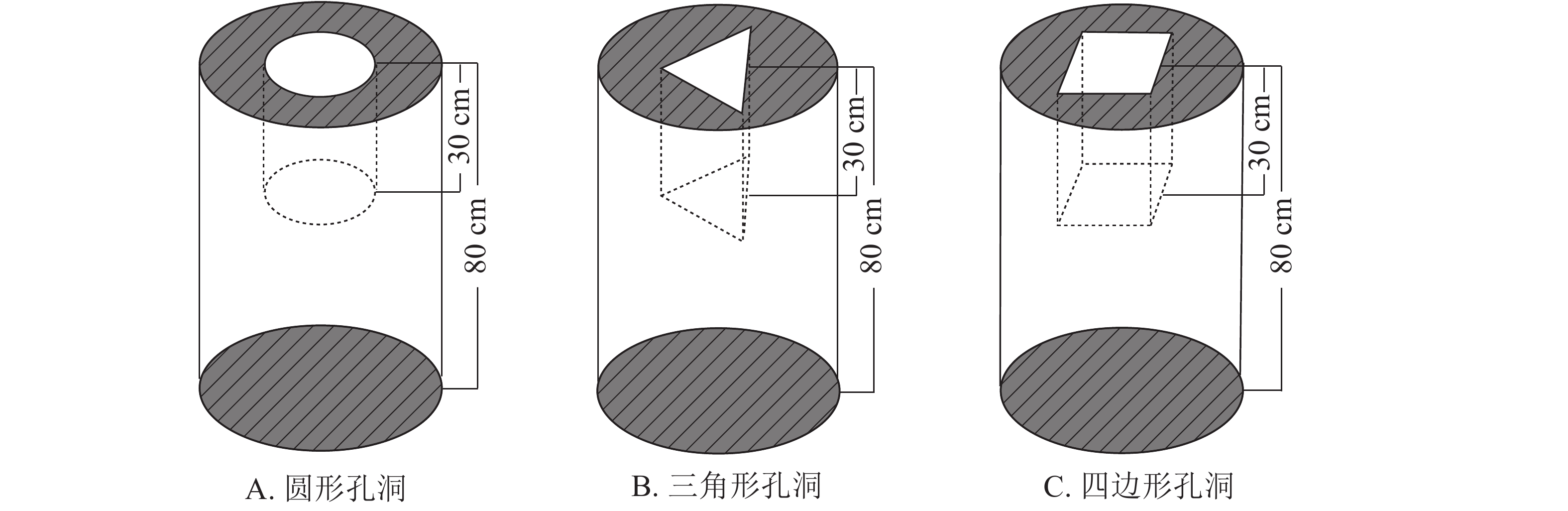
马尾松和杉木原木伐倒后,自树根位置80 cm起按每段长约80 cm逐个截取木段,以模拟短柱;截断后的部分木段通风干燥3 a,使其达到北京地区的平衡含水率。对干燥后的木段进行挑选并分别编号为S1、S2、S3、S4、S5、S6和C1用于实验研究,每个木段标记其起始检测点,其中S1~S6为马尾松试件,C1为杉木试件。取S2、S3、S4木段在其内部加工大小不一致的圆形孔洞;取S5木段在其内部加工三角形孔洞;取S6木段在其内部加工四边形孔洞;取C1木段在其内部加工圆形孔洞;S1为备用木段。以上木段的孔洞加工均位于中心位置,孔洞深度约30 cm(图1)。
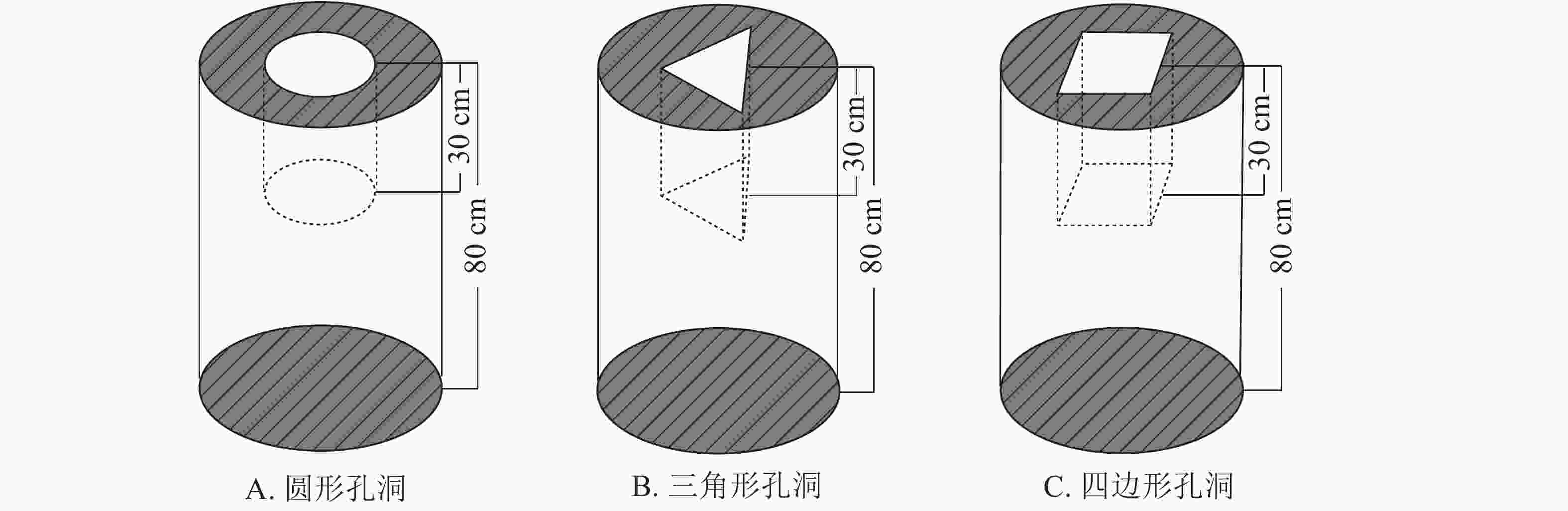
图 1 孔洞加工示意图
Figure 1. Schematic of internal cavity in wood columns
-
采用树木雷达对上述试材进行雷达扫描[13],其中S3、S4、S5、S6为试验用材,S2和C1为数据分析验证试材。本次测试研究采用美国树木雷达检测系统[Tree Radar Unit(TRU)],主要参数设置为:频率900 MHz,最大扫描直径4.2 m,深度分辨率1.9 mm,数据采集间隔5 mm。
-
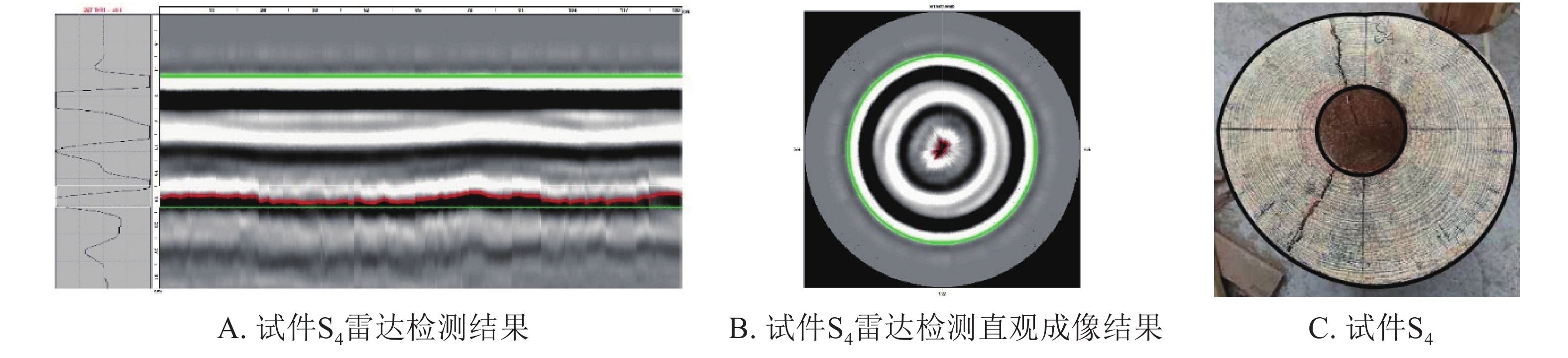
采用雷达扫描技术对带孔试材进行检测。以S4试件为例,通过分析雷达波反射波形和检测图像可以直观判定木材内部孔洞存在(图2A),但雷达预测孔洞面积(图2B)与实际孔洞面积(图2C)存在较大的偏差。
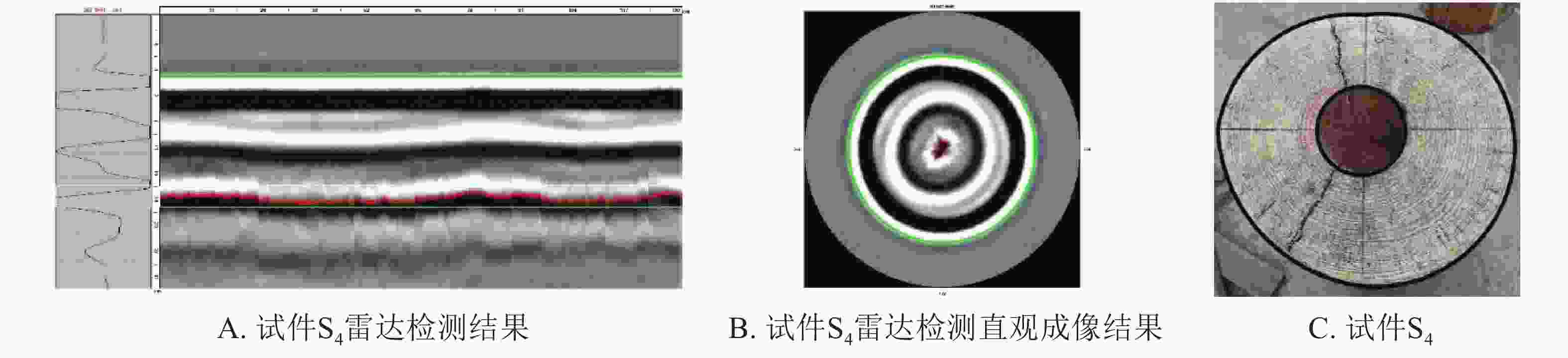
图 2 雷达成像直观结果
Figure 2. Intuitionistic results of the cavity areas in wood columns by TRU
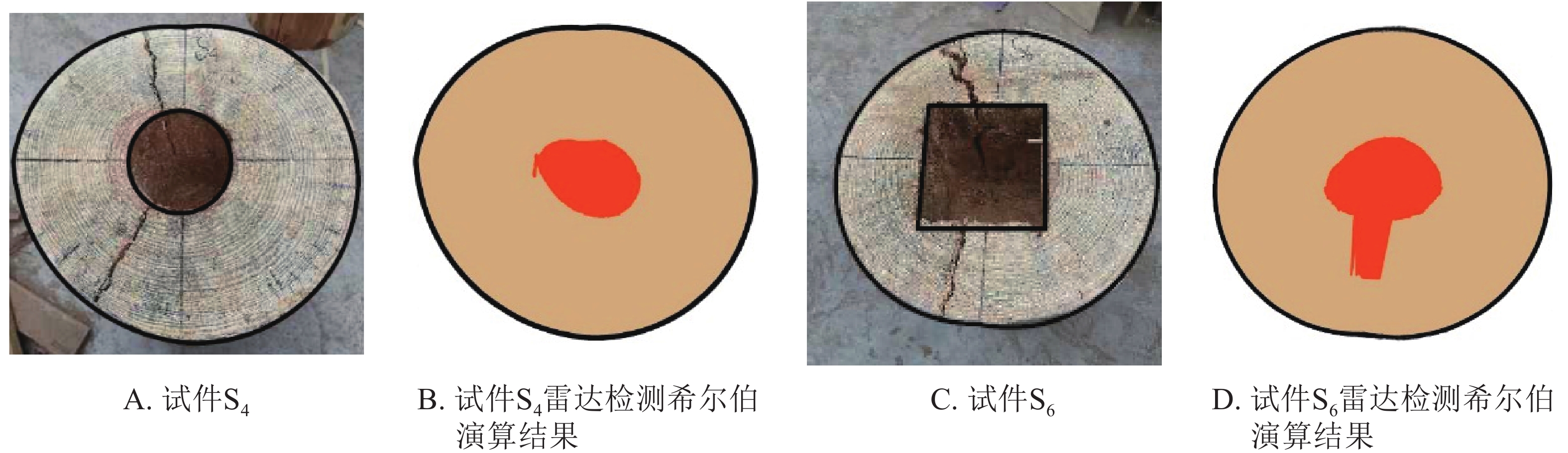
基于直观成像的误差,且希尔伯特积算法相较于阈值法和匹配滤波器法对木质体内部缺陷识别效果更好[12,14],本研究首先对上述检测结果进行希尔伯特积算法修正,其部分检测结果如图3所示。本研究采用层剥反演法获得木材介电常数并对其内部孔洞进行定位,结合检测对象外轮廓数据,可实现内部孔洞较准确定位。
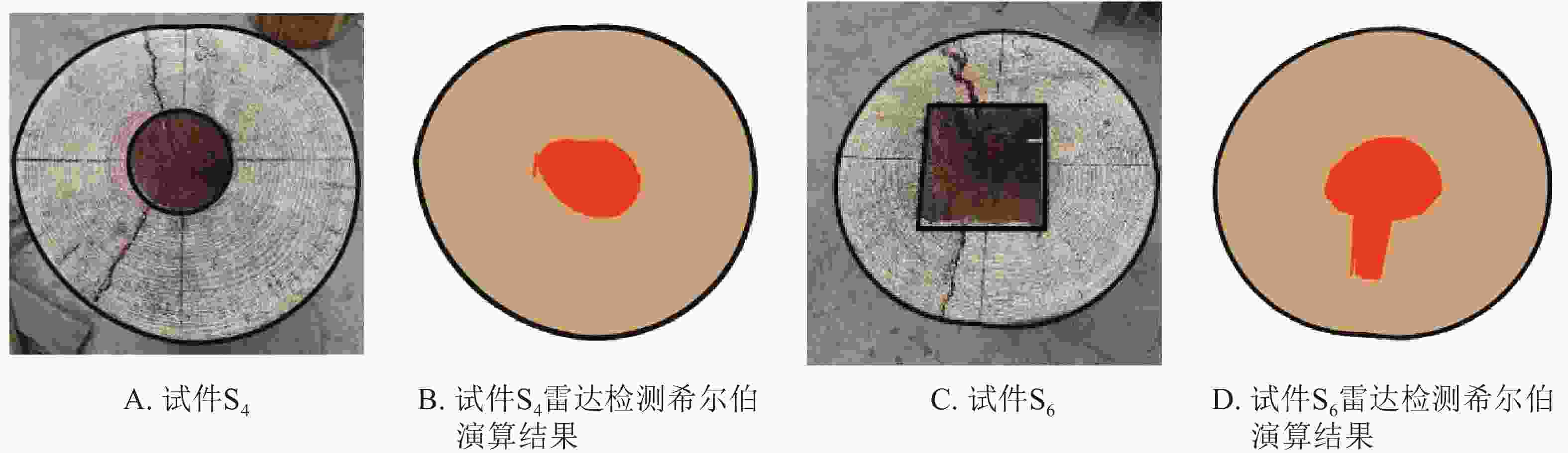
图 3 希尔伯特积算法演算结果
Figure 3. Analysis results of the cavity areas in wood columns by Hilbert
结合图2和图3可以看出:利用希尔伯特积算法对雷达检测图像进行修正,提高了检测精度,但还是存在一定误差,修正面积仍低于实际孔洞面积,其中试件S4修正图像的相对误差为20.57%,试件S6修正图像的相对误差为41.49%。这主要是因为希尔伯特积算法是根据层剥反演获得相对介电常数,从而对木材内部的传输速度进行在线估测,而实际过程该介电常数可能与估算的介电常数存在一定得误差。为此,本研究根据希尔伯特积算法原理,采取提取单路径检测数据,并使用曲线偏移、介电常数修正、辅助系数校正等方法尝试进一步的修正研究。
-
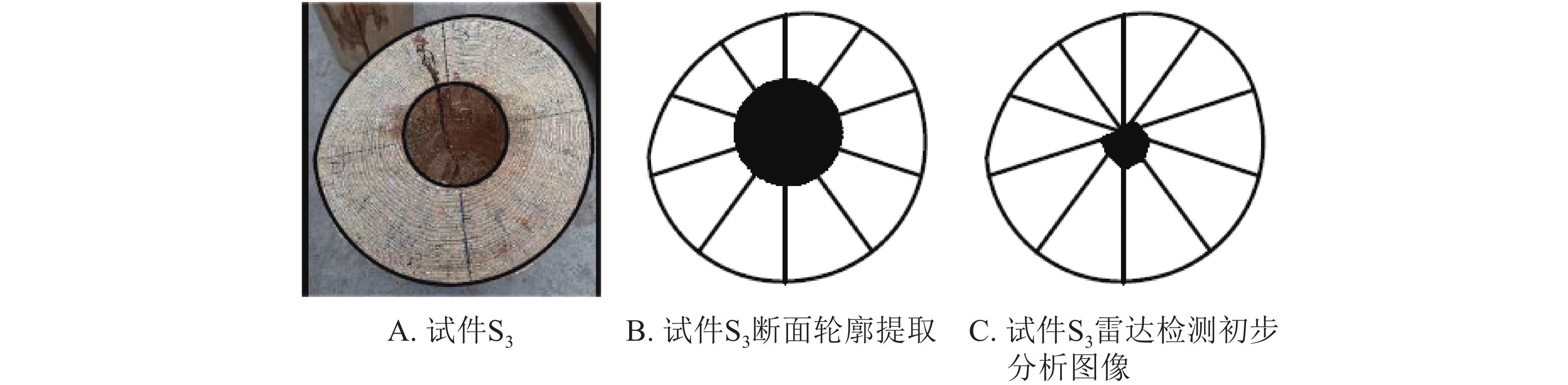
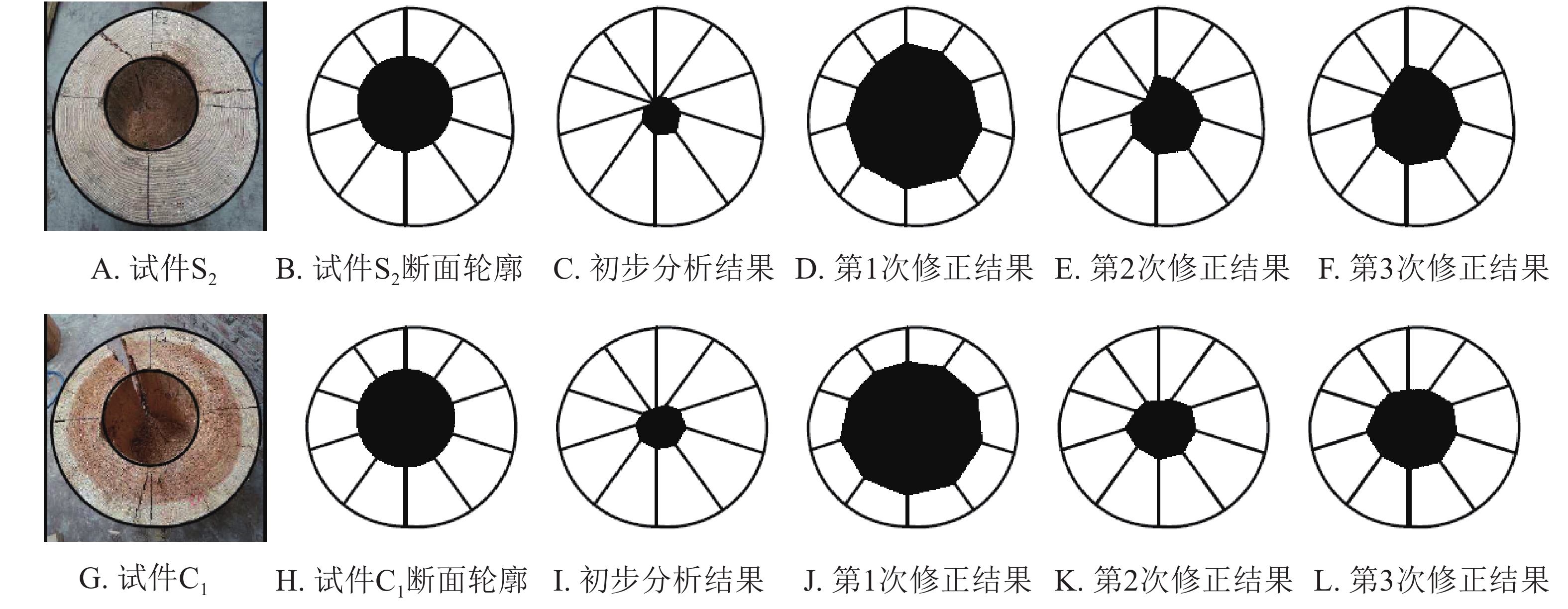
