




 下载:
下载:
-
自工业革命以来,人类大量使用化石燃料及改变土地利用方式等过程驱动大气二氧化碳(CO2)不断升高,导致大气CO2由280 μmol·mol−1上升至目前410 μmol·mol−1,涨幅约46%。按照这一涨幅,预计21世纪末大气CO2将超过700 μmol·mol−1[1]。大气CO2等气体的持续升高将会对各个生态系统产生深远影响,尤其是全球碳的转化与平衡。陆地土壤碳库是地球表面最大的碳储存场所,比植被和大气碳库的总和还要多,其有机碳的储存量约1 200~1 600 Pg,全球0~30 cm土层的有机碳为684~724 Pg,0~1 m土层碳为1 462~1 548 Pg[2]。土壤有机碳(SOC)能够提供植物生长需要的营养元素,有效改善土壤的质量和提高土壤的蓄水保肥能力,因此,SOC是土壤质量和农艺可持续性的重要指标。根据土壤有机碳库的周转速度及对外界因素的敏感程度,可将其分为惰性有机碳库和活性有机碳库,其中活性有机碳库包括可溶性碳(DOC)、微生物量碳(MBC)、轻组有机碳(LFOC)和可矿化碳(MC)等[3]。评价SOC的指标包括碳含量、化学结构组成、分解速率、SOC稳定性等[4],其中碳含量、化学组成结构等目前已有精准的检测方法,分解速率的测定由于高空间异质性、高背景水平、土壤采样策略、采样后处理和较短的实验时间等原因,并不十分准确。SOC稳定性取决于SOC不同组分的构成及其与环境的相互作用,不同土壤中的SOC组分和来源不尽相同且变数很大。目前,有关SOC稳定性的研究逐渐增多,但大气CO2升高对SOC稳定性的影响及其机制研究相对较少。土壤有机碳稳定性指SOC的可矿化性[5],是SOC结构和特定环境的综合反应,是在当前条件下抵抗干扰和恢复原有水平的能力。它是由土壤的理化生物性质所决定的,是自然和人为因素共同作用的结果[6]。大气中CO2与SOC间的转化与平衡是相互影响的,CO2是植物光合作用的原料,大气CO2升高,植物的光合作用会相应地增强,改变植物的生长发育过程,植物地上地下部分的生物量增加,从而提高了土壤中光合有机碳的输入,使土壤成为潜在的碳汇[7]。此外,植物地下部分增加分泌的生物量也会为微生物的生长提供能量,使微生物的活动更加活跃,呼吸作用增强,可能会导致SOC含量有所下降[8]。大气CO2升高改变SOC含量的同时,还可能改变SOC的可矿化性,从而间接影响植物的生长。然而,目前的相关研究主要关注大气CO2升高对SOC储量、化学结构组成、分解速率等的影响,较少涉及其对稳定性的影响研究。因此,本研究基于现有的研究成果,利用多种有机碳稳定性指标来讨论大气CO2升高以及CO2和外源氮交互作用对SOC稳定性的影响,探讨大气CO2升高对SOC稳定性影响的主要机制及时间尺度效应,以期对相关领域的研究起到一定的推动作用。
-
生物指标主要包括SOC矿化率、微生物熵、微生物代谢熵等,其数值越低,表明SOC稳定性越高[5]。土壤有机碳矿化是SOC在微生物作用下被分解为简单无机化合物并释放CO2的过程,SOC矿化率是指单位有机碳下SOC累积矿化量的大小,可以反映SOC矿化潜力及其稳定性[9]。MARHAN等[10]通过矿化实验发现:大气CO2持续升高3 a(580 μmol·mol−1)显著提高了SOC矿化率,然而当土壤中存在小麦Triticum aestivum残渣时,残渣的分解影响到了SOC的矿化效果,使SOC矿化率降低了18.6%。刘娟等[11]通过SOC矿化培育实验发现:FACE处理(大气CO2升高200 μmol·mol−1)后的SOC矿化率为65.83 mg·kg−1·d−1,显著提高了35%。陈栋等[12]在刘娟等[11]研究的基础上,继续以FACE平台长期定位试验水稻Oryza sativa土为研究对象。通过60 d土壤有机碳矿化培养试验发现:14 a内高CO2处理下的SOC含量增加了14.6%~18.6%,SOC的矿化速率提高了33.1%~34.0%,SOC稳定性显著下降。在森林生态系统中,TANEVA等[13]以8 a内大气CO2持续升高200 μmol·mol−1的土壤为研究对象发现:土壤的呼吸速率显著高于对照组,其中表层(0~5 cm)土壤的有机碳矿化速率显著提高了31.7%~46.8%,深层(5~15 cm)土壤提高了7.0%~12.2%,SOC稳定性大大减弱。
微生物熵是指单位有机碳下微生物量碳(MBC)的大小,微生物代谢熵是指单位MBC下土壤呼吸强度,表征了微生物呼吸消耗碳的比例大小[14]。微生物熵和微生物代谢熵均反映了微生物对有机碳的利用效率,其数值升高,则SOC稳定性降低[15]。龙凤玲等[16]发现:大气CO2升高[(700±20) μmol·mol−1]2 a后,森林土壤总有机碳含量显著提高,土壤微生物熵大幅增加了51.05%~244.05%,表明SOC稳定性显著下降。赵光影等[17]发现:大气CO2升高(700 μmol·mol−1)2 a后,湿地土壤微生物熵增加了2%~5%,土壤微生物活性略微增加,SOC稳定性减弱不显著。有研究表示:微生物对大气CO2升高的反应可能取决于土壤质地[18]。BUTTERLY等[19]首次研究了大气CO2升高对3种不同土壤类型中MBC的影响,发现MBC受土壤特异类型和深度的影响较大,在浅层(0~5 cm)土壤中,石灰质土壤的微生物熵提高了10.91%,色溶胶土壤和改性土壤分别降低了40.00%和52.03%;在深层(5~15 cm)土壤中,石灰质土壤和色溶胶土壤的微生物熵分别提高了282.2%和18.7%,改性土壤中降低了41.0%。CO2升高导致石灰质土壤中的SOC稳定性显著下降,微生物在色溶胶土壤和改性土壤中并不活跃。XU等[20]也研究了大气CO2升高对不同土壤质地的SOC稳定性的影响,结果与BUTTERLY等[19]的研究类似,SOC稳定性的降低仅仅存在于石灰质土壤中,其稳定性从小到大依次为改性土壤、色溶胶土壤、石灰质土壤。
