




-
立地因子是影响树木生长发育的环境因子,包括气候、地形和土壤等[1−2]。在一定气候区内,森林的差异主要受地形、土壤等因子的影响[3]。林木更新是指林木在时间、空间尺度上的延续、发展和演替[4],对恢复森林生态系统、维持生物多样性具有重要意义[5],同时也是推动森林演替发展的重要驱动因子[6]。林木天然更新是一个受到诸多因素影响的复杂过程,除了受到林木自身的遗传学和生态学特征[7]影响外,林分结构[8]、地形因子[9]、土壤因子[10]和环境干扰[11]等对林木更新也产生重要影响。地形和土壤作为影响林木生长的两大重要立地因子,通过影响林分内的水热状况和土壤养分等对林木更新产生显著影响,在林木更新过程中的作用不容忽视[12]。黄朗等[13]在研究湖南栎类天然次生林时,发现海拔与湖南亮叶水青冈Fagus lucida、多脉青冈Quercus multinervis混交林幼树密度具有显著的正相关性。DYDERSKI等[14]研究认为:较低pH的土壤有利于挪威云杉Picea asperata幼苗密度的增加。
亚热带针阔混交林是由针叶林向常绿阔叶林过渡的一种森林植被类型,是亚热带常绿阔叶林次生演替过程中一个重要环节[15]。亚热带针阔混交林在浙闽丘陵一带广泛分布,无论是在调节气候、涵养水源等生态效益方面,还是在林产品供给、生态旅游等社会经济效益方面都有着巨大的价值[16],但目前对亚热带针阔混交林林木天然更新影响因子的研究较少。杨娟等[17]在对卧龙自然保护区针阔混交林林隙更新规律的研究中发现:土壤因子是影响优势树种天然更新和更新物种多样性的潜在因素,但并未分析土壤中各种养分及地形对林木更新的作用。本研究以浙江天目山国家级自然保护区的针阔混交林为研究对象,采用Spearman 相关性分析、单因素方差分析和线性回归模型,分析亚热带针阔混交林林木更新与地形、土壤因子之间的关系,为亚热带针阔混交林的经营管理提供理论依据。
-
浙江天目山国家级自然保护区地处浙江省杭州市临安区,30°18′30″~30°24′55″N, 119°23′47″~119°28′27″E。该区气候类型为典型亚热带季风气候,年平均气温为8.8~14.8 ℃,最冷月平均气温为2.6~3.4 ℃,最热月平均气温为19.9~28.1 ℃,无霜期为209.0~235.0 d,气候温和。受海洋暖湿气流影响,降水充足,年降水量为1 390.0~1 870.0 mm,形成浙江西北部的多雨中心。山体土壤随海拔升高由亚热带红壤向温带黄壤过渡。森林类型丰富,是中国亚热带高等植物较丰富的地区[18]。
-
2006年,在研究区选择具有代表性的地段,设置100 m×100 m的针阔混交林固定样地,用相邻格子法将样地分为100个10 m×10 m的网格单元。在每一网格单元内,对样地内胸径<5 cm的林木进行每木检尺,记录树种并测定胸径、树高、冠幅等,并采用激光对中全站仪(徕卡 TCR702Xrange)测量林木空间坐标(x, y, z)。自2006年设置样地开始,每隔5 a对该样地进行1次复查。本研究采用2021年复查数据进行分析。
-
采用网格单元中更新树种总株数和更新树种种数表示样地林木更新的物种多样性水平。
-
采用重要值分析法筛选样地优势树种[19]。
-
根据激光对中全站仪所测量得到的网格单元顶点坐标(x, y, z),计算网格单元的坡度、坡向、粗糙度[20−21]。根据样地内地形因子的数据,将各地形因子按表1标准进行分级[22]。
表 1 各地形因子分级标准
Table 1. Classification criteria for each terrain factor
分级 坡度/(°) 坡向/(°) 海拔/m 粗糙度/m Ⅰ ≥50 >202.5(西南坡) ≥680(高海拔) ≥6 Ⅱ 40~50 157.5~202.5(南坡) 650~680(中高海拔) 4~6 Ⅲ 30~40 112.5~157.5(东南坡) 620~650(中海拔) 2~4 Ⅳ <30 <112.5(东坡) <620(低海拔) <2 说明:表中数值范围采用上限排除法。 -
土壤因子包括土壤pH以及有机质、碱解氮、速效钾和有效磷质量分数5项指标。土壤pH采用复合电极法测定,有机质质量分数采用重铬酸钾容量法测定,碱解氮质量分数采用碱解扩散法测定,速效钾质量分数采用乙酸铵提取火焰光度计法测定,有效磷质量分数采用硫酸-高氯酸消煮法测定[23]。根据样地内各土壤因子的数据分布情况,将各土壤因子按表2标准进行分级。
表 2 各土壤因子分级标准
Table 2. Classification criteria for each soil factor
分级 pH 有机质质量分数/(g·kg−1) 碱解氮质量分数/(mg·kg−1) 有效磷质量分数/(mg·kg−1) 速效钾质量分数/(mg·kg−1) Ⅰ ≥5.5(弱酸) ≥200 ≥300 ≥20 ≥200 Ⅱ 5.0~5.5(酸) 100~200 200~300 10~20 150~200 Ⅲ <5.0(强酸) 50~100 100~200 5~10 <150 Ⅳ − <50 <100 <5 − 说明:表中数值范围采用上限排除法。−表示无此项。 -
分别以2个林木更新指标为因变量,以地形因子和土壤因子为自变量,构建全地形因子模型、全土壤因子模型以及全地形因子和全土壤因子的综合模型。具体构建的线性回归方程为:
$$ {{y}} = {a_0} + {k_1}\sum\limits_{i = 1}^5 {{a_i}{x_i}} + {k_2}\sum\limits_{i = 6}^9 {{a_i}{x_i}} + \varepsilon 。 $$ 其中:y表示更新树种总株数或更新树种种数;k1和k2取值为0或1;ai为回归系数(i=0,1,…,9);xi为变量(i=1,2,3,4,5时,变量为土壤因子,依次为土壤pH以及有机质、碱解氮、有效磷、速效钾质量分数;i=6,7,8,9时,变量为地形因子,依次为坡度、坡向、粗糙度、海拔);ɛ为残差。
-
采用Excel 2023对调查数据进行预处理,计算林分更新指标;利用Origin 2018绘制单元网格更新树种三维堆积图;利用SPSS 26对立地变量和林木更新因子进行正态性检验,其中更新树种总株数、更新树种种数、土壤pH、坡度、坡向、海拔以及土壤速效钾、有效磷质量分数满足正态分布,有机质、碱解氮质量分数以及地表粗糙度不满足正态分布;利用Spearman相关性分析和单因素方差分析方法分析不同地形、土壤因子等级下,样地林木更新指标的差异性;利用SPSS 26对林木更新因子构建线性回归模型。
-
通过样地更新调查,统计出更新林木共计2 991株,79个树种。筛选重要值大于2的树种为优势树种,其中阔叶树种9种,重要值占78.02%;针叶树种1种,重要值占2.78% (表3)。
表 3 样地内主要优势树种
Table 3. Main dominant species in the sample plot
树种 株数/株 株数占比/% 重要值 树种 株数/株 株数占比/% 重要值 连蕊茶Camellia fraterna 1462 48.87 35.55 榧树Torreya grandis 41 1.37 2.78 青冈Cyclobalanopsis glauca 495 16.54 15.98 山胡椒Lindera glauca 63 2.10 2.77 短尾柯Lithocarpus brevicaudatus 209 6.98 7.34 柃木Eurya japonica 46 1.53 2.28 小叶青冈Cyclobalanopsis myrsinifolia 121 4.04 5.92 豹皮樟Litsea coreana 34 1.13 2.19 润楠Machilus pingii 68 2.27 3.11 优势树种总计 2 603 87.03 80.82 马银花Rhododendron ovatum 64 2.13 2.86 对每一网格单元的林木更新情况进行统计,得到了单元网格树种更新的三维堆积图(图1)。可见,不同网格单元林木更新情况差异较大,其中连蕊茶、青冈等优势树种广泛分布在样地中。
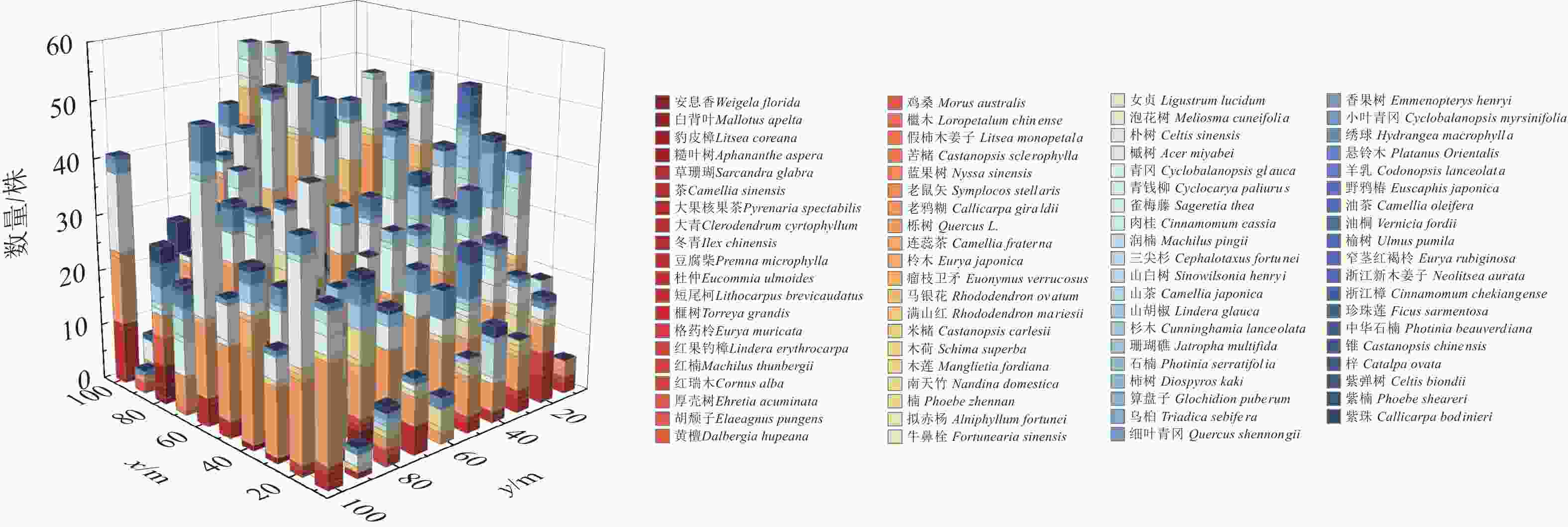
图 1 单元网格更新树种情况
Figure 1. Updated species condition of the cell grid
-
