




-
人是空间活动的主体。在以往研究[1-4]中,空间活力的外在表征被定义为城市空间中的人及其行为活动[5],人群活动特征能直接反应空间的活力情况。刘颂等[6-7]以大数据为基础总结黄浦江滨水空间活力时空变化,提出将人群、时间、空间作为影响空间活力的3个维度。陈菲[8]依据城市形态活力理论[9]研究严寒城市公共空间景观活力环境,杨春侠等[10]采用行为地图记录驻留活动数据,探讨滨水空间要素对驻留活动的显著影响因子并提出对策;刘瑞雪等[11]采用行为观察法研究植物景观空间活力及其与环境因素的关系。这些研究均为景观空间活力特征研究提供了方法借鉴。上塘河作为世界文化遗产京杭大运河的重要支流,是杭州段最古老的运河,其滨河景观带的活力现状关系着沿岸区域的发展和人民生活的改善。探讨杭州上塘河滨河景观带活力特征,可为激活运河城市滨河景观带空间活力提供理论支撑。
-
本研究范围为上塘河杭州段。上塘河作为线性空间地物,纵向空间多样,水系跨越杭城内外,流经城镇、郊野与乡村。本研究着重关注大运河遗产保护中一级缓冲区内的景观空间,因此选取上塘河滨河景观带中的城北体育公园(A1)、东新门广场(A2)、永丰公园(A3)、望宸公园(A4)、水陆院遗址(A5)、千桃园(A6)、天都公园(A7)、西洋桥公园(A8)作为研究对象,以视觉感知距离,以道路、围栏、绿篱等地物要素作为观测边界,截取面积40 m×200 m的8个典型样方并进行编号。
-
城市空间活力的外在表征包括人群基本属性和时空活力行为特征[6],因此,本研究采用行为注记法,围绕活力表征的人群、时间和空间维度展开活力特征调查。3个维度评价指标的具体量化方法如表1所示。行为注记法是结构化观察法的重要分支,在环境行为学研究[12-13]中得到广泛运用,由 ITTELSON等[14]发展而来,包括场地的图形化表现、观察行为的定义、观察与记录的间隔、观察时的规范、数据编码与统计等。以上5项要素确保观察数据具有较高的准确度。行为注记法作为行之有效的调查方法,在国外已有广泛的实践运用,国内戴菲等[15]引申为结构式行为观察,并探讨了具体的调查过程。本研究将行为注记、平面绘图和数据统计相结合,全面客观地评价上塘河滨河景观带的活力特征。调查从调研资料收集、各维度评价指标、数据采集与处理、调查结果分析4个阶段开展。
表 1 各维度评价指标
Table 1. Evaluation indexes of each dimension
活力表征维度 评价指标 量化分析方法 人群维度 人群多元化 不同年龄的人群混合度公 式计算 行为多样性 基于辛普森多样性的行为 多样性公式计算 时间维度 高峰活动频数 某空间高峰时的活动数量 动态波动系数 某时段内活动人数的增长 率偏差情况(标准差) 活动持续程度 空间内人群活动持续时长 空间维度 活动频数 空间内活动数量累计总和 聚集密度强度 空间内活动频数总和/该空 间铺地面积 活动行为分布 各空间内的行为注记图中 的行为分布类型 夏季和冬季由于经常受极端气温的影响,游人活动呈现较大的不稳定性,因此,本研究数据采集时间选择在杭州最适合出游的春季(平均气温10~22 ℃)进行。根据天气情况,在2021年3月12日、3月14日、3月26日、3月28日,对每个调研样地进行连续2周,每周1个工作日和1个双休日的实地调研。为了解不同时间段的空间活力情况,调查时段为6:00—20:00,共观测5个时段,每个时段观测15 min。由8组成员采用行为注记法的5项要素原则对8个样地中的人群属性和时空分布信息进行观察记录,4个观察日共搜集6 176个样本数据。预调研阶段先了解样地功能、路线和人群活动,之后确定样地范围、观测步骤和活动类别。正式调研阶段,观测员采用行为注记法和快照法,通过观测表、注记图、两步路全球定位系统(GPS)等记录人群活动信息。数据采集完成后,在地理空间分析软件ArcGIS 10.4中对典型样地的人群活动数据建立缓冲区,并将其与所处位置的地理信息进行叠加分析,生成人群活动聚集密度分布图,用于探讨滨河空间人群聚集分布特征。
-
人群多元化是城市活力空间中不同性别、各年龄层级和特点的人群相互区别和混合的特性。为确定样地内人群多元化程度,将人群年龄层级划分为0~3岁幼儿、3~14岁儿童、15~44岁青年、45~59岁中年、60岁以上老年人。不同的人群从属于不同社会属性类别,多元混合的人群对空间的使用需求度不一样,尤其是弱势群体(老人和儿童)的生理和心理需求、行为习惯与青年、中年不同。这就要求空间具有容纳多元社会群体的能力。因此,引用统计学对信息论的统计方式,度量随机人群在整个群体的不确定程度。公式为:
$$ H\left(x\right)=-\sum _{i=1}^{n}{P}_{i}\mathrm{lg}{P}_{i}\mathrm{。} $$ (1) 式(1)中:H(x)表示随机变量x的熵,Pi为 xi的概率。熵值越大, 混合度越高;熵值越小, 混合度越低。
人群多元性是前提,活动多样化是目的。本研究根据现场调查结果将人群日常行为分为六大类:动态户外类(骑行、跑步、散步遛弯、广场舞),静态休闲类(静坐闲聊、下棋、驻足观赏、拍照),家庭亲子类(烧烤野餐、亲子活动、亲水活动),日常通勤类(工作通行、工作清理、维护检修),文化艺术类(绘画书法、自然认知、曲艺表演),娱乐游戏类(球类、健身设施、儿童娱乐)。充满活力的空间有着容纳多样化活动的特性[4]。借鉴生态学中物种多样性概念,对人群多样的行为活动,引入“行为多样性”概念[11]。行为多样性可反映空间环境与人群行为的相关性,是活动内容在空间中丰富度和均匀度的综合指标,采用Simpson’s多样性指数来衡量某一样地中人群行为活动的相对丰度。其公式为:
$$ D=1-\sum _{i=1}^{S}{\left(\frac{{n}_{i}}{N}\right)}^{2}\mathrm{。} $$ (2) 式(2)中:D表示行为多样性,S表示活动内容的种类,ni表示第i个活动内容的数量,N表示样地内所有活动内容的总人数。
-
空间活力中时间维度具有动态性和持续性的特征。相关文献[6-7, 16]也曾提到:人群活动的波动系数、活动数量、持续时长等都是时间维度的重要参考指标。空间维度的最大特征为聚集性,空间强度可以从活动频数和聚集密度进行量化分析。根据观测记录的各样地的活动频数和可活动面积,可计算样地的聚集密度和活动频数。活动行为分布和聚集密度形态可反映空间与人群活动的空间分布关系。采用核密度估算法(KDE法)能更好地研究空间分布情况,此方法能将活动的强度转化为密度并进行估值,从而使空间中的活动点产生光滑的表面密度[17]。本研究借助行为注记图和拍摄时照片附带的GPS信息,在ArcGIS 10.4中建立典型样地的空间行为数据缓冲区,并将其与所处位置的地理信息进行叠加,生成人群活动聚集密度分布图,探讨滨河空间人群聚集分布特征。
-
如图1所示:上塘河整体滨河空间人群多元混合度较好,但普遍为中、青年群体占比多,弱势群体占比较少,仅A1、A3、A6样地各年龄层占比较为多元、均衡。人群多元化指数表明(表2):各样地多元混合度从大到小依次为A3、A6、A1、A7、A5、A2、A8、A4。2种数据分析结果基本一致。主要原因是A1、A3、A6样地除了滨河游步道外,还设有具有通勤功能的水上巴士、适合老人的健身设施、休憩空间和儿童娱乐玩耍的活动场地,场地水岸空间功能更多元复合。由此可见,多元复合功能的空间人群多元性会更高。

图 1 各样地行为主体占比图
Figure 1. Chart of the proportion of actors in various plots
表 2 各维度空间活力量化指数
Table 2. Spatial vitality indexes of each dimension
样地 样地基本信息 人群维度 时间维度 空间维度 观测范围/m2 可活动面积/m2 人群多元化 行为多样性 高峰频数/人 平均波动系数/% 活动频数/人 聚集密度/(人·m−2) A1 25 247 2 643 0.611 0.798 210 21.7 1 058 0.400 A2 16 842 2 470 0.551 0.817 159 47.6 808 0.327 A3 21 252 4 392 0.627 0.781 242 39.3 870 0.198 A4 30 446 3 801 0.448 0.667 36 11.7 185 0.048 A5 24 130 4 162 0.581 0.837 226 40.0 680 0.163 A6 20 964 4 208 0.621 0.882 589 298.9 1 240 0.294 A7 24 130 4 922 0.587 0.843 150 86.4 858 0.174 A8 14 771 2 752 0.460 0.795 107 21.1 477 0.173 -
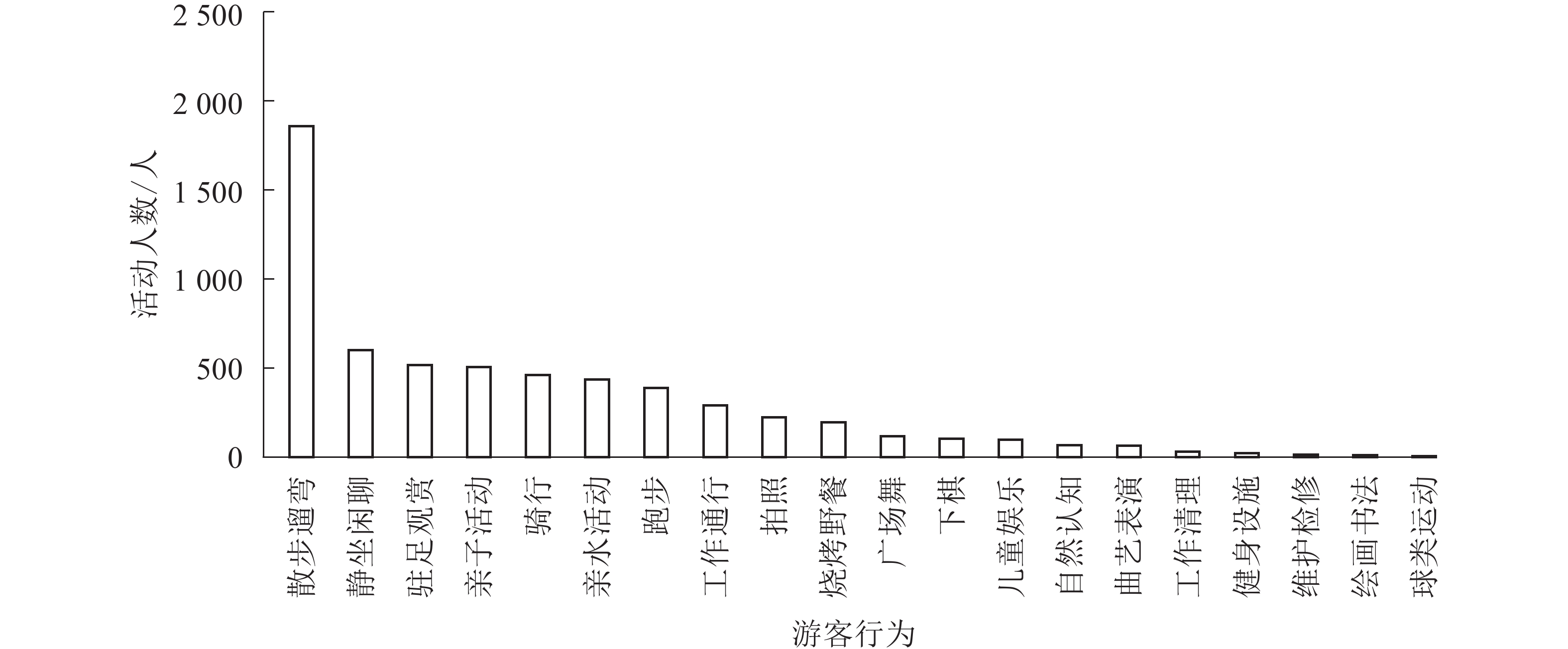