-
化学指标包括土壤活性有机碳/总有机碳比、碳氮比、烷基化程度、芳香度等,其中土壤活性有机碳包括溶解性有机碳(DOC)、易氧化有机碳(ROC)、大团聚体结合碳和颗粒有机碳(POC)等。土壤活性有机碳/总有机碳比值越大,即SOC的活性越大,微生物越容易分解,SOC稳定性也越低[5]。大部分研究表明:自然生态系统中随着大气CO2长期升高,SOC活性组分比率增加,SOC转化速度加快,有机碳活性增大,因此有机碳稳定性相应下降[21-24]。赵光影等[17]发现:大气CO2持续升高(350 μmol·mol−1)2 a后,湿地生态系统中植株根系分泌物增多,土壤微生物活跃,因此易分解有机碳的含量显著增加,其中DOC和ROC分别增加了8.30%和4.50%,DOC/SOC和ROC/SOC分别增加0.03%~5.70%和0.20%~1.40%,其SOC稳定性轻微减弱。PROCTER等[18]模拟3种草地生态系统发现:大气CO2持续升高4个生长期,SOC含量几乎都没有变化,却加快了其更新循环过程,其中黑色黏土POC含量大幅增加(上升约4倍),沙质土中POC增加50%,不稳定碳比例增加,SOC稳定性相应下降;在粉质黏土中,最古老和最顽固的矿物结合碳含量在土层垂直梯度下降了23%。农田生态系统的研究也发现:中短期大气CO2升高(<10 a)提高了表层土壤(0~5 cm)中DOC、ROC、POC、大团聚体结合碳等易分解有机碳的比率,有机碳活性增大容易被土壤中的微生物分解,有机碳的稳定性有所下降[12, 25]。关松等[26]发现:大气CO2持续升高1 a(570 μmol·mol−1),水稻土壤表层(0~15 cm)及深层(15~30 cm)有机碳含量均增加9.90%左右,表层土壤中DOC增加了42.90%,DOC/SOC增加30.40%,即表层SOC稳定性显著下降,而深层SOC稳定性几乎没有变化。种植不同品种农作物的土壤对大气CO2升高的响应可能存在区别。HU等[27]利用运行10 a的FACE平台检测了2个水稻品种土壤对CO2升高的响应,结果表明:高响应水稻品种分泌的根系生物量更大,土壤中的DOC增多,其DOC/SOC增加10.00%~67.00%,SOC稳定性下降。也有部分研究表明:短期内大气CO2升高对SOC活性组分无显著影响,如陈婧等[28]研究发现:在干旱农田生态系统中,1 a内大气CO2持续升高(275.0±6.6) μmol·mol−1后,正常和升高温度下土壤中的SOC和DOC含量均无显著差异。
土壤烷基化程度(即烷基碳/烷氧碳)和芳香度[即芳香碳/(烷基碳+烷氧碳+芳香碳)]可以反映SOC的降解和稳定程度[29-30]。烷基碳和芳香碳化学结构稳定,是SOC中比较稳定的部分,比例越高则SOC稳定性越好。目前,关于大气CO2升高方面的研究较少。陈栋[31]利用FACE平台研究了中长期(14 a)尺度下的农田土壤烷基化程度和芳香度的变化,发现大气CO2升高对各年代土壤均无显著影响,从时间尺度整体来看,不考虑CO2处理的影响,2004−2015年SOC芳香度显著降低了19.05%~22.73%,烷基化程度无显著差异。总的来说,SOC中稳定性组分减少,降低了SOC稳定性。
-
同位素标记法被用来定量SOC的转化率,可以直接反映SOC稳定性。基于15N在有机质分解、碳氮比逐渐降低过程中的分馏作用,和有机质分解产物与矿质颗粒结合形成稳定的SOC稳定机制。CONEN等[32]建立了自然生态系统中适用的SOC稳定性定量评价方法,该方法灵敏可靠,所测定结果被称为SOC的相对稳定性。CLERCQ等[33]证实该方法同样适用于农田SOC稳定性的定量评价。陈栋[31]基于新型有机质转化概念模型[颗粒有机质(POM)转化成矿物结合态有机质(mOM)]计算SOC相对稳定性,结果发现中长期大气CO2升高条件下,SOC相对稳定性显著降低了34.74%~67.11%,而在短期尺度下,SOC稳定性却有所增加(2.43~3.74倍)。
热指标主要包括SOC热重比,即单位有机碳下SOC矿化前后热重量差值的大小,是1种新兴土壤有机物稳定性评估指标,目前还未在大气CO2升高领域被广泛应用[5]。显然,大部分中长期模拟试验表明:大气CO2升高提高了农田、自然湿地和森林等生态系统SOC稳定性间接指标(SOC矿化率、微生物熵、DOC/SOC、POC/SOC等)[21, 34-35],降低了SOC稳定性,即不管SOC含量升高与否,中短期尺度下大气CO2升高会降低稳定老碳的比率,导致不同程度的激发效应,降低了SOC稳定性。长期CO2浓度升高降低了SOC的可分解性,可能使活性碳提高引起的激发效应削弱,即长期CO2升高降低SOC稳定性的趋势将逐渐减缓[4]。
