





-
薄壳山核桃Carya illinoensis是一种重要的木本油料树种,又是珍贵的干果树种[1]。中国的亚热带东部地区、长江流域和云南等10多个省均建立了薄壳山核桃引种区,并已建成种质资源库、采穗圃、品种园和早实丰产示范林[2]。到目前为止,薄壳山核桃生产在国内仍未实现产业化,坚果消费大部分依赖进口,造成这种现象的一个重要原因是建园时未科学配置授粉树,导致授粉不良,生理落果严重[3]。薄壳山核桃是典型的雌雄同株、雌雄异熟植物,在建园时需要注意品种搭配,选择雌雄花期相匹配并且能够相互亲和的品种搭配种植,必要时在花期进行人工辅助授粉[4]。在生产中,薄壳山核桃散粉期,仅2~3 d,而雌花发育进程不一,且有等待授粉习性,短期内不能充分授粉,人工辅助授粉是一项重要的丰产措施[5]。所以花粉的采集和有效保存在生产中具有重要意义。薄壳山核桃‘马汉’‘Mahan’品种是当前中国栽培最为广泛的品种之一,也是常用的授粉品种。本研究以品种‘马汉’为研究对象,通过研究不同散粉期花粉活力差异以及不同储藏条件和储藏时间对花粉活力影响的研究,找出最佳的花粉采集时间和最适储藏条件,为人工辅助授粉提供最优保证。
-
试验地位于浙江省建德市莲花镇齐平村薄壳山核桃早实丰产试验林基地,29°34′42″N,119°18′21″E,海拔为110 m,年平均气温为16.0 ℃,年降水量1 818.8 mm。供试材料为薄壳山核桃‘马汉’品种12年生健康植株。散粉期花粉活力测定材料取即将散粉期(花药由绿变黄)、散粉初期、散粉盛期、散粉末期雄花序散出花粉,不同储藏条件花粉活力测定的花粉取自散粉盛期雄花序散出的花粉。
-
调查方法参考《经济林研究法》《果树研究法》中有关生物学调查部分和山核桃的一些性状调查方法[6]。在基地挑选3株营养水平相近、长势基本一致的植株作为标准株,取每植株东、南、西、北4个方向,每个方向选取10个结果枝进行挂牌,观察记录雄花芽萌动日期、雄花序生长日期、花萼开裂期、花药由绿变黄、散粉初期(以25 %小花花药散开为标准)、散粉盛期(以50 %以上小花花药散开为标准)、散粉末期(以花药完全开裂,花粉基本散尽,花序变黑褐色为标准)及小花脱落的变化过程。
-
①花粉采集。采集即将散粉期、散粉初期、散粉盛期、散粉末期4个时期的雄花序。将各时期雄花序平摊在硫酸纸上,将已经散出的花粉收集,花序置避风阳光下晾晒,每隔半小时收集1次花粉。收集的花粉用100目筛子过筛去除杂质。②花粉保存。4个散粉时期花粉保存:常温干燥保存,每天测花粉活力。不同储藏条件下花粉保存:将薄壳山核桃散粉盛期的花粉随机分成6份,装入密封袋内,置于常温密封、常温密封干燥;4 ℃低温密封和4 ℃低温密封干燥;-70 ℃超低温密封和-70 ℃超低温密封干燥内(干燥条件即在密封袋内装入500 g变色硅胶)。储藏2,4,6,8,10,30,50,70,90,120,360 d后测各组的花粉活力。③花粉活力测定方法(FCR荧光染色法)[7-8]。a.配制母液1(SS1):1.75 mol·L-1蔗糖,3.32 mmol·L-1硼酸,3.05 mmol·L-1硝酸钙,3.33 mmol·L-1硫酸镁,1.98 mmol·L-1硝酸钾,蒸馏水定容后,置4℃低温保存备用。另外,为了避免由渗透压引起的花粉破裂,可以适当增加蔗糖浓度,还可以通过增加盐的浓度增强荧光效果。b.配制母液2(SS2):把双乙酸荧光素溶于丙酮中,配成浓度为7.21 mmol·L-1后,置棕色玻璃瓶,4 ℃低温保存备用。c.工作液:使用时,取8~12滴SS2于10 mL SS1中,混匀直到混合液变为轻乳状即为工作液。操作时,取2~3滴工作液滴于花粉上,盖上盖玻片,2 min后置荧光显微镜下观察。重复3次·处理-1,观察5个视野·重复-1,统计每个视野内花粉颗粒总数(明场光照下)和被荧光染色花粉颗粒总数。④花粉活力计算方法:有生活力花粉百分数=(视野内变绿色花粉颗粒数目/视野内花粉颗粒总数)×100%。
-
试验数据采用Excel 2007计算和分析,对百分率数据进行反正弦平方根转换,其独立样本t检验和方差分析处理均采用SPSS 18.0版统计软件进行统计学分析处理,多重比较采用Duncan修复极差法分析。
-