根据雷达检测原理,树木雷达工作时雷达天线绕该木柱一周相当于虚拟钻孔机对该木柱进行了360°的“钻孔”操作。本研究对检测对象进行圆周10等分,并提取等分点上每一次的“钻孔”结果。每一次“钻孔”结果即对应雷达检测中每一个位置的回波图形。通过回波图形分析可获得对应位置木材边缘与孔洞边缘距离,其中孔洞边缘即高介电常数物质(木材)与低介电常数物质(孔洞)交界面,交界面位置在回波图形中的特征为谷—峰—谷,叠加后在雷达检测图像中表现的形态为黑—白—黑。根据“钻孔”检测结果,将得到10个孔洞边界点,将边界点进行连线,即形成了内部孔洞轮廓。以试件S3为例,结果如图4。

图 4 雷达检测初步分析结果
Figure 4. Preliminary results of the cavity areas in wood columns by TRU
图4A为试件S3;图4B为图4A的断面轮廓提取,其目的是便于检测对象横断面及孔洞面积的计算;图4C为雷达检测初步分析图像。为了便于数据比对,本研究对试件S3、S4、S5、S6的雷达初步检测结果进行了测算和统计。结果见表1。
表 1 雷达检测初步测算与统计
Table 1. Preliminary calculation and statistics of the cavity areas in wood columns by TRU
序号 孔洞类型 实际孔洞百分比/% 雷达测算孔洞百分比/% S3 圆形孔洞 14.67 2.15 S4 圆形孔洞 9.66 0.84 S5 三角形孔洞 12.82 1.59 S6 四边形孔洞 20.26 2.53 从表1和图4可以看出:通过雷达检测可以快速判定木材内部孔洞的存在,但其预估的孔洞面积与实际面积相差较大。这主要是因为雷达检测内部孔洞的轮廓线产生了一定的偏移,且实际检测对象的介电常数与预设常数相差较大,导致预估孔洞面积与实际孔洞面积相差甚远。从表1和图4结果也可以看出:雷达检测预估内部孔洞面积若不加以修正,其数值远小于实际孔洞面积,无法直接应用于古建筑木构件内部残损的定量评估。
-
雷达波在介质中的传播速度可以简化为
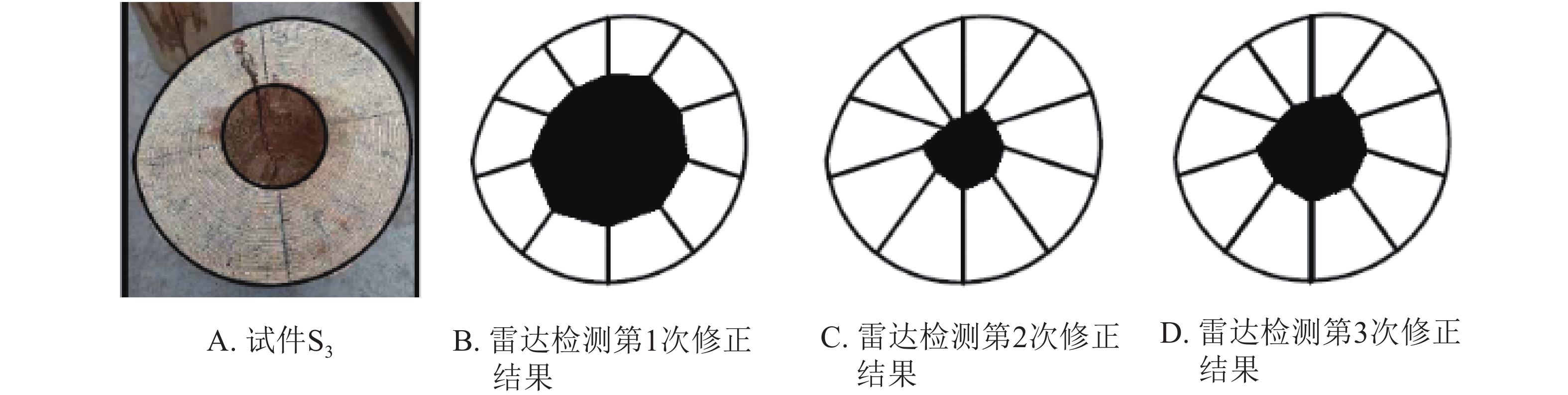
$ v=\dfrac{c}{\sqrt{{\rm{\varepsilon }}}} $ ,其中:c为雷达波在真空中的传播速度;ε为介质的介电常数[15]。由此,木材边缘与孔洞边缘的距离可以简化为$ s=\dfrac{{\rm{c}}t}{2\sqrt{{\rm{\varepsilon }}}} $ ,其中:t为雷达波传播时间。假设“钻孔”操作中木材边缘与孔洞边缘的距离为sr,实测偏移量为a,则雷达检测中木材边缘与孔洞边缘距离第1次修正为s1c=sr−a。同时,根据雷达检测系统预设介电常数ε1和马尾松木材介电常数ε2[16-17],设定介电常数修正系数为$ {k}_{1}=\dfrac{\sqrt{{{\rm{\varepsilon }}}_{1}}}{\sqrt{{{\rm{\varepsilon }}}_{2}}} $ ,则雷达检测中木材边缘与孔洞边缘距离第2次修正为s2c=(sr−a)k1。以试件S3为例,修正结果见图5。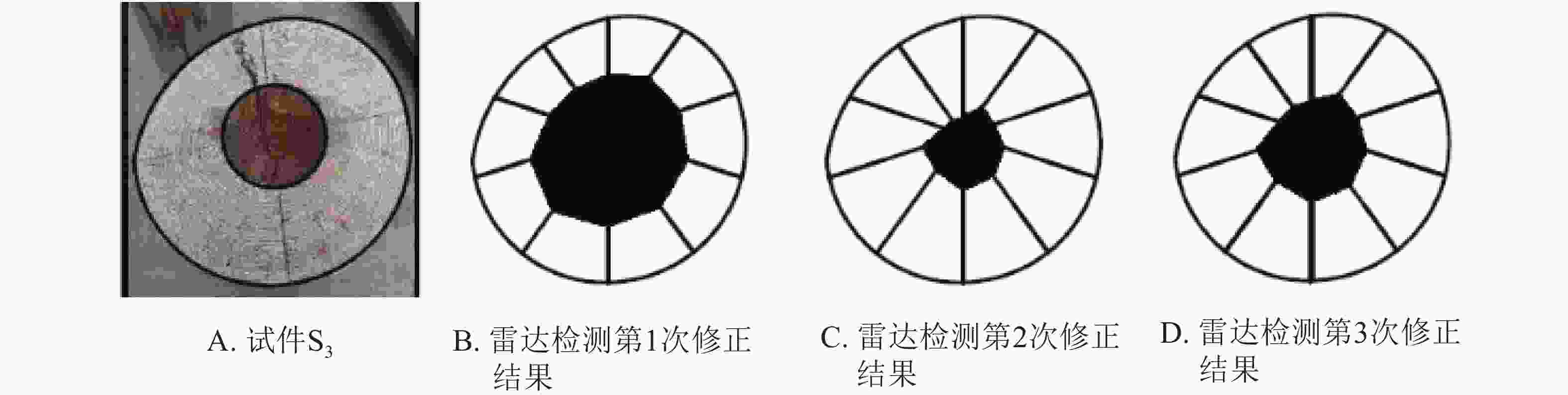
图 5 雷达检测数据修正结果
Figure 5. Corrected results of the cavity areas in wood columns by TRU