各样地行为多样性从大到小依次为A6、A7、A5、A2、A1、A8、A3、A4 (表2)。上塘河滨河景观带行为多样性总体均值为0.857,其中A6行为多样性最高,为0.882,A4行为多样性最低,为0.667。各样地中行为多样性存在较大差距。各项活动内容的活动人数结果(图2)显示:散步遛弯、静坐闲聊等自发行为活动占54.7%,工作通行、清理等必要性行为占5.5%,亲子活动、亲水活动、静坐闲聊、广场舞等社会性行为占39.8%。整体活动情况为自发性行为最多,社会性行为次之,必要性行为最少。主要原因是游客出于休闲、健身等活动需求,会开展自发性活动,如A6样地内设有特色的皋亭桃林,A7样地有法兰西风格的雕塑、小品等景观设施,能吸引更多的游客。由此可见,具有历史价值的文化景源和景观性设施能增加游客自发性行为。
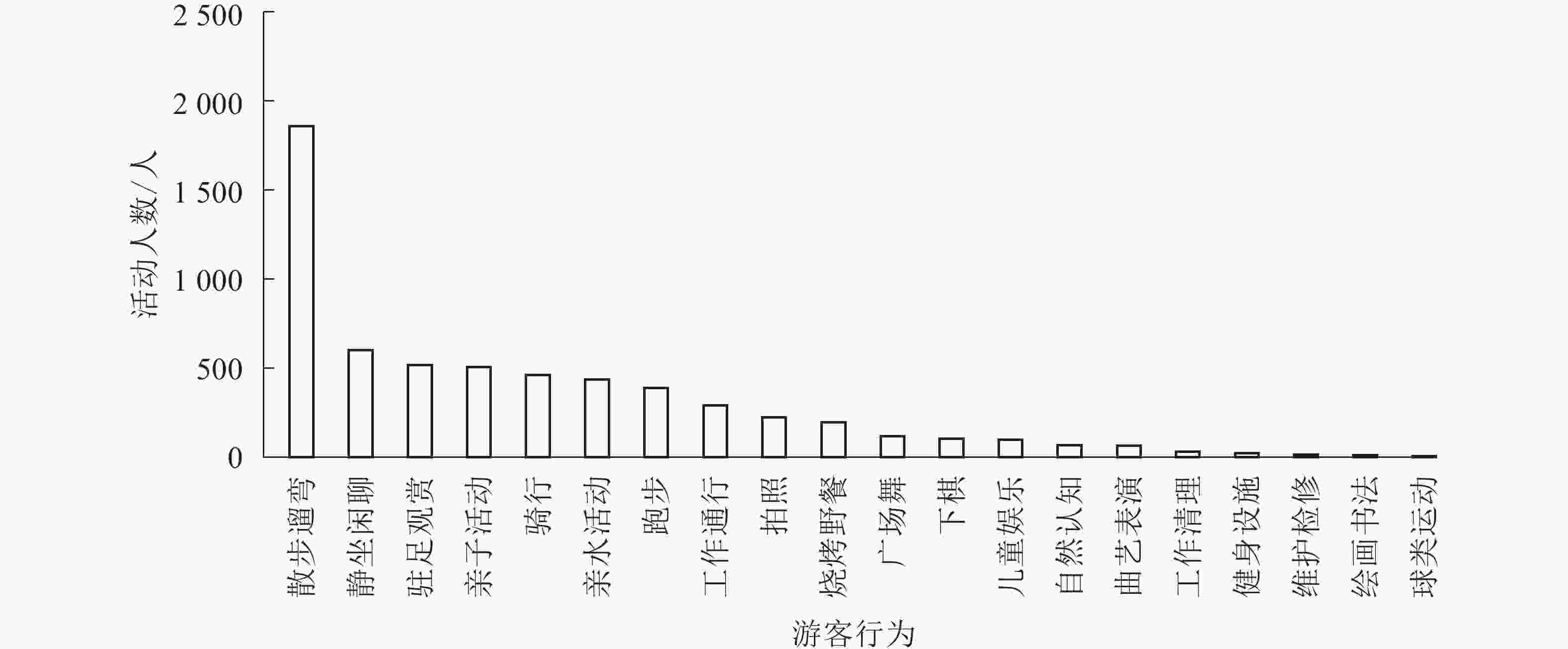
图 2 各项活动内容统计图
Figure 2. Statistics of various activities
-
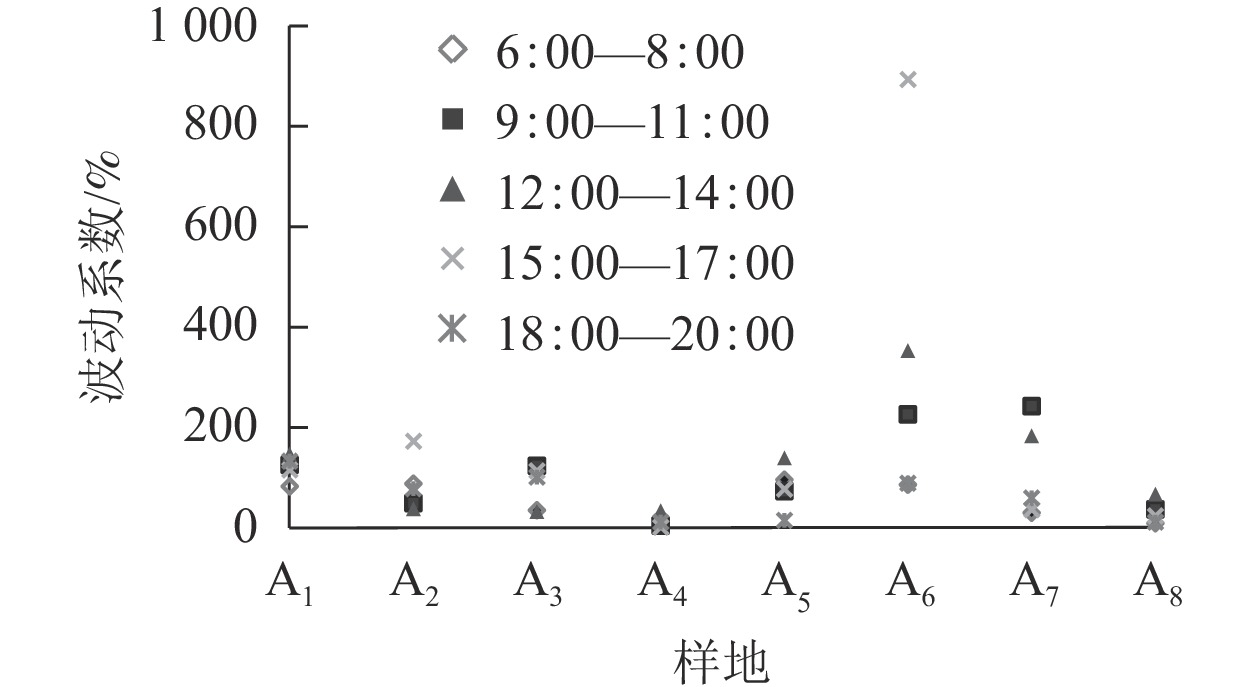
如表2所示。上塘河各样地单位时间内的波动系数差异较大,各样地波动系数从大到小依次为A6、A7、A2、A5、A3、A1、A8、A4。虽然A5、A6、A7日波动系数变化趋势大,空间内的活动强度不稳定,但空间活动频数增加,空间更具活力。A1、A3、A6波峰出现在15:00—17:00,A3、A7波峰出现在9:00—11:00,A1、A5波峰出现在12:00—14:00。A6在各时间段的波动系数都比其他空间大,A3出现双波峰,A8、A4整体无明显变化(图3)。这主要是由于A5、A6、A7等样地沿线会进行夜间景观亮化美化管理,通过整合闸坝、桥梁、建筑等光源,能渲染出夜间热闹的滨水氛围。且在节假日,A6、A7通过举行特色水岸文化活动,吸引更多人前来,因此空间波动系数较大,空间更具活力。

图 3 各样地不同时段的波动系数对比图
Figure 3. Comparison of fluctuation coefficients in various plots
-
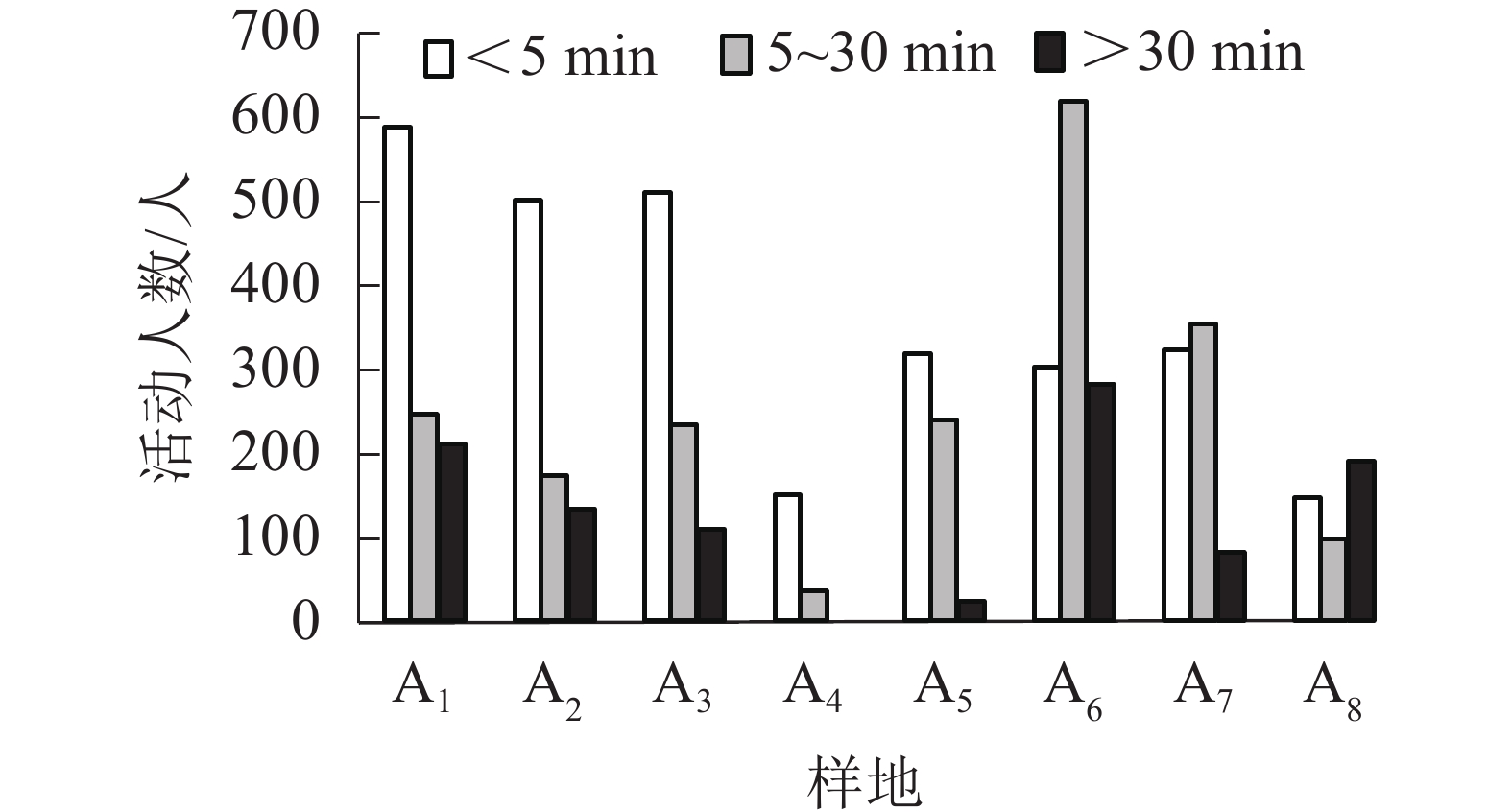
从图4可知:上塘河不同活动持续时间的人数从多到少依次为<5 min、5~30 min、>30 min。各样地活动时长在<5 min的人数从多到少依次为A1、A3、A2、A7、A5、A6、A8、A4,活动时长为5~30 min的人数从多到少依次为A6、A7、A1、A5、A3、A2、A8、A4,活动时长>30 min的人数从多到少依次为A6、A1、A8、A2、A3、A7、A5、A4。主要由于上塘河整体缺乏完善的配套设施和休闲设施,而活动持续性强的A6、A1样地内配套设施完善,空间功能综合性高,有可供人群长时间停留的座椅、活动设施、观景亭廊和亲水平台,A1样地还设有水岸广场,能开展曲艺表演、绘画下棋等文化活动,参与活动的人群多且驻留时间长。由此可见,完善场地配套设施、休闲设施能增加活动频数,能延长人群活动。

图 4 各样地活动时间持续性统计图
Figure 4. Statistics of the duration of activities in various plots
-
从表2可知:各样地的活动频数从高到低依次为A6、A1、A3、A7、A2、A5、A8、A4,表明A6、A1的活动频数较高,空间活力较高。单位时间段的人群聚集密度从高到低依次为A1、A2、A6、A3、A5、A7、A8、A4。其中,最高的是A1,为0.400人·m−2;其次是A2,为0.327人·m−2;A6为0.294人·m−2;最低的是A4,为0.049 人·m−2。A1和A6在空间强度的2组量化指标具有较大差异。
比较A6、A1的样地基本信息(表2)发现:空间内可活动面积是影响人群聚集密度的关键。A6相对A1的空间尺度更大,内部基本设施齐全,功能完善,拥有容纳大量人群活动的草坪空间,而A1样地空间形态狭长,多为滨水绿道。人际交往需要保持相互之间的距离。EDWARD[18]曾将人际交往空间分为4种距离:亲密距离、个人距离、社交距离、公众距离,但当陌生人进入个人距离46~76 cm范围内会让人形成自我空间被触犯、感觉不舒服的抵触反应。由此可见,在空间强度中不能一味追求过高的聚集密度,单位空间内合理的人群聚集密度能够带来更大的活力。如A1、A2等狭长的线性空间,当活动频数处于峰值时,过高的聚集度会让人不舒服,此时,应适当在局部拓宽活动场地面积,增加空间距离。
-
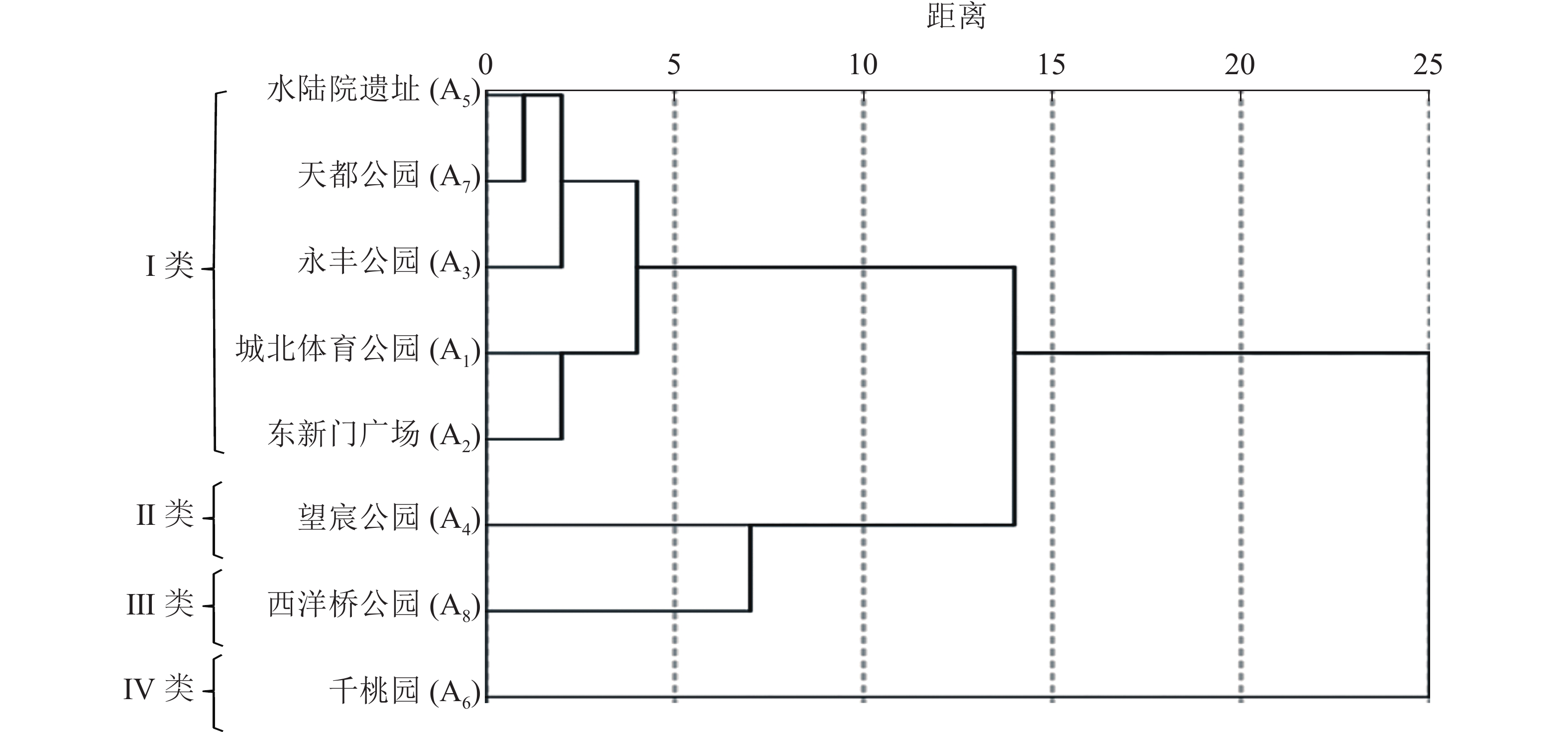
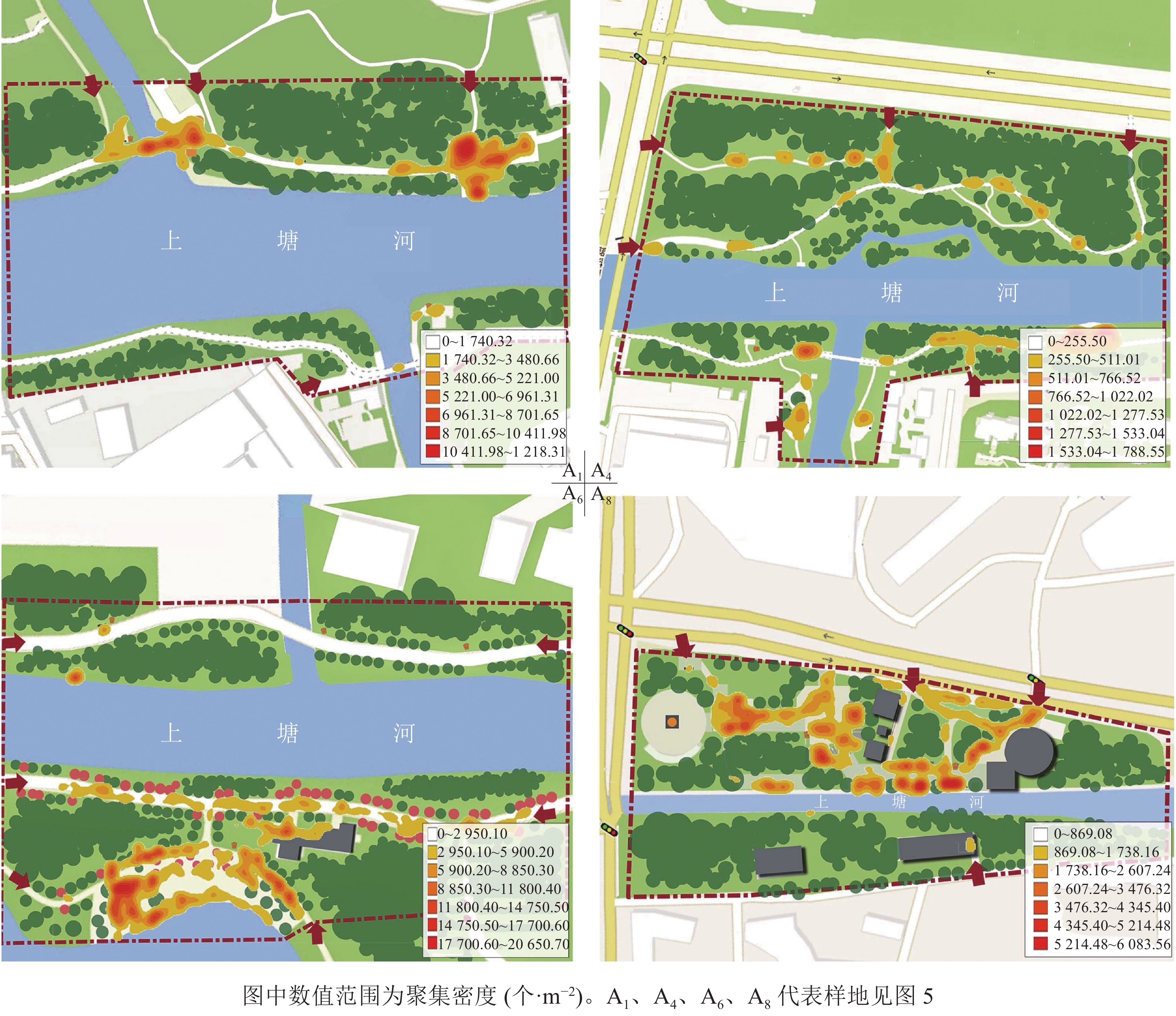
单一空间维度的数值能对空间进行纵向比较,但无法具体判读空间活力状况。为更好地区别各样地的活力状况,本研究进行了多指标分类分析。把8个样地的各维度指标数值(表2)标准化处理后运用SPSS 21.0进行系统聚类分析。在标定距离为5的水平上,将不同活力状况的样地划分为4种类型(图5),得到上塘河滨河景观带4种空间活力类型,并对其典型样地A1、A4、A6、A8运用ArcGIS 10.4软件绘制人群活动聚集密度分布图(图6)。