因部分变量并不符合正态分布,故对各指标进行Spearman相关分析(表4)。结果表明:更新树种总株数与土壤速效钾呈现极显著负相关(P<0.01),与海拔呈现极显著正相关(P<0.01);更新树种种数与土壤速效钾呈现极显著负相关(P<0.01),与土壤有机质呈现显著负相关(P<0.05)。说明土壤因子和地形因子影响林木更新。
表 4 相关性分析结果
Table 4. Correlation analysis
指标 S Nu pH SOM N P K Sl As SR A S 1 Nu 0.407** 1 pH 0.027 0.087 1 SOM 0.003 −0.235* −0.445** 1 N 0.001 −0.180 −0.462** 0.774** 1 P −0.168 −0.115 −0.650** 0.365** 0.412** 1 K −0.336** −0.258** −0.300** 0.531** 0.579** 0.462** 1 Sl 0.180 0.040 −0.156 0.090 −0.041 −0.004 -0.112 1 As 0.165 0.046 0.096 −0.123 −0.194 −0.146 -0.191 0.071 1 SR 0.113 −0.046 0.034 0.196 0.181 −0.143 0.065 0.102 0.146 1 A 0.551** −0.013 0.093 0.406** 0.335** −0.203* −0.087 0.187 0.017 0.327** 1 说明:*. P<0.05; **. P<0.01。S. 更新树种总株数; Nu. 更新树种种数; pH. 土壤酸碱度; SOM. 土壤有机质; N. 土壤碱解氮; P. 土壤有效磷; K. 土壤速效钾; Sl. 坡度; As. 坡向; SR. 粗糙度; A. 海拔。 -
因更新树种总株数和更新树种种数均为正态数据,故本研究采用单因素方差分析方法进行地形因子对林木更新因子影响的差异性分析。
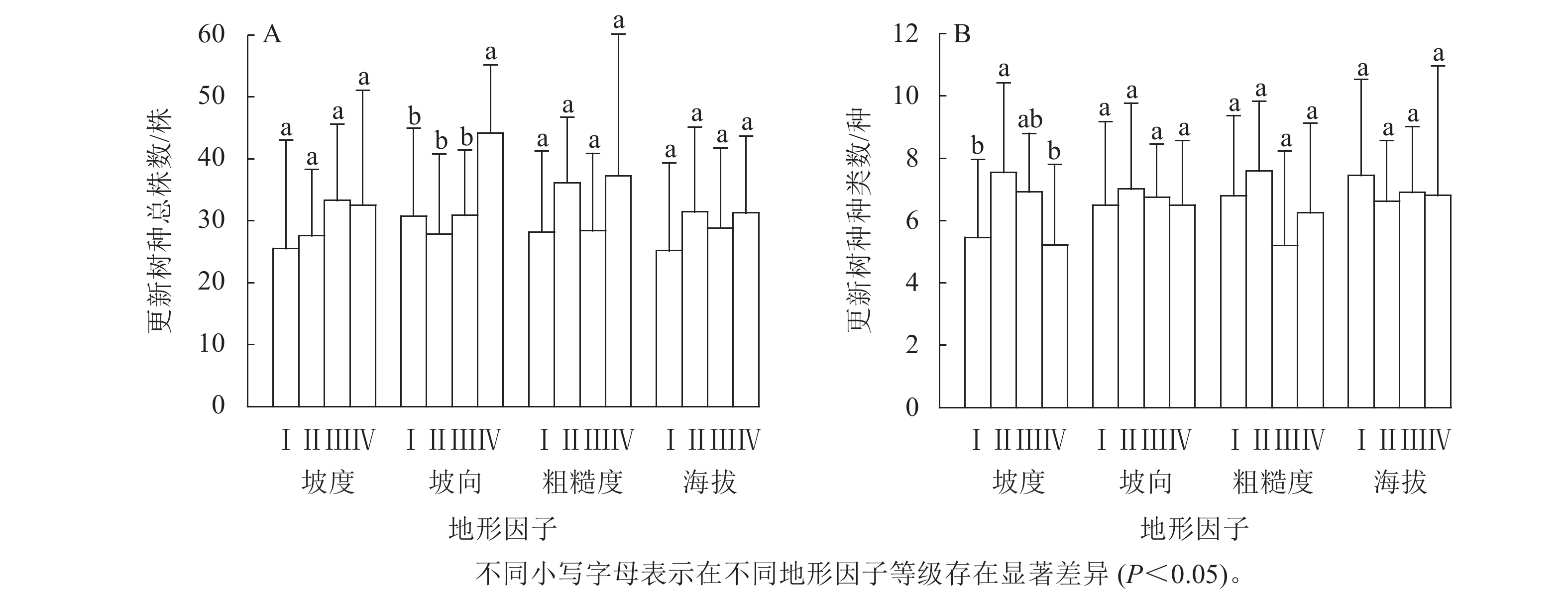
对不同坡度等级的林木更新情况进行差异性分析。结果显示:不同坡度等级的更新树种总株数的差异不显著(图2A),说明坡度对更新树种总株数的影响较小。不同坡度等级的更新树种种数具有显著差异(P<0.05,图2B)。其中,Ⅱ级坡度的更新树种种数与Ⅰ级坡度和Ⅳ级坡度的更新树种种数间均存在显著差异(P<0.05),表明坡度影响林木更新的种数。随着坡度的增加,更新树种种数先增加再减少。

图 2 地形因子对更新树种总株数和更新树种种数的影响
Figure 2. Impact of terrain factors on the total number of renewed tree species and the number of renewed tree species
对不同坡向指数的林木更新情况进行单因素方差分析。结果表明:不同坡向指数的更新树种总株数具有显著差异(P<0.05,图2A)。其中,西南坡的更新树种总株数与南坡、东南坡和东坡均间存在显著差异(P<0.05),表明坡向指数影响林木更新总株数,西南坡的更新树种总数显著高于南坡、东南坡和东坡。不同坡度指数的更新树种种数差异均不显著(图2B),说明坡向指数对更新树种种数影响较小。
对不同海拔和粗糙度等级的林木更新情况进行单因素方差分析。结果表明:海拔和粗糙度指标对更新树种总株数和更新树种种数均没有显著的影响。
-
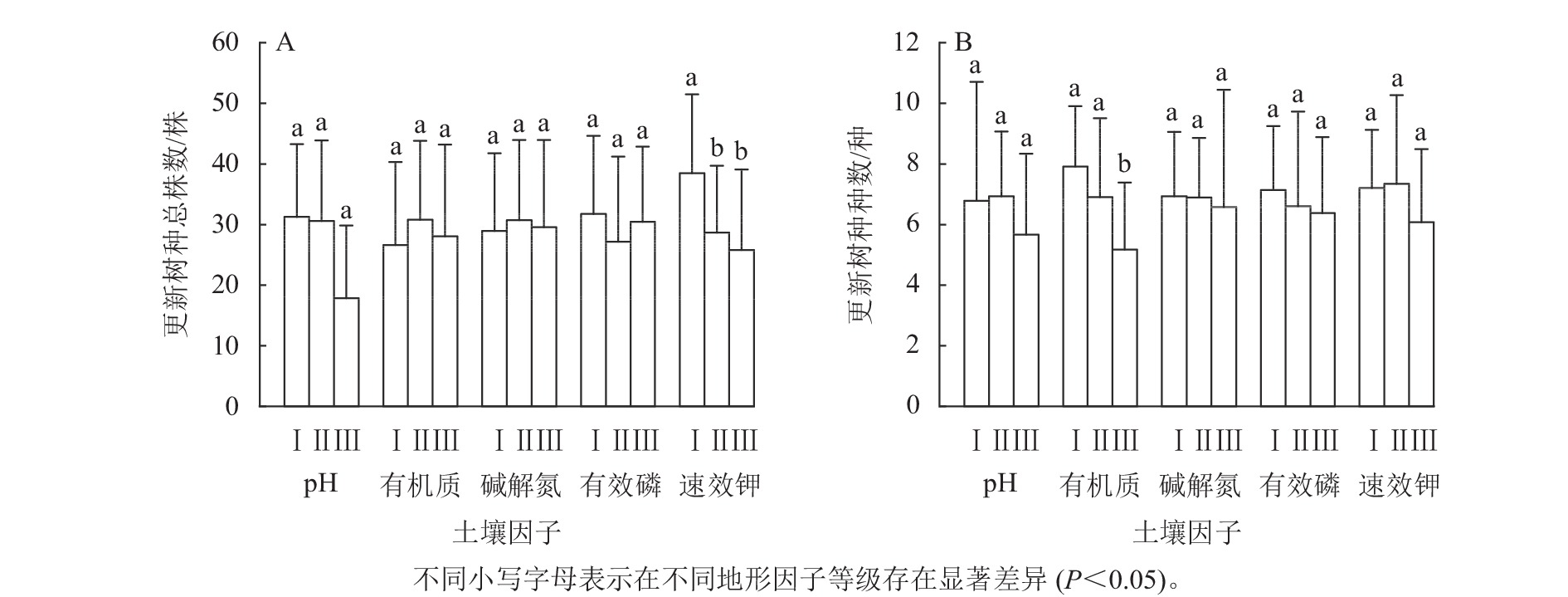
因林木更新因子均为正态性数据,故利用单因素方差分析法分析土壤因子对林木更新因子的影响。对不同有机质等级土壤的林木更新情况进行单因素方差分析。结果显示:不同有机质等级土壤的更新树种总株数均不显著(图3A),表明有机质对林木更新树种总株数无显著影响。不同有机质等级的林木更新种种数存在显著差异(图3B),其中Ⅲ级更新树种种数与Ⅰ级和Ⅱ级间均存在显著差异(P<0.05),这表明有机质影响林木更新的更新树种种数。随着有机质等级的增大,更新树种种数逐渐减小。

图 3 土壤因子对更新树种总株数和更新树种种数的影响
Figure 3. The impact of soil factors on the total number of renewed tree species and the number of renewed tree species
对不同速效钾等级土壤的林木更新情况进行单因素方差分析。结果显示:不同速效钾等级的更新树种总株数存在显著差异(图3A),其中Ⅰ级更新树种总株数与Ⅱ级和Ⅲ级间均存在显著差异(P<0.05),这表明速效钾显著影响林木更新树种总株数。随着速效钾等级的增大,更新树种的总株数逐渐减小。不同速效钾等级的更新树种种数差异不显著(图3B),说明速效钾对更新树种种数无显著性影响。
对不同土壤pH、碱解氮、有效磷等级的林木更新情况进行单因素方差分析。结果表明:土壤pH、碱解氮、有效磷指标对更新树种总株数和更新树种种数均没有显著影响。
-
分别以更新树种总株数和更新树种种数为因变量,以地形因子和土壤因子为自变量构建回归模型(表5)。
表 5 回归模型结果
Table 5. Result of the regression model
回归参数 更新树种总株数 更新树种种数 模型1 模型2 模型3 模型4 模型5 模型6 a0 62.073 −180.728 −135.979 10.657 4.338 7.142 a1 −2.937 −8.725 −0.260 −0.182 a2 0.025 −0.061 −0.011 −0.012 a3 0.040 0.010 0.004 0.004 a4 −0.209 0.302 0.015 0.016 a5 −0.143** −0.071* −0.013 −0.013 a6 0.145 0.109 0.009 a7 0.050 0.030 0.002 −0.001 a8 −1.083 −0.150 0.004 −0.083 a9 0.318** 0.339** −0.245 0.004 决定系数(R2) 0.186 0.362 0.475 0.111 0.004 0.116 调整后决定系数 0.142 0.336 0.422 0.064 0.020 0.027 F 4.290** 13.501** 9.032** 2.359* 0.491 1.306 说明:a0为回归方程截距,a1、a2、a3、a4、a5、a6、a7、a8、a9分别为回归方程中土壤pH、有机质、碱解氮、有效磷、速效钾、坡度、坡向、粗糙度、海拔项的回归系数。*. P<0.05;**. P<0.01。 以更新树种总株数为因变量的模型中,比较模型1和模型3,发现含有地形因子的模型比不含地形因子的模型拟合精度高,且模型中速效钾指标的回归系数均呈现显著负相关(P<0.05),说明速效钾会显著抑制林木更新树种总株数;比较模型2和模型3,发现含有土壤因子的模型比不含土壤因子的模型拟合精度高,且海拔的回归系数均呈现极显著正相关(P<0.01),说明海拔显著正向影响林木更新的更新树种总株数。含有土壤与地形因子的模型3拟合度最好,可以较好地解释更新树种总株数47.5%的变化原因,说明地形因子海拔和土壤因子速效钾是影响林木更新树种总株数的主要因子。
以更新树种种数为因变量的模型中,仅有模型4的F检验显著,但拟合度不高,无显著自变量,故本组线性回归模型无统计意义。以平均胸径为因变量的模型中,由于F检验均不显著,故所建模型无统计意义。
-
本研究表明:地形因子对林木更新产生了显著影响,尤其是海拔因素。随着海拔的升高,林木的更新总株数显著增加。徐振邦等[24]研究了长白山红松Pinus koraiensis阔叶混交林的天然更新条件,认为海拔显著影响林木更新,在一定海拔范围内,林木更新数量随海拔的上升而明显增加。李文博等[25]在对河南主要山区栎类天然次生林幼树更新特征及影响因子的研究中也发现:在未达到栎类次生林生长的最适海拔时,林木更新情况与海拔呈正相关。本研究进一步证实了此结论。但也有研究表明:海拔对更新有副作用。HAQ等[26]研究了喜马拉雅山主要森林类型树木更新随海拔梯度的变化,发现高海拔的森林类型树种丰富度有下降趋势,说明海拔与更新有负相关关系。TIWARI等[27]在研究海拔和坡向对加尔瓦尔山脉湿润温带森林更新的影响中也发现:树种丰富度遵循传统格局即随海拔增加而降低。此外,李进等[28]在研究华北落叶松Larix principis-rupprechtii天然次生林的更新影响因素中发现,海拔对华北落叶松和云杉的更新无显著影响。说明不同区域不同林分的更新受海拔的影响不同。随海拔的上升,更新树种种数有减小的趋势,可能是由于物种间竞争随海拔的上升而增强,环境条件逐渐变差[29],使得物种间的存活能力差异扩大,能够更新的树种种数逐步减少。