图5A为试件S3;图5B为根据偏移量进行第1次修正后的孔洞图像;图5C为根据介电常数进行第2次修正后的孔洞图像;图5D为3次修正后的孔洞图像。为了便于分析,同样对试件S3、S4、S5、S6的雷达检测数据修正结果进行了计算和统计。从表2和图5可以看出:根据偏移量进行第1次修正后,识别精度得到一定得提升,但其与实际孔洞相比均表现为偏大,这主要是因为修正距离s1c是根据雷达检测系统预设介电常数ε1计算得来,但实际检测对象的介电常数为ε2,在气干木材的检测中预设介电常数ε1大于检测对象介电常数ε2,故而s1c距离偏小,导致孔洞识别偏大。从表2和图5也可以看出:根据介电常数进行第2次修正后,其检测修正面积与实际孔洞面积仍存在一定偏差。从图5也可以看出:无论是否修正,雷达识别孔洞形状的能力均较差,这可能是因为外部形状等因素引起了内部图像的扭曲。
表 2 雷达检测数据修正与统计
Table 2. Corrected calculation and statistics of the cavity areas in wood columns by TRU
序号 残损类型 第1次修正后孔洞百分比/% 第2次修正后孔洞百分比/% 第3次修正后孔洞百分比/% S3 圆形孔洞 30.21 6.85 13.26 S4 圆形孔洞 26.51 6.39 12.49 S5 三角形孔洞 38.03 8.70 15.30 S6 四边形孔洞 37.71 8.75 15.29 上述研究结果表明:单独依靠介电常数比值进行修正,其孔洞识别面积与实际孔洞仍存在一定偏差。为此,本研究设置了另一辅助修正系数k2,即雷达检测中木材边缘与孔洞边缘距离按照s3c=(sr−a)k1k2进行修正,其中辅助修正系数k2为常数。经过前期的研究分析和数据拟合,可初步确定常数k2取值为0.80~0.90,本研究k2取值为0.85。从表1和表2可以看出:雷达检测结果进行3次修正后,其识别精度得到明显提升,总体相对误差小于30%。
-
为验证上述结论,本研究选取了具有不同大小孔洞马尾松试材S2和杉木试材C1进行补充试验,并根据上述研究步骤获取其断面轮廓、雷达检测初步结果、第1次修正结果、第2次修正结果和第3次修正结果,经数据整理见图6。
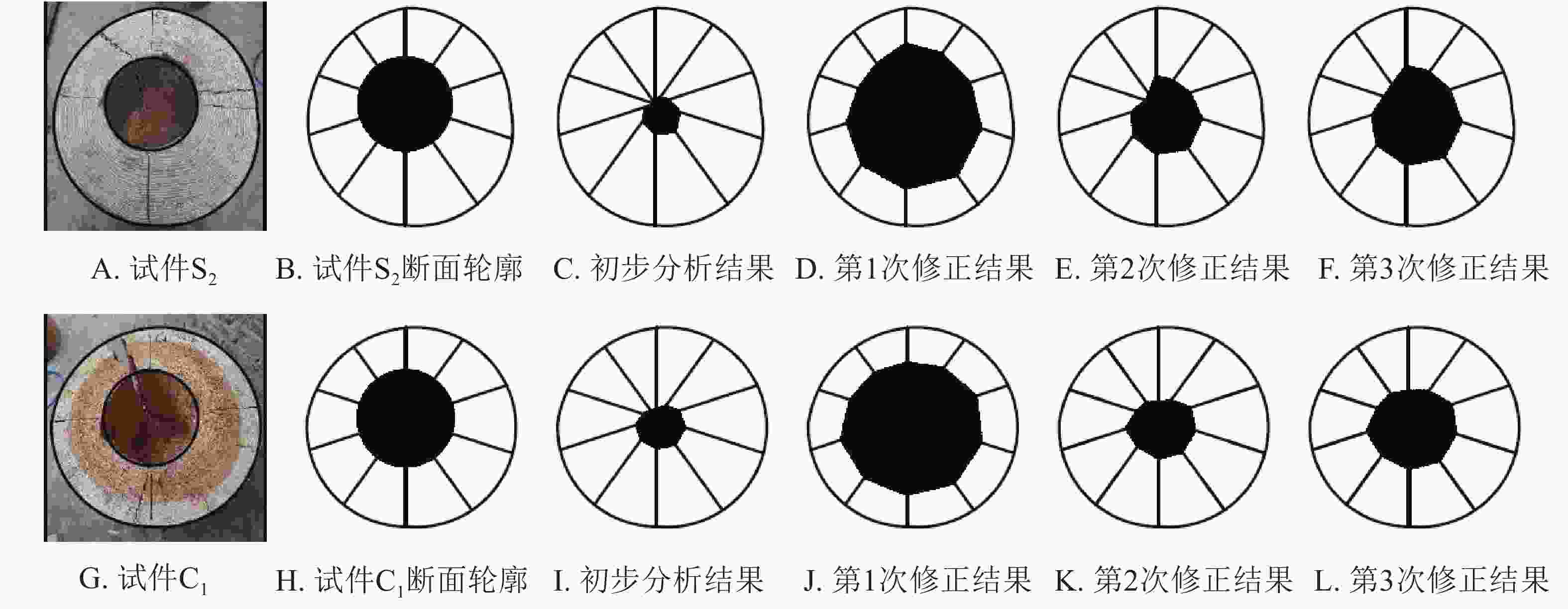
图 6 雷达检测数据修正过程
Figure 6. Corrected process of the cavity areas in wood columns by TRU
从验证试验可以得出同样结论,未修正或按照一定含水率下特定树种的介电常数修正的雷达图像其预估孔洞面积与孔洞实际面积均存在一定误差,按照s3c=(sr−a)k1k2对图像进行3次修正后精度较高。3次修正后,试材S2雷达检测预估孔洞面积相对误差为9.47%,试材C1预估相对误差为28.72%,检测结果可能会因为个体差异产生一定的偏差,但大致趋势相同。
-
通过分析雷达回波图形和检测图像可以快速判定松木和杉木木材内部孔洞的存在,但其直观成像技术在定量评估方面误差较大,需进行一定的数据处理和修正。①对带孔木材进行雷达扫描,未经修正的雷达图像其预估孔洞面积小于实际孔洞面积,按介电常数进行修正的雷达图像其预估孔洞面积与实际孔洞面积仍存在一定的误差。②无论是否进行数据处理和修正,利用雷达扫描进行内部孔洞的形状预估难度较大,难以准确识别三角形、四边形、圆形等各种形状的孔洞,应开展进一步研究。③实际应用中,根据树种和含水率所得到的介电常数应进行再次修正,即实际应用中根据树种和含水率所得到的介电常数仅为多参数校正中的一个因素。④利用该雷达检测与修正方法进行木材内部孔洞探测,综合树种和含水率等参数,可确保孔洞面积的识别误差小于30%。此项技术可应用于古建筑木构件内部残损的检测评估。⑤在开展古建筑木构件内部残损的雷达检测与定量评估前,应首先调查分析木结构建筑常用的树种类型,并建立常用树种、含水率与介电常数之间的关系模型。