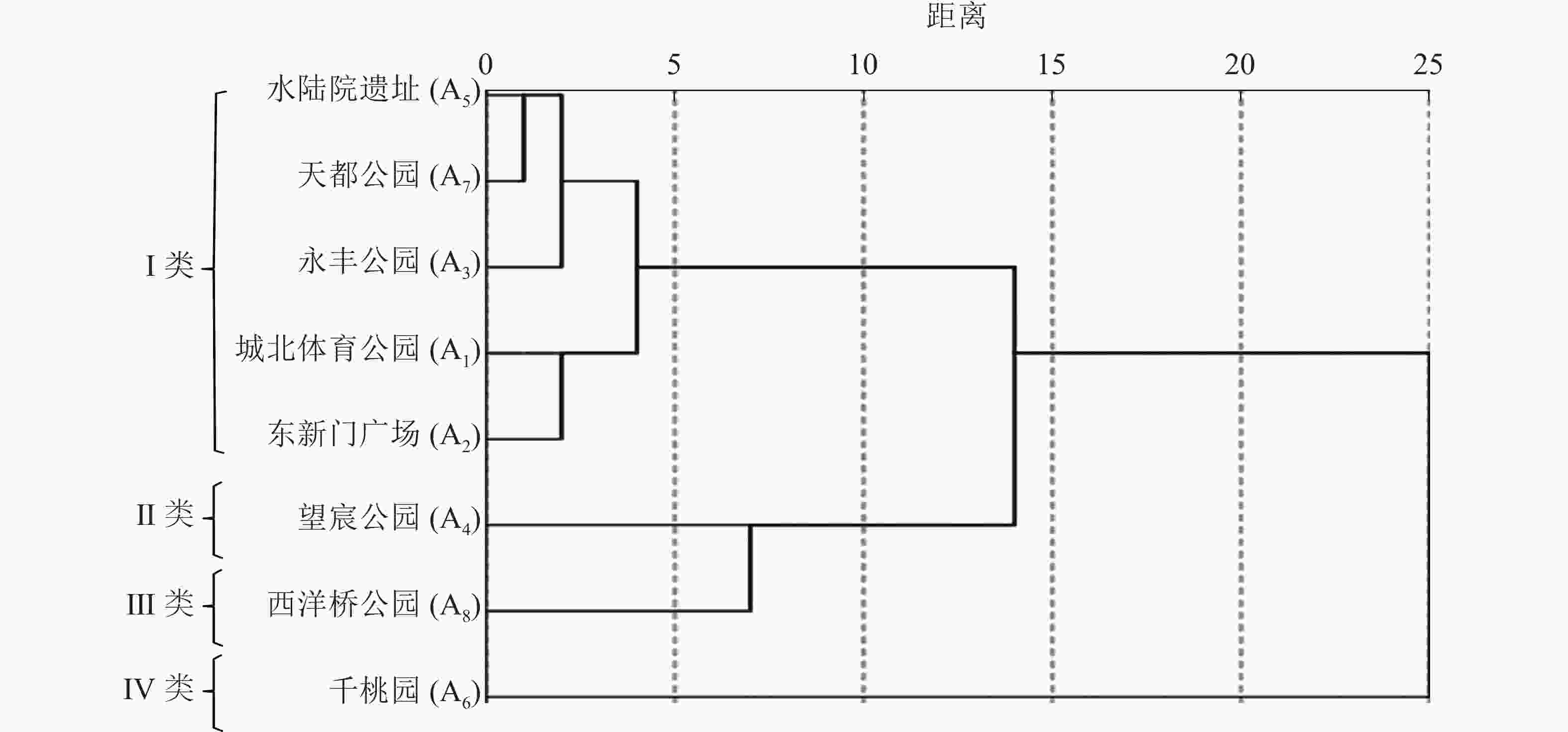
图 5 各空间树状分析图
Figure 5. Analysis of each spatial cluster
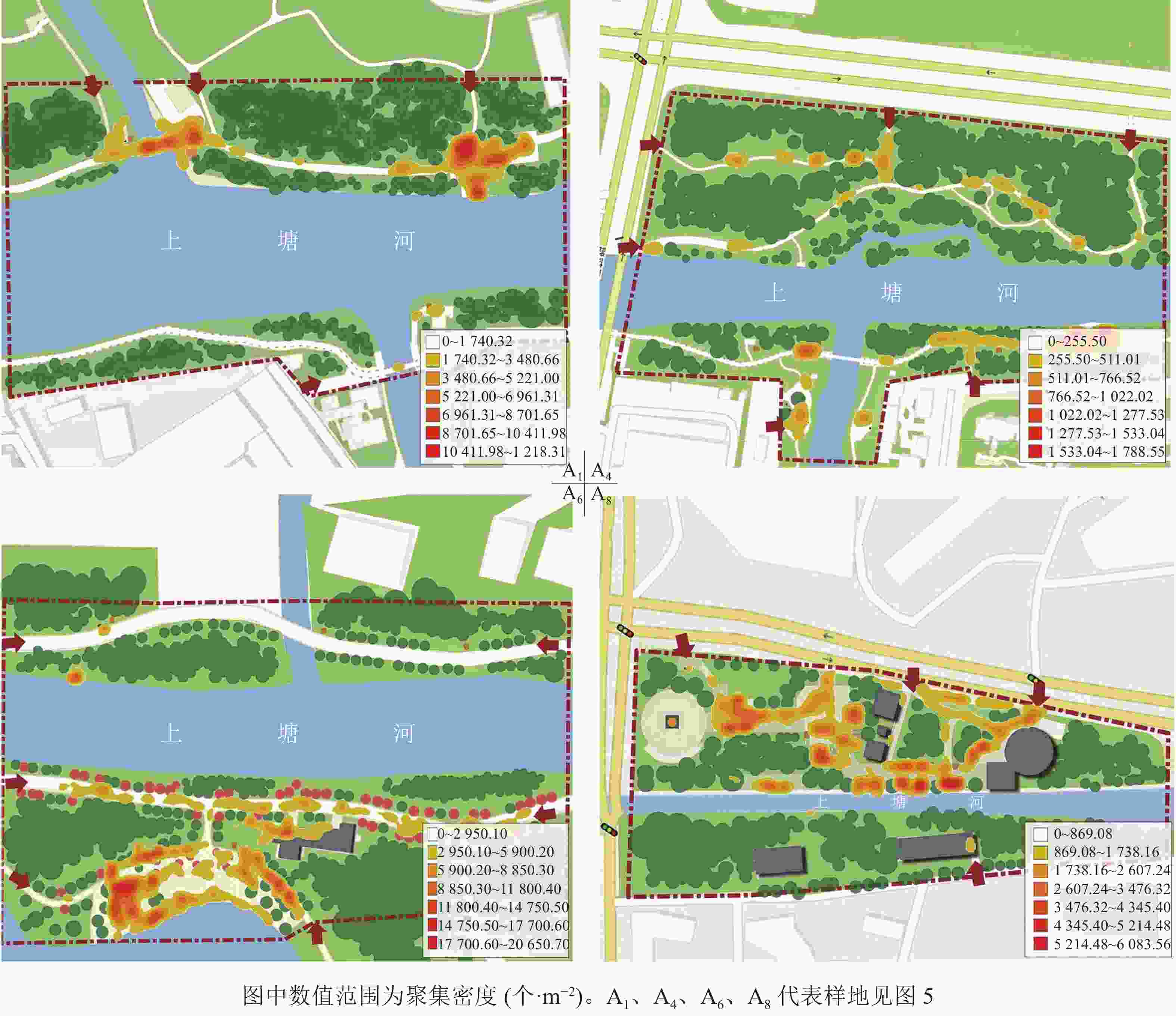
图 6 4类典型样地人群活动聚集密度分布
Figure 6. Distribution of population activity density in 4 typical sample plots
4种滨河景观带的空间活力类型分别是①高密度-高多样性空间。这类滨河空间分布特征为人群行为活动范围大,活力点聚集密度值高,聚集形态大部分为面状、线状,四周也有点状活力点。此类空间多为乡村类景观和自然类景观,分布在上塘河建设完成的主题滨河公园内。本研究中典型空间为A6。从图6的A6空间聚集密度分布可看出:活动空间与乔木、铺装场地及草地的边界处人群聚集度较高。主要是由于游客对于心理安全的追求,通常休息时选择可依靠的边界处驻留,活动时则更倾向开阔、滨河风光优美又有高大乔木遮阴的空间。②高密度-低多样性空间。这类滨河空间分布特征为人群行为分布均匀,活力点聚集密度值高,聚集形态多为大块的面状,点、线性状较少。这类空间多为现代城镇生活段景观,分布在上塘河沿岸两侧居住区附近。本研究中的典型空间为A1。受周边居住用地因素的限制,多为滨河绿道,空间狭长且活动面积小,在特定时间段内人口密度高、流动快、活动量大,从而形成瞬时聚集密集,但通常只能开展几类固定的活动项目。低多样性是由于沿河绿道能够满足中老年群体早、晚时段散步遛弯的日常需求。③低密度-高多样性空间。这类滨河空间分布特征为人群行为分布随意,活力点分散;聚集密度值偏低,活力范围小,聚集形态为点、线状,空间内有多处点状活力聚集地。这类空间多为现代城镇类景观,分布在城镇生活段和工业段的交界处。本研究中的典型空间为A8。这类空间内小尺度活动场地多、功能多样,在绿道基础上还建有开放、半开放的休憩平台和亭廊,人们更容易在围合感强的空间中开展活动。④低密度-低多样性空间。这类滨河空间分布特征为人群行为分布随意,零星几个活力点,整体聚集密度值低,活力范围小,聚集形态呈散点状分布。这类空间多为现代城镇工业段景观,分布在城市灰色区域或在建地块周边。本研究中的典型空间为A4。该样地一侧为高速公路,另一侧为小区内部绿地,所以滨河绿道贯通受阻,造成步行交通可达性差,且绿地整体以防护和生态功能为主,缺乏活动空间和景观维护,是活力最弱的空间。
从上塘河的4种空间活力类型分析可知:空间边界、空间活动面积、空间围合感、休闲设施、景观维护程度等内部因素,以及步行便捷度、周边用地性质、周边人口密度等外部因素都是影响滨河空间活力的因素。
-
从人群维度、时间维度和空间维度,对杭州上塘河滨河景观带活力特征的研究结果表明:营建复合滨河景观,促进空间多元功能融合,挖掘山水、人文景源,增设景观装置小品,完善基础配套设施,营造活力水岸夜景,举办文化互动活动能够提升上塘河滨河景观带的活力。
基于上述结论,对杭州上塘河滨河景观带空间活力提升提出以下建议:①促进空间多元功能融合,形成复合滨河景观。应优化空间各项功能,满足综合服务需求,塑造与功能相符的多样化、复合式特色空间,如在水岸休闲空间中,融入娱乐、文化、休闲、科普、运动等功能,满足多年龄层次的多元化康体文娱需求,促进多样化的人群活动。②挖掘山水、人文景源,提升各区段自身吸引力。要明确滨河空间活力提升应与周围环境相适宜,空间活力暨受自身要素影响,也受周边用地性质、发展潜力影响,没有必要全段建成高活力空间。通过挖掘的景观资源,如皋亭山景、临平山景等自然景源和运河文化、诗词等人文景源,可激活低活力空间景观特色,从而提升自身吸引力。③完善基础配套设施建设。对铺装、道路、环卫、座椅设施等维护整修,加强公园内部的特色景观性建设。将运河文化融入到配套设施中时,还应将当地文化图案融入到老旧的拱桥、闸坝附近活动场所的设计中,合理改造沿线景观节点。④营造水岸互动模式,活化水岸功能。应增设水岸码头、水上巴士,恢复河道航运功能,营建滨河夜景、水岸舞台,促进人与水互动。这不仅能改善沿岸群众生产生活,还能实现航运功能与休闲生活功能融合,将单一的滨河绿化转变为具有可观、可留、可动的多元化空间。此外,还应完善沿岸夜景照明,从而引导游人前进,保障场地内活动的安全开展。⑤创新运河文化体验,分段设计主题景观。开展放花灯、赛龙舟等特色民俗活动,唤醒人们对运河往事的记忆。创造“一段一景一文化”的滨河景观,以特色文化景观吸引更多年轻人了解上塘河,发挥其服务社会、弘扬优秀传统文化的社会功能。