-
大气CO2的持续升高间接影响植物-土壤-微生物系统的氮循环过程,改变土壤氮的供应能力,可能造成土壤氮限制[36]。长期CO2升高通过作用于硝化和反硝化过程,可能会促进、降低或不改变土壤氮净转化速率[37-39]。同时,氮源的增加也会引起生态系统的结构和功能发生变化,加快土壤酸化等问题,影响SOC稳定性[40]。目前,有关大气CO2升高与外源氮对SOC稳定性影响的文献主要涉及2个方面:①施加氮肥,主要研究农田和草原生态系统;②大气中的氮沉降,主要研究森林生态系统。
-
氮转化改变的方向和程度主要取决于土壤中氮的水平和类型。在氮限制土壤中,长期CO2升高降低了土壤中的有效态氮,即提高土壤有机质的碳氮比,氮的相对缺乏导致激发效应加剧,加速土壤中稳定碳的分解,从而将有机氮转化释放供植物吸收利用[20, 41],也就是说,在氮限制的条件下,长期CO2升高将会导致SOC稳定性的降低。因此,通过添加氮肥来改善CO2升高引起的土壤有效氮限制,从而抑制土壤中稳定碳的分解,减少总有机碳的损失,使SOC能够保持其稳定性[42]。然而,当土壤中含有充足氮源的时候,施氮将会促进作物、植物根系生物量和分泌物的增多,促使土壤团聚体的形成和稳定,进而保护SOC组分不被分解,甚至有可能增强SOC稳定性[25]。
PATERSON等[43]以自然生长的黑麦草Lolium perenne为研究对象,发现高浓度CO2和氮肥添加单独处理时,SOC矿化速率均显著增加,两者共同处理时SOC矿化率更高,SOC稳定性也随之降低。大量研究表明:施肥和CO2升高会促进植物的光合作用,提高植物产量和根系分泌物,导致土壤中不稳定有机碳增多[44]。寇太记等[45]研究了大气CO2升高和不同氮肥水平对水稻—小麦轮作制中SOC更新的影响,发现高CO2促进了作物向土壤中输送更多的碳,高CO2高氮处理比高CO2低氮处理的SOC更新率高2%左右,SOC含量增加。但是也有研究表明:施肥和CO2升高显著降低了土壤中DOC等不稳定有机碳成分,这是由于不稳定碳是土壤微生物的主要能量来源,根系分泌物的增多使微生物活跃起来,土壤微生物尤其是根际微生物的大量活动会导致土壤不稳定有机碳大量损失[46]。肖列等[47]发现:对于种植多年生C4草本植物白羊草Bothrioch loaischaemum的草地来说,CO2、施氮水平及两者交互作用对根际和非根际土壤DOC含量和DOC/TOC均无显著影响。近期农田13C-CO2连续标记实验发现:大气CO2升高条件下,促进了光合同化碳向地下的输入,促进了SOC的更新与循环,然而氮肥的施用仅仅促进了植物地上部分的生长,但并不能促进13C在土壤中的积累,反而降低了光合13C在土壤中的分配比例[48]。
-
世界范围内的化石燃料燃烧、工业发展、含氮化肥使用等,导致全球的氮循环加速,生态系统中的氮沉降也显著增加。在没有氮肥输入的自然和半自然生态系统中,氮沉降加速残落物和根的分解,减缓了大气CO2升高引起的氮限制,从而抑制土壤中稳定碳的分解,减少总有机碳的损失[49]。然而,当系统中的氮超过植物及土壤微生物对氮的蓄积能力时,生态系统将出现氮饱和的情况,导致土壤硝酸盐淋溶、土壤酸化等问题,进一步影响到陆地生态系统中的碳循环过程[50]。
CHEN等[21]和ZHANG等[23]利用开顶箱(OTC)发现:CO2升高和氮沉降显著提高了森林生态系统土壤中的ROC和POC,ROC/SOC显著增加了7.8%~44.6%,POC/SOC增加了0.3%~12.6%,活性有机碳比例增多,SOC稳定性下降。张继舟等[51]在模拟氮沉降和大气CO2增加对三江平原小叶章Deyenxia angustifolia群落土壤总有机碳的影响时发现:短期内大气CO2升高和氮沉降并不会影响到土壤总有机碳含量,且一定量的大气CO2升高会增加土壤中氮素的含量,过量的大气CO2反而减少土壤氮素。有研究认为:植物根系生物量的增加会引起碳输入的增多,但同时也会使土壤微生物的活动变得活跃,导致SOC加速分解,输入与分解达到平衡时,SOC含量看似并未发生变化,若是土壤中稳定碳被分解则其稳定性减弱[52]。龙凤玲等[16]在模拟大气CO2上升和氮添加对南亚热带模拟森林生态系统的影响时发现:高CO2高氮处理下,微生物熵显著增加了11.49%~29.97%,SOC稳定性降低。SAIYA-CORK等[53]研究表明:氮沉降使凋落物和土壤酶活性大部分增加,加速了生态系统中凋落物的分解,却抑制了土壤有机物的分解,因此可能使SOC含量增加,然而稳定性降低。
大气CO2升高和外源氮对生态系统的影响并不是单一孤立的,两者共同影响着不同生态系统中SOC的稳定性,其影响因素是多样的,如土壤理化性质、土壤微生物、凋落物成分、植物根系生长等[38, 54-56]。目前,碳氮交互作用对SOC稳定性影响的研究有限,存在很大的不确定性,还需要今后不断地深入研究。
-