本研究发现:坡度也显著影响林木更新。更新树种种数在Ⅱ级坡度后随坡度的增大显著减小,这是由于坡度增大会增加地表径流通量和流速,改变土壤结构和植物所需营养物质,不利于植物存活[30],尤其不利于适应性低的物种更新,从而降低更新树种种数。在Ⅱ级坡度下,林木更新的效果较好,可能是因为这种坡度等级在环境中处于中等范围,有助于生态因子(如光、热和水)的均匀分布,从而促进幼苗和幼树种群数量的增长和扩展[31]。
此外,坡向通过影响林木更新的水热条件来影响林木更新[32]。本研究中,西南坡更新树种最多。但在研究地形对陕北黄土区衰退沙棘Hippophae rhamnoides人工林天然更新的影响时,张恰咛等[33]发现:当坡向为阴坡时,林木更新效果最好。连相汝[34]在对泰山侧柏 Platycladus orientalis人工林天然更新动态及幼树分布规律的研究中发现:处于阴坡的样地更新效果显著高于阳坡,说明坡向对林木更新的影响会受不同树种以及所处环境干湿状况而产生差异。本研究的样地处于亚热带季风气候区,气候暖湿,水热条件不是限制树种生长的主要原因,加上优势树种为连蕊茶、青冈等喜阳树种,故有丰富光照的阳坡更适宜生长。但在气候较干旱的陕北、泰山,水分条件是影响林木更新的关键因素,处于阴坡的树种,土壤中水分蒸发较少,有相对较好的水分条件,因而更有利于更新[35]。SEWERNIAK[36]研究了波兰内陆沙丘的坡向效应对天然更新和人工栽植樟子松Pinus sylvestris var. mongolica的影响,发现南坡的幼松更新效果好,可能是由于南坡有较好的光照,在缺水的内陆不利于杂草生长,不会对幼松的生长产生过度竞争以及影响幼松对水的需求。
已有研究表明:林地土壤是林木更新的基础,土壤的理化性质在很大程度上影响林木的更新[37]。本研究中,土壤速效钾对林木更新有显著影响,物种丰富度随土壤速效钾质量分数上升而下降。张丽珍等[38]对关帝山寒温性针叶林土壤营养状况与林下更新关系进行研究,结果表明:云杉林更新幼苗数量与土壤速效钾质量分数呈正相关。陈兵权等[39]在研究天山花楸Sorbus pohuashanensis更新影响因子时发现:土壤中钾能提高林木抗逆性,增强光合作用,与林木更新呈显著正相关。说明不同气候区,土壤速效钾对林木更新的影响差异较大。本研究发现:土壤有机质对更新树种种数有负向影响,而魏玉龙等[40]对兴安落叶松Larix gmelinii林缘天然更新与立地环境因子的相关分析中发现:土壤有机质显著正向影响兴安落叶松林缘天然更新。说明不同气候区、不同立地条件下,林木更新受土壤有机质影响也不同。
本研究未发现土壤pH、碱解氮和有效磷会对林木更新有显著影响。但许多学者研究表明:土壤pH、碱解氮、有效磷会对更新产生影响。曾思齐等[41]在研究木荷Schima superba次生林林木更新与土壤特征的相关性时,观察到物种的丰富度与土壤pH呈负相关关系,即pH越小,土壤酸性越强时,木荷的更新情况越好。PGAM等[42]对越南猫巴国家公园石灰岩林树木更新特征的研究表明:该区域树种更新与土壤因子中的pH指标呈负相关。说明pH对不同树种更新的影响程度不同。黄婷等[43]在三江源地区对3种天然圆柏(大果圆柏Juniperus tibetica、垂枝祁连圆柏J. przewalskii、密枝圆柏J. convallium)的更新进行调查时,发现土壤中有效磷质量分数对大果圆柏的更新具有显著影响,而土壤中的全氮质量分数则成为影响密枝圆柏更新的主要因素。SUKHBAATAR等[44]在研究维持欧洲赤松Pinus sylvestris林土壤特性和促进天然更新的最佳择伐强度时发现,土壤氮、pH均对欧洲赤松更新有抑制作用。卫舒平等[45]对不同密度华北落叶松林天然更新及其影响因子的研究中发现:林分更新指数与土壤全氮质量分数呈显著正相关,而与土壤全磷质量分数呈显著负相关。说明在不同立地条件、不同林分结构下,有效磷和碱解氮对更新有不同程度的影响,具体林区的更新情况受对应气候区、立地条件等因子的综合影响。
本研究还存在一些不足之处。许多研究表明:空间结构[46]、经营措施[47]等对林木更新也有显著影响。董莉莉等[48]研究认为:林分的空间结构对辽东山区蒙古栎Quercus mongolica林幼苗更新的物种多样性有显著影响。当林分中存在更大程度的混交,且垂直结构更为复杂时,可以观察到更新幼苗的种数更加丰富,导致物种多样性的提高。曾德慧等[49]研究认为:在樟子松的天然更新过程中实施必要的人工促进措施,如埋土覆盖、人工灌水、合理采伐、适当封育等,可以很好地为天然更新提供保障。此外,本研究未充分考虑两两指标的交互效应对更新的影响,也没有区别不同树种进行更新情况分析,各更新树种在促进群落正向演替过程中的作用也值得进一步探讨。
-
在浙江天目山国家级自然保护区对典型针阔混交林立地因子对林木更新影响的研究,得出以下主要结论:①样地内更新株数共2 991株·hm−2,分属79个不同树种,连蕊茶、青冈、短尾柯、小叶青冈、润楠、马银花、榧树、山胡椒、柃木、豹皮樟等10种为优势树种。②速效钾与更新总株数之间呈极显著负相关,海拔与更新总株数之间呈极显著正相关;速效钾与更新树种种数之间呈极显著负相关,有机质与更新树种种数之间呈显著负相关。③不同等级坡向和不同速效钾质量分数土壤的更新总株数有显著差异;不同等级坡度和不同有机质质量分数土壤的更新树种种数有显著差异。④以更新总株数为因变量的线性回归模型中,综合土壤与地形因子的模型拟合效果最好,其中速效钾的线性回归系数呈现显著负相关,海拔的线性回归系数呈现极显著正相关。
Impact of site factors on forest regeneration in Mount Tianmu mixed coniferous and broad-leaved forests
-
摘要:
目的 研究天目山针阔混交林立地因子对林木更新的影响,为亚热带针阔混交林的经营管理提供理论依据。 方法 以天目山针阔混交林为研究对象,将林木更新因子与土壤因子、地形因子进行Spearman相关分析,再利用单因素方差方法分析不同土壤、地形因子间林木更新的差异性,构建线性回归模型来定量描述林木更新与地形因子和土壤因子之间的关系。 结果 ①天目山针阔混交林更新树种为79种,其中优势树种为连蕊茶Camellia fraterna、青冈Cyclobalanopsis glauca、短尾柯Lithocarpus brevicaudatus、小叶青冈Cyclobalanopsis myrsinifolia、润楠Machilus pingii、马银花Rhododendron ovatum、榧树Torreya grandis、山胡椒Lindera glauca、柃木Eurya japonica、豹皮樟Litsea coreana。②速效钾与更新树种总株数呈现极显著负相关(P<0.01),海拔与更新树种总株数呈现极显著正相关(P<0.01),有机质与更新树种种数呈现显著负相关(P<0.05),速效钾与更新树种种数呈现极显著正相关(P<0.01)。③不同等级坡向和速效钾的更新树种总株数具有显著差异(P<0.05);不同等级坡度和有机质的更新树种种数有显著差异(P<0.05)。④在以更新树种总株数为因变量的线性回归模型中,速效钾、海拔的回归系数显著(P<0.05)。 结论 在天目山针阔混交林中,海拔、坡度、坡向、有机质和速效钾对林木的更新产生显著影响。