Calculation and correction of wood internal cavity area based on radar detection
-
摘要:
目的 开展木材内部孔洞的雷达检测研究,探讨该检测手段下孔洞成像的准度和精度,实现木材内部孔洞的雷达快速识别和定量表征。 方法 以古建筑木结构常用的马尾松Pinus massoniana木材和杉木Cunninghamia lanceolata木材为研究对象,采取人工模拟的方法在木段端部制作异型孔洞,并利用雷达扫描仪对带孔木段进行检测,比较分析直观成像技术与希尔伯特积算法、单路径数据提取合成以及成像轮廓多参数校正等处理方法之间的差异,提出基于雷达检测的木材内部孔洞面积测算与可行的修正方法。 结果 利用雷达无损检测技术可以对马尾松和杉木木材内部孔洞的存在进行快速识别,但其直观成像技术在定量评估方面误差较大,需进行相关的数据处理和修正;未经修正的雷达图像其预估孔洞面积小于实际孔洞面积,通过希尔伯特积算法演算或介电常数修正的雷达图像其预估孔洞面积与实际孔洞面积仍存在一定的误差;木材边缘与孔洞边缘的距离应按s3c=(sr-a)k1k2进行修正,即实际应用中根据树种和含水率所得到的介电常数仅为多参数校正中的一个因素;无论是否进行数据处理和修正,雷达检测难以准确识别孔洞的具体形状,应开展进一步研究。 结论 该雷达检测及修正方法应用于木材内部孔洞探测,综合树种和含水率数值,可确保孔洞面积识别误差小于30%。图6表2参17 Abstract:Objective The objective of this study is to realize the rapid recognition and quantitative characterization of the internal cavities in wood by evaluating the accuracy and precision of the cavity imaging under radar detection method. Method Pinus massoniana and Cunninghamia lanceolata, commonly used in ancient building timber structure, were taken as the research object. Artificial simulation was used to make special-shaped cavities at the end of the logs to simulate the internal damages, and then the radar scanner was used to detect the cavities. The differences between the visual imaging technology and the processing methods such as the Hilbert