‘马汉’为典型雌蕊先熟型品种,雄花芽于3月中旬开始萌动,经9~12 d后芽开始绽开,芽绽开后长出3束葇荑花序,花序直挺,0.5~1.1 cm左右,斜上生长,此为初花期。4月中旬花序伸长,开始软垂,雄花序迅速伸长,小花形成,并由深绿色变成浅绿色。4月下旬,花苞开放,花药发育,每个花序由114~126朵小花组成。
雄蕊散粉期经过花萼开裂→即将散粉期(花药由绿变黄)→散粉初期(25%小花花药开裂)→散粉盛期(50%以上小花花药开裂)→散粉末期(花药完全开裂,花序变黑,花粉基本散尽)→小花脱落的变化过程。雄蕊散粉周期短,且发育进程不一致。如图 1所示,5月6日花药由绿变黄,占雄花比例的95.22%。此后,散粉雄花序比列逐渐增大,雄蕊进入散粉期。5月7日,散粉初期花序占44.79%,散粉盛期花序占11.90%,进入散粉的雄花序占到56.69%比例。5月8日,各散粉期比例呈明显正态分布,散粉盛期花序比例增大,占46.60%,散粉比例达到75.29%,此时进入散粉盛期。5月9日,即将散粉期花序仅有4.57%,散粉盛期比例最大,达52.09%。5月10日,雄蕊进入散粉末期,比例为80.57%。散粉后,小花枯萎脱落,进入落花期。

图 1 薄壳山核桃散粉期雄花所处发育状态比例
Figure 1. Ratio of male flowers during four development stages
-
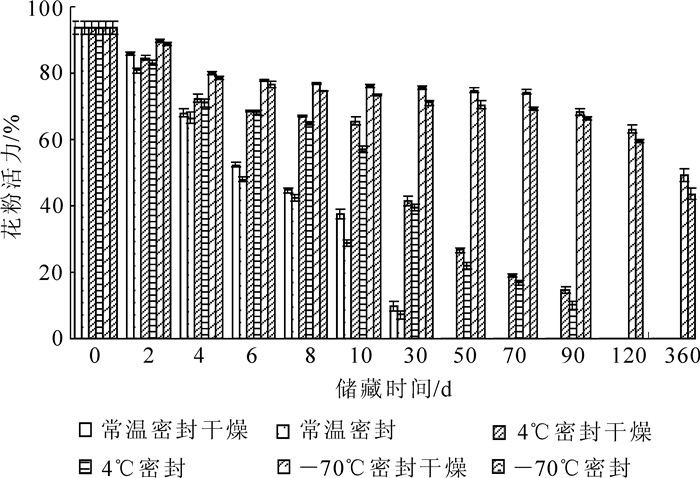
如表 1和表 2所示:不同散粉期花粉生活力差异显著,各时期花粉的耐储藏性也差异显著,表现为即将散粉期>散粉初期>散粉盛期>散粉末期。在整个散粉过程中,即将散粉期和散粉初期花粉活力最高,达到90%以上,两者之间无显著性差异;散粉盛期花粉活力次之,花粉活力为88.39%;散粉末期花粉活力最差,仅为79.60%,与花药黄色时期和散粉初期差异显著。将4个时期的花粉在常温干燥条件下储藏,进行花粉活力比较(图 2),发现花药黄色时期花粉活力下降最慢,与其他3个时期的花粉相比更耐储藏,4 d之后花粉活力仅下降24.00%;散粉初期和散粉盛期花粉活力下降次之,两者之间无显著性差异,4 d后分别下降41.89%和44.11%,但与黄色花药时期差异显著;散粉末期花粉活力下降最快,4 d之后花粉下降74.00%,与之前3个时期花粉储藏活力达到极显著差异。

图 2 不同储藏条件下花粉活力比较
Figure 2. Comparison of pollen viability under different storage conditions
表 1 薄壳山核桃散粉期花粉生活力
Table 1. Pollen viability of pecan during pollination
散粉状态 不同储藏时间的花粉生活力/% 0 1 2 3 4 d 即将散粉 93.00±0.31a 90.51±0.29 a 85.88±1.52 a 79.07±1.17 a 70.48±0.41 a 散粉初期 92.75±1.33 a 90.86±0.59 ab 83.03±0.55 ab 73.93±0.88 b 53.90±2.26 b 散粉盛期 88.39±1.29 b 88.03±0.82 b 80.17±0.93 b 70.39±0.79 b 49.40±1.35 b 散粉末期 79.60±1.44 c 72.27±1.20 c 58.45±0.56 c 45.28±2.31 c 20.47±1.12 c 说明:表中同列不同字母表示差异达显著性水平(P<0.05)。 表 2 散粉期花粉活力方差分析
Table 2. Comparison of pollen viability during pollination
差异来源 平方和 自由度 均方 F值 置信度 储藏时间 0.563 4 0.141 49.966 0.000 散粉期 0.313 3 0.104 36.984 0.000 误差 0.034 12 0.003 总计 0.910 19 说明:置信度<0.05差异显著,置信度<0.01差异极显著。 -
