





-
楠木是一类珍贵用材树种,包括樟科Lauraceae润楠属Machilus和楠属Phoebe植物及一些近缘类群,部分种类为国家二级重点保护野生植物。长期的人为破坏[1],以及其自身天然更新不良等因素,中国楠木数量锐减。楠木零星散生于各类天然林,少有成片分布。天然林保护是在一定的生命周期和范畴内,为使资源继续顺应生态循环生长,同时能够保持资源价值不断提高的情况下,对各种森林资源进行持续的利用[2]。种间联结是植物群落重要的数量和结构特征之一,群落的种间关系能反映群落的稳定性。种间关系一般被划分为正联结、负联结和无关联。正联结是由于物种选择相同的生境或者2种物种相互依赖形成的,负联结是由于2种物种存在资源上的竞争或者根系中存在物理或化学因素的相互影响而形成的[3]。随着演替,森林群落结构趋于复杂和稳定[4],群落内物种种间关系越来越复杂,生态位分化越合理,种间关系趋于正联结[5]。但森林群落的演替并不是一味向着正关联的方向进行的[6]。对群落种间联结的相关研究展现的多是种间竞争的结果或群落的现状[7],关于群落种间联结的动态报道较少。目前,有关楠木的相关研究多在生态生理特征方面,如木材、育苗、叶片特征,而关于群落结构、种间关系等的相关研究较多的针对红楠Machilus thunbergii和紫楠Phoebe sheareri群落[8-9],缺少楠木天然林群落动态特征和种间关系研究。浙江省建德市寿昌林场绿荷塘林区内存有亚洲最大的天然楠木林,其主要种类有红楠、紫楠、刨花润楠Machilus pauhoi、薄叶润楠Machilus leptophylla等,是研究天然林群落楠木发育和演替的理想场所。本研究分别以楠木作为优势树种和伴生树种的天然林样地作为对象,对调查样地内的楠木在不同林分条件下的乔木层、灌木层、草本层种间联结性进行了定量分析,揭示它们在天然林群落发育和演替过程中种间关联性动态变化规律,为楠木天然林的科学管理提供理论依据。
-
研究区位于浙江省建德市寿昌林场绿荷塘林区内(29°25′45″~29°27′01″N,119°08′45″~119°11′0″E),属千里岗山脉,地貌为低山丘陵,坡度为25°~45°,海拔200~700 m,顶峰海拔804 m。该林区面积约450 hm2,分布约370 hm2的天然常绿阔叶林,有木本植物30科300余种。该地区属亚热带季风气候,温暖湿润,四季分明,雨量充足。年平均气温为17.6 ℃,极端最低温−13.9 ℃,极端最高温42.2 ℃。年降水量1 700 mm,集中在4−6月,平均相对湿度为82%,年日照时数为1 762 h,全年无霜期265 d。土壤以黄壤为主,局部为侵蚀性红壤。土层厚度30~80 cm,呈酸性。
如图1所示:在以楠木为优势种的林内建立100 m×100 m样地(A),并在相距1.35 km生境相似的地区,在以楠木为伴生种的林内建立样地(B),由于场地限制,样地面积为100 m×40 m。2013年对楠木优势林(样地A)进行了1次割灌处理,对非目标树种的灌木进行伐除。对楠木伴生林样地B未采取任何经营管理措施。
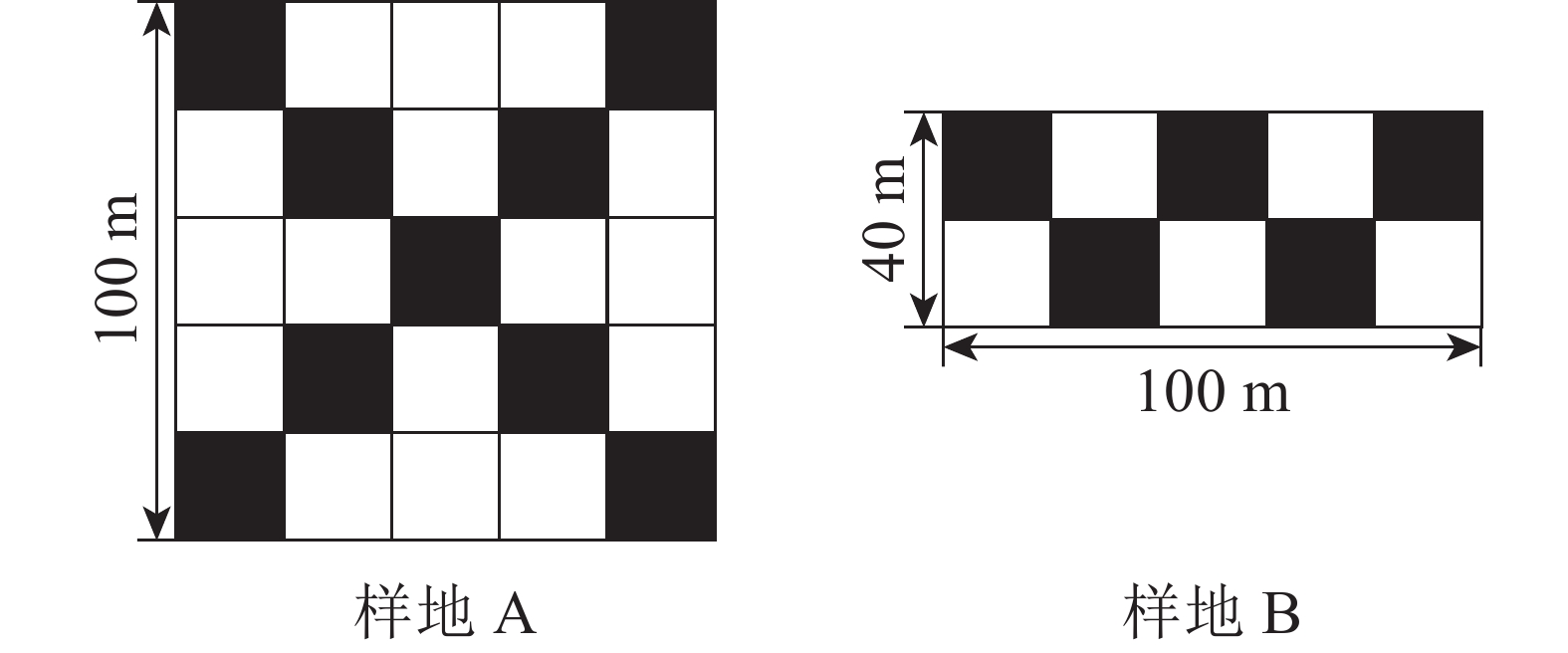
图 1 样地设置
Figure 1. Sample plots for shrub layer survey
参照中国林业科学研究院多功能森林经营样地监测方法开展样地植被调查工作。将每个样地划分为20 m×20 m的乔木样方,标定所有样方内胸径≥5.0 cm的乔木。每个样地按照图1的方法在涂黑的乔木样方右下角设置1个5 m×5 m 的灌木层小样方,记录胸径<5.0 cm,树高≥1.5 m 的木本植物,记录树种名称、胸径、树高。在每个灌木层小样方沿对角线设置3个1 m×1 m的草本层小样方,对树高<1.5 m的幼苗和草本植物进行调查,调查指标包括植物种类、盖度、丛(株)数、平均高度等。为了方便计算,将楠木幼树划分至灌木层,幼苗划分至草本层。
利用2015和2019年的2次群落调查数据,分析群落乔木层、灌木层、草本层主要物种的种间关系。
-
采用SCHLUTER[10]提出的方差比率法(
$ V_{\rm{R}} $ )检验群落物种间的总体关联性。$$ {V}_{{\rm{R}}}={S}_{{\rm{w}}}^{2}/{\delta }_{{\rm{w}}}^{2}\text{。} $$ (1) 式(1)中:VR为方差比率;总样方物种数方差
$S_{\rm{w}}^2 = \dfrac{1}{N}\displaystyle\sum\limits_{j = 1}^N {{{({T_j} - t)}^2}}$ ,$ t=\left({T}_{1}+{T}_{2}+{ \cdots +T}_{N}\right)/N $ ,$ N $ 为总样方数,t为样方物种数平均值,$ {T}_{j} $ 为第$ j $ 个样方物种数;总物种频度方差${\delta }_{{\rm{w}}}^{2}=\displaystyle\sum\limits_{i=1}^{S}{P}_{i}(1-{P}_{i})$ ,$ S $ 为总的物种数,$ {P}_{i} $ 为第$ i $ 个物种出现的样方数($ {n}_{i} $ )占总样方数($ N $ )的比例,$ {P}_{i}={n}_{i}/N $ ,$ {n}_{i} $ 为物种$ i $ 出现的样方数。在独立零假设下,VR期望值为1,表示群落种间整体无关联,VR>1表示整体正联结,VR<1表示整体负联结。采用统计量$ \omega $ ($ \omega =V_{\rm{R}} N $ )检验$ V_{\rm{R}} $ 偏离1的显著性,当${\textit{χ}}_{0.95}^{2}\left(N\right) \text{<} \omega \text{<}$ $ {\textit{χ}}_{0.05}^{2}\left(N\right) $ 时,认为整体关联不显著[11]。 -
根据物种在样方内是否出现的二元数据矩阵作为原始数据,以χ2检验为基础,结合联结系数(AC)和共同出现百分率(PC)对群落进行种间联结分析。
$ a $ 为2个物种都出现的样方数,$ b $ 、$ c $ 为只出现1个物种的样方数,$ d $ 为2个物种都不出现的样方数,在计算χ2及联结系数时,为避免$ a $ 、$ c $ 、$ d $ 为0,导致χ2及联结系数无法比较关联程度的情况,把$ a $ 、$ c $ 、$ d $ 为0的数值都加权为1[12]。χ2统计量能比较客观准确地判断种对联结的显著性,而共同出现百分率和联结系数却能体现出χ2检验不显著种对的联结性及其大小,同时共同出现百分率还能较准确地反映物种间正联结性的强弱[13]。 -
采用YATES连续矫正公式[14]:
$$ {\textit{χ} }^{2}=\frac{({\left|ad-bc\right|-0.5n)}^{2}n}{(a+b)(a+c)(b+d)(c+d)}\text{。} $$ (2) 若χ2
$ {\text{<}} 3.841(P {\text{>}} 0.05) $ 表示种对间无联结性,2种间基本独立;若3.841≤χ2≤6.635(0.01≤P≤0.05) ,表示种对间联结性显著;若χ2$ {\text{>}} 6.635(P{\text{<}} 0.01) $ ,表示种对间联结性极显著。 -