图3表5参49 Abstract:Objective The objective is to study the impact of site factors on forest regeneration in the mixed coniferous and broad-leaved forests in Mount Tianmu, and provide a theoretical basis for the management of subtropical mixed coniferous and broad-leaved forests. Method A mixed coniferous and broad-leaved forest in Mount Tianmu was taken as the research object. Spearman’s correlation analysis was conducted between forest regeneration factors, soil factors and terrain factors. One-way ANOVA was used to analyze the differences in forest regeneration among different soil and terrain factors. A linear regression model was constructed to quantitatively describe the relationship between forest regeneration and terrain and soil factors. Result (1) There were 79 species of mixed coniferous and broad-leaved forests in Mount Tianmu, among which the dominant species were Camellia fraterna, Cyclobalanopsis glauca, Lithocarpus brevicaudatus, Cyclobalanopsis myrsinifolia, Machilus pingii, Rhododendron ovatum, Torreya grandis, Lindera glauca, Eurya japonica, and Litsea coreana. (2) There was a highly significant negative correlation between available potassium and the total number of updated tree species (P<0.01), a highly significant positive correlation between altitude and the total number of updated tree species (P<0.01), a significant negative correlation between organic matter and the number of updated tree species (P<0.05), and a highly significant positive correlation between available potassium and the total number of updated tree species (P<0.01). (3) There was a significant difference in the total number of updated tree species with different slope orientations and available potassium levels (P<0.05). There were significant differences in the number of updated tree species with different grades of slope and soil organic matter (P<0.05). (4) In the linear regression model with the total number of upgraded tree species as the dependent variable, the regression coefficients of available potassium and altitude were significant (P<0.05). Conclusion In the mixed coniferous and broad-leaved forests in Mount Tianmu, altitude, slope, aspect, soil organic matter and available potassium have significant effects on forest regeneration. [Ch, 3 fig. 5 tab. 49 ref.] -
Key words:
- forest regeneration /
- terrain factor /
- soil factor /
- one-way variance analysis /
- regression analysis /
- Mount Tianmu
-
图 2 地形因子对更新树种总株数和更新树种种数的影响
Figure 2 Impact of terrain factors on the total number of renewed tree species and the number of renewed tree species
图 3 土壤因子对更新树种总株数和更新树种种数的影响
Figure 3 The impact of soil factors on the total number of renewed tree species and the number of renewed tree species
表 1 各地形因子分级标准
Table 1. Classification criteria for each terrain factor
分级 坡度/(°) 坡向/(°) 海拔/m 粗糙度/m Ⅰ ≥50 >202.5(西南坡) ≥680(高海拔) ≥6 Ⅱ 40~50 157.5~202.5(南坡) 650~680(中高海拔) 4~6 Ⅲ 30~40 112.5~157.5(东南坡) 620~650(中海拔) 2~4 Ⅳ <30 <112.5(东坡) <620(低海拔) <2 说明:表中数值范围采用上限排除法。  下载: 导出CSV
下载: 导出CSV
表 2 各土壤因子分级标准
Table 2. Classification criteria for each soil factor
分级 pH 有机质质量分数/(g·kg−1) 碱解氮质量分数/(mg·kg−1) 有效磷质量分数/(mg·kg−1) 速效钾质量分数/(mg·kg−1) Ⅰ ≥5.5(弱酸) ≥200 ≥300 ≥20 ≥200 Ⅱ 5.0~5.5(酸) 100~200 200~300 10~20 150~200 Ⅲ <5.0(强酸) 50~100 100~200 5~10 <150 Ⅳ − <50 <100 <5 − 说明:表中数值范围采用上限排除法。−表示无此项。
下载: 导出CSV
表 3 样地内主要优势树种
Table 3. Main dominant species in the sample plot
树种 株数/株 株数占比/% 重要值 树种 株数/株 株数占比/% 重要值 连蕊茶Camellia fraterna 1462 48.87 35.55 榧树Torreya grandis 41 1.37 2.78 青冈Cyclobalanopsis glauca 495 16.54 15.98 山胡椒Lindera glauca 63 2.10 2.77 短尾柯Lithocarpus brevicaudatus 209 6.98 7.34 柃木Eurya japonica 46 1.53 2.28 小叶青冈Cyclobalanopsis myrsinifolia 121 4.04 5.92 豹皮樟Litsea coreana 34 1.13 2.19 润楠Machilus pingii 68 2.27 3.11 优势树种总计 2 603 87.03 80.82 马银花Rhododendron ovatum 64 2.13 2.86
下载: 导出CSV
表 4 相关性分析结果
Table 4. Correlation analysis