有机碳在土壤中是否稳定主要取决于土壤环境因子(如土壤微生物、活性矿物表面、团聚体、养分、湿度、温度等)对有机碳的作用,是一个很复杂的过程,各种环境因子间相互影响[57]。大气CO2升高时,对环境因子产生影响进而改变了SOC的含量及稳定性。目前,研究中的观点有:①大气CO2升高可以提高部分植物的光合速率和净初级生产量,增加植物根系分泌物、根生物量和其他根际沉积物向土壤的碳输入,这可能会直接刺激土壤中微生物的生长和活性,从而加强土壤的呼吸作用,增加了SOC的矿化速率[58-59],甚至改变土壤微生物群落结构,最终可能促进土壤中稳定碳的分解,甚至产生显著的激发效应,改变SOC的含量和组成。②大气CO2升高改变植物体内有机物的化学成分[60],例如植株氮含量降低、碳氮比升高等[25, 27],从而改变输入土壤有机物的化学组成,可能导致土壤微生物可利用能源减弱,降低微生物碳的利用效率,影响SOC的分解速率[61-62],甚至增强激发效应[63-65]。③大气CO2升高促进了全球的碳循环,增加了土壤的碳通量和碳氮比,进而导致土壤微生物缺乏分解、合成等活动所需要的氮源,抑制了微生物的呼吸作用,土壤中的碳得到了积累。然而,当土壤中出现极端氮限制时,也会引起土壤碳的激发效应,促进SOC分解[66]。④大气CO2升高带来的综合影响可能会改变土壤的原有质地,而部分研究认为:土壤质地对SOC的存储起着重要作用[67],进而影响SOC稳定性,例如土壤团聚体越大,其内部土壤有机质越多,同时大颗粒团聚体避免了有机质的可接触性,有利于SOC的保存[68]。此外,土壤垂直深度也会影响SOC的稳定性,深层缺氧环境下的SOC转化速率慢,相应的也更稳定[69]。
总体而言,大气CO2升高对提高多种生态系统中SOC能力是有限的[70-71],但是会逐渐改变SOC结构组成,甚至改变分解有机碳的微生物群落结构,从而影响SOC稳定性[4, 72]。
-
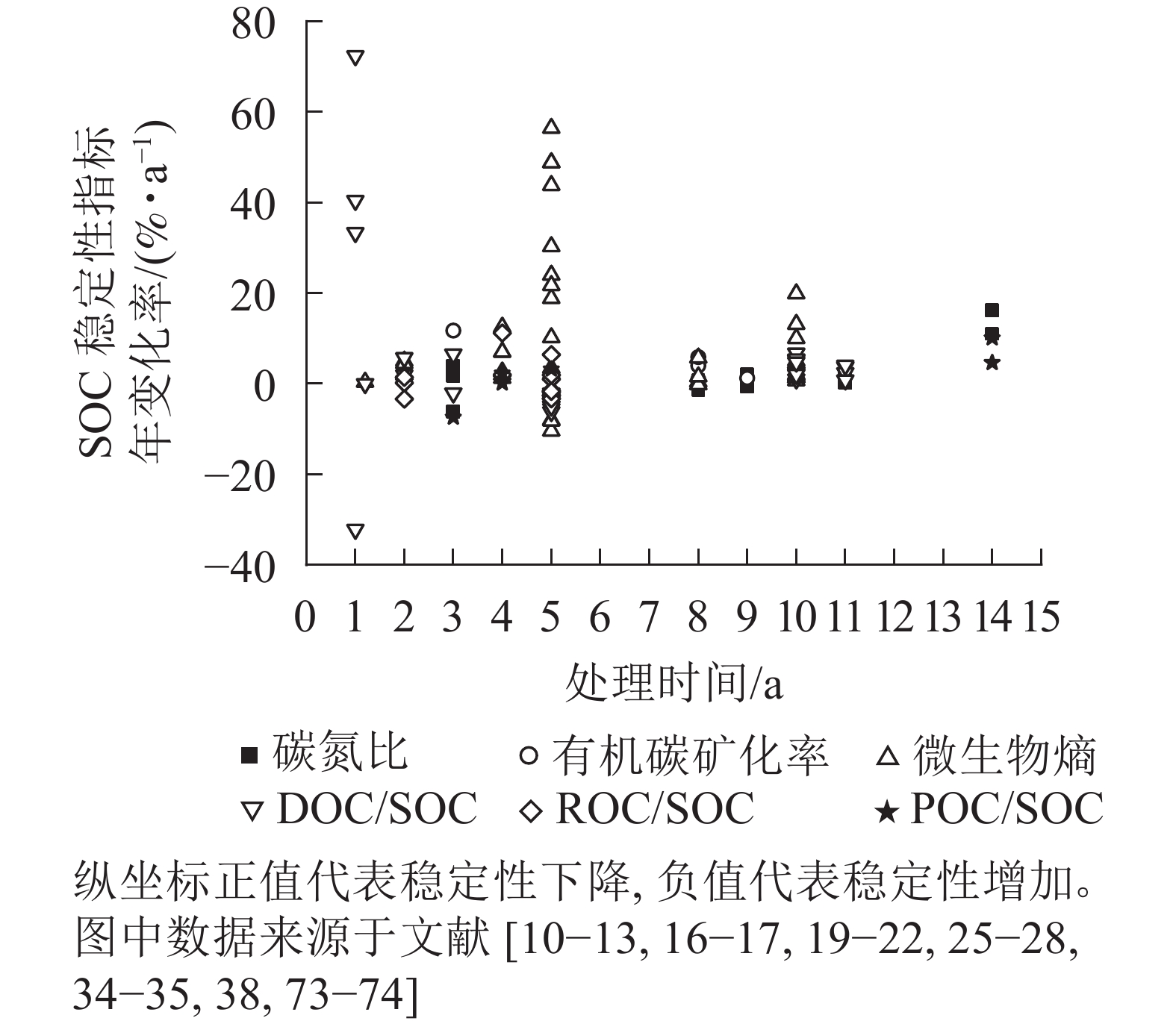
综上所述,在中长期尺度下,大气CO2升高能够改变土壤中有机碳总量及其稳定性。目前,已有的SOC稳定性间接性指标很多(生物指标、化学指标等),总结归纳出不同时间尺度下不同SOC稳定性指标的年变化率,将其作为SOC稳定性变化程度。如图1所示:以处理时间为自变量,SOC稳定性变化程度为因变量,可观察不同时间尺度对SOC稳定性的影响程度。
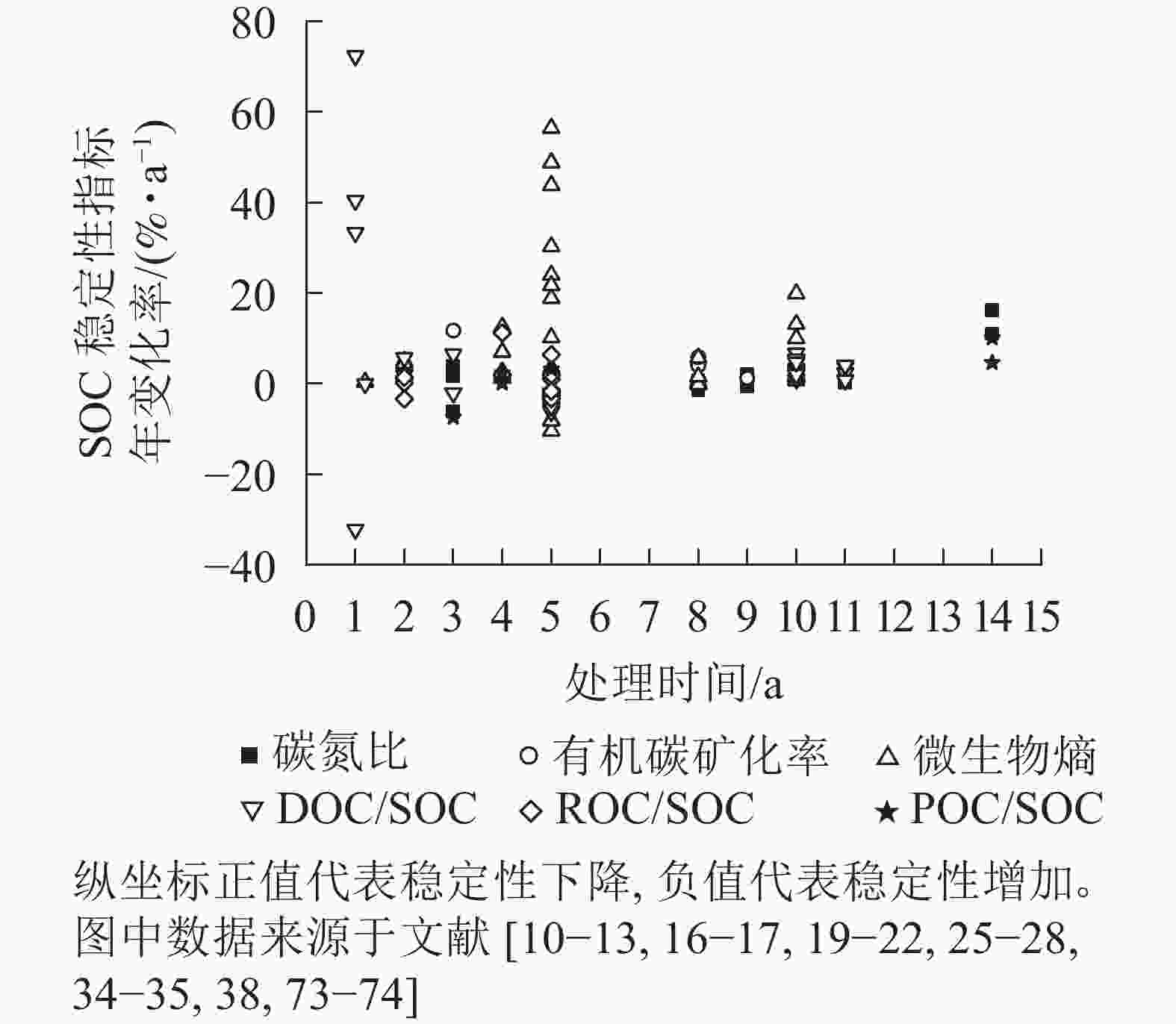
图 1 大气CO2升高对SOC稳定性影响的时间尺度效应
Figure 1. Time scale effect of elevated atmospheric CO2 on SOC stability
土壤中活性有机碳组分增多,则SOC稳定性下降。从图1可以看出:在大部分情况下,大气CO2升高导致了SOC稳定性变差,然而,随着处理时间的延长,SOC稳定性的变化逐渐变小。由此可见,土壤本身是具有适应能力的,适应了CO2升高的条件,SOC稳定性逐渐回归。也有部分数据表示:SOC稳定性是增加的,或许与土壤本质及种植的植物类型有关[62, 70]。据近年来的报告显示:大气CO2逐年上升且涨幅也在逐年增加[1],因此,长期大气CO2升高如何改变SOC稳定性还需要进一步研究。
-
土壤有机碳库是地球上最大且最具活动性的生态系统碳库之一,其微小变化将直接影响到其他的生态系统碳库。大气CO2升高促进了植物的光合作用,增加了植物向土壤中的光合碳输送,然而这并不意味着SOC含量是增加的,光合碳的过多输入也会刺激植物根部的生长,根部生物量的增加促进了微生物活性,进而提高了SOC的周转速率。大气CO2升高还可能改变植物体内有机物成分,改变土壤中碳氮交互作用,改变土壤质地,同样影响SOC的分解和转化,最终改变SOC稳定性。目前,有关大气CO2升高对SOC稳定性的研究大部分是基于自然生态系统的研究,CO2升高能带来土壤有机碳库的快速积累,但这种积累是短暂的,而且并不稳定。从中长期尺度来看,CO2升高改变了SOC稳定性,由于土壤的自我调节适应性,SOC稳定性降低的速率逐渐减缓。当大气CO2升高与氮耦合时,SOC稳定性下降,而氮素充足时则对SOC影响不显著。
SOC稳定性的改变通过影响土壤物质循环间接反馈到植物的生理生长,然而目前这方面的研究很少,从而潜在低估了生态系统水平上大气CO2升高带来的影响[4]。目前,农业生产系统提出“减氮增效”措施,但从稳定性的角度来看,减氮可能会使未来高CO2条件下的SOC稳定性降低加剧,意味着土壤的可持续能力可能受到挑战。如何采取有效措施保持SOC的稳定性,还需在今后的研究中去探索。
Effects of the mole fraction of elevated atmospheric CO2 on soil organic carbon stability
-