$${\text{若}}ad {\text{≥}} bc,\;{\text{则}}{A_{\rm{C}}} = \frac{{ad - bc}}{{(a + d)(b + d)}}\text{;}$$ (3) $${\text{若}}bc {\text{>}} ad{\text{且}}d {\text{≥}} a,\;{\text{则}}{A_{\rm{C}}} = \frac{{ad - bc}}{{(a + b)(a + c)}}\text{;}$$ (4) $${\text{若}}bc {\text{>}} ad{\text{且}}d {\text{<}} a,\;{\text{则}}{A_{\rm{C}}} = \frac{{ad - bc}}{{(c + d)(b + d)}}\text{。}$$ (5) 式(3)~(5)中:AC为联结系数,其值域为[−1, 1]。AC越接近1,物种间正关联越强;越接近−1;物种间负关联越强;为AC=0,则表明物种间完全独立。
-
$$ {P}_{{\rm{C}}}=\frac{a}{a+b+c}\text{。} $$ (6) 式(6)中:PC为共同出现百分率,表示物种间的联结程度[15],可以避免联结系数受d影响太大而造成偏差。其值域为[0, 1],PC=0时,表明种对无关联;PC=1时,表明种对关联程度最紧密。
-
2015和2019年,楠木优势林样地的乔木层均记录到18个种,隶属8科15属,无物种退出或进入;2015年灌木层记录到7个种,隶属5科7属,2019年记录到11个种,隶属8科10属,2个物种退出灌木层,6个物种进入灌木层;2015年草本层记录到12个种,隶属9科11属,2019年记录到5个种,隶属4科5属,7个物种退出草本层。
2015年楠木伴生林样地的乔木层记录到27个种,隶属18科23属,2019年记录到25个种,隶属16科21属,2个物种退出乔木层;2015年灌木层记录到25个种,隶属17科24属,2019年记录到14个种,隶属为7科12属,17个物种退出灌木层,6个物种进入灌木层;2015年草本层记录到6个种,隶属6科6属,2019年记录到5个种,隶属4科4属,3个物种退出草本层,2个物种进入草本层。
-
采用重要值[16]筛选群落乔木层的主要物种,只要某物种在2次群落调查中出现过1次,重要值≥1.0%,则把它认作群落中的主要物种。由表1可见:样地A有11个主要物种,2015年楠木个体占总个体数的84.76%,2019年占85.49%。样地B有16个主要物种,2015年楠木个体数占15.61%,2019年占16.10%。
表 1 2015和2019年绿荷塘林区A、B样地主要乔木物种及其重要值
Table 1. Dominant species and importance value of the tree layer of plot A and plot B in 2015 and 2019
楠木优势林(样地A) 楠木伴生林(样地B) 编号 物种 2015年 2019年 2015年 2019年 重要值/
%个体密度/
(株·m−2)重要值/
%个体密度/
(株·m−2)重要值/
%个体密度/
(株·m−2)重要值/
%个体密度/
(株·m−2)1 刨花润楠 Machilus pauhoi 46.98 162 47.93 161 2 紫楠 Phoebe sheareri 22.86 104 22.68 98 1.69 8 1.75 8 3 秀丽锥 Castanopsis jucunda 8.64 24 8.12 22 2.92 23 2.92 20 4 甜槠 Castanopsis eyrei 3.59 9 3.48 8 16.64 145 17.14 143 5 青冈 Cyclobalanopsis glauca 3.12 8 3.19 8 16.67 183 16.93 170 6 红楠 Machilus thunbergii 2.38 5 2.43 5 5.83 50 6.05 50 7 黄檀 Dalbergia hupeana 2.06 4 1.59 3 8 薄叶润楠 Machilus leptophylla 2.65 7 2.70 7 8.51 90 9.05 85 9 樟树 Cinnamomum camphora 1.45 2 1.47 2 10 栾树 Koelreuteria paniculata 1.14 2 1.17 2 11 石栎 Lithocarpus glaber 1.03 1 1.05 1 16.45 165 16.18 148 12 赤杨叶 Alniphyllum fortunei 5.21 45 4.56 38 13 花榈木 Ormosia henryi 1.01 5 1.05 5 14 冬青 Ilex chinensis 1.02 5 1.06 5 15 杉木 Cunninghamia lanceolata 9.79 165 10.13 160 16 厚皮香 Ternstroemia gymnanthera 3.08 35 3.23 35 17 苦槠 Castanopsis sclerophylla 1.90 8 1.98 8 18 八角枫 Alangium chinense 1.00 5 1.03 5 19 老鼠矢 Symplocos stellaris 0.99 5 1.02 5 20 檫木 Sassafras tzumu 1.22 10 0.70 3 -
由表2可见:样地A,2015年灌木层有7个树种,楠木个体数量占比16.13%,2019年有11个树种,楠木个体数量占比7.14%;样地B,2015年灌木层有25个物种,楠木个体数量占比18.35%,2019年有14个物种,楠木个体数量占比0.91%。
表 2 2015和2019年绿荷塘林区A、B样地灌木层物种及其重要值
Table 2. Species and importance value of the shrub layer of plot A and plot B in 2015 and 2019
编号 物种 重要值/% 编号 物种 重要值/% 样地A 样地B 样地A 样地B 2015年 2019年 2015年 2019年 2015年 2019年 2015年 2019年 1 菝葜 Smilax china 1.45 19 毛花连蕊茶 Camellia fraterna 15.04 2 薄叶润楠 Machilus leptophylla 10.90 20 刨花润楠 Machilus pauhoi 7.18 7.02 3 茶 Camellia sinensis 1.61 10.84 21 青冈 Cyclobalanopsis glauca 13.29 1.31 27.80 4 赤楠 Syzygium buxifolium 4.65 22 山矾 Symplocos sumuntia 7.03 2.44 5 赤杨叶 Alniphyllum fortunei 2.75 23 杉木 Cunninghamia lanceolata 4.16 6 豆腐柴 Premna microphylla 2.47 24 石斑木 Rhaphiolepis indica 1.36 7 杜茎山 Maesa japonica 6.78 25 石栎 Lithocarpus glaber 2.89 13.33 8 格药柃 Eurya muricata 25.59 1.87 26 甜槠 Castanopsis eyrei 6.80 9 枸杞 Lycium chinense 1.66 27 乌药 Lindera aggregata 10.19 3.05 10 红楠 Machilus thunbergii 4.50 28 秀丽锥Castanopsis