Research on vitality characteristics of riverside landscape belt of Shangtang River in Hangzhou based on tourist behavior
-
摘要:
目的 具有千年历史的杭州上塘河正遭遇活力不足的困境。探讨杭州上塘河滨河景观带活力特征,可为激活运河城市滨河景观带空间活力提供理论支撑。 方法 以上塘河一级缓冲区内的8个样地[城北体育公园(A1)、东新门广场(A2)、永丰公园(A3)、望宸公园(A4)、水陆院遗址(A5)、千桃园(A6)、天都公园(A7)、西洋桥公园(A8)]景观空间为研究对象,采用行为注记法对上塘河滨河景观带进行了活力特征调查,采用聚类分析和核密度估算法(KDE法)对调查结果进行分析。 结果 ①从人群维度来看,人群多元化指标从大到小依次为A3、A6、A1、A7、A5、A2、A8、A4,行为多样性从大到小依次为A6、A7、A5、A2、A1、A8、A3、A4,主要原因是A3、A6水岸空间多元复合,历史文化遗址、景观性设施等吸引人们开展自发性活动。②从时间维度来看,人群活动的波动系数从大到小依次为A6、A7、A2、A5、A3、A1、A8、A4 ,活动时长大于30 min的人数从多到少依次为A6、A1、A8、A2、A3、A7、A5、A4。主要原因为A6、A1内有滨河夜景、文化活动和完善的配套设施能延长人群驻留时间。③从空间维度来看,活动强度从大到小依次为A6、A1、A3、A7、A2、A5、A8、A4,单位时间段的人群聚集密度从高到低依次为A1、A2、A6、A3、A5、A7、A8、A4。主要原因是人群空间分布会受到空间边界、空间活动面积、空间围合感、周边用地性质、周边人口密度等多因素影响。 结论 营建复合滨河景观,促进空间多元功能融合,挖掘山水、人文景源,增设景观装置小品,完善基础配套设施,营造活力水岸夜景,举办文化互动活动,是提升上塘河滨河景观带活力的有效措施。图6表2参18 Abstract:Objective With a history of thousands of years, the Shangtang River in Hangzhou is facing the challenge of insufficient vitality. The purpose of this study is to explore the vitality characteristics of the riverside landscape belt of the Shangtang River, so as to provide research support for activating the spatial vitality of the riverside landscape belt of the canal city. Method The landscape space of 8 sample plots [Chengbei Sports Park (A1), Dongxinmen Square (A2), Yongfeng Park (A3), Wangchen Park (A4), Shuiluyuan Site (A5), Qiantaoyuan (A6), Tiandu Park (A7), and Xiyangqiao Park (A8)] in the first-level buffer zone of the Shangtang River were taken as the research objects, and the vitality characteristics of the riverside landscape belt were investigated by behavioral annotation method. The cluster analysis and kernel density estimation (KDE) were used to analyze the survey results. Result (1) From the population dimension, the order of population diversity indicators from large to small were A3 A6, A1, A7, A5, A2, A8, and A4, and the behavior diversity