integral method, single path data extraction and synthesis, and the multi-parameter correction of the image contour were compared and analyzed. The calculation and feasible correction method of wood internal cavity area based on radar detection was proposed. Result The internal cavity of P. massoniana and C. lanceolata wood could be recognized rapidly by the radar nondestructive testing technology, but the visual imaging technology had a large error in quantitative evaluation, which required further relevant data processing and correction. The estimated cavity area of the unmodified radar image was smaller than that of the actual one. There was still a certain error between the actual cavity area and the estimated cavity area of the radar image calculated by Hilbert integral method or modified by dielectric constant. The distance between the wood edge and the cavity edge should be corrected according to the formula s3c=(sr−a)k1k2, that was to say, the dielectric constant obtained by tree species and moisture content in practical application was only one factor in the multi-parameter correction. Therefore, it was difficult for radar scanner to accurately recognize the specific shape of the cavity, regardless of data processing and correction, and further research should be carried out. Conclusion The radar detection and correction technology can be applied to the detection of cavities in wood and the error of cavity area identification can be less than 30% by integrating tree species and moisture content values. [Ch, 6 fig. 2 tab. 17 ref.] -
Key words:
- wood science /
- ancient building timber structure /
- cavity /
- radar /
- quantitative evaluation /
- dielectric constant
-
图 2 雷达成像直观结果
Figure 2 Intuitionistic results of the cavity areas in wood columns by TRU
图 3 希尔伯特积算法演算结果
Figure 3 Analysis results of the cavity areas in wood columns by Hilbert
图 4 雷达检测初步分析结果
Figure 4 Preliminary results of the cavity areas in wood columns by TRU
表 1 雷达检测初步测算与统计
Table 1. Preliminary calculation and statistics of the cavity areas in wood columns by TRU
序号 孔洞类型 实际孔洞百分比/% 雷达测算孔洞百分比/% S3 圆形孔洞 14.67 2.15 S4 圆形孔洞 9.66 0.84 S5 三角形孔洞 12.82 1.59 S6 四边形孔洞 20.26 2.53  下载: 导出CSV
下载: 导出CSV
表 2 雷达检测数据修正与统计
Table 2. Corrected calculation and statistics of the cavity areas in wood columns by TRU
序号 残损类型 第1次修正后孔洞百分比/% 第2次修正后孔洞百分比/% 第3次修正后孔洞百分比/% S3 圆形孔洞 30.21 6.85 13.26 S4 圆形孔洞 26.51 6.39 12.49 S5 三角形孔洞 38.03 8.70 15.30 S6 四边形孔洞 37.71 8.75 15.29
下载: 导出CSV
-
[1] RAZAFINDRATSIMA S, SBARTAI Z M, FRANCOIS D. Permittivity measurement of wood material over a wide range of moisture content [J]. Wood Sci Technol, 2017, 51(6): 1421 − 1431. [2] MAI T C, RAZAFINDRATSIMA S, SBARTAI Z M, et al. Non-destructive evaluation of moisture content of wood material at GPR frequency [J]. Constr Build Mater, 2015, 77: 213 − 217. [3] 吕静霞. 基于雷达波的木材内部缺陷检测方法研究[D]. 