指标 S Nu pH SOM N P K Sl As SR A S 1 Nu 0.407** 1 pH 0.027 0.087 1 SOM 0.003 −0.235* −0.445** 1 N 0.001 −0.180 −0.462** 0.774** 1 P −0.168 −0.115 −0.650** 0.365** 0.412** 1 K −0.336** −0.258** −0.300** 0.531** 0.579** 0.462** 1 Sl 0.180 0.040 −0.156 0.090 −0.041 −0.004 -0.112 1 As 0.165 0.046 0.096 −0.123 −0.194 −0.146 -0.191 0.071 1 SR 0.113 −0.046 0.034 0.196 0.181 −0.143 0.065 0.102 0.146 1 A 0.551** −0.013 0.093 0.406** 0.335** −0.203* −0.087 0.187 0.017 0.327** 1 说明:*. P<0.05; **. P<0.01。S. 更新树种总株数; Nu. 更新树种种数; pH. 土壤酸碱度; SOM. 土壤有机质; N. 土壤碱解氮; P. 土壤有效磷; K. 土壤速效钾; Sl. 坡度; As. 坡向; SR. 粗糙度; A. 海拔。
下载: 导出CSV
表 5 回归模型结果
Table 5. Result of the regression model
回归参数 更新树种总株数 更新树种种数 模型1 模型2 模型3 模型4 模型5 模型6 a0 62.073 −180.728 −135.979 10.657 4.338 7.142 a1 −2.937 −8.725 −0.260 −0.182 a2 0.025 −0.061 −0.011 −0.012 a3 0.040 0.010 0.004 0.004 a4 −0.209 0.302 0.015 0.016 a5 −0.143** −0.071* −0.013 −0.013 a6 0.145 0.109 0.009 a7 0.050 0.030 0.002 −0.001 a8 −1.083 −0.150 0.004 −0.083 a9 0.318** 0.339** −0.245 0.004 决定系数(R2) 0.186 0.362 0.475 0.111 0.004 0.116 调整后决定系数 0.142 0.336 0.422 0.064 0.020 0.027 F 4.290** 13.501** 9.032** 2.359* 0.491 1.306 说明:a0为回归方程截距,a1、a2、a3、a4、a5、a6、a7、a8、a9分别为回归方程中土壤pH、有机质、碱解氮、有效磷、速效钾、坡度、坡向、粗糙度、海拔项的回归系数。*. P<0.05;**. P<0.01。
下载: 导出CSV
-
[1] 张万儒, 盛炜彤, 蒋有绪, 等. 中国森林立地分类系统[J]. 林业科学研究, 1992, 5(3): 251 − 262. ZHANG Wanru, SHENG Weitong, JIANG Youxu, et al. Classification of forest site system in China [J]. Forestry Research, 1992, 5(3): 251 − 262. [2] 王高峰. 森林立地分类研究评介[J]. 南京林业大学学报, 1986, 9(3): 108 − 124. WANG Gaofeng. A review of the study of forest site classification [J]. Journal of Nanjing Forestry University, 1986, 9(3): 108 − 124. [3] 周政贤, 杨世逸. 试论我国立地分类理念基础[J]. 林业科学, 1987, 23(1): 61 − 67. ZHOU Zhenxian, YANG Shiyi. A discussion on the basic theory of site classification in China [J]. Scientia Silvae Sinicae, 1987, 23(1): 61 − 67. [4] 金永焕, 李敦求, 姜好相, 等. 长白山区次生林恢复过程中天然更新的动态[J]. 南京林业大学学报:自然科学版, 2005, 29(5): 65 − 68. JIN Yonghuan, LEE D K, KANG H S, et al. Quantitative dynamics on natural regeneration of secondary forest during the restoration period in Changbai Mountain Area [J]. Journal of Nanjing Forestry University (Natural Science Edition), 2005, 29(5): 65 − 68. [5] ALAKÄRPPÄ E, TAULAVUORI E, VALLEDOR L, et al. Early growth of Scots pine seedlings is affected by seed origin and light quality [J]. Journal of Plant Physiology, 2019, 237: 120 − 128. [6] 韩豪. 闽楠天然林种群特征及其天然更新研究[D]. 贵阳: 贵州大学, 2020. HAN Hao. A Research on the Population Characteristics of Natural Forest of Phoebe bournei and Its Regeneration [D]. Guiyang: Guizhou University, 2020. [7] TINYA F, MARIALIGETI S, BIDLO A, et al. Environmental drivers of the forest regeneration in temperate mixed forests [J]. Forest Ecology and Management, 2019, 433: 720 − 728. [8] 欧芷阳, 庞世龙, 谭长强, 等. 林分结构对桂西南蚬木种群天然更新的影响[J]. 应用生态学报, 2017, 28(10): 3181 − 3188. OU Zhiyang, PANG Shilong, TAN Changqiang, et al. Effects of forest structure on natural regeneration of Excentrodendron hsienmu population in Southwest Guangxi, China [J]. Chinese Journal of Applied Ecology, 2017, 28(10): 3181 − 3188. [9] LE A V, PAULL D J, GRIFFIN A L. Exploring the inclusion of small regenerating trees to improve above-ground forest biomass estimation using geospatial data [J/OL]. Remote Sensing, 2018, 10(9): 1446[2023-07-25]. doi: 10.3390/rs10091446. [10] SOHRABI H, JOURGHOLAMI M, TAVANKAR F, et al. Post-harvest evaluation of soil physical properties and natural regeneration growth in steep-slope terrains [J/OL]. Forests, 2019, 10(11): 1034[2023-07-25]. doi: 10.3390/f10111034. [11] PRÉVOSTO B, GAVINET J, RIPERT C, et al. Identification of windows of emergence and seedling establishment in a pine Mediterranean forest under controlled disturbances [J]. Basic and Applied Ecology, 2015, 16(1): 36 − 45. [12] 孙敬然. 泰山麻栎林天然更新特征及影响因素分析[D]. 