摘要: 随着工业的不断发展,全球大气二氧化碳(CO2)呈明显增加趋势。大气CO2的增加将会影响土壤有机碳(SOC)转化和更新,进而改变土壤碳的稳定性。研究大气CO2升高对SOC稳定性的影响,不但是评价陆地生态系统对气候变化反馈效应的重要环节,也对实现碳元素在土壤中的有效储存,对保持土壤肥力的可持续性具有重要意义。利用现有的文献资料,综述了大气CO2升高对SOC稳定性的影响及其稳定性指标(生物指标、化学指标、其他指标等),外源氮和大气CO2升高的交互作用对SOC稳定性的影响,以及SOC稳定性随时间尺度的变化趋势等。总结发现:大气CO2升高导致活性有机碳(溶解性有机碳、颗粒性有机碳、易氧化有机碳等)比例增多,SOC稳定性降低,尤其在氮限制的环境中,SOC稳定性更差。总结近年的研究成果发现:随着高CO2处理时间的加长,SOC稳定性降低速率逐渐减小,表明土壤本身具有一定的适应能力和自我恢复能力。最后展望了SOC稳定性变化对植物生理、生长的反馈影响。未来大气CO2升高对SOC稳定性的影响研究,应该着力于提高农田生态系统土壤肥力可持续性及提高农作物的产量产能。图1参74Abstract: The fast development of the industry has been accompanied with a significant increase of the global atmospheric CO2, which will affect the transformation and renewal of soil organic carbon (SOC), and then its stability. Therefore, an exploration of the effects of elevated atmospheric CO2 on SOC stability is not only an important attempt to evaluate the feedback effect of terrestrial ecosystem on climate change, but also of great significance to the effective storage of element C in soil and the sustainability of soil fertility. With an overview of previous researches, this study is aimed at a summary of the effects of elevated atmospheric CO2 on SOC stability and its stability indexes (biological index, chemical index, other index, etc.), the interaction between exogenous N and elevated atmospheric CO2 on SOC stability as well as the variation trend of SOC stability over time. The results showed that elevated atmospheric CO2 resulted in an increase in the proportion of labile organic carbon (readily oxidized carbon, particulate organic carbon, dissolved organic carbon, etc.), and a decrease in SOC stability, especially in nitrogen limitation environment. It was also found, with a summary of the research findings in recent decades, that there was a gradual decrease in the SOC stability reduction rate with the increase of high CO2 treatment time, indicating that the soil itself is equipped with the capacity to adapt and recover on its own. In conclusion, given the the feedback effect of SOC stability variation on plant physiology and growth, future researches on the effects of elevated atmospheric CO2 on SOC stability should be focused on promoting the sustainability of soil fertility in farmland ecosystem and increasing crop production and productivity. [Ch, 1 fig. 74 ref.]