jucunda 29.23 4.12 5.50 7.37 11 厚皮香 Ternstroemia gymnanthera 6.27 3.40 29 野桐 Mallotus japonicus 3.65 0.89 12 花榈木 Ormosia henryi 1.06 30 油茶 Camellia oleifera 25.60 10.05 5.50 13 黄檀 Dalbergia hupeana 6.11 31 油桐 Vernicia fordii 0.89 14 檵木 Loropetalum chinense 6.11 7.24 1.61 32 浙江楠 Phoebe chekiangensis 13.69 4.05 15 荚蒾 Viburnum dilatatum 0.89 33 栀子 Gardenia jasminoides 4.12 3.03 16 矩形叶鼠刺 Itea oblonga 6.51 6.86 3.70 34 朱砂根 Ardisia crenata 4.38 17 老鸦糊 Callicarpa giraldii 1.24 35 紫楠 Phoebe sheareri 12.47 15.25 18 毛冬青 Ilex pubescens 1.19 -
由表3可见:样地A,2015年草本层有12个物种,楠木个体数量占比1.23%;2019年有5个物种,楠木占比29.58%;样地B,2015年草本层有6个物种,楠木个体数量占比21.28%;2019年有5个物种,楠木占比5.54%。
表 3 2015和2019年绿荷塘林区A、B样地草本层物种及其重要值
Table 3. Species and importance value of the herb layer of plot A and plot B in 2015 and 2019
编号 物种 重要值/% 编号 物种 重要值/% 样地A 样地B 样地A 样地B 2015年 2019年 2015年 2019年 2015年 2019年 2015年 2019年 1 白结香 Edgeworthia albiflora 5.31 9 鳞毛蕨 Dryopteris sp. 22.11 2 薄叶润楠 Machilus leptophylla 26.96 4.50 10 蔓赤车 Pellionia scabra 3.95 3 狗脊蕨 Woodwardia unigemmata 25.84 11 刨花润楠 Machilus pauhoi 46.04 12.54 4 寒莓 Rubus buergeri 0.80 1.37 31.22 12 三穗薹草Carex tristachya 5.05 16.97 14.26 5 红楠 Machilus thunbergii 1.03 5.92 13 细柄薯蓣Dioscorea tenuipes 1.09 6 蕨 Pteridium aquilinum var. latiusculum 5.38 14.77 2.60 35.31 14 鱼腥草Houttuynia cordata 0.95 7 紫萁 Osmunda japonica 10.52 15 苎麻Boehmeria nivea 16.95 15.42 8 里白 Hicriopteris glauca 5.51 16 紫楠Phoebe sheareri 2.92 22.02 -
根据表4可知:2015和2019年,楠木优势林样地乔木层种间总体关联性呈显著正联结;灌木层2015和2019年种间总体均呈不显著负联结;草本层2015年呈不显著正联结,2019年呈显著负联结,种间关系由正转负。说明乔木层物种结构相对稳定,灌木层种间关系松散,草本层结构遭到破坏。楠木伴生林样地乔木层2015年种间总体关联性呈不显著正联结,2019年呈不显著负联结;灌木层2015年种间呈不显著正联结,2019年呈不显著负联结;草本层2015呈不显著负联结,2019年呈显著负联结。乔木层和灌木层种间关系由正转负,说明群落稳定性遭到破坏;草本层种间关系松散,结构更加不稳定。
表 4 绿荷塘林区A、B样地2015和2019年种间总体关联性
Table 4. Overall associations of plot A and plot B in 2015 and 2019
样地 层次 年份 方差比率 检验统计量 临界值 结果 楠木优势林(样地A) 乔木层 2015 1.69 42.30 [14.61, 37.65] 显著正联结 2019 1.69 42.25 [14.61, 37.65] 显著正联结 灌木层 2015 0.45 4.05 [3.33, 16.92] 不显著负联结 2019 0.78 7.02 [3.33, 16.92] 不显著负联结 草本层 2015 1.30 35.18 [16.15, 40.11] 不显著正联结 2019 0.22 5.98 [16.15, 40.11] 显著负联结 楠木伴生林(样地B) 乔木层 2015 1.12 11.16 [3.94, 18.31] 不显著正联结 2019 0.99 9.88 [3.94, 18.31] 不显著负联结 灌木层 2015 1.22 6.09 [1.15, 11.07] 不显著正联结 2019 0.41 2.05 [1.15, 11.07] 不显著负联结 草本层 2015 0.80 11.99 [7.26, 25.00] 不显著负联结 2019 0.24 3.57 [7.26, 25.00] 显著负联结 -
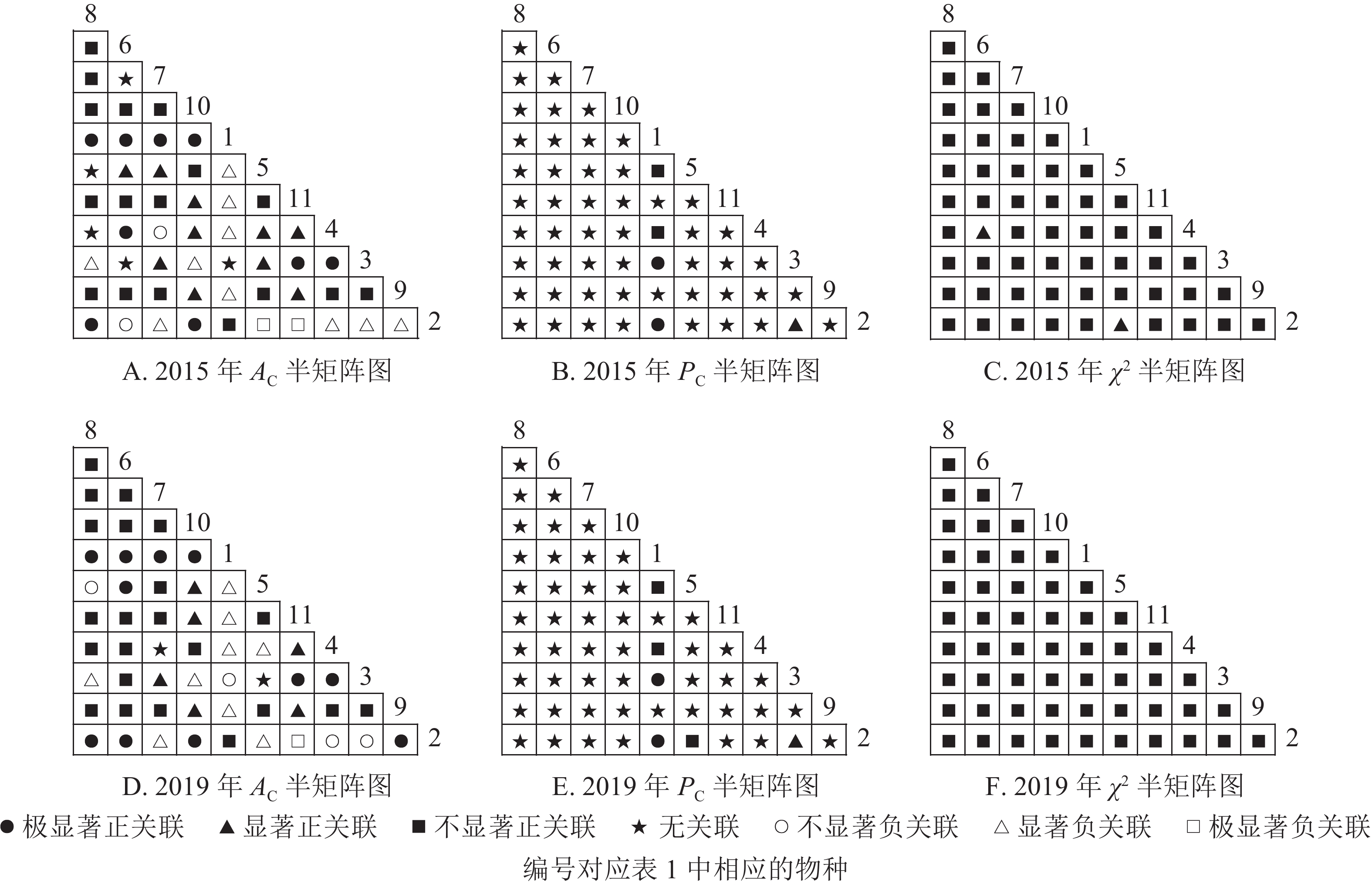
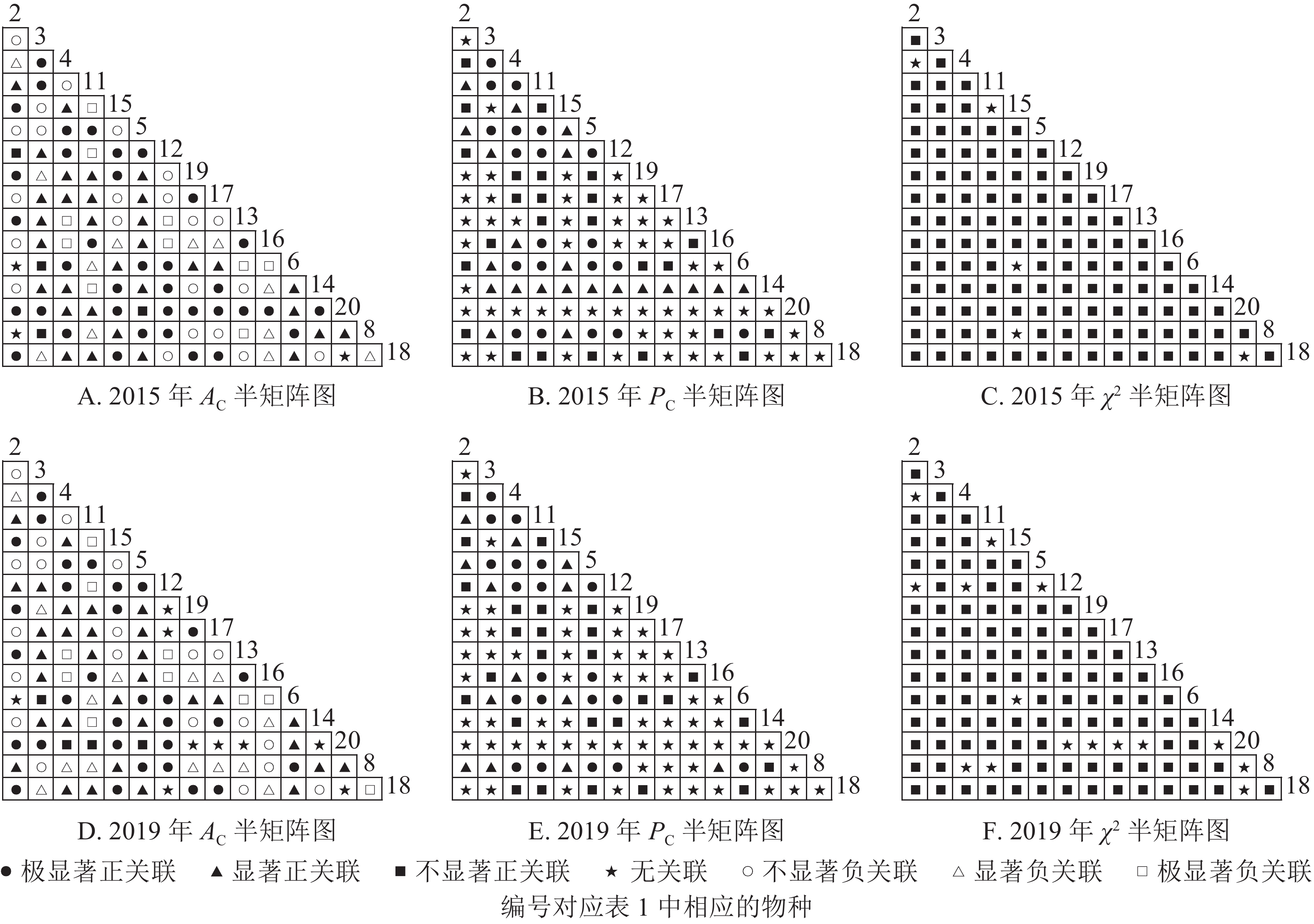
从图2可知:2015和2019年无主要树种退出乔木层,11个主要树种组成55个种对。χ2检验结果显示:2015年55个种对均表现为正关联,2019年55个种对均表现为不显著正关联。联结系数(
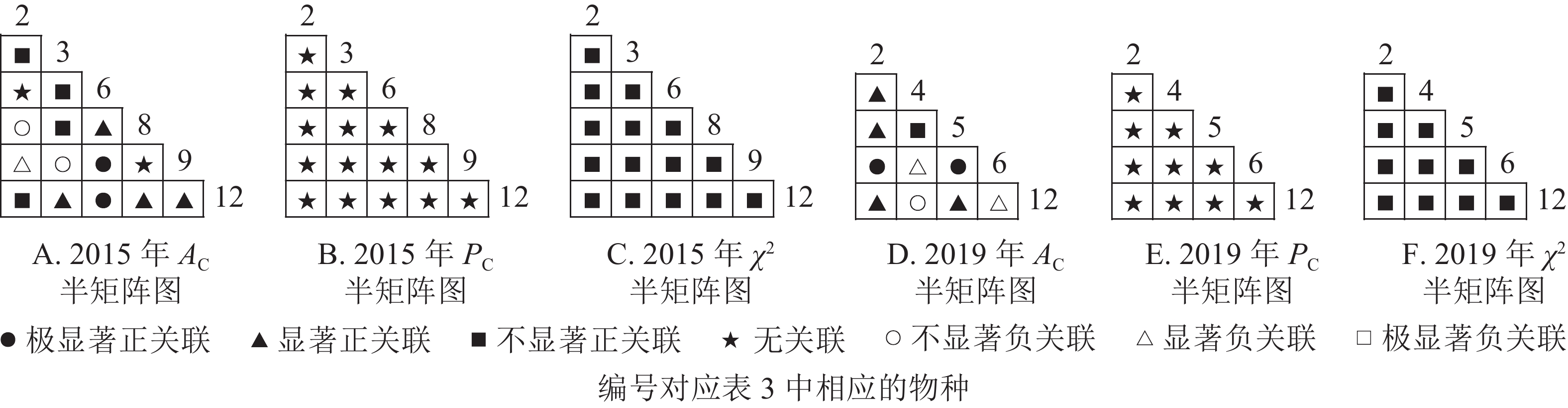
$ A_{\rm{C}} $ )结果显示:正联结种对减少8对,负联结种对减少7对。共同出现百分率($ P_{\rm{C}} $ )结果显示:2015和2019年,均有2个种对表现为极显著关联,1个种对表现为显著关联。可见,2015和2019年,乔木层中刨花润楠与其他树种负关联增强;红楠与甜槠的正联结减弱,与黄檀、紫楠正联结增强;薄叶润楠与乔木层其他树种的关联性相对稳定;紫楠与其他树种的正负关联比增大,与红楠、青冈、樟树正关联增强。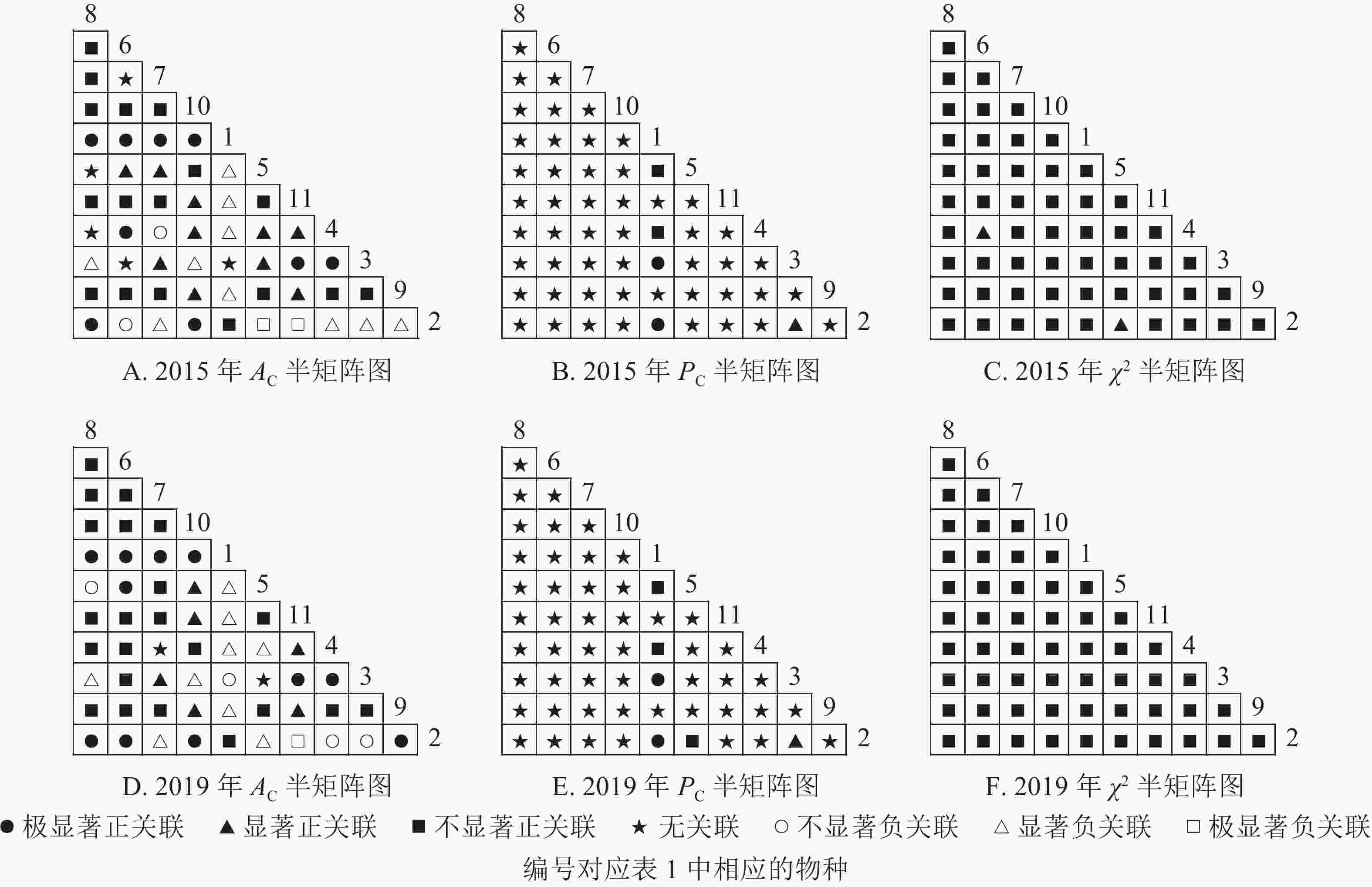
图 2 楠木优势林样地乔木层主要树种种间关联半矩阵图
Figure 2. Semi-matrix of inter-specific correlations of the tree layer of plot A
图3显示:灌木层2015年由7个主要树种组成21个种对,2019年由11个主要树种组成55个种对。χ2检验结果显示,2015年21个种对均呈不显著正关联,2019年55个种对均呈不显著正关联。联结系数结果中,2015年,11个种对呈显著正关联;1个种对呈显著负联结。2019年,8个种对呈极显著正关联,16个种对呈显著正关联;1个种对呈显著负关联。共同出现百分率结果显示,2015和2019年,均有1个种对呈不显著关联,其余种对呈无关联。以上结果表明:2015和2019年,灌木层的楠木主要为刨花润楠和紫楠。刨花润楠与其他灌木树种均表现为正关联,其中与檵木、秀丽锥的正联结加强;紫楠与其他灌木树种正关联性加强,但与檵木的联结性由正转负。