泰安: 山东农业大学, 2022. SUN Jingran. Natural Regeneration Characteristics and Influencing Factors of Quercus acutissima Forest in Taishan [D]. Tai’an: Shandong Agricultural University, 2022. [13] 黄朗, 朱光玉, 康立, 等. 湖南栎类天然次生林幼树更新特征及影响因子[J]. 生态学报, 2019, 39(13): 4900 − 4909. HUANG Lang, ZHU Guangyu, KANG Li, et al. Regeneration characteristics and related factors affecting saplings in Quercus spp. natural secondary forests in Hunan Province, China [J]. Acta Ecologica Sinica, 2019, 39(13): 4900 − 4909. [14] DYDERSKI M K, GAZDA A, HACHULKA M , et al. Impacts of soil conditions and light availability on natural regeneration of Norway spruce Picea abies (L.) H. Karst. in low-elevation mountain forests [J/OL]. Annals of Forest Science, 2018, 75(4): 91[2023-07-25]. doi: 10.1007/s13595-018-0775-x. [15] 王周平, 李旭光, 石胜友, 等. 重庆缙云山针阔混交林林隙树木更替规律研究[J]. 植物生态学报, 2001, 25(4): 399 − 404. WANG Zhouping, LI Xuguang, SHI Shengyou, et al. Gap phase tree replacement in a conifer-broadleaved mixed forest in Jinyun Mountain, Chongqing [J]. Acta Phytoecologica Sinica, 2001, 25(4): 399 − 404. [16] 吴强, 张合平. 中国亚热带4种森林服务功能价值与补偿标准研究[J]. 中南林业科技大学学报, 2017, 37(7): 140 − 148. WU Qiang, ZHANG Heping. Four kinds of China subtropical forest service function value and compensation standard research [J]. Journal of Central South University of Forestry and Technology, 2017, 37(7): 140 − 148. [17] 杨娟, 葛剑平, 刘丽娟, 等. 卧龙自然保护区针阔混交林林隙更新规律[J]. 植物生态学报, 2007, 31(3): 425 − 430. YANG Juan, GE Jianping, LIU Lijuan, et al. Gap-phase regeneration rules of conifer-broadleaved mixed forest in Wolong Nature Reserve, China [J]. Journal of Plant Ecology, 2007, 31(3): 425 − 430. [18] 汤孟平, 周国模, 施拥军, 等. 天目山常绿阔叶林优势种群及其空间分布格局[J]. 植物生态学报, 2006, 30(5): 743 − 752. TANG Mengping, ZHOU Guomo, SHI Yongjun, et al. Study of dominant plant populations and their spatial patterns in evergreen broadleaved forest in Tianmu Mountain, China [J]. Journal of Plant Ecology, 2006, 30(5): 743 − 752. [19] FEROZ S M, YOSHIMURA K, HAGIHARA A. Stand stratification and woody species diversity of a subtropical forest in limestone habitat in the northern part of Okinawa Island [J]. Journal of Plant Research, 2008, 121(3): 329 − 337. [20] 汤孟平, 周国模, 施拥军, 等. 不同地形条件下群落物种多样性与胸高断面积的差异分析[J]. 林业科学, 2007, 43(6): 27 − 31. TANG Mengping, ZHOU Guomo, SHI Yongjun, et al. Difference analysis of community biodiversity and basal area under different terrain [J]. Scientia Silvae Sinicae, 2007, 43(6): 27 − 31. [21] 黄杏元, 马劲松. 地理信息系统概论[M]. 北京: 高等教育出版社, 2008. HUANG Xingyuan, MA Jinsong. Introduction to Geographic Information Systems [M]. Beijing: Higher Education Press, 2008. [22] 龙俊松, 汤孟平. 天目山常绿阔叶林空间结构与地形因子的关系[J]. 浙江农林大学学报, 2021, 38(1): 47 − 57. LONG Junsong, TANG Mengping. Relationship between spatial structure and terrain factors of evergreen broad-leaved forest in Mount Tianmu [J]. Journal of Zhejiang A&F University, 2021, 38(1): 47 − 57. [23] 马鑫茹, 郑旭理, 郑春颖, 等. 毛竹扩张对常绿阔叶林土壤微生物群落的影响[J]. 应用生态学报, 2022, 33(4): 1091 − 1098. MA Xinru, ZHENG Xuli, ZHENG Chunying, et al. Effects of moso bamboo (Phyllostachys edulis) expansion on soil microbial community in evergreen broad-leaved forest [J]. Chinese Journal of Applied Ecology, 2022, 33(4): 1091 − 1098. [24] 徐振邦, 代力民, 陈吉泉, 等. 长白山红松阔叶混交林森林天然更新条件的研究[J]. 生态学报, 2001, 21(9): 1413 − 1420. XU Zhenbang, Dai Limin, CHEN Jiquan, et al. Natural regeneration condition in Pinus koraiensis broad-leaved mixed forest [J]. Acta Ecologica Sinica, 2001, 21(9): 1413 − 1420. [25] 李文博, 石君杰, 张向阳, 等. 河南主要山区栎类天然次生林幼树更新特征及影响因子[J]. 