-
Key words:
- soil science /
- soil organic carbon /
- stability /
- elevated atmospheric CO2 /
- N addition
-
[1] IPCC. Climate Change 2013: The Physical Science Basis. Working Group I Contribution to the Fifth Assessment Report of the Intergovernmental Panel on Climate Change[R]. Cambridge: Cambridge University Press, 2013. [2] BATJES N H. Total carbon and nitrogen in the soils of the world [J]. Eur J Soil Sci, 1996, 47(2): 151 − 163. [3] 何姗, 刘娟, 姜培坤, 等. 经营管理对森林土壤有机碳库影响的研究进展[J]. 浙江农林大学学报, 2019, 36(4): 818 − 827. HE Shan, LIU Juan, JIANG Peikun, et al. Effects of forest management on soil organic carbon pool: a review [J]. J Zhejiang A&F Univ, 2019, 36(4): 818 − 827. [4] CARRILLO C, DIJKSTRA F, LECAIN D, et al. Elevated CO2 and warming cause interactive effects on soil carbon and shifts in carbon use by bacteria [J]. Ecol Lett, 2018, 21(11): 1639 − 1648. [5] PLANTE A F, FERNÁNDEZ J M, HADDIX M L, et al. Biological, chemical and thermal indices of soil organic matter stability in four grassland soils [J]. Soil Biol Biochem, 2011, 43(5): 1051 − 1058. [6] 吴庆标, 王效科, 郭然. 土壤有机碳稳定性及其影响因素[J]. 土壤通报, 2005, 36(5): 743 − 747. WU Qingbiao, WANG Xiaoke, GUO Ran. Soil organic carbon stability and influencing factors [J]. Chin J Soil Sci, 2005, 36(5): 743 − 747. [7] 邢军会, 倪红伟, 王建波. 二氧化碳浓度升高与氮沉降对三江平原小叶章群落生物量累积及其分配格局的影响[J]. 中国农学通报, 2011, 27(13): 49 − 54. XING Junhui, NI Hongwei, WANG Jianbo. Effects of elevated CO2 concentration and N deposition on plant biomass accumulation and allocation in the communities of Deyeuxia angustifolia in Sanjiang Plain [J]. Chin Agric Sci Bull, 2011, 27(13): 49 − 54. [8] FONTAINE S, BARDOUX G, ABBADIE L, et al. Carbon input to soil may decrease soil carbon content [J]. Ecol Lett, 2004, 7(4): 314 − 320. [9] BHATTACHARYA S S, KIM K H, DAS S, et al. A review on the role of organic inputs in maintaining the soil carbon pool of the terrestrial ecosystem [J]. J Environ Manage, 2016, 167: 214 − 227. [10] MARHAN S, DEMIN D, ERBS M, et al. Soil organic matter mineralization and residue decomposition of spring wheat grown under elevated CO2 atmosphere [J]. Agric Ecosyst Environ, 2008, 123(1/3): 63 − 68. [11] 刘娟, 韩勇, 蔡祖聪, 等. FACE系统处理3年后淹水条件下土壤CH4和CO2排放变化[J]. 生态学报, 2007, 27(6): 19 − 20. LIU Juan, HAN Yong, CAI Zucong, et al. Changes of CH4 and CO2 emissions from soils under flooded condition after exposed to FACE (free-air CO2 enrichment) for three years [J]. Acta Ecol Sin, 2007, 27(6): 19 − 20. [12] 陈栋, 郁红艳, 邹路易, 等. 大气CO2浓度升高对不同层次水稻土有机碳稳定性的影响[J]. 应用生态学报, 2018, 29(8): 2559 − 2565. CHEN Dong, YU Hongyan, ZOU Luyi, et al. Effects of elevated atmospheric CO2 concentration on the stability of soil organic carbon in different layers of a paddy soil [J]. Chin J Appl Ecol, 2018, 29(8): 2559 − 2565. [13] TANEVA L, GONZALEZ-MELER M A. Decomposition kinetics of soil carbon of different age from a forest exposed to 8 years of elevated atmospheric CO2 concentration [J]. Soil Biol Biochem, 2008, 40(10): 2670 − 2677. [14] 聂阳意, 王海华, 李晓杰, 等. 武夷山低海拔和高海拔森林土壤有机碳的矿化特征[J]. 应用生态学报, 2019, 28(3): 748 − 756. NIE Yiyang, WANG Haihua, LI Xiaojie, et al. Characteristics of soil organic carbon mineralization in low altitude and high altitude forests in Wuyi Mountains, southeastern China [J]. Chin J Appl Ecol, 2019, 28(3): 748 − 756. [15] 康熙龙. 生物质炭施用对土壤有机碳矿化和分配及团聚体组成的影响[D]. 南京: 南京农业大学, 2015. KANG Xilong. Effects of Bllochar Amendment on Soil Organic Carbon Minerazation and Soil Aggregate Distribution and Organic Carbon Composition[D]. Nanjing: Nanjing Agricultural University, 2015. [16] 龙凤玲, 李义勇, 方熊, 等. 大气CO2浓度上升和氮添加对南亚热带模拟森林生态系统土壤碳稳定性的影响[J]. 植物生态学报, 2014, 38(10): 1053 − 1063. LONG Fengling, LI Yiyong, FANG Xiong, et al. Effects of elevated CO2 concentration and nitrogen addition on soil carbon stability in southern subtropical experimental forest ecosystems [J]. Chin J Plant Ecol, 2014, 38(10): 1053 − 1063. [17] 赵光影, 刘景双, 王洋, 等. CO2浓度升高对三江平原湿地活性有机碳及土壤微生物的影响[J]. 地理与地理信息科学, 2011, 27(2): 96 − 100. ZHAO Guangying, LIU Jingshuang, WANG Yang, et al. Effects of elevated CO2 concentration on active carbon in Sanjiang Plain freshwater marsh [J]. Geogr Geo-Inf Sci, 2011, 27(2): 96 − 100. [18] PROCTER A C, GILL R A, FAY P A, et al. Soil carbon responses to past and future CO2 in three Texas prairie soils [J]. Soil Biol Biochem, 2015, 83: 66 − 75. [19] BUTTERLY C R, PHILLIPS L A, WILTSHIRE J L, et al. Long-term effects of elevated CO2 on carbon and nitrogen functional capacity of microbial communities in three contrasting soils [J]. Soil Biol Biochem, 2016, 97: 157 − 167. [20] XU Qiao, JIN Jian, WANG Xiaojun, et al. Susceptibility of soil organic carbon to priming after long-term CO2 fumigation is mediated by soil texture [J]. Sci Total Environ, 2019, 657: 1112 − 1120. [21] CHEN Xiaomei, LIU Juxiu, DENG Qi, et al. Effects of elevated CO2 and nitrogen addition on soil organic carbon fractions in a subtropical forest [J]. Plant Soil, 2012, 357(1/2): 25 − 34. [22] HOFMOCKEL K S, ZAK D R, MORAN K K, et al. Changes in forest soil organic matter pools after a decade of elevated CO2 and O3 [J]. Soil Bio Biochem, 2011, 43(7): 1518 − 1527. [23] ZHANG Jing, TANG Xuli, HE Xinhua, et al. Glomalin-related soil protein responses to elevated CO2 and nitrogen addition in a subtropical forest: potential consequences for soil carbon accumulation [J]. Soil Biol Biochem, 2015, 83: 142 − 149. [24] KEIDEL L, LENHART K, MOSER G, et al. Depth-dependent response of soil aggregates and soil organic carbon content to long-term elevated CO2 in a temperate grassland soil [J]. Soil Biol Biochem, 2018, 123: 145 − 154. [25] WU Qicong, ZHANG Congzhi, YU Zhenghong, et al. Effects of elevated CO2 and nitrogen addition on organic carbon and aggregates in soil planted with different rice cultivars [J]. Plant Soil, 2018, 432(1/2): 245 − 258. [26] 关松, 窦森, 张大军, 等. 