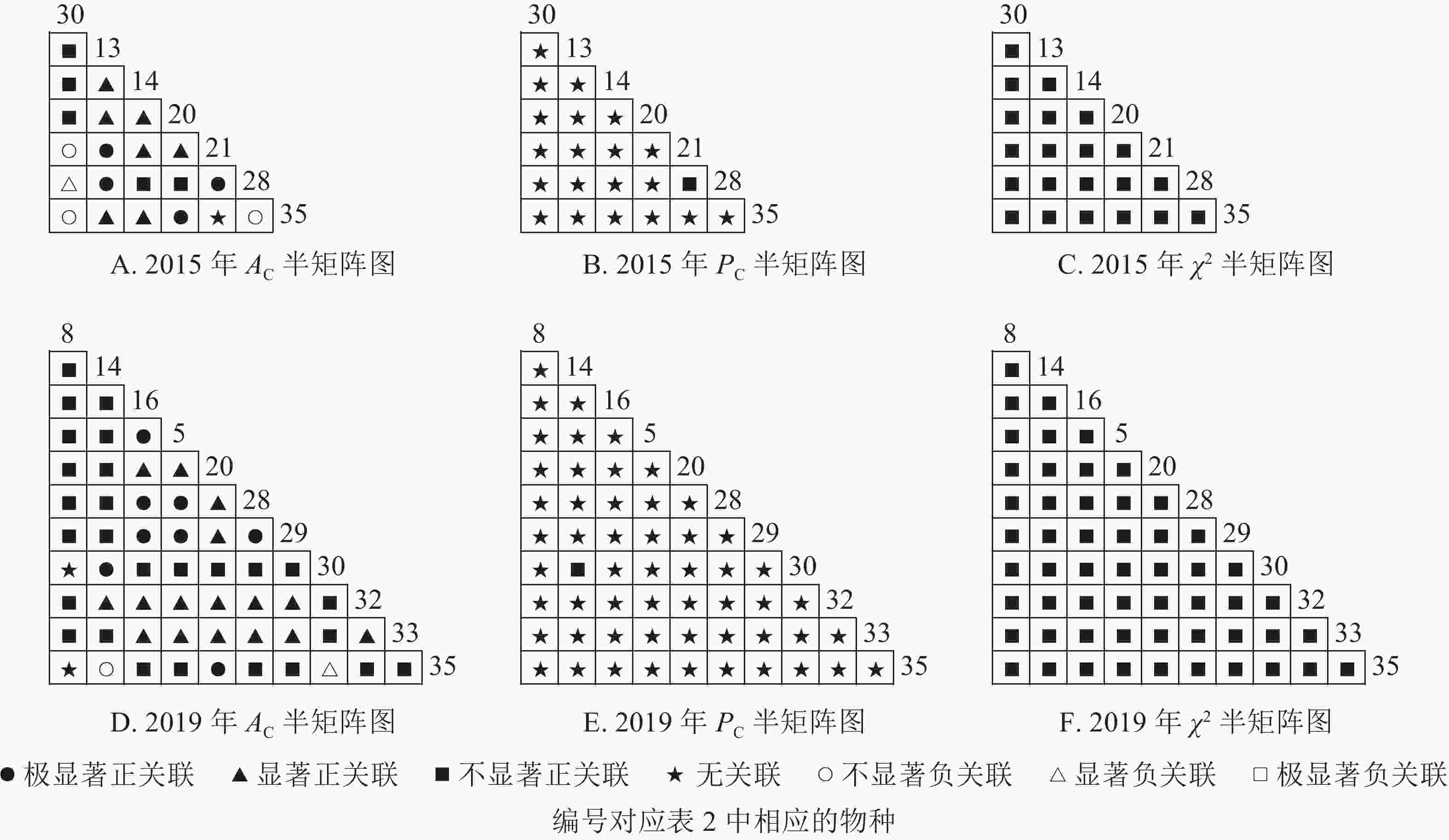
图 3 楠木优势林样地灌木层主要树种种间关联半矩阵图
Figure 3. Semi-matrix of inter-specific correlations of the shrub layer of plot A
从图4可知:草本层中,2015年由12个主要树种组成66个种对,2019年由5个主要树种组成10个种对。χ2检验结果显示:2015年3个种对呈显著正关联,2019年10个种对均呈不显著正关联。联结系数结果中:2015年,13个种对呈显著正关联,18个种对呈显著负关联;2019年,5个种对呈显著正关联,3个呈显著负联结。共同出现百分率结果显示:2015年,2个种对呈极显著关联,1个种对呈显著关联;2019年,10个种对均呈不显著正关联。以上结果表明:2015−2019年,草本层的物种变化较大,2015年主要的楠木树种有红楠、刨花润楠、紫楠,2019年红楠退出草本层。刨花润楠与大部分草本层物种呈负联结,与苎麻、紫楠的负联结增强。紫楠与刨花润楠的负联结加强,与蕨和苎麻的正联结相对稳定。
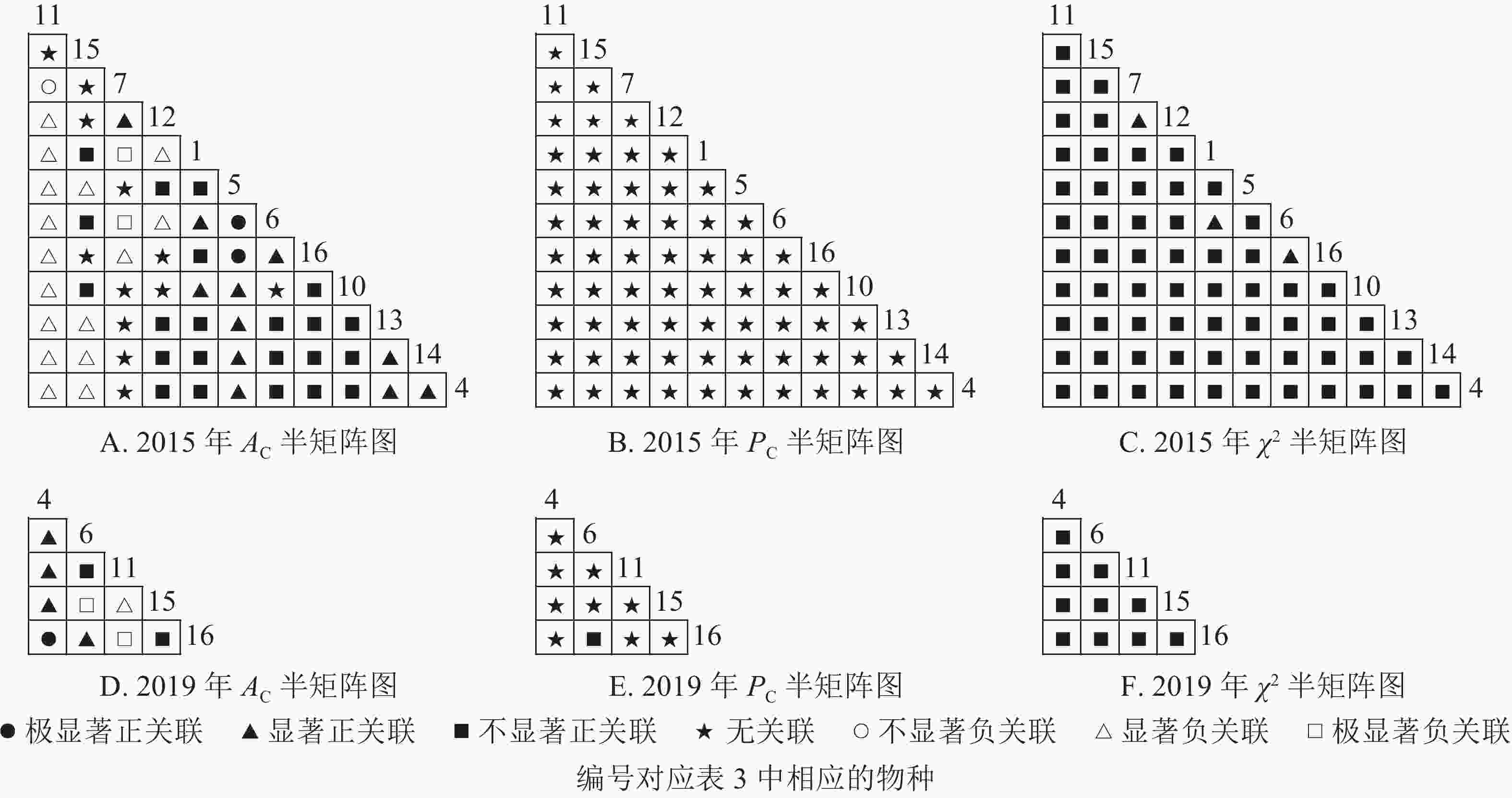
图 4 楠木优势林样地草本层主要树种种间关联半矩阵图
Figure 4. Semi-matrix of inter-specific correlations of the herb layer of plot A
-
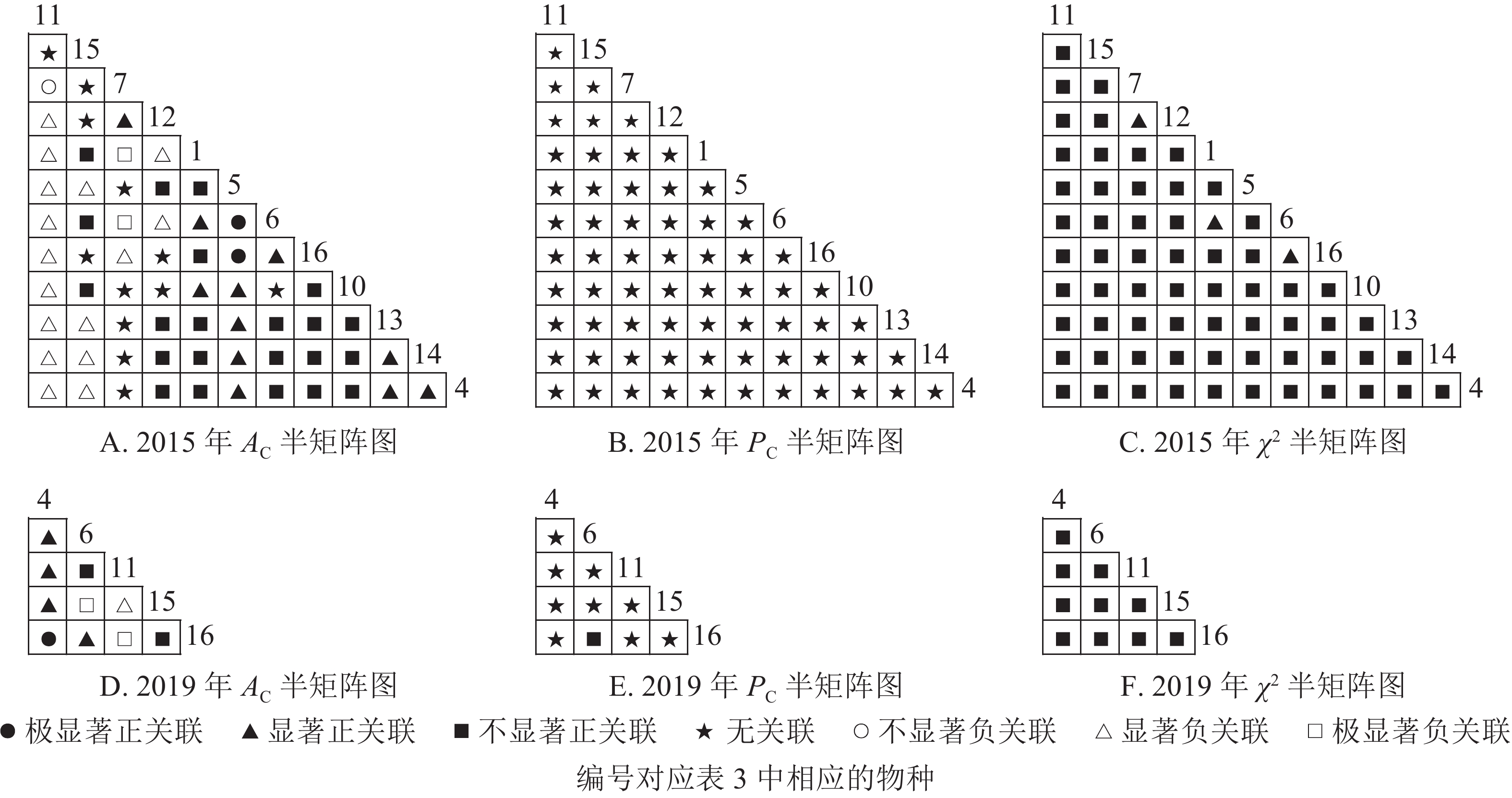
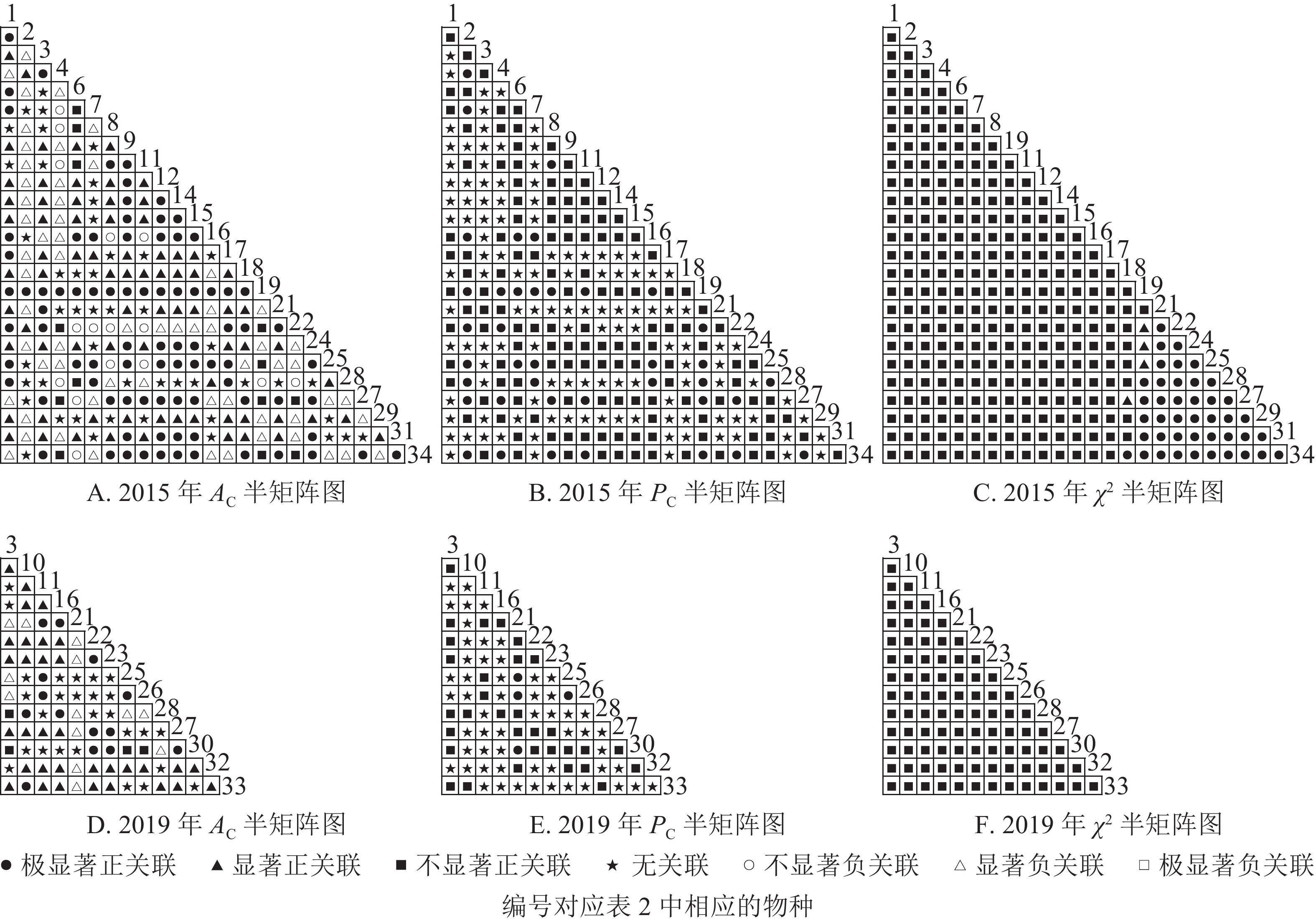
根据图5:2015和2019年,乔木层无主要树种退出,16个主要树种组成120个种对。χ2检验结果显示:2015年,115个种对呈不显著正关联,5个种对呈无关联;2019年,105个种对均呈不显著正关联,15个种对呈无关联。联结系数结果显示:正联结种对6对,负联结种对为44对。共同出现百分率结果显示:2015年,20个种对呈极显著关联,11个种对呈显著正关联;2019年,20个种对呈极显著关联,13个种对呈显著关联。可见,2015和2019年,乔木层中,红楠与其他树种之间的正负关联比大于1,且关联相对稳定;紫楠与其他树种之间的正负关联比增大;薄叶润楠与其他树种的正负关联比2015年大于1,2019年小于1,与秀丽锥、甜槠、老鼠矢、苦槠、花榈木、八角枫的负关联增强。
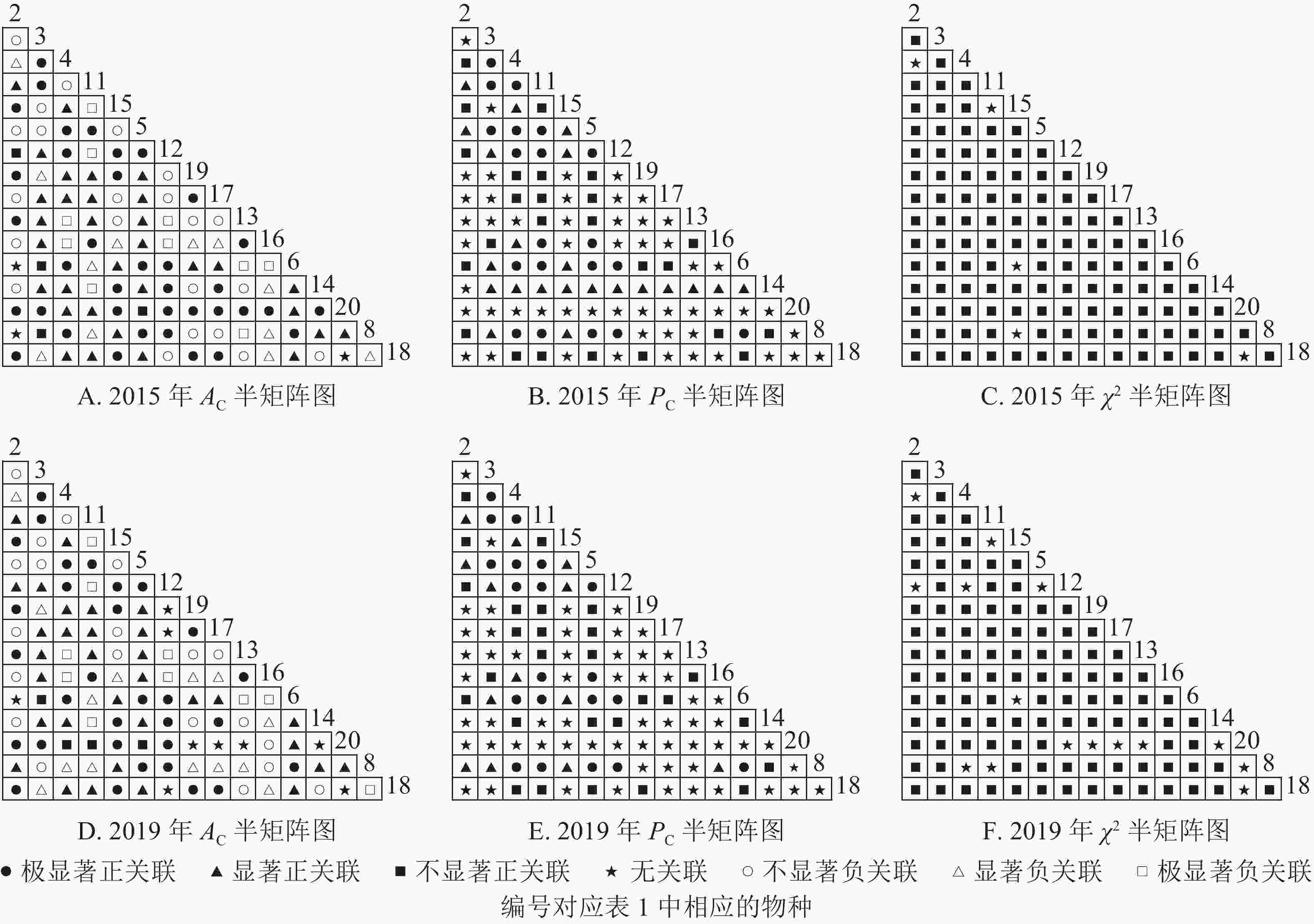
图 5 楠木伴生林样地乔木层主要树种种间关联半矩阵图
Figure 5. Semi-matrix of inter-specific correlations of the tree layer of plot B
根据图6:灌木层中,2015年由25个主要树种组成300个种对,2019年由14个主要树种组成91个种对。χ2检验结果显示:2015年46个种对呈显著正关联;2019年,105个种对均呈不显著正关联。联结系数结果显示:2015年161个种对呈显著正关联,62个种对呈显著负关联;2019年47个种对呈显著正关联,13个种对呈显著负关联。共同出现百分率结果显示:2015年,39个种对呈极显著关联;2019年,4个种对呈极显著关联。可见,2015和2019年,灌木层的树种变化较大,2015年灌木层的楠木树种主要为薄叶润楠,2019年主要为红楠。2015年,薄叶润楠与其他树种的正负关联比小于1。2019年,红楠与青冈、石栎、甜槠呈显著负关联,与其余树种呈正关联。

图 6 楠木伴生林样地灌木层主要树种种间关联半矩阵图
Figure 6. Semi-matrix of inter-specific correlations of the shrub layer of plot B
根据图7:草本层中,2015年由6个主要树种组成15个种对,2019年由5个主要树种组成10个种对。χ2检验结果显示:2015年15个种对均呈不显著正关联;2019年,10个种对均呈不显著正关联。联结系数结果中:2015年6个种对呈显著正关联,1个种对呈显著负关联;2019年8个种对呈显著正关联,2个种对呈显著负关联。共同出现百分率结果显示:2015和2019年所有种对均呈无关联。表明2015和2019年薄叶润楠幼苗与草本层其他的植物正负关联比增加;2019年红楠进入草本层,与其他植物均呈正联结。
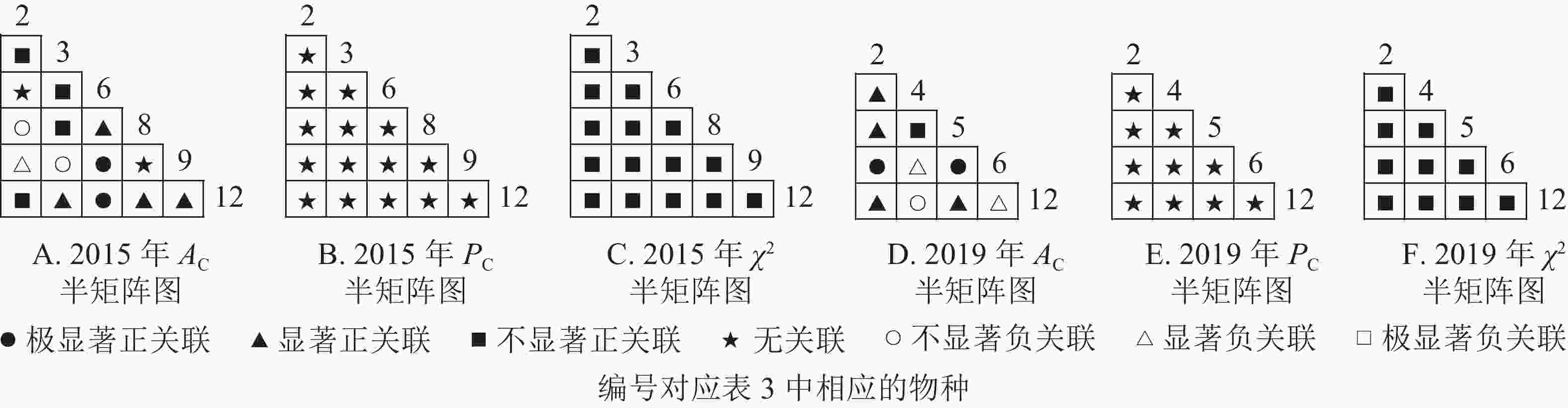
图 7 楠木伴生林样地草本层主要树种种间关联半矩阵图
Figure 7. Semi-matrix of inter-specific correlations of the herb layer of plot B
-
经总体关联性检验、χ2检验、联结系数和共同出现百分率的分析,研究区楠木优势林样地和伴生林样地的乔木层、灌木层、草本层均处于不稳定阶段。在天然楠木优势林样地中,薄叶润楠与乔木层其他树种之间的关联性相对稳定,但缺少更新层。红楠在乔木层与其他树种之间的关联性相对稳定,但在灌木层和草本层出现了更新不良的状况,这可能由于其幼苗的存活率依赖于充足的阳光所造成的[17]。紫楠在乔木层与其他树种的正关联关系加强,拥有良好的更新层,可能是由于其幼苗的光补偿点较低,对弱光的利用能力较强,在光强弱的环境下更有利于正常生长[18]。刨花润楠拥有完整的更新层,作为乔木层的优势种,与其他树种负联结增强,其幼苗能在光照强度较弱的环境下正常生长,但在草本层与其他物种之间的负联结明显,可能是由于其幼苗抗旱能力较差,与其他物种竞争土壤水分导致的[19]。
在楠木伴生林样地中,薄叶润楠作为乔木层的优势树种之一,与其他物种负联结增强,在灌木层和草本层也表现出更新不良的状况。红楠在乔木层与其他物种正联结增强,少形成负联结,在进入灌木层和草本层后也与其他物种的呈正联结,可见红楠能够和大多数物种稳定共存,具有良好的更新能力。紫楠在乔木层与其他树种正关联增强,但是缺少更新层。此前也有研究表明:在常绿阔叶林中优势树种的种内竞争比种间竞争激烈,种内竞争最激烈的优势树种与其他优势树种的种间竞争强度也是最高的[18],和其他物种在空间和资源上竞争激烈[20]。
通过对比2个样地内主要楠木树种与其他树种之间的联结性发现:在楠木优势林样地中形成负联结种对的物种,却在楠木伴生林样地中形成正联结种对。如2015年薄叶润楠与秀丽锥在天然楠木优势林呈负联结,但在天然楠木伴生林样地内呈正联结。这可能是由于群落生境的异质性和微环境的变化,使同一种植物在不同的林分具有不同的生态位,从而影响它与其他物种之间的关系[21]。
种对间的正关联主要是由于2个物种具有相似的环境需要、相似的生物学特征或相似的生态位;负联结主要是由于物种之间存在竞争,相互排斥。林分中下层的植物个体的生长和演替,除了受到土壤、水分等环境因子影响外,还要受到乔木层的影响,常绿阔叶林郁闭度较高,林下光照弱,可能会加剧林分中下层对光照的竞争,从而出现林分断层现象。林分各层次的正关联性越强,群落物种之间的互补性越强,能够更加充分地利用资源,增强群落的稳定性[22]。根据这2个群落种间联结动态变化的特点,在天然楠木林中,将楠木作为目标树种进行管理与保护时,可采用间伐、择伐等抚育方法,保留与楠木有显著正关联的树种,伐除负关联的树种,缓和种间矛盾;疏伐上层高大乔木,增加林窗,以满足林分更新层对光照的需求,保护其更新层,以形成完整的垂直结构。