北京: 北京林业大学, 2015. LÜ Jingxia. Research on Wood Internal Defects Detection Method Based on the Radar Wave[D]. Beijing: Beijing Forestry University, 2015. [4] HANS G, REDMAN D, LEBLON B, et al. Determination of log moisture content using early-time GPR signal [J]. Wood Mater Sci Eng, 2015, 10(1): 112 − 129. [5] LORENZO C H, PÉREZ G M de la V, NOVO A, et al. Forestry applications of ground-penetrating radar [J]. For Syst, 2010, 19(1): 5 − 17. [6] 邸向辉, 王立海. 探地雷达(GPR)在木材无损检测应用中的可行性探讨[J]. 无损检测, 2013, 35(11): 51 − 54. DI Xianghui, WANG Lihai. Study on the feasibility about GPR applied on wood nondestructive testing [J]. Nondestr Test, 2013, 35(11): 51 − 54. [7] JEZOVA J, MERTENS L, LAMBOT S. Ground-penetrating radar for observing tree trunks and other cylindrical objects [J]. Constr Build Mater, 2016, 123: 214 − 225. [8] WU Xi, LI Guanghui, JIAO Zhi, et al. Reliability of acoustic tomography and ground-penetrating radar for tree decay detection[J]. Appl Plant Sci, 2018, 6(10): e1187. doi: 10.1002/aps3.1187. [9] DEVARU D, HALABE U B, GOPALAKRISHNAN B, et al. Algorithm for detecting defects in wooden logs using ground penetrating radar [J]. Proc SPIE- Int Socr Opt Eng, 2005, 5999: 110 − 121. [10] BUTNOR J R, PRUYN M L, SHAW D C, et al. Detecting defects in conifers with ground penetrating radar: applications and challenges [J]. For Pathol, 2009, 39(5): 309 − 322. [11] 李伟林, 文剑, 肖中亮, 等. 颐和园古树内部异常的雷达波检测[J]. 福建农林大学学报(自然科学版), 2017, 46(6): 665 − 670. LI Weilin, WEN Jian, XIAO Zhongliang, et al. Detection of internal anomaly for trees in the Summer Palace based on radar [J]. J Fujian Agric For Univ Nat Sci Ed, 2017, 46(6): 665 − 670. [12] 肖夏阳, 文剑, 肖中亮, 等. 