from large to small was A6, A7, A5, A2, A1, A8, A3, and A4. The main reason lied in that the waterfront space of A3 and A6 was more diverse and complex, and historical and cultural sites and landscape facilities attracted people to carry out spontaneous activities. (2) From time dimension, the fluctuation coefficients of crowd activities from large to small were A6, A7, A2, A5, A3, A1, A8, and A4. The number of people with activity duration longer than 30 minutes was A6, A1, A8, A2, A3, A7, A5, and A4 in descending order. The main reason was that there were riverside night scenes, cultural activities and perfect supporting facilities in A6 and A1, which attracted people to extend their stay. (3) From spatial dimension, the activity intensity was A6, A1, A3, A7, A2, A5, A8, and A4 in descending order, and the crowd density per unit time period from high to low was A1, A2, A6, A3, A5, A7, A8, and A4. The main reason was that the spatial distribution of the population would be affected by internal and external factors such as spatial boundary, spatial activity area, sense of spatial enclosure, nature of surrounding land, and surrounding population density. Conclusion Constructing composite riverside landscape, promoting the integration of multiple spatial functions, tapping the source of landscape and cultural scenery, adding landscape installation sketches, improving infrastructure, creating a vibrant waterfront nightscape, and holding cultural interactive activities are effective measures to enhance the vitality of the riverside landscape belt of the Shangtang River. [Ch, 6 fig. 2 tab. 18 ref.] -
Key words:
- tourist behavior /
- riverside landscape belt /
- spatial vitality /
- vitality characteristics
-
图 6 4类典型样地人群活动聚集密度分布
Figure 6 Distribution of population activity density in 4 typical sample plots
表 1 各维度评价指标
Table 1. Evaluation indexes of each dimension