Interspecific association dynamics of Nanmu natural forest in Jiande, Zhejiang Province
-
摘要:
目的 以浙江省建德市寿昌林场绿荷塘林区内楠木优势程度不同的天然林作为研究对象,进行种间关系的分析,揭示楠木在天然林群落发育和演替过程中种间关联性动态变化规律。 方法 分别在2015和2019年,对2个楠木优势程度不同的天然林样地进行调查,基于2×2联列表,通过χ2检验、联结系数、共同出现百分率,研究调查样地内楠木在不同林分条件下,乔木层、灌木层、草本层的种间联结性。 结果 2015和2019年,天然楠木优势林样地乔木层总体种间关联性均呈显著正联结,灌木层和草本层种间总体关联性趋向于负联结;天然楠木伴生林乔木层、灌木层、草本层种间总体关联性趋向于负联结。在这2种天然林中,楠木在乔木层与其他树种之间的联结相对独立,在灌木层和草本层的种间竞争较为激烈,且均有楠木在垂直方向上表现出林分断层现象。 结论 在天然林中,将楠木作为目标树种进行管理与保护时,可采用间伐、择伐等抚育方法,保留与楠木有显著正关联的树种,伐除负关联的树种,缓和种间矛盾;疏伐上层高大乔木,增加林窗,以满足林分更新层对光照的需求,保护其更新层,以形成完整的垂直结构。图7表4参22 Abstract:Objective This study aims to investigate the interspecific relationship of natural forest of Nanmu with different dominant degrees in Shouchang Forest Farm of Jiande City, Zhejiang Province, so as to reveal the dynamic change rules of the interspecific association in the process of community development and succession. Method In 2015 and 2019, two natural forest plots with different dominance of Nanmu were investigated. Based on the 2×2 contingency table, the interspecific association of tree layer, shrub layer and herb layer of Nanmu under different stand conditions was studied by χ2 test, association coefficient, and percentage of co-occurrence. Result In 2015 and 2019, the overall interspecific association of tree layer was significantly positive, while that of shrub layer and herb layer tended to be negative. In the natural stands with Nanmu as associated tree, the correlation among tree layer, shrub layer and herb layer tended to be negative. In both stands, Nanmu showed stand fault phenomenon in vertical direction. In the tree layer, the association between Nanmu and other species was relatively independent, while the interspecific competition between Nanmu and other species in shrub layer and herb layer was intense. Conclusion In natural forests, thinning, selective cutting and other tending methods can be adopted when Nanmu is taken as the target tree species for management and protection. The tree species with significant positive correlation with Nanmu are retained and the tree species with negative correlation are cut down to ease the interspecific contradiction. Thinning tall trees in the upper layer and increasing the forest gap can meet the needs of illumination for renewal layer. Protecting the renewal layer is conducive to the formation of a complete vertical structure. [Ch, 7 fig. 4 tab. 22 ref.] -
Key words:
- Nanmu /
- interspecific association /
- natural forest /
- Jiande of Zhejiang Province
-
图 2 楠木优势林样地乔木层主要树种种间关联半矩阵图
Figure 2 Semi-matrix of inter-specific correlations of the tree layer of plot A
图 3 楠木优势林样地灌木层主要树种种间关联半矩阵图
Figure 3 Semi-matrix of inter-specific correlations of the shrub layer of plot A
图 4 楠木优势林样地草本层主要树种种间关联半矩阵图
Figure 4 Semi-matrix of inter-specific correlations of the herb layer of plot A
图 5 楠木伴生林样地乔木层主要树种种间关联半矩阵图
Figure 5 Semi-matrix of inter-specific correlations of the tree layer of plot B
图 6 楠木伴生林样地灌木层主要树种种间关联半矩阵图
Figure 6 Semi-matrix of inter-specific correlations of the shrub layer of plot B
图 7 楠木伴生林样地草本层主要树种种间关联半矩阵图
Figure 7 Semi-matrix of inter-specific correlations of the herb layer of plot B
表 1 2015和2019年绿荷塘林区A、B样地主要乔木物种及其重要值