河南林业科技, 2023, 43(2): 1 − 5. LI Wenbo, SHI Junjie, ZHANG Xiangyang, et al. Regeneration characteristics and related factors affecting saplings in Quercus secondary forest of main mountainous areas in Henan Province [J]. Journal of Henan Forestry Science and Technology, 2023, 43(2): 1 − 5. [26] HAQ S M, CALIXTO E S, RASHID I, et al. Tree diversity, distribution and regeneration in major forest types along an extensive elevational gradient in Indian Himalaya: Implications for sustainable forest management [J/OL]. Forest Ecology and Management, 2022, 506: 119968[2023-07-25]. doi: 10.1016/j.foreco.2021.119968. [27] TIWARI O P, SHARMA C M, RANA Y S. Influence of altitude and slope-aspect on diversity, regeneration and structure of some moist temperate forests of Garhwal Himalaya [J]. Tropical Ecology, 2020, 61: 278 − 289. [28] 李进, 石晓东, 高润梅, 等. 华北落叶松天然次生林更新及影响因素[J]. 森林与环境学报, 2020, 40(6): 588 − 596. LI Jin, SHI Xiaodong, GAO Runmei, et al. Regeneration and affecting factors of Larix principis-rupprechtii natural secondary forests [J]. Journal of Forest and Environment, 2020, 40(6): 588 − 596. [29] 王志鸣, 郭秋菊, 艾训儒, 等. 恩施州不同海拔马尾松天然次生林林分结构特征研究[J]. 西南林业大学学报(自然科学), 2019, 39(1): 114 − 122. WANG Zhiming, GUO Qiuju, AI Xunru, et al. Structure characteristics of Pinus massoniana natural secondary forest at different altitude of Enshi [J]. Journal of Southwestern Forestry University (Natural Science), 2019, 39(1): 114 − 122. [30] 赵家豪, 孙晓丹, 叶钰倩, 等. 微地形对江西武夷山南方铁杉针阔混交林幼树更新的影响[J]. 生态学报., 2022, 42(6): 2357 − 2367. ZHAO Jiahao, SUN Xiaodan, YE Yuqian, et al. Effect of micro-topography on the saplings regeneration in the coniferous (Tsuga chinensis var. tchekiangensis) and broadleaf mixed forest in the Wuyishan, Jiangxi Province [J]. Acta Ecologica Sinica, 2022, 42(6): 2357 − 2367. [31] 董丽, 郭东罡, 段毅豪, 等. 灵空山辽东栎-油松林更新空间分布格局及其与地形因子的关系[J]. 应用与环境生物学报, 2013, 19(6): 914 − 921. DONG Li, GUO Donggang, DUAN Yihao, et al. Spatial distribution patterns of regenerating Quercus wutaishanica-Pinus tabulaeformis in relation to topographic factors in the Lingkong Mountain [J]. Chinese Journal of Applied and Environmental Biology, 2013, 19(6): 914 − 921. [32] 佘萍. 六盘山华北落叶松林天然更新影响因子与人工促进方法初步研究[D]. 银川: 宁夏大学, 2022. SHE Ping. Preliminary Study on the Influencing Factors of Natural Regeneration and Artificial Stimulating of Larix principis-rupprechtii Mayr in Liupan Mountains [D]. Yinchuan: Ningxia University, 2022. [33] 张恰咛, 朱清科, 任正龑, 等. 地形对陕北黄土区衰退沙棘人工林天然更新的影响[J]. 林业科学研究, 2017, 30(2): 300 − 306. ZHANG Qianing, ZHU Qingke, REN Zhengyan, et al. Modeling the effects of topography on the natural renewal of declining sea-buckthorn forest in Loess Plateau of northern Shaanxi Province [J]. Forestry Research, 2017, 30(2): 300 − 306. [34] 连相汝. 泰山侧柏人工林天然更新动态及幼树分布规律研究[D]. 泰安: 山东农业大学, 2014. LIAN Xiangru. Reseach on Natural Regeneration Dynamics and Sapling Distribution of Oriental Arborvitae Plantation of Mount Tai [D]. Tai’an: Shandong Agricultural University, 2014. [35] 卢彦磊, 张文辉, 杨斌, 等. 秦岭中段不同坡向锐齿栎种子雨、土壤种子库与幼苗更新[J]. 应用生态学报, 2019, 30(6): 1965 − 1973. LU Yanlei, ZHANG Wenhui, YANG Bin, et al. Seed rain, soil seed bank and seedling regeneration of Quercus aliena var. acureserrata in different slope directions on the middle Qinling Mountains, China [J]. Chinese Journal of Applied Ecology, 2019, 30(6): 1965 − 1973. [36] SEWERNIAK P. Differences in early dynamics and effects of slope aspect between naturally regenerated and planted Pinus sylvestris woodland on inland dunes in Poland [J]. iForest-Biogeosciences and Forestry, 2016, 9: 875 − 882. [37] DAVIS E L, HAGER H A, GEDALOF Z. Soil properties as constraints to seedling regeneration beyond alpine treelines in the Canadian Rocky Mountains [J/OL]. Arctic, Antarctic, and Alpine Research, 2018, 50(1): e1415625[2023-07-25]. doi: 10.1080/15230430.2017.1415625. [38] 张丽珍, 牛伟, 郭晋平, 等. 关帝山寒温性针叶林土壤营养状况与林下更新关系研究[J]. 