土壤腐殖质组成对大气二氧化碳浓度升高的响应[J]. 水土保持学报, 2006, 20(5): 186 − 188. GUAN Song, DOU Sen, ZHANG Dajun, et al. Responses of fractions of soil humus to free-air CO2 enrichment [J]. J Soil Water Conserv, 2006, 20(5): 186 − 188. [27] HU Zhengkun, ZHU Chunwu, CHEN Xiaoyun, et al. Responses of rice paddy micro-food webs to elevated CO2 are modulated by nitrogen fertilization and crop cultivars [J]. Soil Biol Biochem, 2017, 114: 104 − 113. [28] 陈婧, 陈法军, 刘满强, 等. 温度和CO2浓度升高下转Bt水稻种植对土壤活性碳氮和线虫群落的短期影响[J]. 生态学报, 2014, 34(6): 1481 − 1489. CHEN Jing, CHEN Fajun, LIU Manqiang, et al. Short-term effects of CO2 concentration elevation, warming and transgenic Bt rice cropping on soil labile organic carbon and nitrogen, and nematode communities [J]. Acta Ecol Sin, 2014, 34(6): 1481 − 1489. [29] 付琳琳. 生物质炭施用下稻田土壤有机碳组分、腐殖质组分及团聚体特征研究[D]. 南京: 南京农业大学, 2013. FU Linlin. The Study of Characteristics of Paddy Soil Organic Carbon Fractions, Huminstruvture and Aggregates under Different Biochar Amendments[D]. Nanjing: Nanjing Agricultural University, 2013. [30] 赵亚南. 长期不同施肥下紫色水稻土有机碳变化特征及影响机制[D]. 重庆: 西南大学, 2016. ZHAO Yanan. Characteristic and Mechanism of Organic Carbon Sequestration of Purple Paddy Soil under Long-Term Fertilization[D]. Chongqing: Southwest University, 2016. [31] 陈栋. 稻田土壤有机碳稳定性对大气CO2浓度升高的响应研究[D]. 无锡: 江南大学, 2018. CHEN Dong. Response of Soil Organic Carbon Stability under Atmospheric CO2 Concentration in a Paddy Field[D]. Wuxi: Jiangnan University, 2018. [32] CONEN F, ZIMMERMANN M, LEIFELD J, et al. Relative stability of soil carbon revealed by shifts in δ15 N and C∶N ratio [J]. Biogeosciences, 2008, 4(4): 123 − 128. [33] CLERCQ T D, HEILING M, DERCON G, et al. Predicting soil organic matter stability in agricultural fields through carbon and nitrogen stable isotopes [J]. Soil Biol Biochem, 2015, 88: 29 − 38. [34] INUBUSHI K, CHENG W, MIZUNO T, et al. Microbial biomass carbon and methane oxidation influenced by rice cultivars and elevated CO2 in a Japanese paddy soil [J]. Eur J Soil Sci, 2011, 62(1): 69 − 73. [35] AL-MALIKI S, JONES D L, GODBOLD D L, et al. Elevated CO2 and tree species affect microbial activity and associated aggregate stability in soil amended with litter[J]. Forests, 2017, 8(3): 70. doi: 10.3390/f8030070. [36] YIQI L, BO S, CURRIE W S, et al. Progressive nitrogen limitation of ecosystem responses to rising atmospheric carbon dioxide [J]. Bioscience, 2004, 54(8): 731 − 739. [37] CHENG Yi, ZHANG Jinbo, ZHU Jianguo, et al. Ten years of elevated atmospheric CO2 doesn’t alter soil nitrogen availability in a rice paddy [J]. Soil Biol Biochem, 2016, 98: 99 − 108. [38] 陈娜, 刘毅, 肖谋良, 等. CO2倍增和施氮对水稻不同生长期土壤反硝化细菌丰度的影响[J]. 环境科学研究, 2019, 32(4): 683 − 691. CHEN Na, LIU Yi, XIAO Mouliang, et al. Effect of elevated CO2 and nitrogen application on the abundance of soil denitrifying bacteria in different growth stages of rice [J]. Res Environ Sci, 2019, 32(4): 683 − 691. [39] USYSKIN-TONNE A, HADAR Y, YERMIYAHU U, et al. Elevated CO2 has a significant impact on denitrifying bacterial community in wheat roots[J]. Soil Biol Biochem, 2019, 142: 107697. doi: 10.1016/j. soilbio. 2019.107697. [40] 樊后保, 袁颖红, 王强, 等. 氮沉降对杉木人工林土壤有机碳和全氮的影响[J]. 福建林学院学报, 2007, 27(1): 1 − 6. FAN Houbao, YUAN Yinghong, WANG Qiang, et al. Effects of nitrogen deposition on soil organic carbon and total nitrogen beneath Chinese fir plantations [J]. J Fujian Coll For, 2007, 27(1): 1 − 6. [41] 邵兴华, 王爱斌. 施氮对水田和旱地有机碳和黑炭的影响[J]. 浙江农林大学学报, 2014, 31(4): 554 − 559. SHAO Xinghua, WANG Aibin. Organic carbon and black carbon with fertilization in paddy and upland soils [J]. J Zhejiang A&F Univ, 2014, 31(4): 554 − 559. [42] YAN Guoyong, XING Yajuan, WANG Jianyu, et al. Sequestration of atmospheric CO2 in boreal forest carbon pools in northeastern China: effects of nitrogen deposition [J]. Agric For Meteorol, 2018, 248: 70 − 81. [43] PATERSON E, THORNTON B, MIDWOOD A J, et al. Atmospheric CO2 enrichment and nutrient additions to planted soil increase mineralisation of soil organic matter, but do not alter microbial utilisation of plant- and soil C-sources [J]. Soil Biol Biochem, 2008, 40(9): 2434 − 2440. [44] ZHAO Guangying, LIU Jingshuang. Effects of elevated CO2 concentration on biomass and active organic carbon of freshwater marsh after two growing seasons in Sanjiang Plain, Northeast of China [J]. J Environ Sci, 2009, 21(10): 1393 − 1399. [45] 寇太记, 朱建国, 谢祖彬, 等. 大气CO2浓度升高和氮肥水平对麦田土壤有机碳更新的影响[J]. 土壤学报, 2009, 46(3): 459 − 465. KOU Taiji, ZHU Jianguo, XIE Zubin, et al. Effect of elevated atmospheric pCO2 and nitrogen level on replacement rate of soil organic carbon in winter wheat field [J]. Acta Pedol Sin, 2009, 46(3): 459 − 465. [46] WANG Xiaoguo, LI Changsheng, LUO Yong, et al. The impact of nitrogen amendment and crop growth on dissolved organic carbon in soil solution [J]. J Mt Sci, 2016, 13: 95 − 103. [47] 肖列, 刘国彬, 张娇阳, 等. CO2浓度升高、干旱胁迫和氮沉降对白羊草光响应曲线的影响[J]. 草地学报, 2016, 24(1): 69 − 75. XIAO Lie, LIU Guobin, ZHANG Jiaoyang, et al. Effects of elevated CO2, drought stress and nitrogen deposition on photosynthesis light response curves of Bothriochloa ischaemum [J]. Acta Agrestia Sin, 2016, 24(1): 69 − 75. [48] 任逸文, 肖谋良, 袁红朝, 等. 水稻光合碳在植物-土壤系统中的分配及其对CO2升高和施氮的响应[J]. 