Table 1. Dominant species and importance value of the tree layer of plot A and plot B in 2015 and 2019
楠木优势林(样地A) 楠木伴生林(样地B) 编号 物种 2015年 2019年 2015年 2019年 重要值/
%个体密度/
(株·m−2)重要值/
%个体密度/
(株·m−2)重要值/
%个体密度/
(株·m−2)重要值/
%个体密度/
(株·m−2)1 刨花润楠 Machilus pauhoi 46.98 162 47.93 161 2 紫楠 Phoebe sheareri 22.86 104 22.68 98 1.69 8 1.75 8 3 秀丽锥 Castanopsis jucunda 8.64 24 8.12 22 2.92 23 2.92 20 4 甜槠 Castanopsis eyrei 3.59 9 3.48 8 16.64 145 17.14 143 5 青冈 Cyclobalanopsis glauca 3.12 8 3.19 8 16.67 183 16.93 170 6 红楠 Machilus thunbergii 2.38 5 2.43 5 5.83 50 6.05 50 7 黄檀 Dalbergia hupeana 2.06 4 1.59 3 8 薄叶润楠 Machilus leptophylla 2.65 7 2.70 7 8.51 90 9.05 85 9 樟树 Cinnamomum camphora 1.45 2 1.47 2 10 栾树 Koelreuteria paniculata 1.14 2 1.17 2 11 石栎 Lithocarpus glaber 1.03 1 1.05 1 16.45 165 16.18 148 12 赤杨叶 Alniphyllum fortunei 5.21 45 4.56 38 13 花榈木 Ormosia henryi 1.01 5 1.05 5 14 冬青 Ilex chinensis 1.02 5 1.06 5 15 杉木 Cunninghamia lanceolata 9.79 165 10.13 160 16 厚皮香 Ternstroemia gymnanthera 3.08 35 3.23 35 17 苦槠 Castanopsis sclerophylla 1.90 8 1.98 8 18 八角枫 Alangium chinense 1.00 5 1.03 5 19 老鼠矢 Symplocos stellaris 0.99 5 1.02 5 20 檫木 Sassafras tzumu 1.22 10 0.70 3  下载: 导出CSV
下载: 导出CSV
表 2 2015和2019年绿荷塘林区A、B样地灌木层物种及其重要值
Table 2. Species and importance value of the shrub layer of plot A and plot B in 2015 and 2019
编号 物种 重要值/% 编号 物种 重要值/% 样地A 样地B 样地A 样地B 2015年 2019年 2015年 2019年 2015年 2019年 2015年 2019年 1 菝葜 Smilax china 1.45 19 毛花连蕊茶 Camellia fraterna 15.04 2 薄叶润楠 Machilus leptophylla 10.90 20 刨花润楠 Machilus pauhoi 7.18 7.02 3 茶 Camellia sinensis 1.61 10.84 21 青冈 Cyclobalanopsis glauca 13.29 1.31 27.80 4 赤楠 Syzygium buxifolium 4.65 22 山矾 Symplocos sumuntia 7.03 2.44 5 赤杨叶 Alniphyllum fortunei 2.75 23 杉木 Cunninghamia lanceolata 4.16 6 豆腐柴 Premna microphylla 2.47 24 石斑木 Rhaphiolepis indica 1.36 7 杜茎山 Maesa japonica 6.78 25 石栎 Lithocarpus glaber 2.89 13.33 8 格药柃 Eurya muricata 25.59 1.87 26 甜槠 Castanopsis eyrei 6.80 9 枸杞 Lycium chinense 1.66 27 乌药 Lindera aggregata 10.19 3.05 10 红楠 Machilus thunbergii 4.50 28 秀丽锥Castanopsis jucunda 29.23 4.12 5.50 7.37 11 厚皮香 Ternstroemia gymnanthera 6.27 3.40 29 野桐 Mallotus japonicus 3.65 0.89 12 花榈木 Ormosia henryi 1.06 30 油茶 Camellia oleifera 25.60 10.05 5.50 13 黄檀 Dalbergia hupeana 6.11 31 油桐 Vernicia fordii 0.89 14 檵木 Loropetalum chinense 6.11 7.24 1.61 32 浙江楠 Phoebe chekiangensis 13.69 4.05 15 荚蒾 Viburnum dilatatum 0.89 33 栀子 Gardenia jasminoides 4.12 3.03 16 矩形叶鼠刺 Itea oblonga 6.51 6.86 3.70 34 朱砂根 Ardisia crenata 4.38 17 老鸦糊 Callicarpa giraldii 1.24 35 紫楠 Phoebe sheareri 12.47 15.25 18 毛冬青 Ilex pubescens 1.19
下载: 导出CSV
表 3 2015和2019年绿荷塘林区A、B样地草本层物种及其重要值
Table 3. Species and importance value of the herb layer of plot A and plot B in 2015 and 2019
编号 物种 重要值/% 编号 物种 重要值/% 样地A 样地B 样地A 样地B 2015年 2019年 2015年 2019年 2015年 2019年 2015年 2019年 1 白结香 Edgeworthia albiflora 5.31 9 鳞毛蕨 Dryopteris sp. 22.11 2 薄叶润楠 Machilus leptophylla 26.96 4.50 10 蔓赤车 Pellionia scabra 3.95 3 狗脊蕨 Woodwardia unigemmata 25.84 11 刨花润楠 Machilus pauhoi 46.04 12.54 4 寒莓 Rubus buergeri 0.80 1.37 31.22 12 三穗薹草Carex tristachya 5.05 16.97 14.26 5 红楠 Machilus thunbergii 1.03 5.92 13 细柄薯蓣Dioscorea tenuipes 1.09 6 蕨 Pteridium aquilinum var. latiusculum 5.38 14.77 2.60 35.31 14 鱼腥草Houttuynia cordata 0.95 7 紫萁 Osmunda japonica 10.52 15 苎麻Boehmeria nivea 16.95 15.42 8 里白 Hicriopteris glauca 5.51 16 紫楠Phoebe sheareri 2.92 22.02