西北植物学报, 2005, 25(7): 1329 − 1334. ZHANG Lizhen, NIU Wei, GUO Jinping, et al. Relationship of soil nutrients with the regeneration of cold temperate coniferous forest in the Guandishan Mountain [J]. Acta Botanica Boreali-Occidentalia Sinica, 2005, 25(7): 1329 − 1334. [39] 陈兵权, 赵善超, 古丽米热·艾合买提, 等. 天山花楸的天然更新及其影响因子[J]. 安徽农业大学学报, 2023, 50(2): 206 − 212. CHEN Bingquan, ZHAO Shanchao, Gulimire Aihemaiti, et al. Natural regeneration of Tianshan rowan Rupr. and its influencing factors [J]. Journal of Anhui Agricultural University, 2023, 50(2): 206 − 212. [40] 魏玉龙, 张秋良. 兴安落叶松林缘天然更新与立地环境因子的相关分析[J]. 南京林业大学学报(自然科学版), 2020, 44(2): 165 − 172. WEI Yulong, ZHANG Qiuliang. Forest edge renewal of Larix gmelinii and its response to the environment [J]. Journal of Nanjing Forestry University (Natural Science Edition), 2020, 44(2): 165 − 172. [41] 曾思齐, 甘静静, 肖化顺, 等. 木荷次生林林木更新与土壤特征的相关性[J]. 生态学报, 2014, 34(15): 4242 − 4250. ZENG Siqi, GAN Jingjing, XIAO Huashun, et al. Changes in soil properties in regenerating Schima superba secondary forests [J]. Acta Ecologica Sinica, 2014, 34(15): 4242 − 4250. [42] PGAM V V, AMMER C, ANNIGHÖFER P, et al. Tree regeneration characteristics in limestone forests of the Cat Ba National Park, Vietnam [J/OL]. BMC Ecology and Evolution, 2022, 22(1): 6[2023-07-25]. doi: 10.1186/s12862-021-01957-9. [43] 黄婷, 郝家田, 杜一尘, 等. 青海三江源地区三种天然圆柏林更新特征[J]. 应用生态学报, 2022, 33(2): 297 − 303. HUANG Ting, HAO Jiatian, DU Yichen, et al. Regeneration characteristics of three natural Juniperus forests in the Three-River Headwater Region of Qinghai Province, China [J]. Chinese Journal of Applied Ecology, 2022, 33(2): 297 − 303. [44] SUKHBAATAR G, BAATARBILEG N, PUREVRAGCHAA B, et al. Which selective logging intensity is most suitable for the maintenance of soil properties and the promotion of natural regeneration in highly continental scots pine forests? – results 19 years after harvest operations in Mongolia [J/OL]. Forests, 2019,10(2): 141[2023-07-25]. doi: 10.3390/f10020141. [45] 卫舒平, 梁文俊, 魏曦, 等. 不同密度华北落叶松林天然更新及其影响因子[J]. 应用生态学报, 2022, 33(10): 2687 − 2694. WEI Shuping, LIANG Wenjun, WEI Xi, et al. Natural regeneration of Larix principis-rupprechtii plantations with different densities and its influencing factors [J]. Chinese Journal of Applied Ecology, 2022, 33(10): 2687 − 2694. [46] 张树梓, 李梅, 张树彬, 等. 塞罕坝华北落叶松人工林天然更新影响因子[J]. 生态学报, 2015, 35(16): 5403 − 5411. ZHANG Shuzi, LI Mei, ZHANG Shubin, et al. Factors affecting natural regeneration of Larix principis-rupprechtii plantations in Saihanba of Hebei, China [J]. Acta Ecologica Sinica, 2015, 35(16): 5403 − 5411. [47] RIBEIRO S, CERVEIRA A, SOARES P S, et al. Natural regeneration of maritime pine: a review of the influencing factors and proposals for management [J/OL]. Forests, 2022, 13(3): 386[2023-07-26]. doi: 10.3390/f13030386. [48] 董莉莉, 汪成成, 赵济川, 等. 辽东山区蒙古栎次生林天然更新特征及其影响因子分析[J]. 西北农林科技大学学报(自然科学版), 2023, 51(1): 94 − 101, 109. DONG Lili, WANG Chengcheng, ZHAO Jichuan, et al. Natural regeneration characteristics and influencing factors of Quercus mongolica secondary forest in the mountainous area of eastern Liaoning [J]. Journal of Northwest A&F University (Natural Science Edition), 2023, 51(1): 94 − 101, 109. [49] 曾德慧, 尤文忠, 范志平, 等. 樟子松人工固沙林天然更新障碍因子分析[J]. 应用生态学报, 2002, 13(3): 257 − 261. ZENG Dehui, YOU Wenzhong, FAN Zhiping, et al. Analysis of natural regeneration barriers of Pinus sylvestris var. mongolica plantation on sandy land [J]. Chinese Journal of Applied Ecology, 2002, 13(3): 257 − 261. -

-
链接本文:
https://zlxb.zafu.edu.cn/article/doi/10.11833/j.issn.2095-0756.20230446
 点击查看大图
点击查看大图
计量
- 文章访问数: 1626
- HTML全文浏览量: 446
- PDF下载量: 34
- 被引次数: 0