花粉散落后,其内部就开始发生一系列的生理代谢反应,活力逐渐衰退。随储藏时间的延长,花粉活力有明显下降趋势。由图 2表明:不同温度处理下花粉活力-70 ℃超低温>4 ℃低温>常温储藏(图 3),在常温、低温、超低温储藏条件下花粉活力下降呈显著性差异(表 3和图 4)。在常温条件下花粉储藏时间最短。常温储藏4 d后花粉活力为66.34%,下降29.14%,10 d后下降69.25%,30 d之后花粉活力下降到7.08%,50 d后花粉已经完全丧失活力;在常温密封干燥条件下花粉活力下降趋势与常温密封下无显著性差异,10 d后,花粉活力下降59.95%,50 d后花粉也已经完全丧失活力。而4 ℃低温条件下花粉活力储藏时间比常温储藏时间更长,低温密封储藏4 d后花粉活力为71.04 %,下降24.22%,与常温密封条件下花粉活力差异显著,10 d后下降39.03%,30 d后下降57.83%,90 d之后花粉活力下降到10.09%,120 d后花粉已经完全丧失活力;低温密封干燥与低温密封保存无显著性差异,120 d后花粉也完全丧失活力。-70 ℃超低温条件下保存效果最好,与常温、低温差异极显著,储藏50 d花粉活力仍有70.40%,仅下降24.91%,储藏120 d花粉活力下降36.62%,储藏360 d花粉活力还保持43.55%的活力,-70 ℃密封干燥要优于-70 ℃密封储藏,但差异也不显著,两者分别下降了53.55%和47.43%。

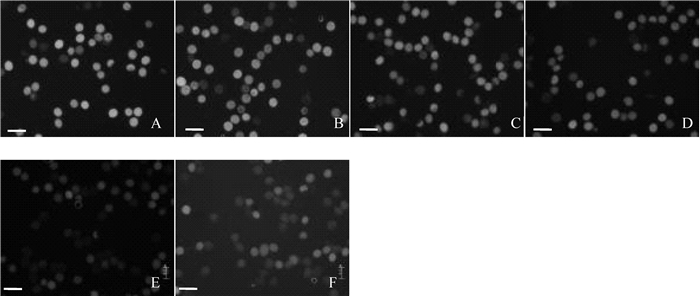
图 3 储藏4 d后4个散粉期花粉活力对比
Figure 3. Pollen vitality of four storage period after storing four days

图 4 储藏6d花粉活力对比
Figure 4. pollen vitality of four storage period after strong six days
表 3 不同储藏条件下花粉活力方差分析
Table 3. Variance analysis of pollen viability under different storage conditions
差异来源 平方和 自由度 均方 F值 置信度 储藏条件 3.781 5 0.756 20.345 0.000 储藏时间 8.037 11 0.731 19.659 0.000 误差 2.044 55 0.037 总计 13.862 71 -
在农林业生产实践中,为了进行人工辅助授粉或杂交授粉育种,需要早期采集花粉和储藏花粉,并对花粉活力进行检测[9-11]。花粉的采集时间是影响花粉活力的一个重要的因素。采集过早,花粉未完全发育, 营养物质积累不够充分,花粉少且活力低;采集时间过迟,则花多已散粉,影响花粉采集量[12]。作者参考曲柏宏等[13]通过花期物候观察确定花粉成熟期,利用荧光染色反应(FCR)测定各散粉期花粉活力。研究结果表明:不同散粉期采集的花粉生活力差异显著,从而确定最佳采粉时间是即将散粉期(花药由绿变黄时期)。作者认为,在即将散粉期,花粉粒已经发育完全,而且花粉粒仍处于花粉囊中,避免花粉与空气接触而导致花粉活力降低,所以相比较散粉期的花粉更有利于运输和保存,但这有待于进一步研究。
温度是影响花粉储藏活力的重要因素[14-16],低温能明显阻止花粉的衰老,主要原因是降低了呼吸强度和酶的活性[17]。通过研究表明:不同储藏条件对薄壳山核桃花粉活力有显著影响,这与许多研究结果一致,而且温度与花粉储藏寿命呈负相关[18]。在室温条件下,储藏4 d花粉活力下降了30.00%,储藏10 d就下降了70.00%。此方法储藏时间短,所以在生产实践上只能短期应用。在4 ℃低温条件下,储藏30 d花粉活力下降58.00%,储藏90 d花粉活力下降90.00%,在一般生产实践上低温保存能够应付花期不遇和杂交授粉等问题。在-70 ℃超低温条件下保存1 a,花粉活力还能达到50.00%,可以用作来年的辅助授粉工作或应对散粉期阴雨天气。近年来,由于应用了超低温和冷冻干燥技术保存花粉,使花粉储藏寿命大幅度延长[19],为杂交育种、种质资源的保存提供了有力支持。
Flowering characteristics and pollen viability of Carya illinoensis 'Mahan'
-