下载: 导出CSV
表 4 绿荷塘林区A、B样地2015和2019年种间总体关联性
Table 4. Overall associations of plot A and plot B in 2015 and 2019
样地 层次 年份 方差比率 检验统计量 临界值 结果 楠木优势林(样地A) 乔木层 2015 1.69 42.30 [14.61, 37.65] 显著正联结 2019 1.69 42.25 [14.61, 37.65] 显著正联结 灌木层 2015 0.45 4.05 [3.33, 16.92] 不显著负联结 2019 0.78 7.02 [3.33, 16.92] 不显著负联结 草本层 2015 1.30 35.18 [16.15, 40.11] 不显著正联结 2019 0.22 5.98 [16.15, 40.11] 显著负联结 楠木伴生林(样地B) 乔木层 2015 1.12 11.16 [3.94, 18.31] 不显著正联结 2019 0.99 9.88 [3.94, 18.31] 不显著负联结 灌木层 2015 1.22 6.09 [1.15, 11.07] 不显著正联结 2019 0.41 2.05 [1.15, 11.07] 不显著负联结 草本层 2015 0.80 11.99 [7.26, 25.00] 不显著负联结 2019 0.24 3.57 [7.26, 25.00] 显著负联结
下载: 导出CSV
-
[1] VENTER O, SANDERSON E W, MAGRACH A, et al. Sixteen years of change in the global terrestrial human footprint and implications for biodiversity conservation [J]. Nat Commun, 2016, 7(1): 1 − 11. [2] 刘世荣, 马姜明, 缪宁. 中国天然林保护、生态恢复与可持续经营的理论与技术[J]. 生态学报, 2015, 35(1): 212 − 218. LIU Shirong, MA Jiangming, MIAO Ning. Achievements in natural forest protection, ecological restoration, and sustainable management in China [J]. Acta Ecol Sin, 2015, 35(1): 212 − 218. [3] ROXBURGH S H, CHESSON P. A new method for detecting species associations with spatially autocorrelated data [J]. Ecology, 1998, 79(6): 2180 − 2192. [4] 周先叶, 王伯荪, 李鸣光, 等. 广东黑石顶自然保护区森林次生演替过程中群落的种间联结性分析[J]. 植物生态学报, 2000, 24(3): 332 − 339. ZHOU Xianye, WANG Bosun, LI Mingguang, et al. An analysis of in terspecific associanons in secondary succession forest communities in Heishiding Natural Reserve Guamgdong Province [J]. Acta Phytoecol Sin, 2000, 24(3): 332 − 339. [5] 彭舜磊, 闫小婷, 齐光, 等. 伏牛山栓皮栎群落优势种群种间关联动态分析[J]. 森林与环境学报, 2018, 38(3): 326 − 333. PENG Shunlei, YAN Xiaoting, QI Guang, et al. Interspecific association dynamic changes among the dominant species of Quercus variabilis community in Funiushan National Nature Reserve [J]. J For Environ, 2018, 38(3): 326 − 333. [6] 王乃江, 张文辉, 陆元昌, 等. 陕西子午岭森林植物群落种间联结性[J]. 生态学报, 2010, 30(1): 67 − 78. WANG Naijiang, ZHANG Wenhui, LU Yuanchang, et al. Interspecific association among the plants communities in the forest at Ziwuling Area in Shaanxi Province [J]. Acta Ecol Sin, 2010, 30(1): 67 − 78. [7] 叶森土, 金超, 吴初平, 等. 浙江松阳县生态公益林群落分类排序及优势种种间关联分析[J]. 浙江农林大学学报, 2020, 37(4): 693 − 701. YE Sentu, JIN Chao, WU Chuping, et al. Classification, ordination and correlation analysis of dominant species of ecological non-commercial forests in Songyang, Zhejiang [J]. J Zhejiang A&F Univ, 2020, 37(4): 693 − 701. [8] 洪利兴, 杜国坚. 杭州市郊黄梅坞林区天然紫楠林的群落结构与生长规律研究[J]. 浙江林业科技, 1989, 9(4): 22 − 33. HONG Lixing, DU Guojian. The structure of the natural forest of Phoebe sheareri in Wangmeiwu forest district of Hangzhou suburb and it’s law of growth [J]. J Zhejiang For Sci Technol, 1989, 9(4): 22 − 33. [9] 高浩杰, 高平仕, 王国明. 舟山群岛红楠林种内和种间竞争研究[J]. 植物研究, 2017, 37(3): 440 − 446. GAO Haojie, GAO Pingshi, WANG Guoming. Intraspecific and interspecific competition of Machilus thunbergii forest in Zhoushan Islands [J]. Bull Bot Res, 2017, 37(3): 440 − 446. [10] SCHLUTER D. A variance test for detecting species associations, with some example applications [J]. Ecology, 1984, 65(3): 998 − 1005. [11] 张金屯. 