活力表征维度 评价指标 量化分析方法 人群维度 人群多元化 不同年龄的人群混合度公 式计算 行为多样性 基于辛普森多样性的行为 多样性公式计算 时间维度 高峰活动频数 某空间高峰时的活动数量 动态波动系数 某时段内活动人数的增长 率偏差情况(标准差) 活动持续程度 空间内人群活动持续时长 空间维度 活动频数 空间内活动数量累计总和 聚集密度强度 空间内活动频数总和/该空 间铺地面积 活动行为分布 各空间内的行为注记图中 的行为分布类型  下载: 导出CSV
下载: 导出CSV
表 2 各维度空间活力量化指数
Table 2. Spatial vitality indexes of each dimension
样地 样地基本信息 人群维度 时间维度 空间维度 观测范围/m2 可活动面积/m2 人群多元化 行为多样性 高峰频数/人 平均波动系数/% 活动频数/人 聚集密度/(人·m−2) A1 25 247 2 643 0.611 0.798 210 21.7 1 058 0.400 A2 16 842 2 470 0.551 0.817 159 47.6 808 0.327 A3 21 252 4 392 0.627 0.781 242 39.3 870 0.198 A4 30 446 3 801 0.448 0.667 36 11.7 185 0.048 A5 24 130 4 162 0.581 0.837 226 40.0 680 0.163 A6 20 964 4 208 0.621 0.882 589 298.9 1 240 0.294 A7 24 130 4 922 0.587 0.843 150 86.4 858 0.174 A8 14 771 2 752 0.460 0.795 107 21.1 477 0.173
下载: 导出CSV
-
[1] GEHL J. Life between Buildings[M]. New York: van Nosrand Reinhold, 2003. [2] LYNCH K A. The Image of the City[M]. Boston: The MIT Press, 1971. [3] JACOBS J. The Death and Life of Great American Cities[M]. New York: Vintage Books, 2012. [4] BENTLEY I. Responsive Environments: A manual for Designers[M]. Princeton: Architectural Press, 1985. [5] 王玉琢. 基于手机信令数据的上海中心城区城市空间活力特征评价及内在机制研究[D]. 南京: 东南大学, 2017. WANG Yuzhuo. Research on Urban Spatial Vitality Characteristic Evaluation and Internal Mechanism of Shanghai Central City Based on Mobile Phone Signaling Data[D]. Nanjing: Southeast University, 2017. [6] 刘颂, 赖思琪. 大数据支持下的城市公共空间活力测度研究[J]. 风景园林, 2019, 26(5): 24 − 28. LIU Song, LAI Siqi. Measurement of urban public space vitality based on big data [J]. Landscape Archit, 2019, 26(5): 24 − 28. [7] 刘颂, 赖思琪. 基于多源数据的城市公共空间活力影响因素研究——以上海市黄浦江滨水区为例[J]. 风景园林, 2021, 28(3): 75 − 81. LIU Song, LAI Siqi. Influence factors of urban public space vitality based on multi-source data: a case study of Huangpu River waterfront area of Shanghai [J]. Landscape Archit, 2021, 28(3): 75 − 81. [8] 陈菲. 严寒城市公共空间景观活力评价研究[D]. 哈尔滨: 哈尔滨工业大学, 2016. CHEN Fei. Research on Landscape Activity Evaluation of Public Space in Winter City[D]. Harbin: Harbin Institute of Technology, 2016. [9] 蒋涤非. 城市形态活力论[M]. 