摘要: 为解决薄壳山核桃Carya illinoensis花期花粉的采集和有效保存等问题, 系统观测雄蕊开花习性, 采用荧光染色反应(FCR)法研究了不同散粉期花粉生活力差异, 以及不同储藏条件(室温密封、室温密封干燥; 4℃密封、4℃密封干燥; -70℃密封、-70℃密封干燥)和储藏时间对花粉活力的影响。结果表明:①4月26日薄壳山核桃雄蕊花萼开裂, 5月6日花药由绿变黄, 5月7-9日雄蕊进入散粉期, 5月8日散粉量最大, 占花粉总量的75%, 至5月10日花粉基本散尽, 花药变黑、小花开始脱落; ②散粉期花粉生活力大小依次为即将散粉期(花药由绿变黄)>散粉初期>散粉盛期>散粉末期, 花粉耐储藏性亦是即将散粉期最优, 即将散粉期>散粉初期>散粉盛期>散粉末期。③储藏条件对花粉活力的保持有显著影响, 利于花粉生活力保持的储藏条件依次为-70℃密封干燥> -70℃密封> 4℃密封干燥> 4℃密封>室温密封干燥>室温密封; 在任一储藏条件下, 花粉活力均随储藏时间而下降, 干燥与不干燥差异不显著。室温下花粉活力下降最快, 50 d后花粉已经完全丧失活力; 4℃下花粉活力次之, 120 d后花粉已经完全丧失活力; -70℃超低温条件下保存效果最好, 与常温、低温储藏差异极显著, 储藏360 d花粉还保持43.55%的活力。Abstract: To promote collection and the effective conservation of Carya illinoensis (pecan) pollen, male flowering habits and pollen vigor in four developmental stages with different storage conditions[a) room temperature, b) room temperature and dry; c) 4℃, d) 4℃ and dry; e) -70℃, and f) -70℃ and dry] were observed using the fluorochromatic reaction (FCR) method. Results showed that on April 26th the bract in the catkins had cracked. By May 6th the anthers had turned from green to yellow; between May 7th and 9th, the pollen sac split shedding catkin pollen. As of May 8th about 75% of the shedding had occurred, and on May 10th when the catkins turned brown and fell from the trees, shedding was complete. The sequence for pollen viability was beginning of shedding> early shedding> heavy shedding> final shedding with storage durability for the pollen having the same order. Pollen viability for different storage conditions followed decay curves, but differences were noted with -70℃ and dry> -70℃> 4℃ and dry> 4℃> room temperature and dry> room temperature. Also, as storage time increased, pollen viability gradually decreased. The fastest decline in pollen viability was at room temperature, and after 50 d the pollen had completely lost its vitality. When stored at 4℃, the pollen had mostly lost its vitality after 120 d. The best condition was preservation at -70℃ where pollen was still 43.55% viable after 360 d.