数量生态学[M]. 北京: 科学出版社, 2004. [12] 张倩媚, 陈北光, 周国逸. 鼎湖山主要林型优势树种种间联结性的计算方法研究[J]. 华南农业大学学报, 2006, 27(1): 79 − 83. ZHANG Qianmei, CHEN Beiguang, ZHOU Guoyi, et al. Interspecific association of the dominant species in two typical communities in Dinghushan, South China [J]. J South China Agric Univ, 2006, 27(1): 79 − 83. [13] 郭志华, 卓正大, 陈洁, 等. 庐山常绿阔叶、落叶阔叶混交林乔木种群种间联结性研究[J]. 植物生态学报, 1997, 21(5): 3 − 41. GUO Zhihua, ZHUO Zhengda, CHEN Jie, et al. Interspecific association of trees in mixed evergreen and deciduous broadleaved forest in Lushan Mountain [J]. Acta Phytoecol Sin, 1997, 21(5): 3 − 41. [14] 王伯荪, 彭少麟. 南亚热带常绿阔叶林种间联结测定技术研究(Ⅰ)种间联结测式的探讨与修正[J]. 植物生态学与地植物学丛刊, 1985, 9(4): 274 − 285. WANG Bosun, PENG Shaolin. Studies on the measuring techniques of interspecific association of lower-subtropical evergreen-broadleaved forests (Ⅰ) the exploration and the revision on the measuring formulas of interspecific association [J]. Acta Phytoecol et Geobotan Sin, 1985, 9(4): 274 − 285. [15] PULLIAM R K R. Island biogeography: effect of geographical isolation on species composition [J]. Ecology, 1993, 74(4): 977 − 981. [16] 易敏, 赖猛, 张露, 等. 人工林刨花楠木材主要特性的径向变异及其对气象因子的响应[J]. 应用生态学报, 2018, 29(11): 3677 − 3684. YI Min, LAI Meng, ZHANG Lu, et al. Radical variation of main wood properties and its relationship to climatic factors of Machilus pauhoi plantation [J]. Chin J Appl Ecol, 2018, 29(11): 3677 − 3684. [17] 任晴, 袁位高, 吴初平, 等. 浙江省红楠生境地群落数量分类和环境解析[J]. 生态学报, 2020, 40(15): 1 − 11. REN Qing, YUAN Weigao, WU Chuping, et al. Community quantitative classification and environmental analysis of the Machilus thunbergii habitat in Zhejiang Province [J]. Acta Ecol Sin, 2020, 40(15): 1 − 11. [18] 陈模芳, 韦小丽, 张怡. 紫楠实生幼苗的光合生理特性[J]. 贵州农业科学, 2013, 41(10): 55 − 58. CHEN Mofang, WEI Xiaoli, ZHANG Yi. Study on photosynthetic physiology characteristics of young Phoebe sheareri seedling [J]. Guizhou Agric Sci, 2013, 41(10): 55 − 58. [19] 刘全勇, 卢锟, 李泽, 等. 水分胁迫对刨花润楠幼苗生长及光合特性的影响[J]. 中南林业科技大学学报, 2016, 36(9): 29 − 35. LIU Quanyong, LU Kun, LI Ze, et al. Effects of water stress on photosynthetic physiological characteristics of Machilus pauhoi seedlings [J]. J Cent South Univ For Technol, 2016, 36(9): 29 − 35. [20] CHAI Zongzheng, SUN Caili, WANG Dexiang, et al. Interspecific associations of dominant tree populations in a virgin old-growth oak forest in the Qinling Mountains, China[J]. Bot Stud, 2016, 57(1): 23. doi. 10.1186/s40529-016-0139-5. [21] 石福习, 赵成章, 高福元, 等. 祁连山北坡自然恢复林地灌木层物种多样性及种间关联动态[J]. 生态学杂志, 2012, 31(9): 2177 − 2183. SHI Fuxi, ZHAO Chengzhang, GAO Fuyuan, et al. Dynamics of species diversity and inter-specific association in shrub layer of naturally restored woodland on northern slope of Qilian Moutains [J]. Chin J Ecol, 2012, 31(9): 2177 − 2183. [22] 娄彦景, 赵魁义. 三江平原毛苔草群落近30年演替过程中的种间联结性分析[J]. 生态学杂志, 2008, 27(4): 509 − 513. LOU Yanjing, ZHAO Kuiyi. Analysis of inter-specific association of Carex lasiocarpa community in recent 30-year succession in Sanjiang Plain [J]. Chin J Ecol, 2008, 27(4): 509 − 513. -

-
链接本文:
https://zlxb.zafu.edu.cn/article/doi/10.11833/j.issn.2095-0756.20200565
 点击查看大图
点击查看大图

计量
- 文章访问数: 2489
- HTML全文浏览量: 454
- PDF下载量: 66
- 被引次数: 0








