应用生态学报, 2018, 29(5): 1397 − 1404. REN Yiwen, XIAO Mouliang, YUAN Hongchao, et al. Allocation of rice photosynthates in plant-soil system in response to elevated CO2 and nitrogen fertilization [J]. Chin J Appl Ecol, 2018, 29(5): 1397 − 1404. [49] ZUO Xiaoan, KNOPS J M H. Effects of elevated CO2, increased nitrogen deposition, and plant diversity on aboveground litter and root decomposition[J]. Ecosphere, 2018, 9(2): e02111. doi: 10.1002/ecs2.2111. [50] BOBBINK R, HICKS K, GALLOWAY J, et al. Global assessment of nitrogen deposition effects on terrestrial plant diversity: a synthesis [J]. Ecol Appl, 2010, 20(1): 30 − 59. [51] 张继舟, 倪红伟, 王建波, 等. 模拟氮沉降和CO2浓度增加对三江平原小叶章群落土壤总有机碳和氮素含量的影响[J]. 地球与环境, 2013, 41(3): 216 − 225. ZHANG Jizhou, NI Hongwei, WANG Jianbo, et al. Effects of simulated nitrogen deposition and elevated CO2 concentration on soil organic carbon and nitrogen of Deyeuxia angustifolia community on the Sanjiang Plain [J]. Earth Environ, 2013, 41(3): 216 − 225. [52] van KESSEL C, HORWATH W R, HARTWIG U, et al. Net soil carbon input under ambient and-elevated CO2 concentrations: isotopic evidence after 4 years [J]. Glob Change Biol, 2000, 6(4): 435 − 444. [53] SAIYA-CORK K R, SINSABAUGH R L, ZAK D R. The effects of long term nitrogen deposition on extracellular enzyme activity in an Acer saccharum forest soil [J]. Soil Biol Biochem, 2002, 34(9): 1309 − 1315. [54] OSANAI Y, TISSUE D T, BANGE M P, et al. Plant-soil interactions and nutrient availability determine the impact of elevated CO2 and temperature on cotton productivity [J]. Plant Soil, 2017, 410(1/2): 87 − 102. [55] BLOOM A J, BURGER M, KIMBALL B A, et al. Nitrate assimilation is inhibited by elevated CO2 in field-grown wheat [J]. Nat Clim Change, 2014, 4(6): 477 − 480. [56] REICH P B, HOBBIE S E, LEE T D. Plant growth enhancement by elevated CO2 eliminated by joint water and nitrogen limitation [J]. Nat Geosci, 2014, 7(12): 920 − 924. [57] SCHMIDT M W I, TORN M S, ABOVEN S, et al. Persistence of soil organic matter as an ecosystem property [J]. Nature, 2011, 478(7367): 49 − 56. [58] 马红亮, 朱建国, 谢祖彬. 植物地上部分对大气CO2浓度升高的响应[J]. 生态环境学报, 2004, 13(3): 390 − 393. MA Hongliang, ZHU Jianguo, XIE Zubin. Study on response of vegetable aboveground to the elevated atmospheric CO2 [J]. Ecol Environ, 2004, 13(3): 390 − 393. [59] 罗艳. 土壤微生物对大气CO2浓度升高的响应[J]. 生态环境学报, 2003, 12(3): 108 − 111. LUO Yan. Response of soil microorganism to elevated atmospheric CO2 concentration [J]. Ecol Environ, 2003, 12(3): 108 − 111. [60] ZHU Chunwu, XU Xi, WANG Dan, et al. Elevated atmospheric [CO2] stimulates sugar accumulation and cellulose degradation rates of rice straw [J]. GCB Bioenergy, 2016, 8(3): 579 − 587. [61] MANZONI S, JACKSON R B, TROFYMOW J A, et al. The global stoichiometry of litter nitrogen mineralization [J]. Science, 2008, 321(5889): 684 − 686. [62] WANG Yanhong, YU Zhenhua, LI Yansheng, et al. Microbial association with the dynamics of particulate organic carbon in response to the amendment of elevated CO2-derived wheat residue into a mollisol [J]. Sci Total Environ, 2017, 607: 972 − 981. [63] PHILLIPS R P, FINZI A C, BERNHARDT E S. Enhanced root exudation induces microbial feedbacks to N cycling in a pine forest under long-term CO2 fumigation [J]. Ecol Lett, 2011, 14(2): 187 − 194. [64] KEILUWEIT M, BOUGOURE J J, NICO P S, et al. Mineral protection of soil carbon counteracted by root exudates [J]. Nat Clim Chang, 2015, 5(6): 588 − 595. [65] PUGH T A M, MÜLLER C, ARNETH A, et al. Key knowledge and data gaps in modelling the influence of CO2 concentration on the terrestrial carbon sink [J]. J Plant Physiol, 2016, 203: 3 − 15. [66] HU Shuijin, CHAPIN F S I, FIRESTONE M K, et al. Nitrogen limitation of microbial decomposition in a grassland under elevated CO2 [J]. Nature, 2001, 409(6817): 188 − 191. [67] RASMUSSEN C, HECKMAN K, WIEDER W R, et al. Beyond clay: towards an improved set of variables for predicting soil organic matter content [J]. Biogeochemistry, 2018, 137(5): 297 − 306. [68] von LÜTZOW M, KÖGEL-KGNABNER I, EKSCHMITT K, et al. Stabilization of organic matter in temperate soils: mechanisms and their relevance under different soil conditions-a review [J]. Eur J Soil Sci, 2010, 57(4): 426 − 445. [69] TFAILY M M, HESS N J, KOYAMA A, et al. Elevated [CO2] changes soil organic matter composition and substrate diversity in an arid ecosystem [J]. Geoderma, 2018, 330: 1 − 8. [70] QIAO Na, SCHAEFER D, BLAGODATSKAYA E, et al. Labile carbon retention compensates for CO2 released by priming in forest soils [J]. Global Change Biol, 2014, 20(6): 1943 − 1954. [71] GROENIGEN K J V, QI Xuan, OSENBERG C W, et al. Faster decomposition under increased atmospheric CO2 limits soil carbon storage [J]. Science, 2014, 344(6183): 508 − 509. [72] YANG Sihang, ZHENG Qiaoshu, YUAN Mengting, et al. Long-term elevated CO2 shifts composition of soil microbial communities in a Californian annual grassland, reducing growth and N utilization potentials [J]. Sci Total Environ, 2019, 652: 1474 − 1481. [73] 徐乔. 大气CO2浓度升高对稻田土壤有机碳的影响[D]. 北京: 中国科学院大学, 2014. XU Qiao. Effects of Elevated Atmospheric CO2 on Soil Organic Carbon in Rice Paddy Field[D]. Beijing: University of Chinese Academy of Sciences, 2014. [74] BLACK C K, DAVIS S C, HUDIBURG T W, et al. Elevated CO2 and temperature increase soil C losses from a soybean-maize ecosystem [J]. Global Change Biol, 2017, 23(1): 435 − 445. -

-
链接本文:
https://zlxb.zafu.edu.cn/article/doi/10.11833/j.issn.2095-0756.20200502
 点击查看大图
点击查看大图
图(1)
计量
- 文章访问数: 2887
- HTML全文浏览量: 483
- PDF下载量: 83
- 被引次数: 0