-
图 2 不同储藏条件下花粉活力比较
Figure 2 Comparison of pollen viability under different storage conditions
图 3 储藏4 d后4个散粉期花粉活力对比
A.即将散粉期; B.散粉初期; C.散粉盛期; D.散粉末期; 比例尺为70 μm。
Figure 3 Pollen vitality of four storage period after storing four days
图 4 储藏6d花粉活力对比
A. -70℃密封干燥; B. -70℃密封; C. 4℃密封干燥; D. 4℃密封; E.常温密封干燥; F.常温密封; 标尺为70 μm。
Figure 4 pollen vitality of four storage period after strong six days
表 1 薄壳山核桃散粉期花粉生活力
Table 1. Pollen viability of pecan during pollination
散粉状态 不同储藏时间的花粉生活力/% 0 1 2 3 4 d 即将散粉 93.00±0.31a 90.51±0.29 a 85.88±1.52 a 79.07±1.17 a 70.48±0.41 a 散粉初期 92.75±1.33 a 90.86±0.59 ab 83.03±0.55 ab 73.93±0.88 b 53.90±2.26 b 散粉盛期 88.39±1.29 b 88.03±0.82 b 80.17±0.93 b 70.39±0.79 b 49.40±1.35 b 散粉末期 79.60±1.44 c 72.27±1.20 c 58.45±0.56 c 45.28±2.31 c 20.47±1.12 c 说明:表中同列不同字母表示差异达显著性水平(P<0.05)。  下载: 导出CSV
下载: 导出CSV
表 2 散粉期花粉活力方差分析
Table 2. Comparison of pollen viability during pollination
差异来源 平方和 自由度 均方 F值 置信度 储藏时间 0.563 4 0.141 49.966 0.000 散粉期 0.313 3 0.104 36.984 0.000 误差 0.034 12 0.003 总计 0.910 19 说明:置信度<0.05差异显著,置信度<0.01差异极显著。
下载: 导出CSV
表 3 不同储藏条件下花粉活力方差分析
Table 3. Variance analysis of pollen viability under different storage conditions
差异来源 平方和 自由度 均方 F值 置信度 储藏条件 3.781 5 0.756 20.345 0.000 储藏时间 8.037 11 0.731 19.659 0.000 误差 2.044 55 0.037 总计 13.862 71
下载: 导出CSV
-
[1] 董凤祥, 王贵禧.薄壳山核桃引种及栽培技术[M].北京:金盾出版社, 2003. [2] 张日清, 李江, 吕芳德, 等.我国引种美国山核桃历程及资源现状研究[J].经济林研究, 2003, 21(4):107-109. ZHANG Riqing, LI Jiang, LÜ Fangde, et al. Historical survey of the introduced pecan tree in China:distribution and resource estimate[J]. Nonwood For Res, 2003, 21(4):107-109. [3] 李雪, 徐迎春, 李勇容, 等.储藏条件对薄壳山核桃4个品系花粉活力影响[J].林业科技开发, 2011, 25(1):70. LI Xue, XU Yingchun, LI Yongrong, et al. Effect of different storage condition on pollen viability of four varieties of Carya illinoensis[J]. China For Sci Technol, 2011, 25(1):70. [4] 耿国民, 周久亚, 王国祥, 等.薄壳山核桃果园良种配置方案初报[J].经济林研究, 2011, 29(2):111-113. GENG Guomin, ZHOU Jiuya, WANG Guoxiang, et al. A preliminary study on improved varieties configuration of pecan orchard[J]. Nonwood For Res, 2011, 29(2):111-113. [5] 胡芳名, 谭晓风, 刘惠民, 等.中国主要经济树种栽培与利用[M].北京:中国林业出版社, 2005:68. [6] 姚小华, 王开良, 任华东, 等.薄壳山核桃优新品种和无性系开花物候特性研究[J].江西农业大学学报, 2004, 26(5):675-680. YAO Xiahua, WANG Kailiang, REN Huadong, et al. A study on flowering phenology of Carya illinoensis new varieties and clones in East China[J]. Acta Agric Univ Jiangxi, 2004, 26(5):675-680. [7] SATO S, KATHO N, IWAI S, et al. Estabilishment of reliable method of in vitro pollen germination and pollen preservation of Brassica rapa[J]. Euphytica, 1998, 103(1):29-33. [8] 周英彪, 彭卓伦, 蒋雄辉, 等.