基于雷达波的树木躯干内部缺陷探测识别[J]. 林业科学, 2018, 54(5): 127 − 134. XIAO Xiayang, WEN Jian, XIAO Zhongliang, et al. Detection and recognition of tree trunk internal structure based on radar [J]. Sci Silv Sin, 2018, 54(5): 127 − 134. [13] 陈勇平, 高甜, 李德山, 等. 马尾松木材内部空洞的雷达检测与定量评估[J]. 林业科学, 2017, 53(10): 139 − 145. CHEN Yongping, GAO Tian, LI Deshan, et al. Detection and quantitative evaluation of internal cavity of Pinus massoniana wood by radar testing technology [J]. Sci Silv Sin, 2017, 53(10): 139 − 145. [14] 文剑, 李伟林, 肖中亮, 等. 活立木内部缺陷雷达波检测研究[J]. 农业机械学报, 2017, 48(10): 185 − 193. WEN Jian, LI Weilin, XIAO Zhongliang, et al. Radar wave detection of standing treesinternal defect [J]. Trans Chin Soc Agric Mach, 2017, 48(10): 185 − 193. [15] 郭秀军, 王淼, 张刚, 等. 高频电磁波传播速度在水及淤积砂土中影响因素实验研究[J]. 地球物理学进展, 2010, 25(5): 1820 − 1824. GUO Xiujun, WANG Miao, ZHANG Gang, et al. Research on influence factors of GPR wave velocity in water and sandy soil in laboratory [J]. Prog Geophys, 2010, 25(5): 1820 − 1824. [16] 吴晔. 木材介电常数的研究[J]. 安徽农学院学报, 1989, 16(1): 59 − 74. WU Ye. Study on the dielectric constant of wood [J]. J Anhui Agric Univ, 1989, 16(1): 59 − 74. [17] 季亚春, 季亚坤. 木材介电常数的测量[J]. 黑龙江电子技术, 1997(5): 24 − 28. JI Yachun, JI Yakun. Measurement of wood dielectric constant [J]. Heilongjiang Electron Tech, 1997(5): 24 − 28. -

-
链接本文:
https://zlxb.zafu.edu.cn/article/doi/10.11833/j.issn.2095-0756.20190709
 点击查看大图
点击查看大图
计量
- 文章访问数: 2064
- HTML全文浏览量: 629
- PDF下载量: 33
- 被引次数: 0