南京: 东南大学出版社, 2007. JIANG Difei. The Theory of City Form Vitality [M]. Nanjing: Southeast University Press, 2007. [10] 杨春侠, 邵彬. 滨水公共空间要素对驻留活力的影响和对策——以上海黄浦江2个典型滨水区为例[J]. 城市建筑, 2018(2): 40 − 47. YANG Chunxia, SHAO Bin. Influence of waterfront public space elements on lingering vitality and strategies: taking two typical waterfronts along Huangpu River, Shanghai as Examples [J]. Urbanism Archit, 2018(2): 40 − 47. [11] 刘瑞雪, 许晓雪. 城市公园植物景观空间活力及环境因素影响研究[J]. 中国园林, 2018, 34(增刊 2): 160 − 164. LIU Ruixue, XU Xiaoxue. Space vitality and environmental impact of plant landscape in urban parks [J]. Chin Landscape Archit, 2018, 34(suppl 2): 160 − 164. [12] FRANCIS M, REIMANN A, LINDSEY P, et al. Urban Open Space: Designing for User Needs[M]. Washington D C: Island Press, 2004. [13] 李斌. 环境行为理论和设计方法论[J]. 西部人居环境学刊, 2017, 32(3): 1 − 6. LI Bin. Environment-behavior theories and design methodology [J]. J Human Settlements West China, 2017, 32(3): 1 − 6. [14] ITTELSON W H, RIVLIN L G, PROSHANSKY H M. Environmental Psychology: Man and His Physical Setting[M]. New York: Holt, Rinehart and Winston, 1976. [15] 戴菲, 章俊华. 规划设计学中的调查方法4——行动观察法[J]. 中国园林, 2009, 25(2): 55 − 59. DAI Fei, ZHANG Junhua. The survey methods in planning and design 4: action observation [J]. Chin Landscape Archit, 2009, 25(2): 55 − 59. [16] 胡佳奕. 共享理念下济南市住区公共空间活力优化策略[D]. 济南: 山东建筑大学, 2019. HU Jiayi. Strategies to Promote Residential Vitality under the Concept of Sharing[D]. Ji’nan: Shandong Jianzhu University, 2019. [17] 曾忠平, 王雅丽, 彭浩轩. 基于SOPARC和KDE的游客游憩行为研究——以武汉东湖绿道为例[J]. 中国园林, 2019, 35(12): 58 − 62. ZENG Zhongping, WANG Yali, PENG Haoxuan. Research on tourist recreation behavior based on SOPARC and KDE: taking Wuhan East Lake Greenway as an example [J]. Chin Landscape Archit, 2019, 35(12): 58 − 62. [18] EDWARD T H. The Hidden Dimension[M]. New York: Anchor Books, 1966. -

-
链接本文:
https://zlxb.zafu.edu.cn/article/doi/10.11833/j.issn.2095-0756.20210352
 点击查看大图
点击查看大图

图(6) / 表(2)
计量
- 文章访问数: 1929
- HTML全文浏览量: 450
- PDF下载量: 35
- 被引次数: 0