白掌花粉活力检测及其超低温保存研究[J].中国农学通报, 2013, 29(1):113-117. ZHOU Yingbiao, PENG Zhoulun, JIANG Xionghui, et al. Study on vitality and cryopreservation of 20 Spathiphyllum floribundum pollen[J]. Chin Agric Sci Bull, 2013, 29(1):113-117. [9] 王钦丽, 卢龙斗, 吴小琴, 等.花粉的保存及其生活力测定[J].植物学通报, 2002, 19(3):365-373. WANG Qinli, LU Longdou, WU Xiaoqin, et al. Pollen preservation and its viability test[J]. Chin Bull Bot, 2002, 19(3):365-373. [10] 刘昊, 高英, 董宁光, 等.核桃优良品种雌雄花授粉活力的研究[J].北京林业大学学报, 2011, 33(6):119-123. LIU Hao, GAO Ying, DONG Ningguang, et al. Study on pollination activity of pistillate and staminate flowers in walnut excellent varieties[J]. J Beijing For Univ, 2011, 33(6):119-123. [11] JÓHANNSSON M H, STEPHENSON A G. Variation in sporophytic and gametophytic vigor in wild and cultivated varieties of Cucurbita pepo and their F1 and F2 generations[J]. Sex Plant Reprod, 1998, 11(5):265-271. [12] 刘宝, 曾杰, 程伟, 等.木本植物花粉采集、储藏与活力检测的研究进展[J].广西林业科学, 2008, 37(2):76-80. LIU Bao, ZENG Jie, CHENG Wei, et al. Research progress in pollen collection, storage and vigor test of woody plant species[J]. Guangxi For Sci, 2008, 37(2):76-80. [13] 曲柏宏, 朴永浩, 金京南.苹果梨授粉用花粉的采集、运输及储藏技术[J].延边大学农学学报, 2003, 25(1):8-12. QU Baihong, PIAO Yonghao, JIN Jingnan. Collection, transportation and storage technique of apple-pear pollen for pollinating[J]. J Agric Sci Yanbian Univ, 2003, 25(1):8-12. [14] 尹佳蕾, 赵惠恩.花粉生活力影响因素及花粉储藏概述[J].中国农学通报, 2005, 21(4):110-113. YIN Jialei, ZHAO Huien. Summary of influencia factors on pollen viability and its preservation methods[J]. Chin Agric Sci Bull, 2005, 21(4):110-113. [15] NEPI M, FRANCHI G G, PACINI E. Pollen hydration status at dispersal:cytophysiological features and strategies[J]. Protoplasnra, 2001, 216(3/4):171-180. [16] van GELDEREN E, FOSSEY A, ROBBERTSE P J. The criteria of measurement of the inorganic acid test of pollen viability[J]. South Afr J Bot, 1995, 61(5):253-259. [17] 张亚利, 尚晓倩, 刘燕.花粉超低温保存研究进展[J].北京林业大学学报, 2006, 28(4):139-147. ZHANG Yali, SHANG Xiaoqian, LIU Yan. Advances in research of pollen cryopreservation[J]. J Beijing For Univ, 2006, 28(4):139-147. [18] RODRIGUEZ-RIANO T, DAFNI A. A new procedure to asses pollen viability[J]. Sex Plant Rep, 2000, 12(4):242-244. [19] SPARKS D, YATS I E. Pecan pollen stored over a decade retains viability[J]. HortScience, 2003, 37(1):176-177. -

-
链接本文:
https://zlxb.zafu.edu.cn/article/doi/10.11833/j.issn.2095-0756.2014.04.006
 点击查看大图
点击查看大图

计量
- 文章访问数: 4032
- HTML全文浏览量: 954
- PDF下载量: 557
- 被引次数: 0
